
A High-Pressure, High-Temperature Flow Reactor Simulating the Hadean Earth Environment, with Application to the Pressure Dependence of the Cleavage of Avocado Viroid Hammerhead Ribozyme

 ,
, 
Abstract
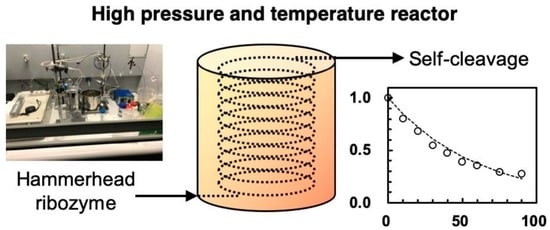
:
1. Introduction
2. Materials and Methods
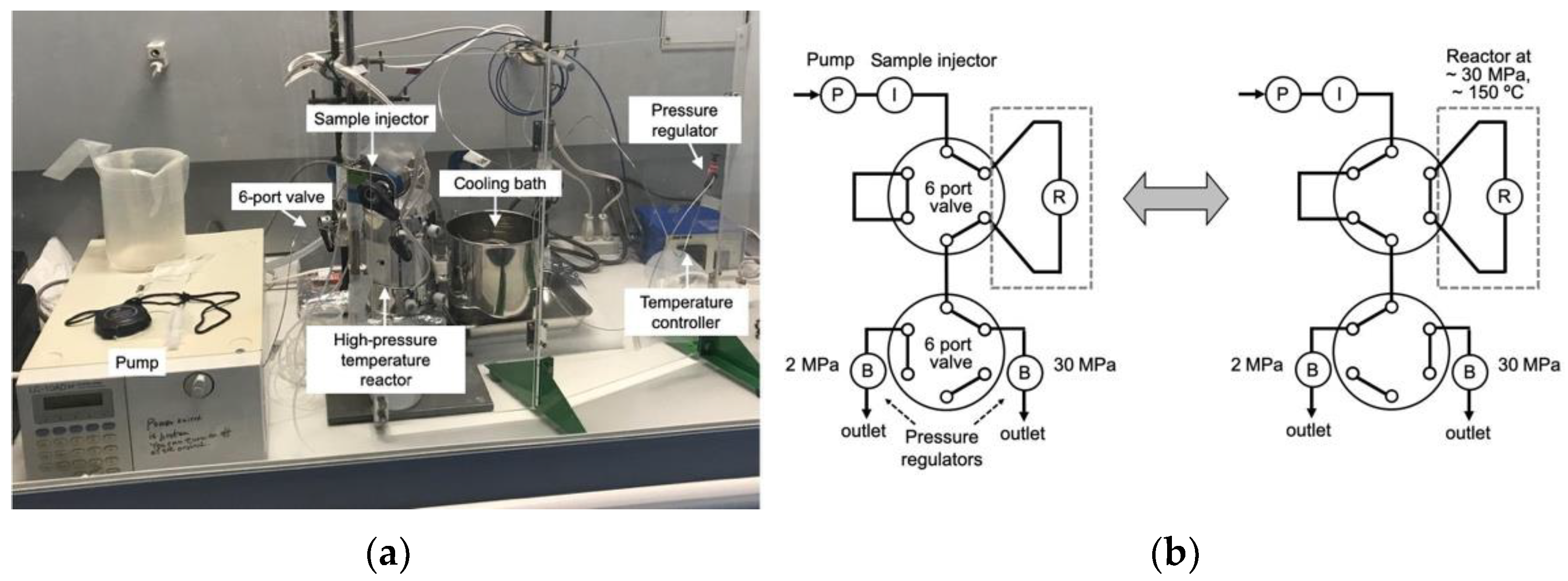
2.1. High-Pressure Reactor (HPR-30)
2.2. Materials
2.3. ASBVd(−):HHR Self-Cleavage at 0.1 MPa
2.4. ASBVd(−):HHR Self-Cleavage at 30 MPa
2.5. HPLC Analysis for the Products of ASBVd(−):HHR Self-Cleavage
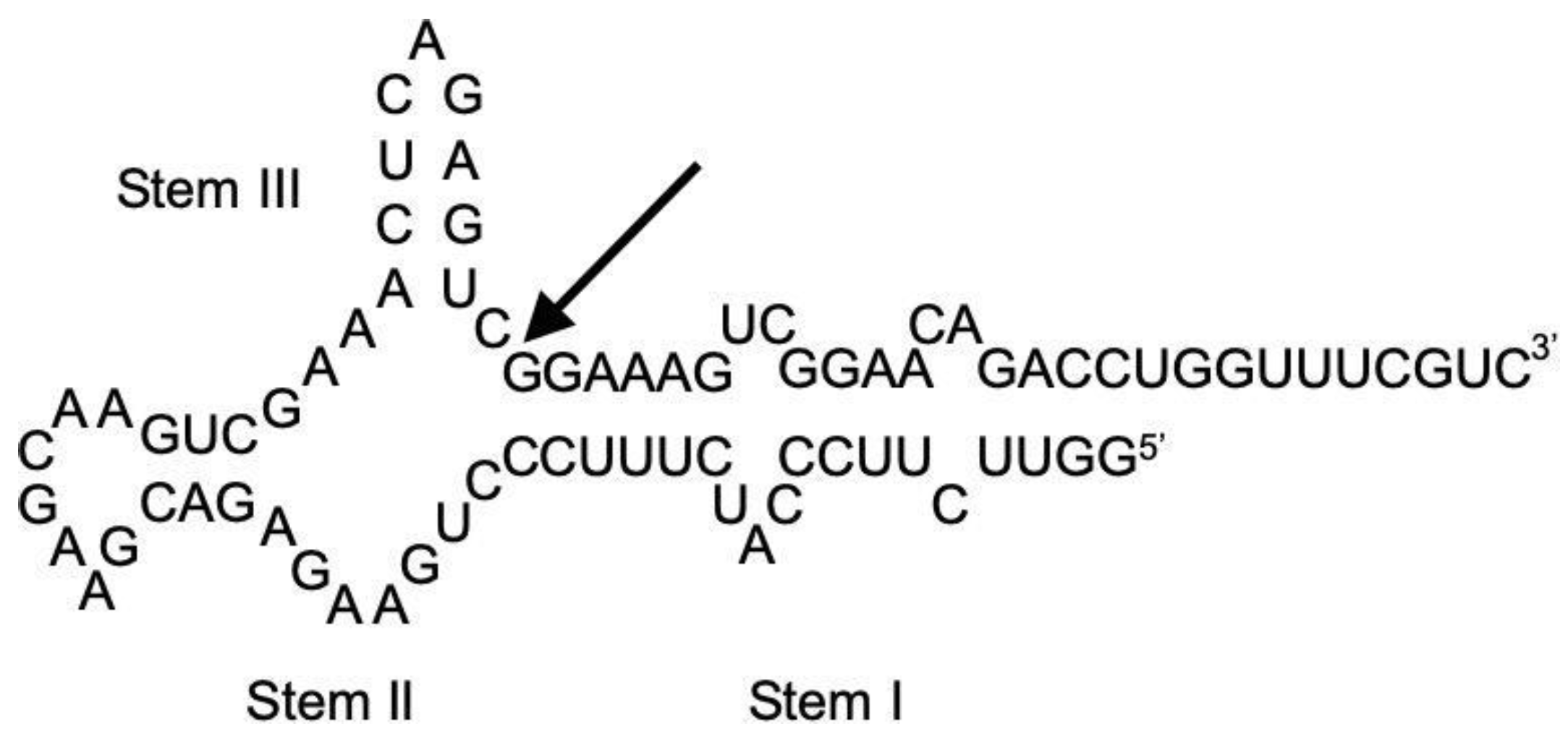
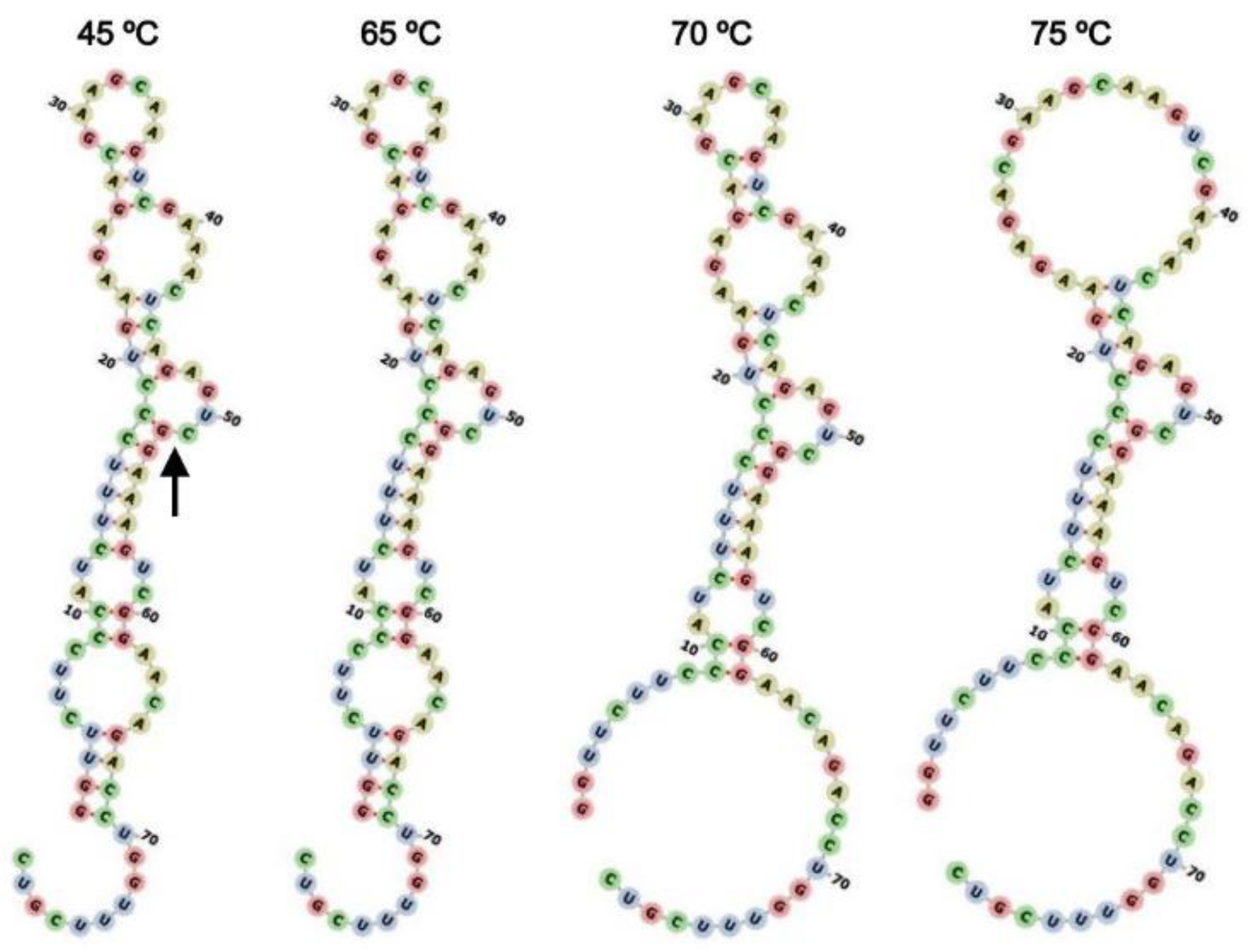
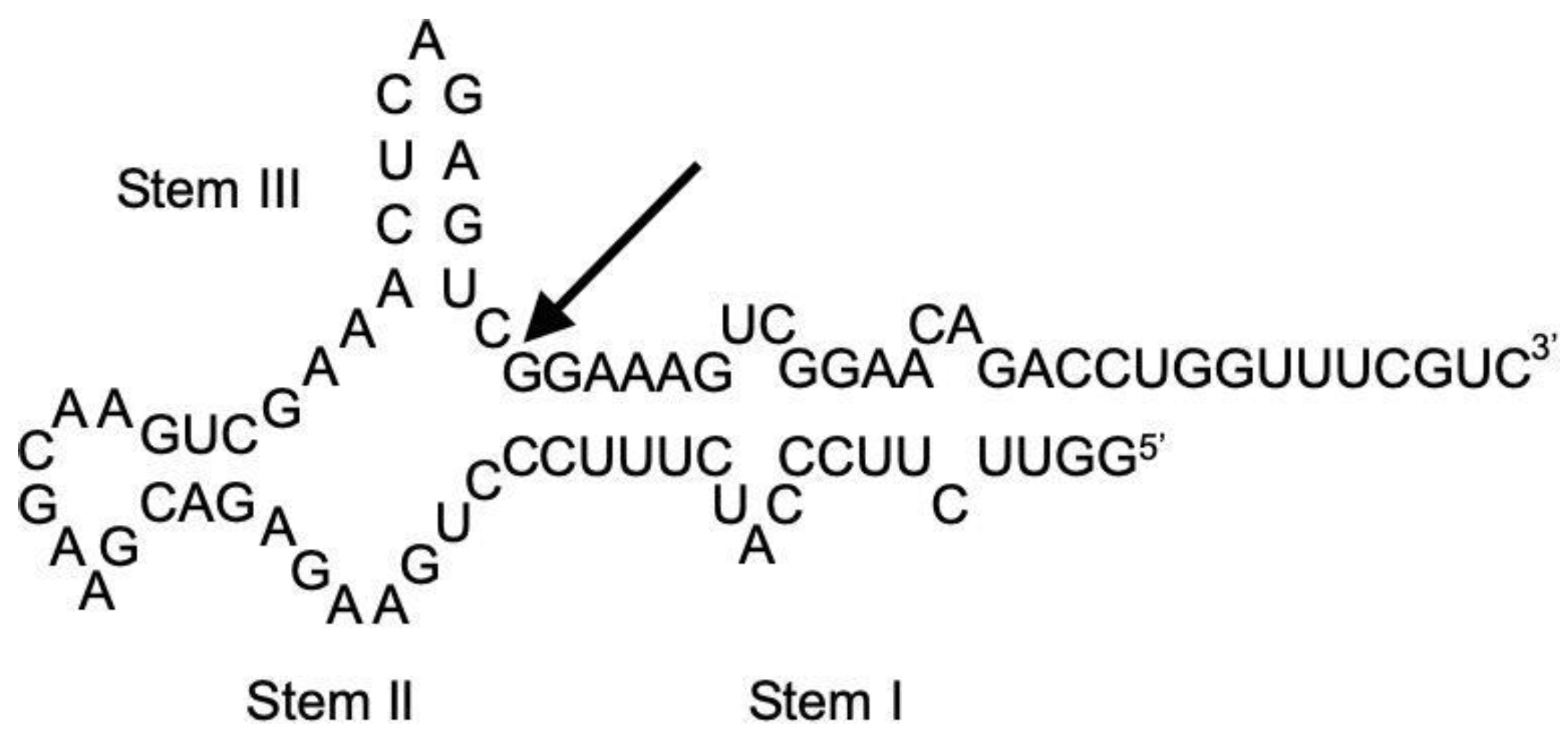
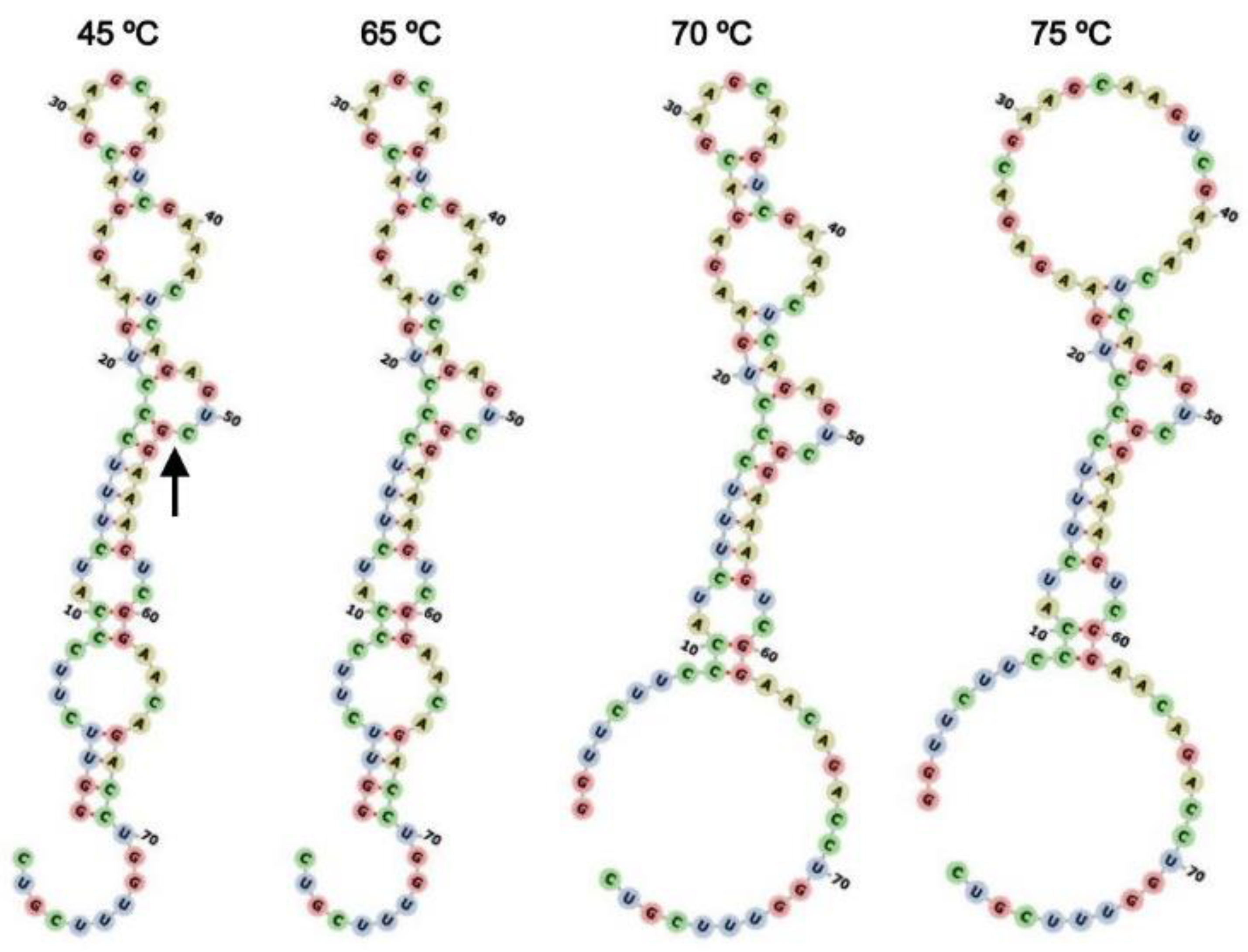
2.6. Secondary Structure Modeling of HHR:ASBVd(−)
3. Results and Discussion
3.1. Construction of High-Pressure Reactor and Performance of HPR-30
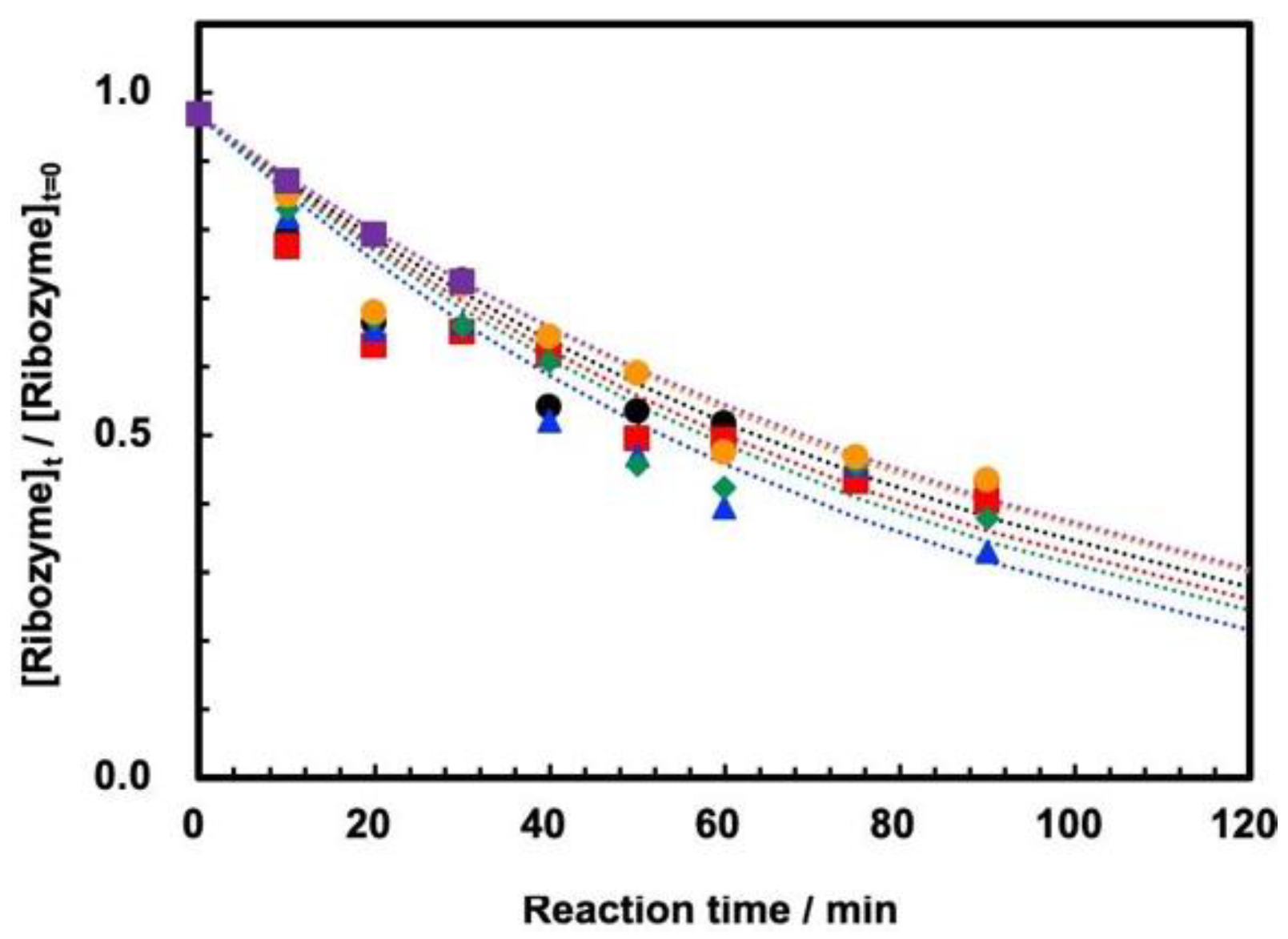
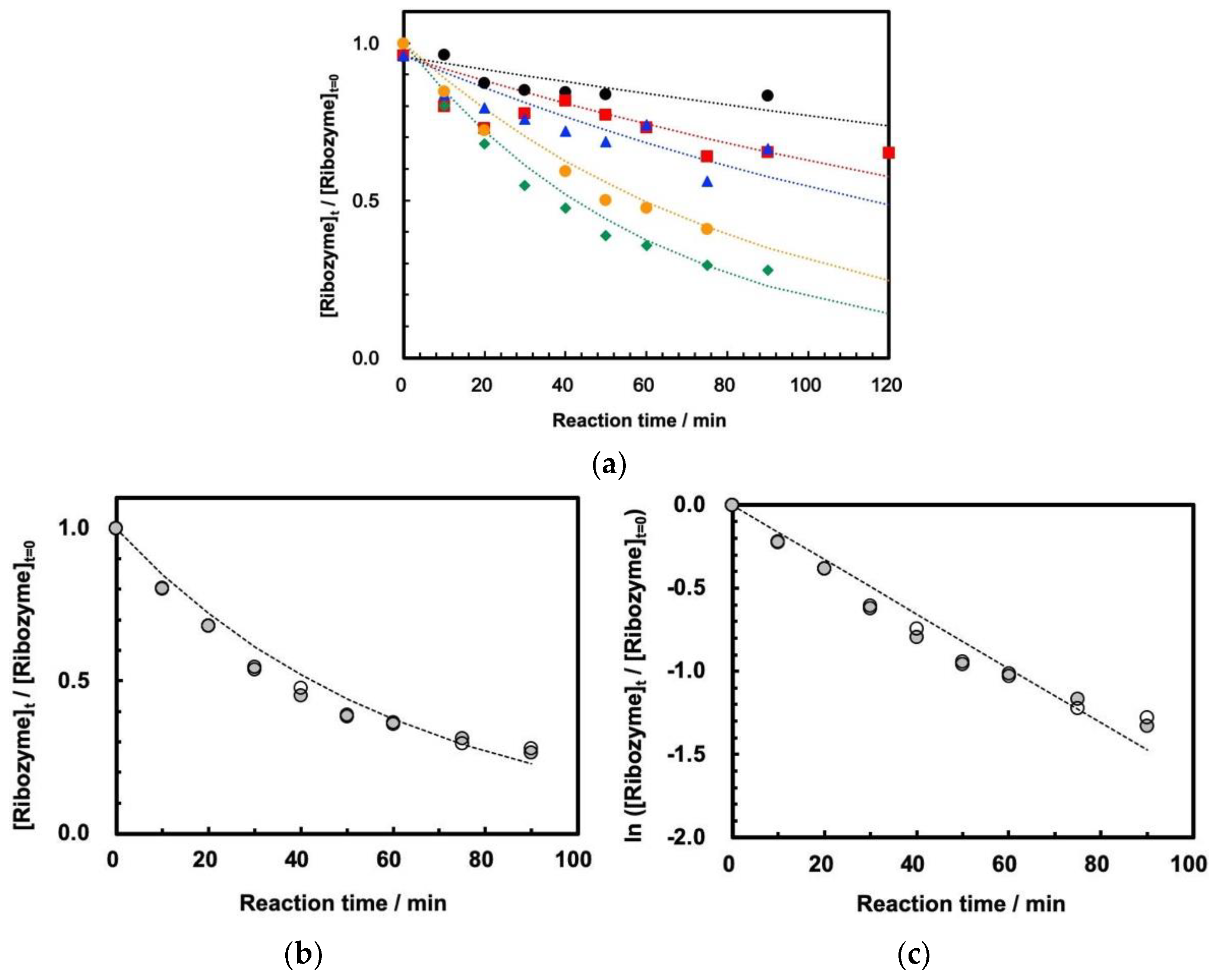
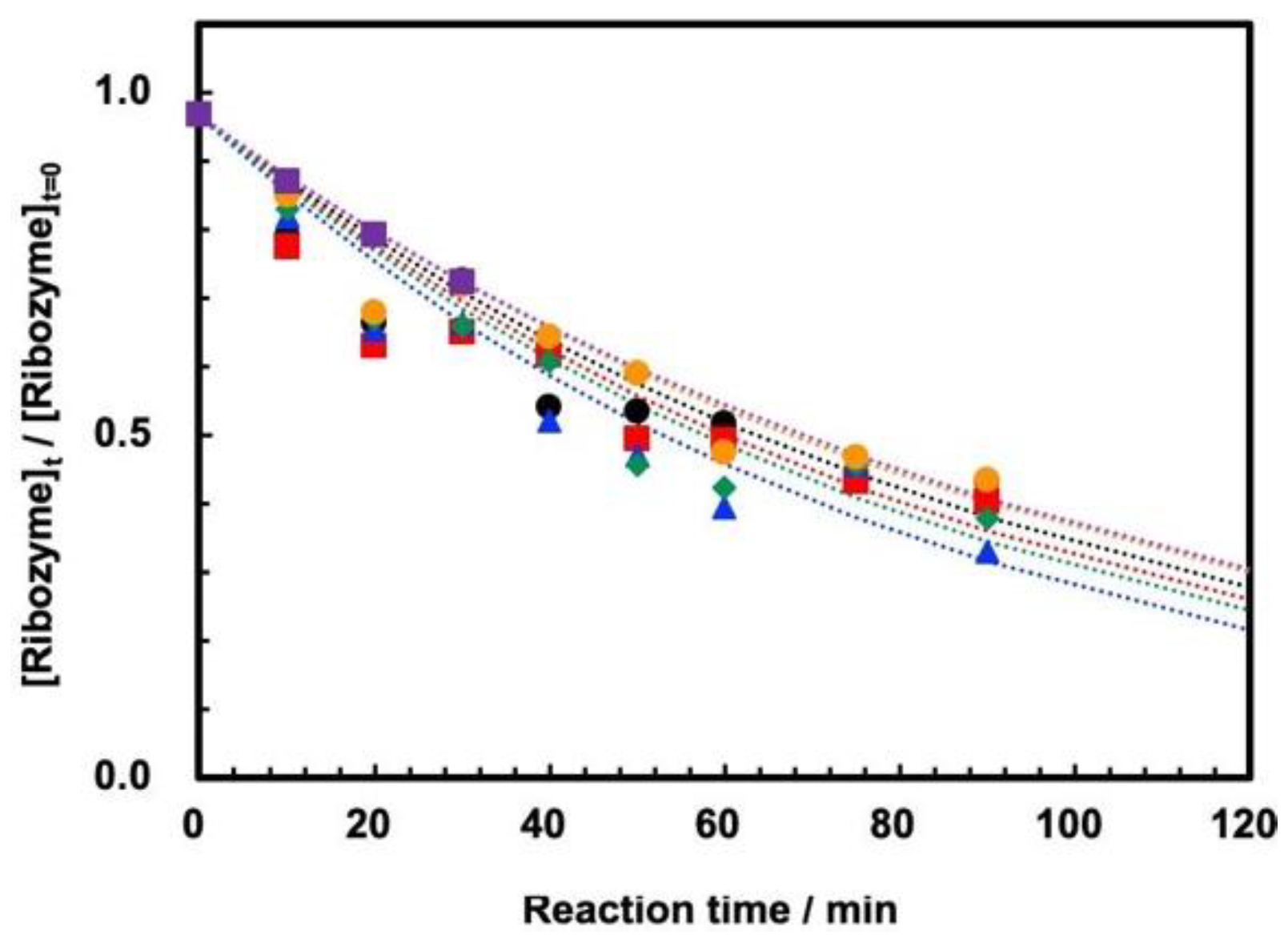
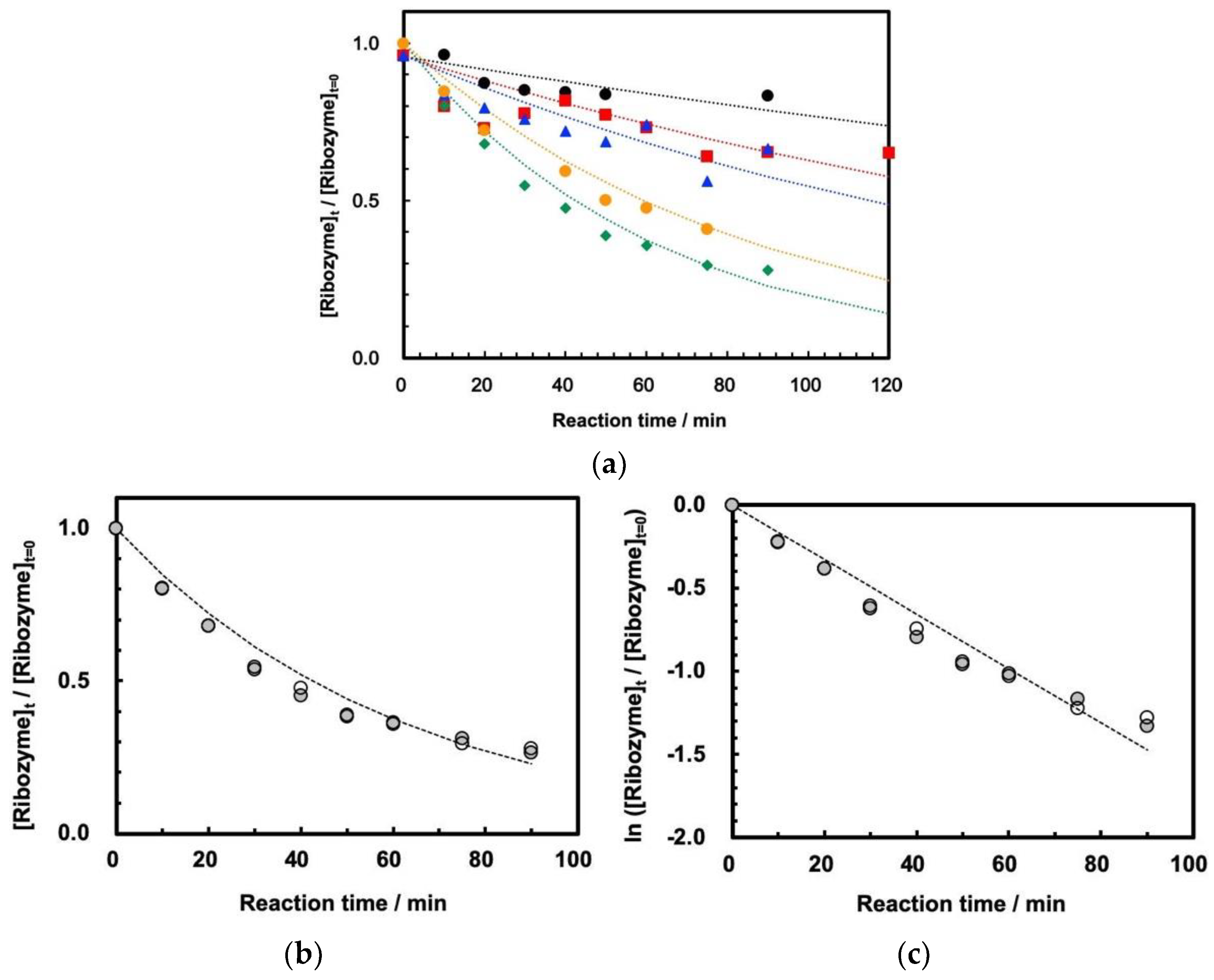
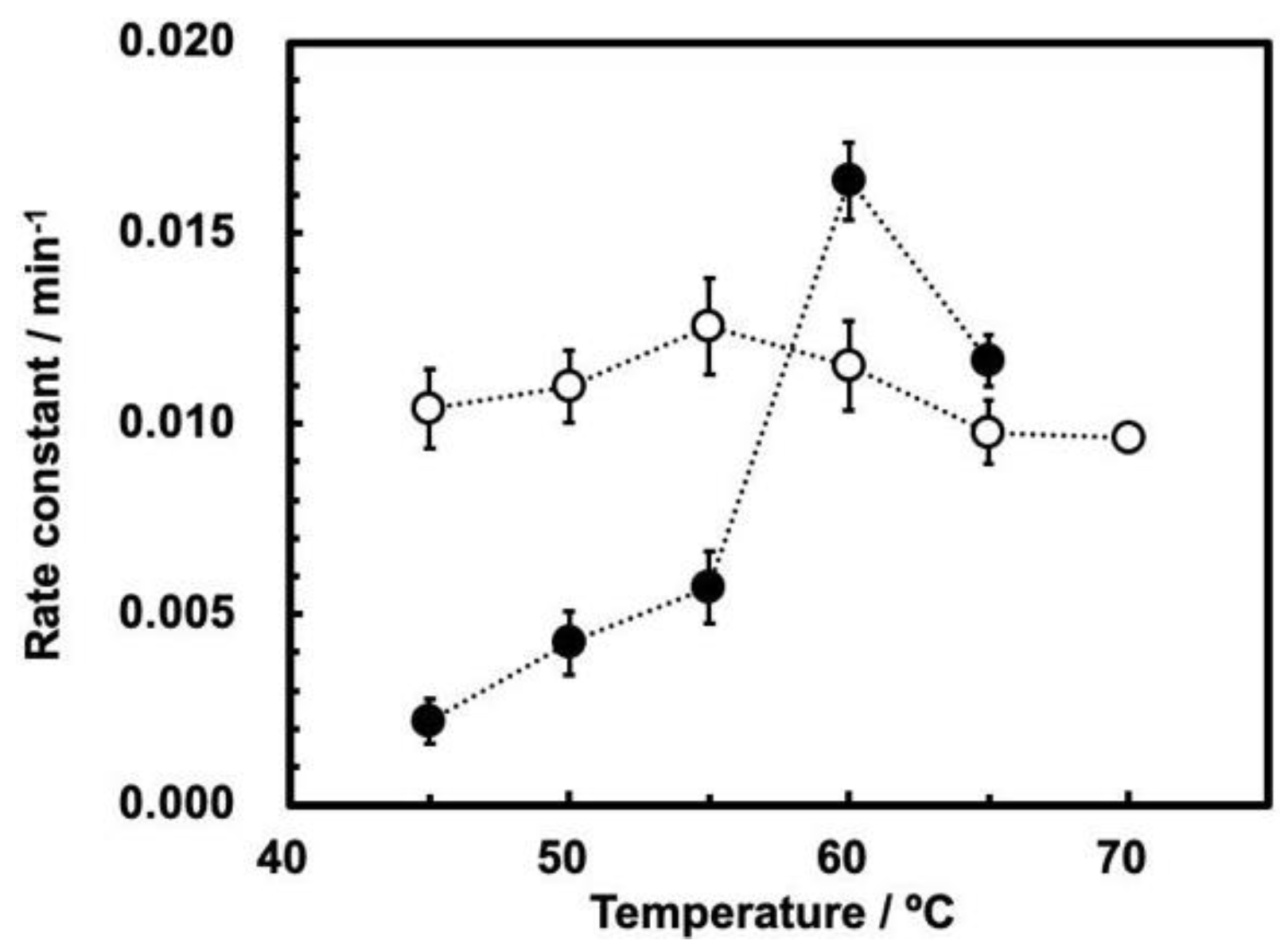
3.2. Cleavage of ASBVd(−):HHR at 0.1 MPa and 30 MPa at 45–70 °C
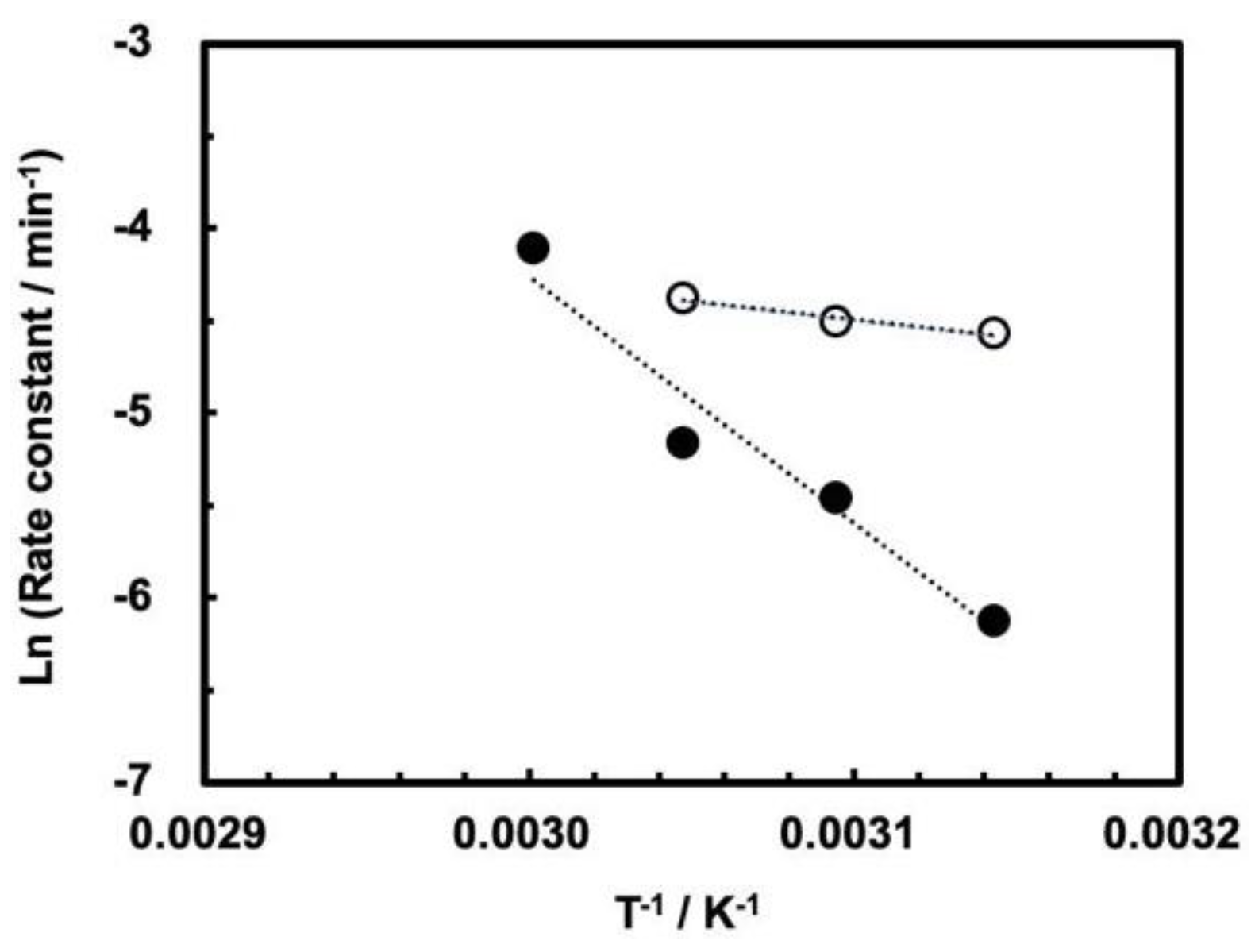
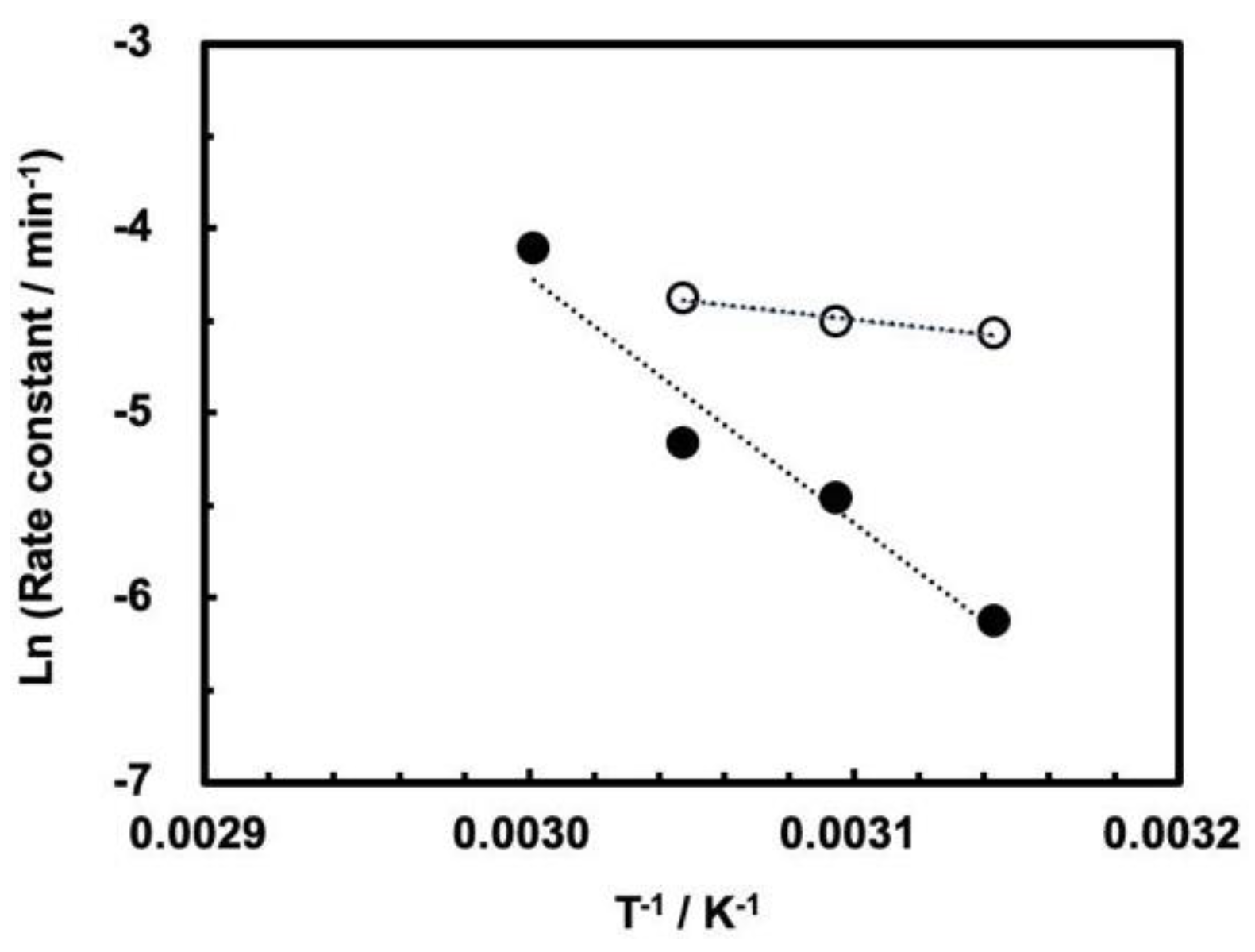
3.3. Influence of High Pressure on the Cleavage Behavior of ASBVd(−):HHR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilbert, W. Origins of life—The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Robertson, M.P.; Joyce, G.F. The Origins of the RNA World. Cold Spring Harb. Perspect. Biol. 2012, 4, a003608. [Google Scholar] [CrossRef]
- Allègre, C.J.; Manhès, G.; Göpel, C. The age of the earth. Geochim. Cosmochim. Acta 1995, 59, 1445–1456. [Google Scholar] [CrossRef]
- Schopf, J.W. Microfossils of the early Archean apex Chert: New evidence of the antiquity of life. Science 1993, 260, 640–646. [Google Scholar] [CrossRef]
- Mojzsis, S.J.; Arrhenius, G.; McKeegan, K.D.; Harrison, T.M.; Nutman, A.P.; Friend, C.R.L. Evidence for life on earth before 3800 million years ago. Nature 1996, 384, 55–59. [Google Scholar] [CrossRef]
- Maruyama, S.; Ikoma, M.; Genda, H.; Hirose, K.; Yokoyama, T.; Santosh, M. The naked planet earth: Most essential pre-requisite for the origin and evolution of life. Geosci. Front. 2013, 4, 141–165. [Google Scholar] [CrossRef]
- Kawamura, K. Hydrothermal microflow technology as a research tool for origin-of-life studies in extreme earth environments. Life 2017, 7, 37. [Google Scholar] [CrossRef]
- Maurel, M.-C.; Leclerc, F.; Hervé, G. Ribozyme chemistry: To be or not to be under high pressure. Chem. Rev. 2020, 120, 4898–4918. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K. Monitoring hydrothermal reactions on the millisecond time scale using a micro-tube flow reactor and kinetics of ATP hydrolysis for the RNA world hypothesis. Bull. Chem. Soc. Jpn. 2000, 73, 1805–1811. [Google Scholar] [CrossRef]
- Kaddour, H.; Vergne, J.; Guy Hervé, G.; Maurel, M.-C. High-pressure analysis of a hammerhead ribozyme from Chrysanthemum chlorotic mottle viroid reveals two different populations of self-cleaving molecule. FEBS J. 2011, 278, 3739–3747. [Google Scholar] [CrossRef] [PubMed]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High pressure effects on protein structure and function. Proteins: Structure. Genetics 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Smeller, L. Pressure–temperature phase diagrams of biomolecules. Biochim. Biophys. Acta-Protein Struct. 2002, 1595, 11–29. [Google Scholar] [CrossRef]
- Giuliodori, A.M.; Di Pietro, F.; Marzi, S.; Masquida, B.; Wagner, R.; Romby, P.; Gualerzi, C.O.; Pon, C.L. The cspA mRNA is a thermosensor that modulates translation of the cold-shock protein cspA. Mol. Cell 2010, 37, 21–33. [Google Scholar] [CrossRef]
- Penchovsky, R. Computational design of allosteric ribozymes as molecular biosensors. Biotechnol. Adv. 2014, 32, 1015–1027. [Google Scholar] [CrossRef]
- Kaddour, H.; Lucchi, H.; Hervé, G.; Vergne, J.; Maurel, M.-C. Kinetic Study of the Avocado Sunblotch Viroid Self-Cleavage Reaction Reveals Compensatory Effects between High-Pressure and High-Temperature: Implications for Origins of Life on Earth. Biology 2021, 10, 720. [Google Scholar] [CrossRef]
- Kawamura, K. Development of micro-flow hydrothermal monitoring systems and their applications to the origin of life study on earth. Anal. Sci. 2011, 27, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Hatanaka, T.; Hamahiga, K.; Ueshima, M.; Nakai, K. In situ UV–VIS–NIR spectrophotometric detection system as a research tool for environment-friendly chemical processes. Environ. Technol. Innov. 2019, 15, 100410. [Google Scholar] [CrossRef]
- Aida, T.M.; Sato, Y.; Watanabe, M.; Tajima, K.; Nonaka, T.; Hattori, H.; Arai, K. Dehydration of D-glucose in high temperature water at pressures up to 80 MPa. J. Super. Fluids 2007, 40, 381–388. [Google Scholar] [CrossRef]
- El-Murr, N.; Maurel, M.-C.; Rihova, M.; Vergne, J.; Hervé, G.; Kato, M.; Kawamura, K. Behavior of a hammerhead ribozyme in aqueous solution at medium to high temperatures. Naturwissenschaften 2012, 99, 731–738. [Google Scholar] [CrossRef]
- Kawamura, K.; Maeda, J. Kinetics and Activation Parameter Analysis for the Prebiotic Oligocytidylate Formation on Na+-Montmorillonite at 0–100 °C. J. Phys. Chem. A 2008, 112, 8015–8023. [Google Scholar] [CrossRef]
- Kanavarioti, A. HPLC methods for purity evaluation of man-made single-stranded RNAs. Sci. Rep. 2019, 9, 1019. [Google Scholar] [CrossRef]
- Bereiter, R.; Himmelstoß, M.; Renard, E.; Mairhofer, E.; Egger, M.; Breuker, K.; Kreutz, C.; Ennifar, E.; Micura, R. Impact of 3-deazapurine nucleobases on RNA properties. Nucleic Acids Res. 2021, 49, 4281–4293. [Google Scholar] [CrossRef]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuböck, R.; Hofacker, I.L. The vienna RNA websuite. Nucleic Acids Res. 2008, 36, W70–W74. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.; Bernhart, S.H.; Zu Siederdissen, C.H.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- Kawamura, K. In situ UV-Vis detection of the association of water-soluble anionic porphyrin and aromatic bases in aqueous solution at high temperatures using a capillary flow hydrothermal reactor system. Anal. Sci. 2003, 19, 1199–1202. [Google Scholar] [CrossRef]
- Dufour, E.; Hervé, G.; Haertle, T. Hydrolysis of β-lactoglobulin by thermolysin and pepsin under high hydrostatic pressure. Biopolymers 1995, 35, 475–483. [Google Scholar] [CrossRef]
- Northrop, D.B. Effects of high pressure on enzymatic activity. Biochim. Biophy. Acta 2002, 1595, 71–79. [Google Scholar] [CrossRef]
- Eisenmenger, M.J.; Reyes-De-Corcuera, J.I. High pressure enhancement of enzymes: A review. Enzym. Microbial. Technol. 2009, 45, 331–347. [Google Scholar] [CrossRef]
- Hervé;, G.; Evans, H.G.; Fernado, R.; Patel, C.; Hachem, F.; Evans, D.R. Activation of latent dihydroorotase from Aquifex aeolicus by pressure. J. Biol. Chem. 2017, 292, 629–637. [Google Scholar] [CrossRef]
- Stage-Zimmermann, T.K.; Uhlenbeck, O.C. Hammerhead ribozyme kinetics. RNA 1998, 4, 875–889. [Google Scholar] [CrossRef] [PubMed]
- Scott, W.G.; Horan, L.H.; Martick, M. The Hammerhead Ribozyme: Structure, Catalysis, and Gene Regulation. Prog. Mol. Biol. Trans. Sci. 2013, 120, 1–23. [Google Scholar] [CrossRef]
- Takagi, Y.; Taira, K. Temperature-dependent change in the rate-determining step in a reaction catalyzed by a hammerhead ribozyme. FEBS Lett. 1995, 361, 273–276. [Google Scholar] [CrossRef]
- Hertel, K.J.; Uhlenbeck, O.C. The internal equilibrium of the hammerhead ribozyme reaction. Biochemistry 1995, 34, 1744–1749. [Google Scholar] [CrossRef]
- Saha, R.; Pohorille, A.; Chen, I.A. Molecular crowding and early evolution. Origins Life Evol. Biosph. 2014, 44, 319–324. [Google Scholar] [CrossRef]
- Tjhunga, K.F.; Shokhireva, M.N.; Horninga, D.P.; Joyce, G.F. An RNA polymerase ribozyme that synthesizes its own ancestor. Proc. Natl. Acad. Sci. USA 2020, 116, 2906–2913. [Google Scholar] [CrossRef]
- Müller, F.; Escobar, L.; Xu, F.; Węgrzyn, E.; Nainytė, M.; Amatov, T.; Chan, C.-Y.; Pichler, A.; Carell, T. A prebiotically plausible scenario of an RNA–peptide world. Nature 2022, 605, 279–284. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | First-Order Rate Constant (min−1) | |
|---|---|---|
| 0.1 MPa 1 | 30 MPa 2 | |
| 45 | (1.04 ± 0.07) × 10−2 | (2.18 ± 0.58) × 10−3 |
| 50 | (1.10 ± 0.09) × 10−2 | (4.25 ± 0.84) × 10−3 |
| 55 | (1.25 ± 0.13) × 10−2 | (5.69 ± 0.94) × 10−3 |
| 60 | (1.15 ± 0.12) × 10−2 | (1.64 ± 0.10) × 10−2 |
| 65 | (9.78 ± 0.84) × 10−3 | (1.17 ± 0.07) × 10−2 |
| 70 | (9.62 ± 0.23) × 10−3 | NA |
| Pressure (MPa) | Ea (kJ mol−1) |
|---|---|
| 0.1 | 16.3 ± 4.0 |
| 30 | 111.5 ± 18.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawamura, K.; Ogawa, M.; Konagaya, N.; Maruoka, Y.; Lambert, J.-F.; Ter-Ovanessian, L.M.P.; Vergne, J.; Hervé, G.; Maurel, M.-C. A High-Pressure, High-Temperature Flow Reactor Simulating the Hadean Earth Environment, with Application to the Pressure Dependence of the Cleavage of Avocado Viroid Hammerhead Ribozyme. Life 2022, 12, 1224. https://doi.org/10.3390/life12081224
Kawamura K, Ogawa M, Konagaya N, Maruoka Y, Lambert J-F, Ter-Ovanessian LMP, Vergne J, Hervé G, Maurel M-C. A High-Pressure, High-Temperature Flow Reactor Simulating the Hadean Earth Environment, with Application to the Pressure Dependence of the Cleavage of Avocado Viroid Hammerhead Ribozyme. Life. 2022; 12(8):1224. https://doi.org/10.3390/life12081224
Chicago/Turabian StyleKawamura, Kunio, Mari Ogawa, Noriko Konagaya, Yoshimi Maruoka, Jean-François Lambert, Louis M. P. Ter-Ovanessian, Jacques Vergne, Guy Hervé, and Marie-Christine Maurel. 2022. "A High-Pressure, High-Temperature Flow Reactor Simulating the Hadean Earth Environment, with Application to the Pressure Dependence of the Cleavage of Avocado Viroid Hammerhead Ribozyme" Life 12, no. 8: 1224. https://doi.org/10.3390/life12081224
APA StyleKawamura, K., Ogawa, M., Konagaya, N., Maruoka, Y., Lambert, J.-F., Ter-Ovanessian, L. M. P., Vergne, J., Hervé, G., & Maurel, M.-C. (2022). A High-Pressure, High-Temperature Flow Reactor Simulating the Hadean Earth Environment, with Application to the Pressure Dependence of the Cleavage of Avocado Viroid Hammerhead Ribozyme. Life, 12(8), 1224. https://doi.org/10.3390/life12081224