
Characterization of NKG2-A/-C, Kir and CD57 on NK Cells Stimulated with pp65 and IE-1 Antigens in Patients Awaiting Lung Transplant

 ,
,  ,
, 
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
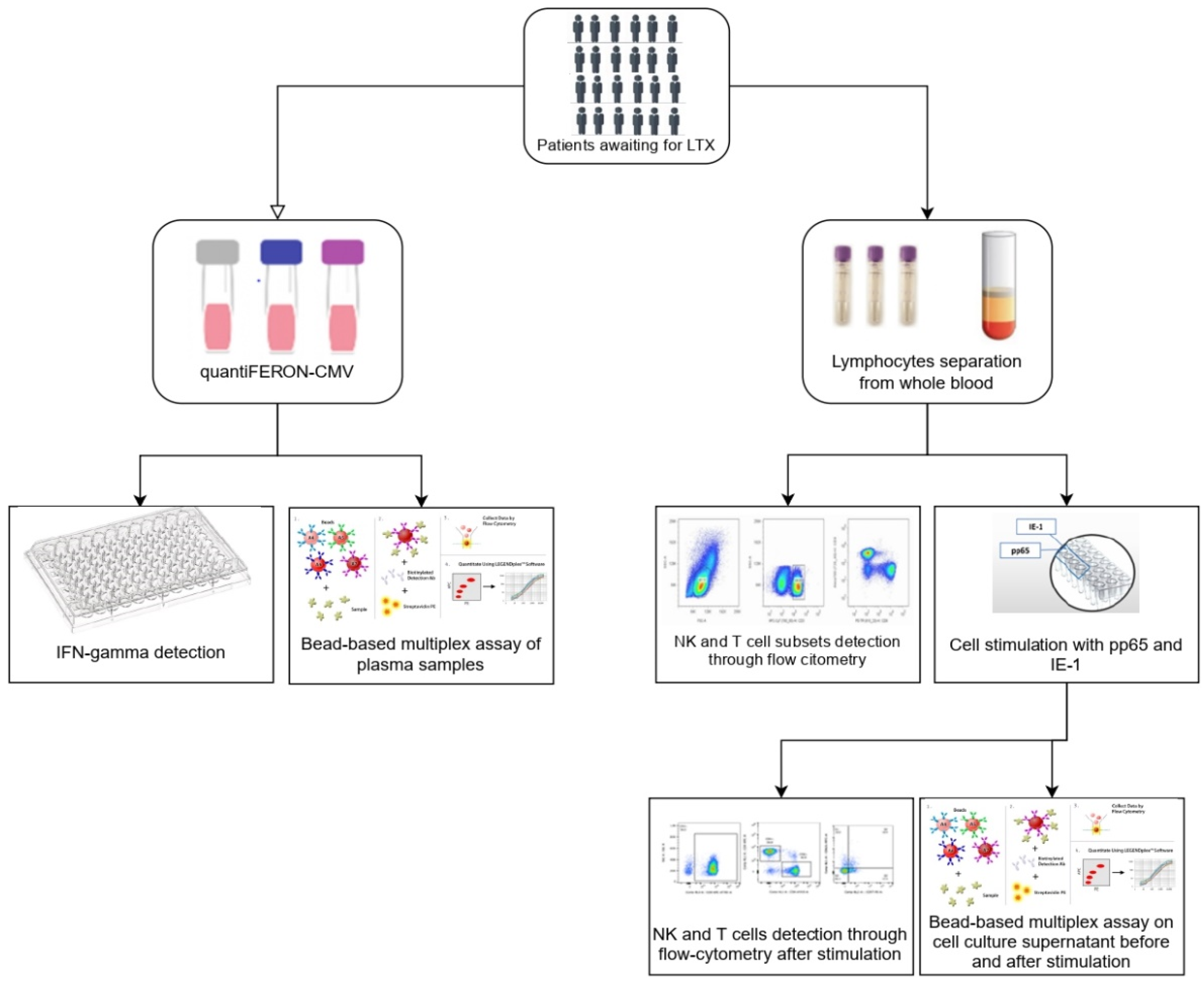
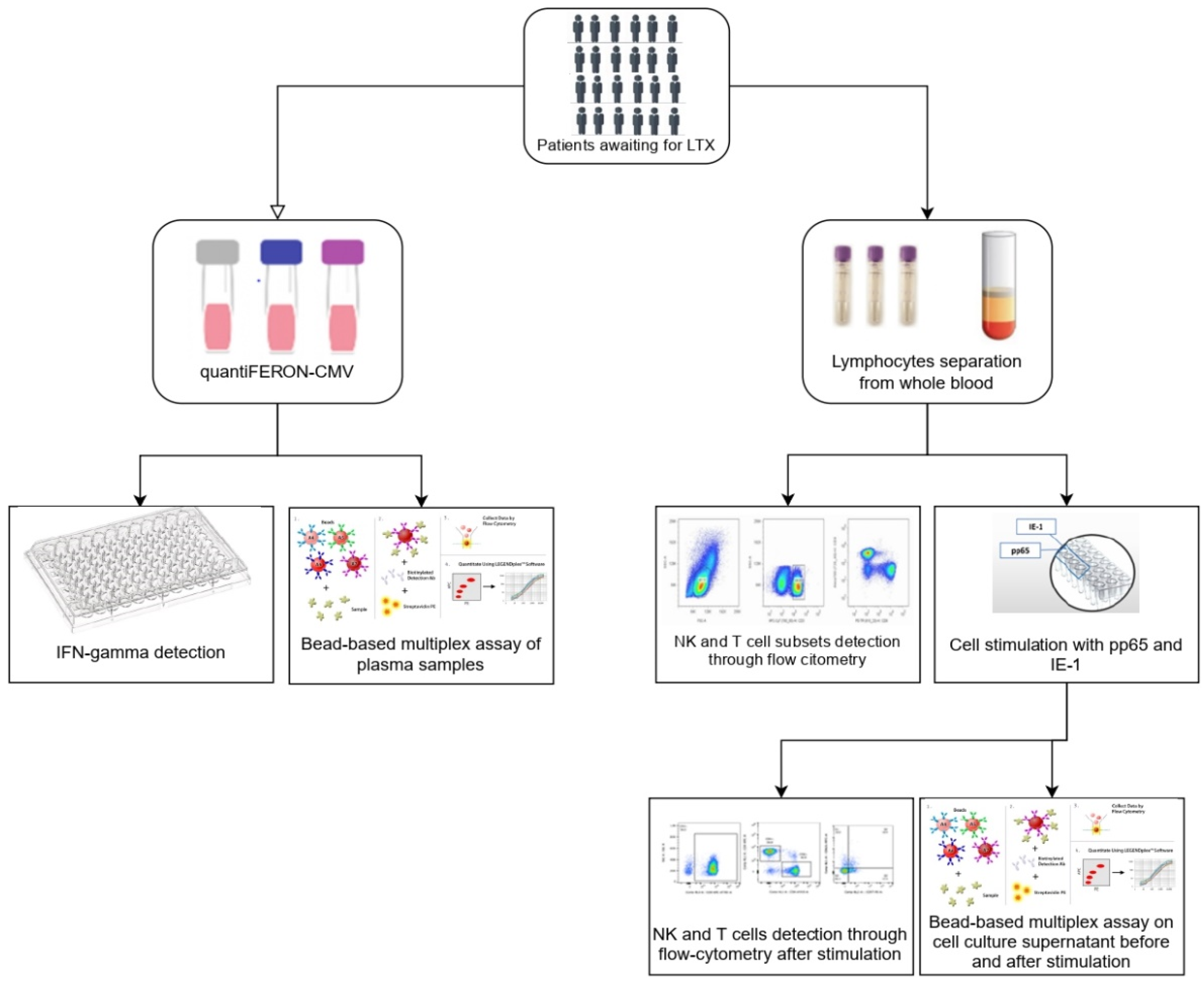
2. Materials and Methods
2.1. Patients and Study Design
2.2. QuantiFERON-CMV Assay and INF-γ Detection
2.3. Preparation and Storage of PBMCs
2.4. Reconstitution of PepTivator® CMV pp65 and IE-1 and Stimulation of PBMCs
2.5. Antibodies
2.6. Flow Cytometric Analysis
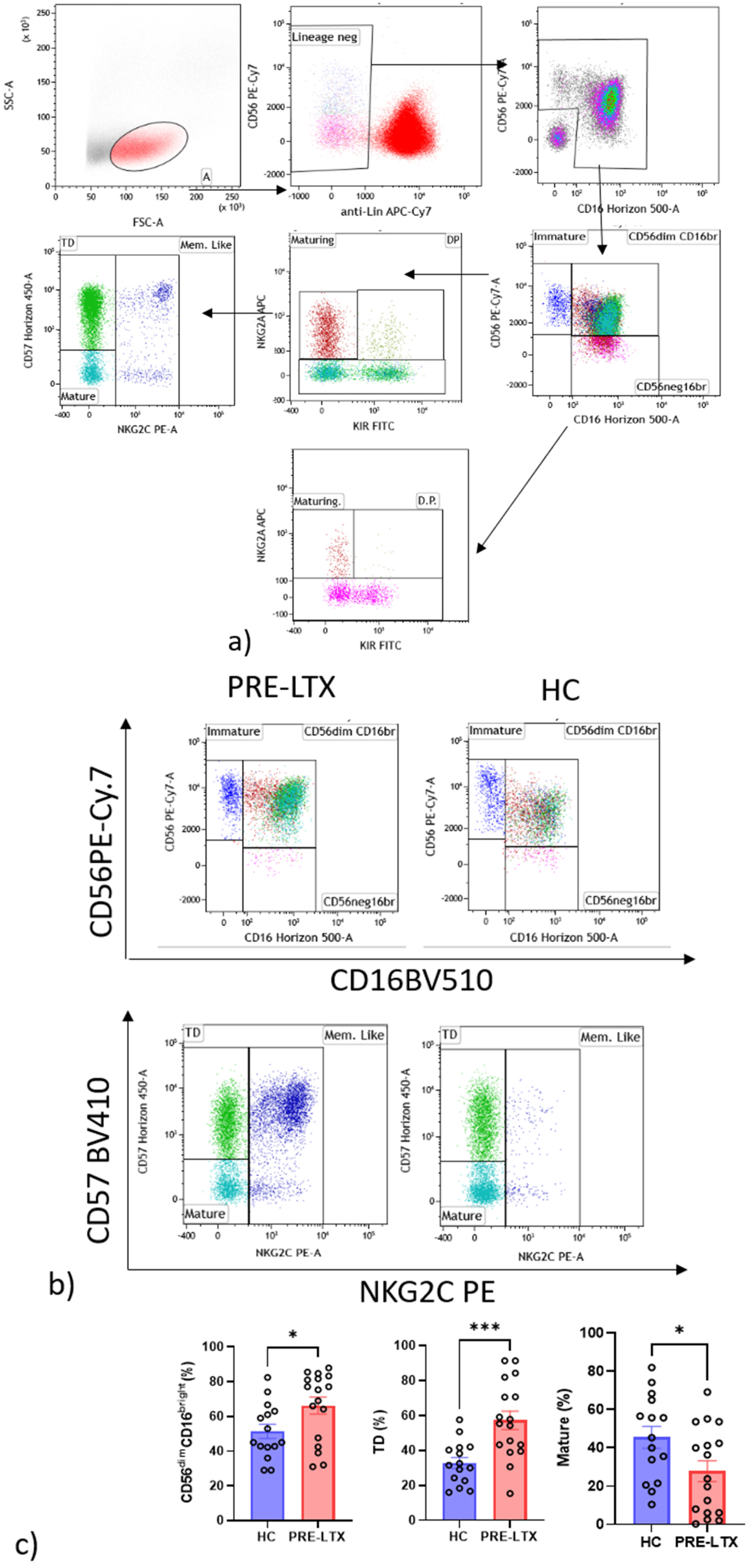
2.6.1. NK Cells Gate
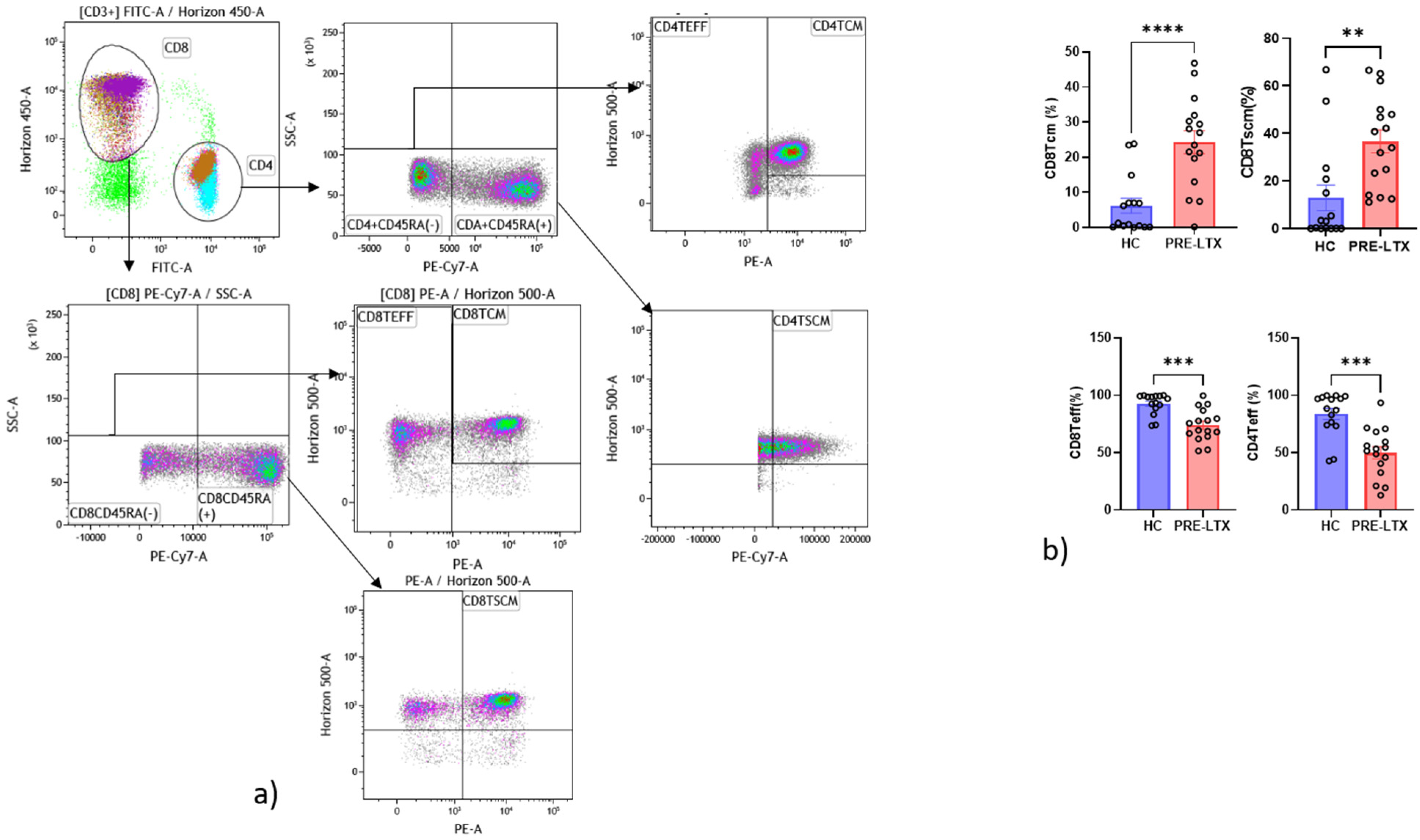
2.6.2. T Cells Gate
2.6.3. Cell Proliferation Dyes
2.7. Immunoassays
2.8. Statistical Analysis
3. Results
3.1. Study Population
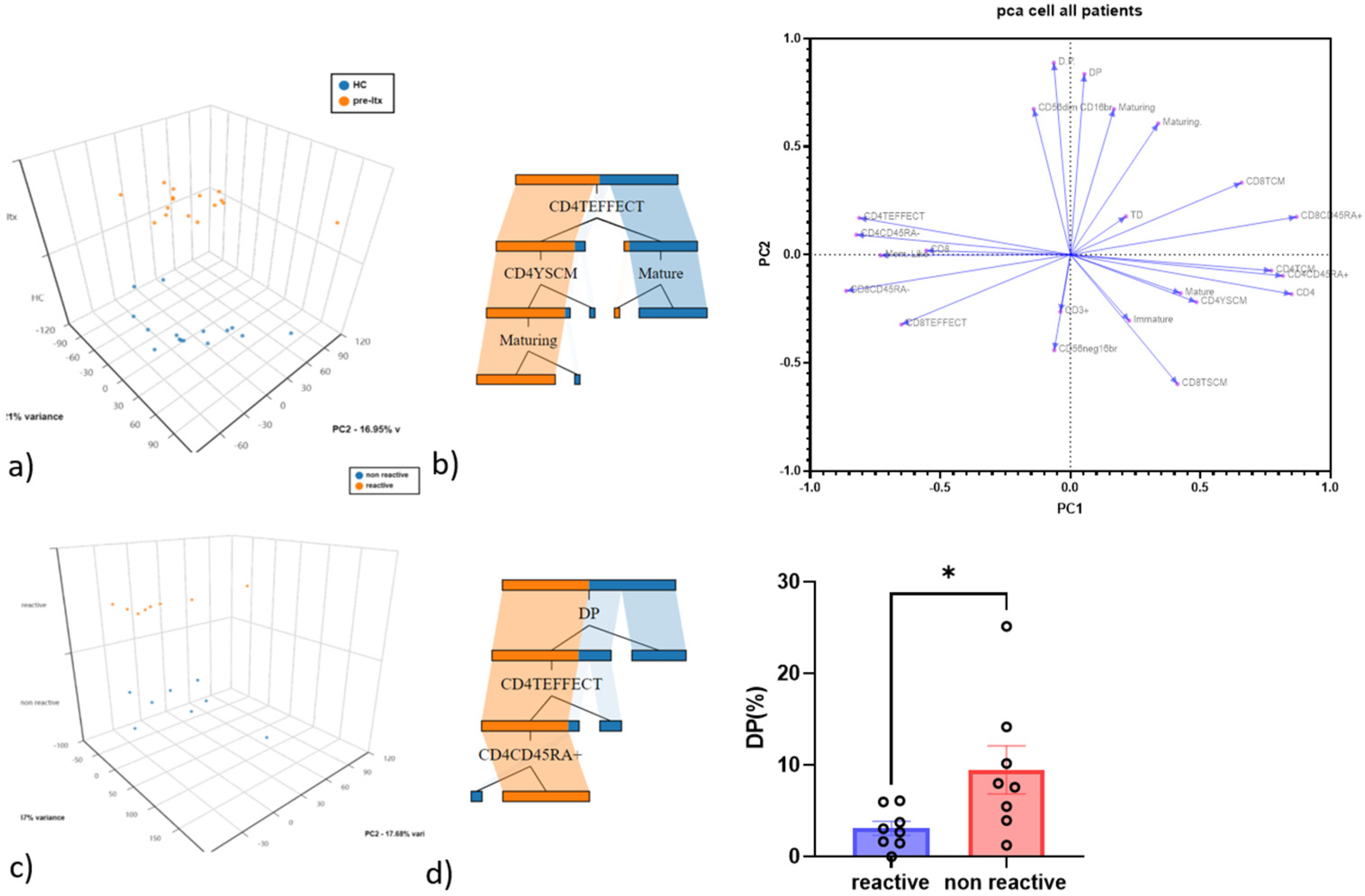
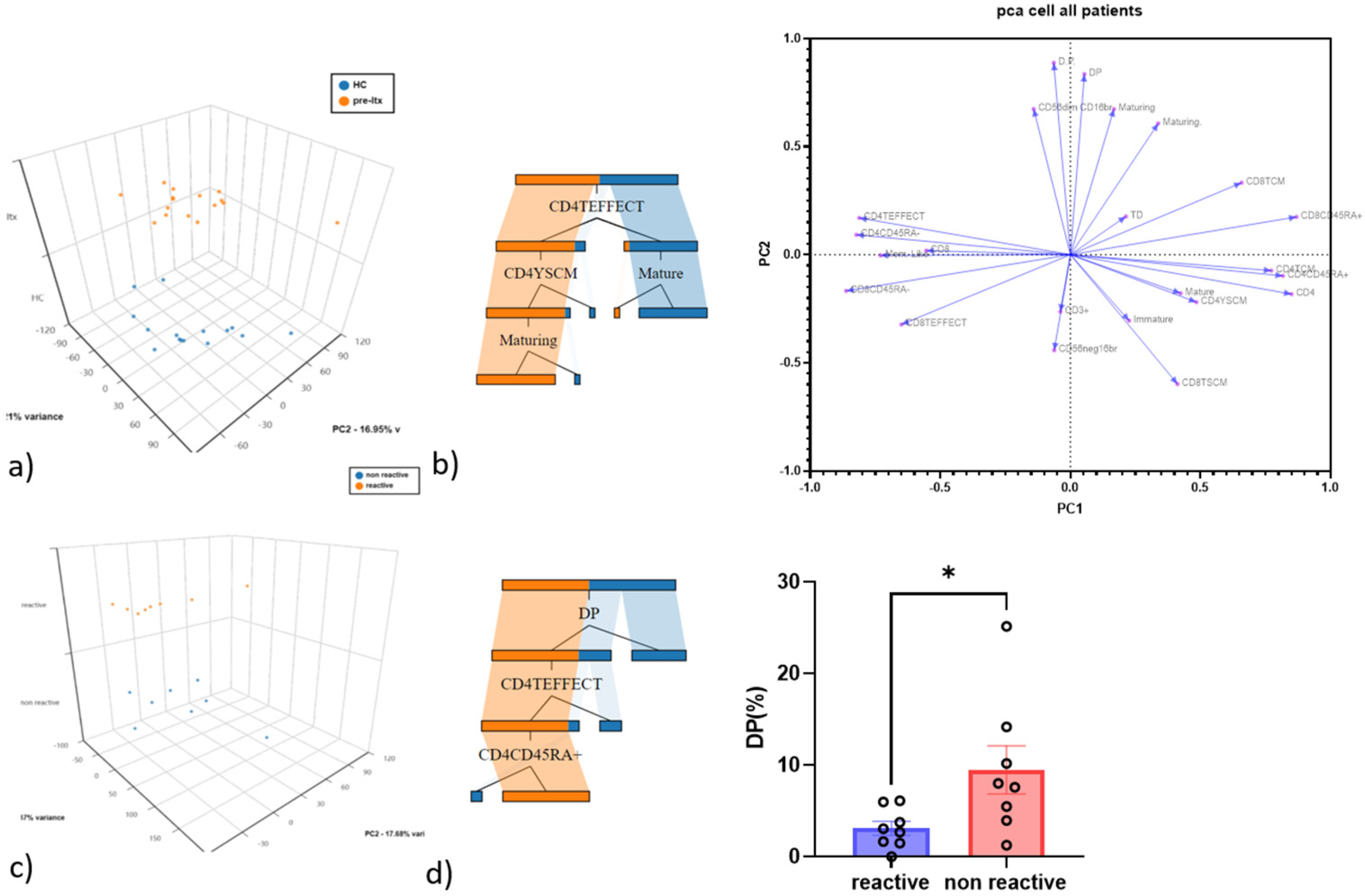
3.2. CD4 Teff and Double-Positive NK Cells as the Best Discriminatory Variables between HC and Pre-LTX and QF+ vs. QF− Patients
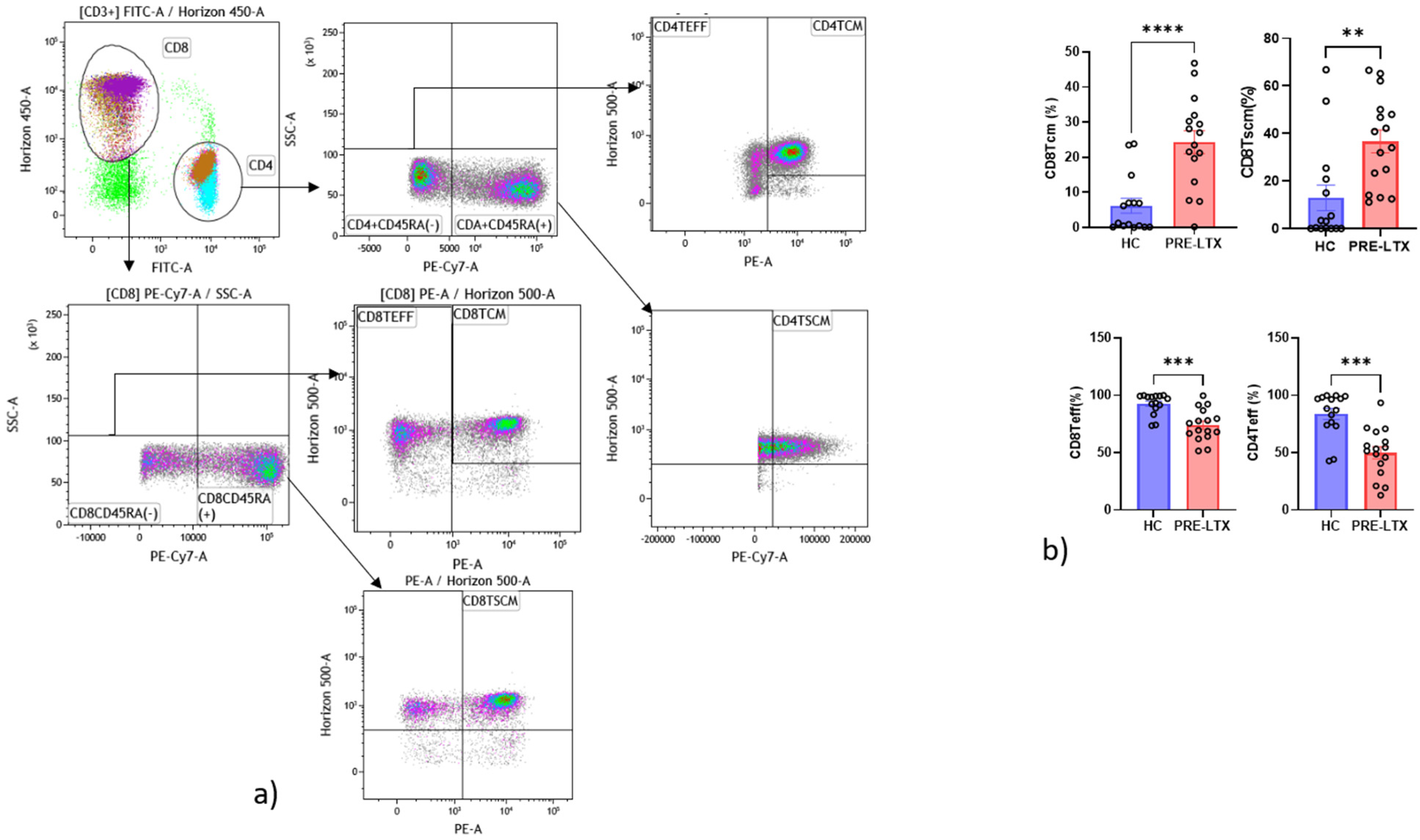
3.3. Comparison of NK and T Cell Counts in HC and Patients
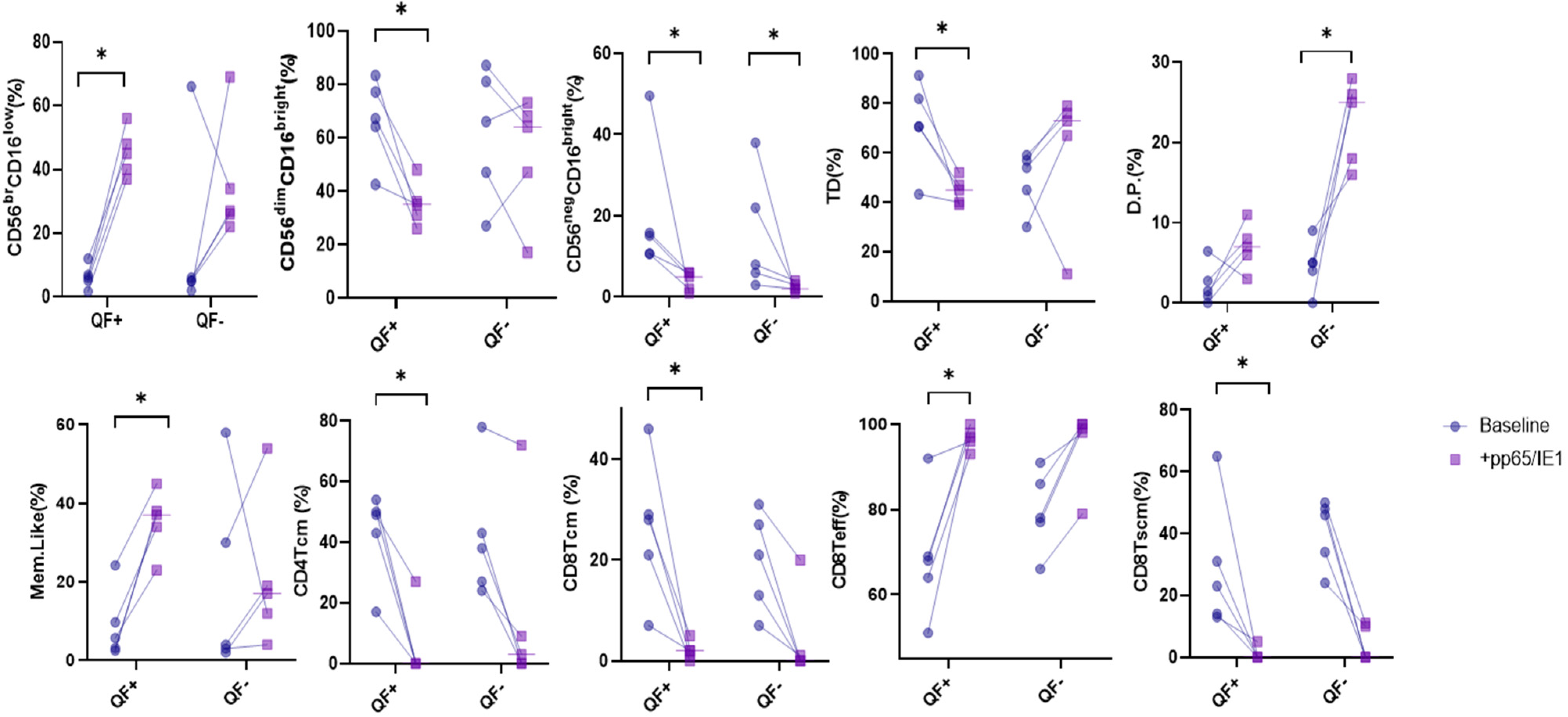
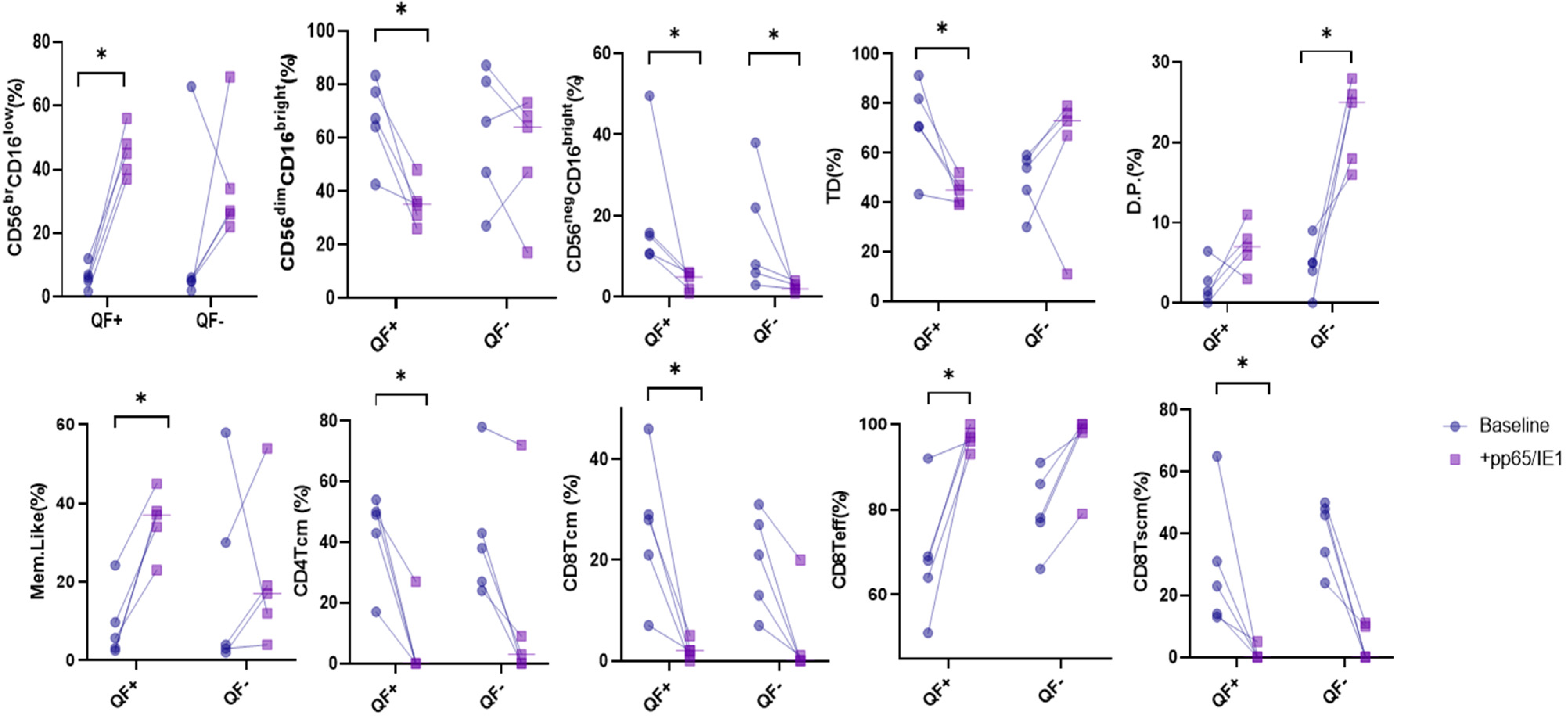
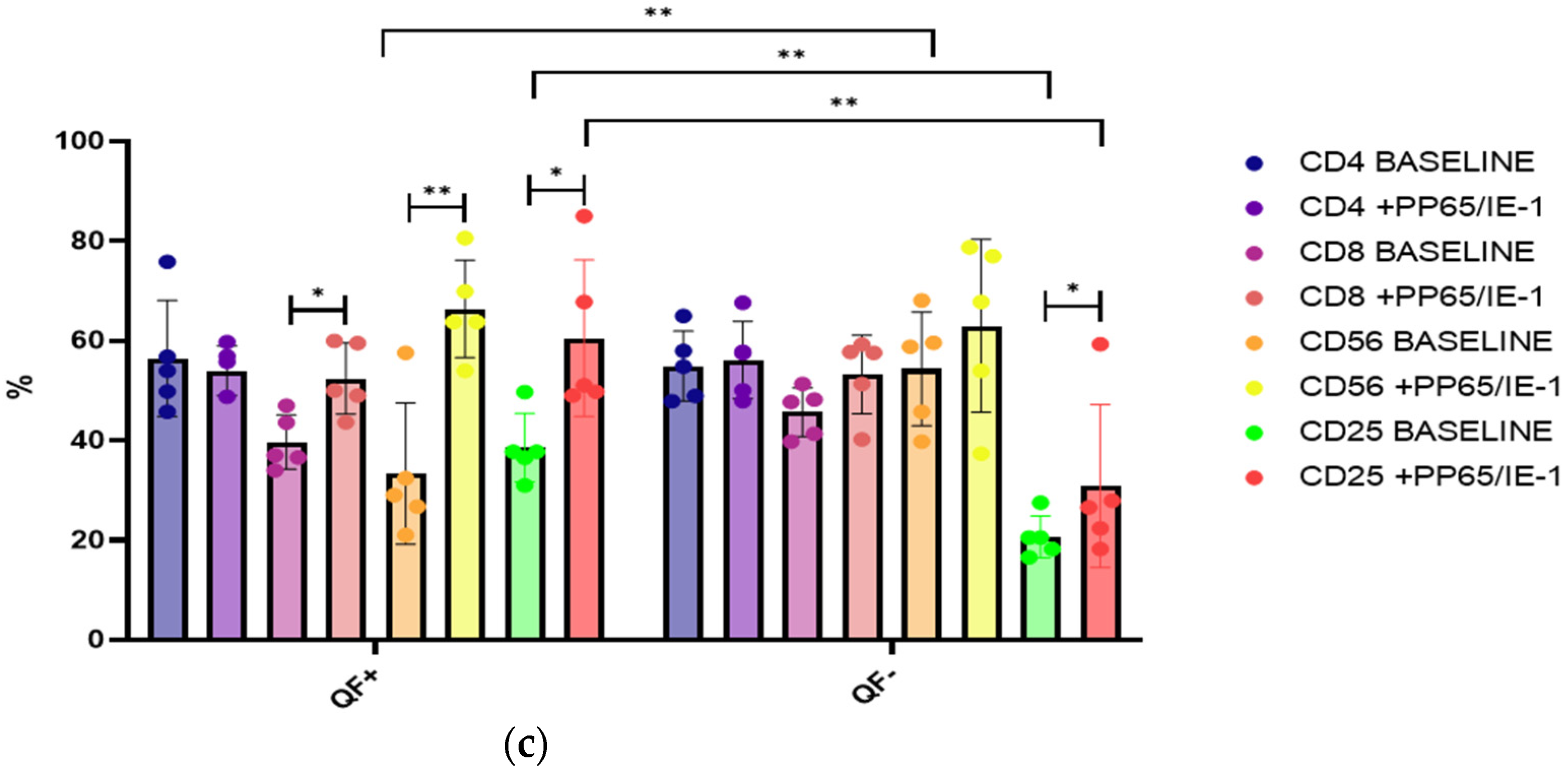
3.4. Stimulation with CMV Antigens, pp65 and IE-1
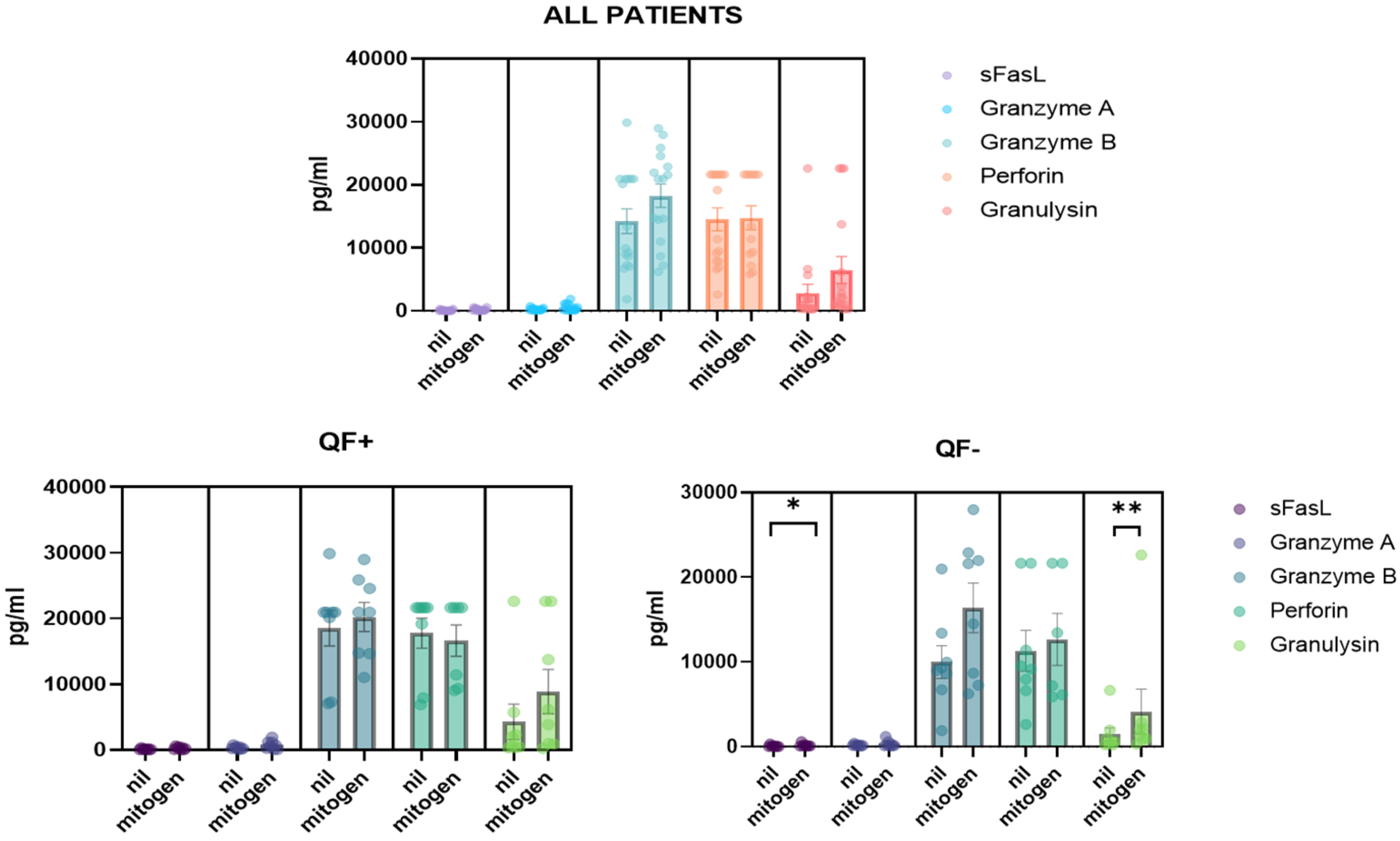
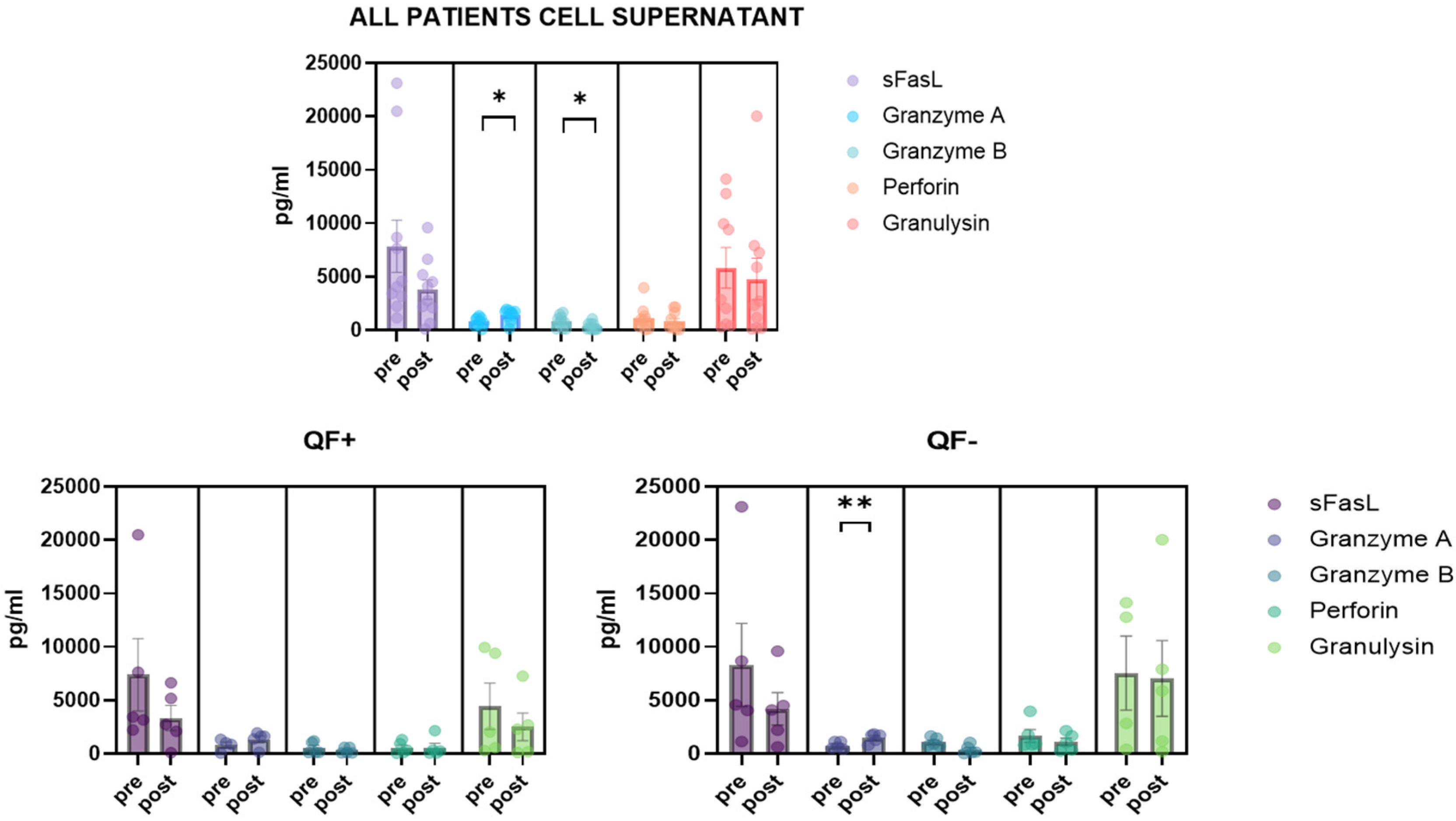
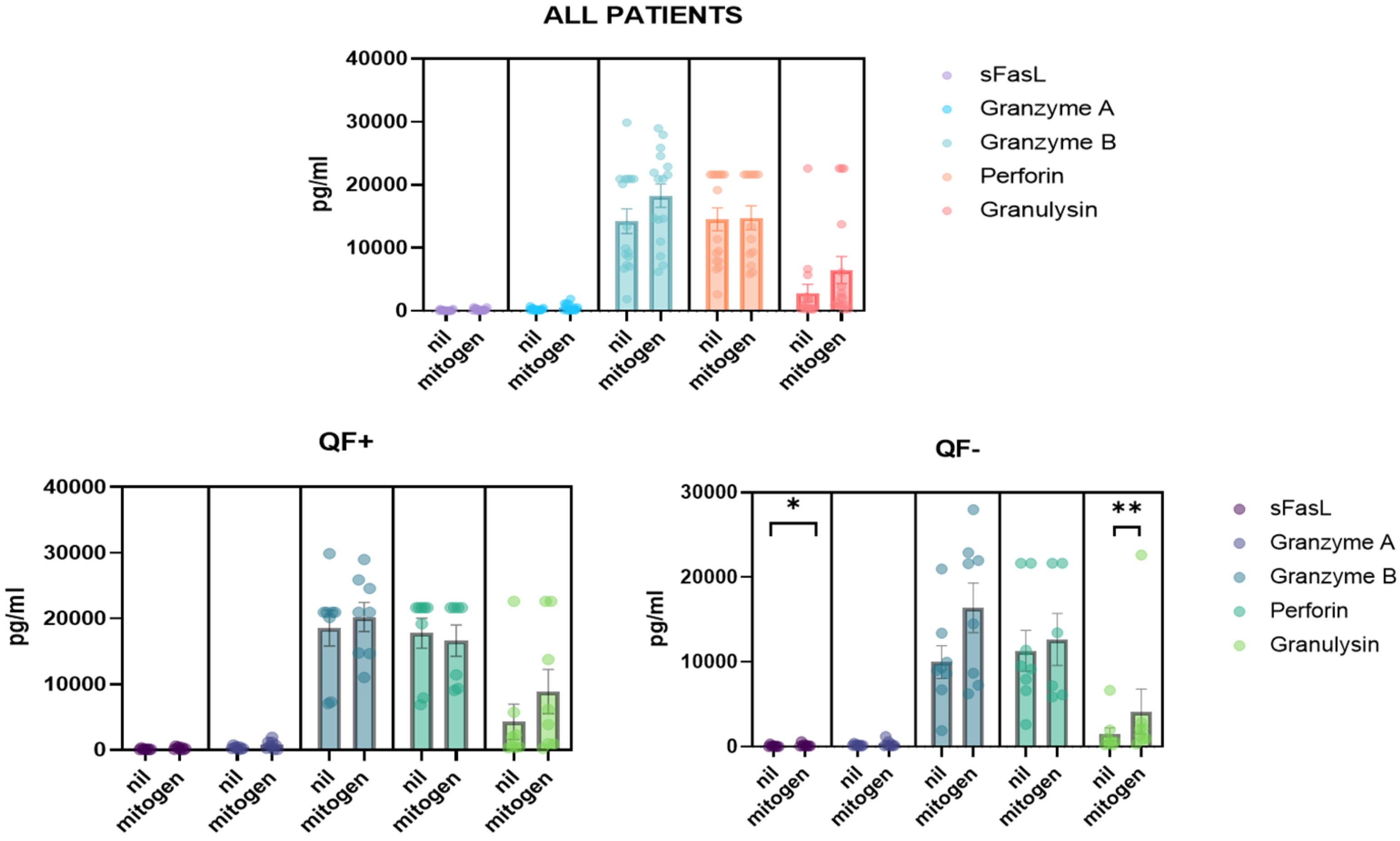
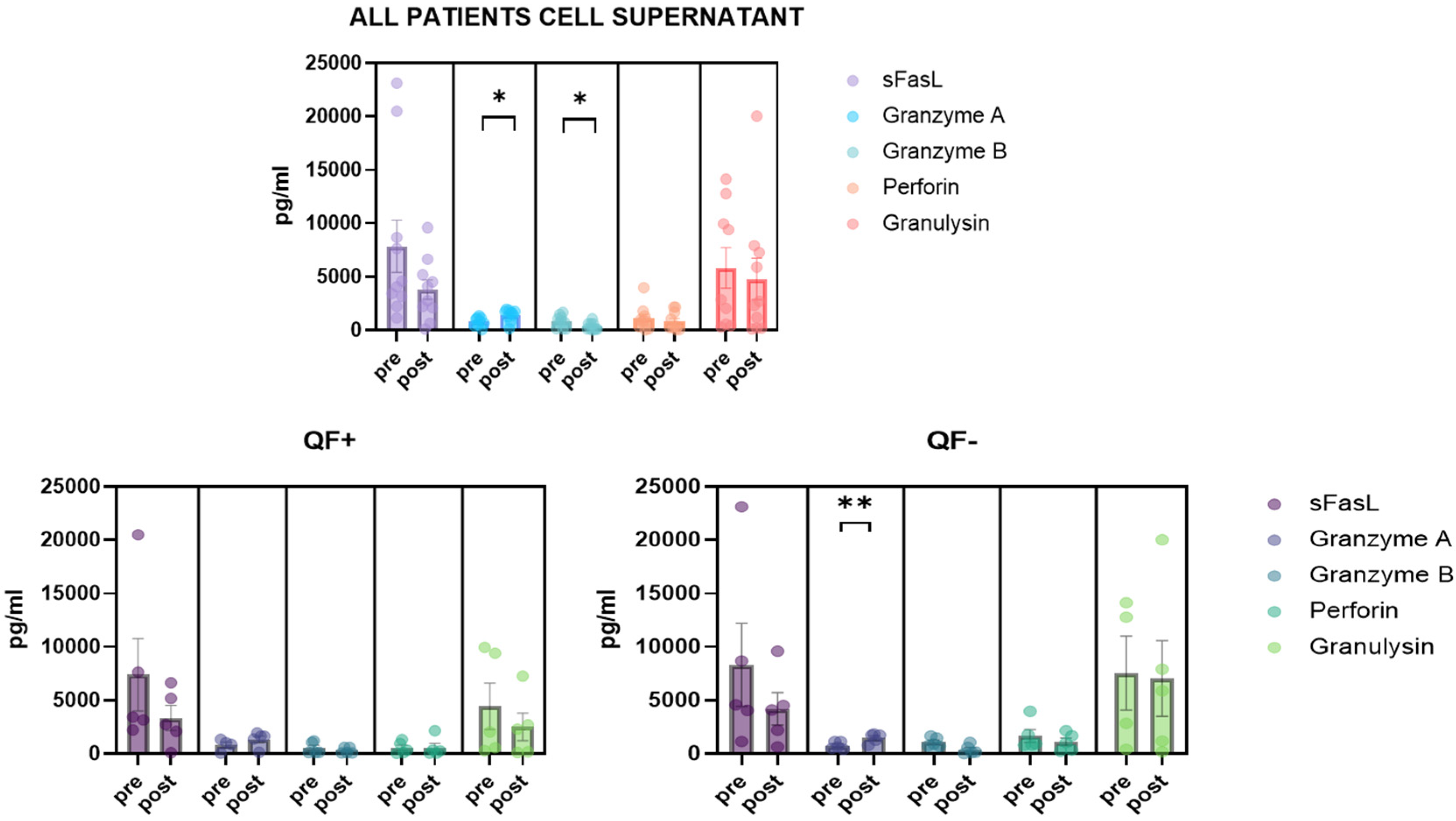
3.5. Detection of CD8/NK Cytokine Release in Serum Samples and Cell Culture Supernatant
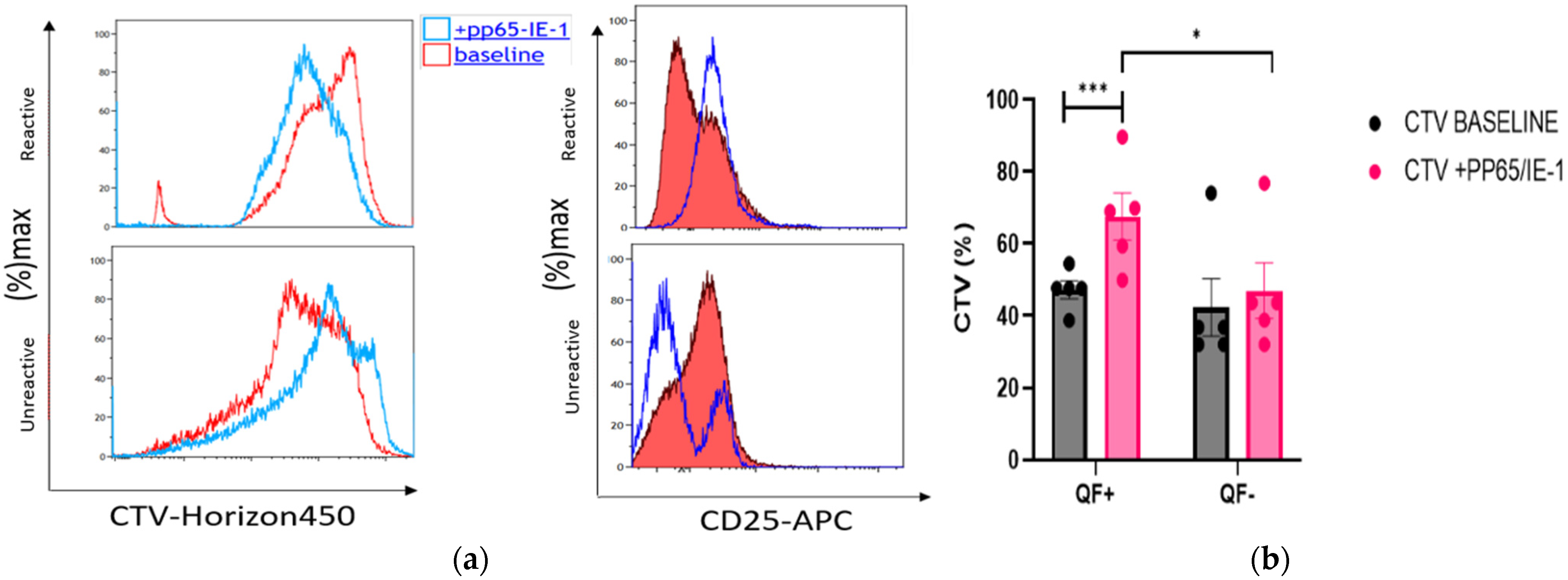
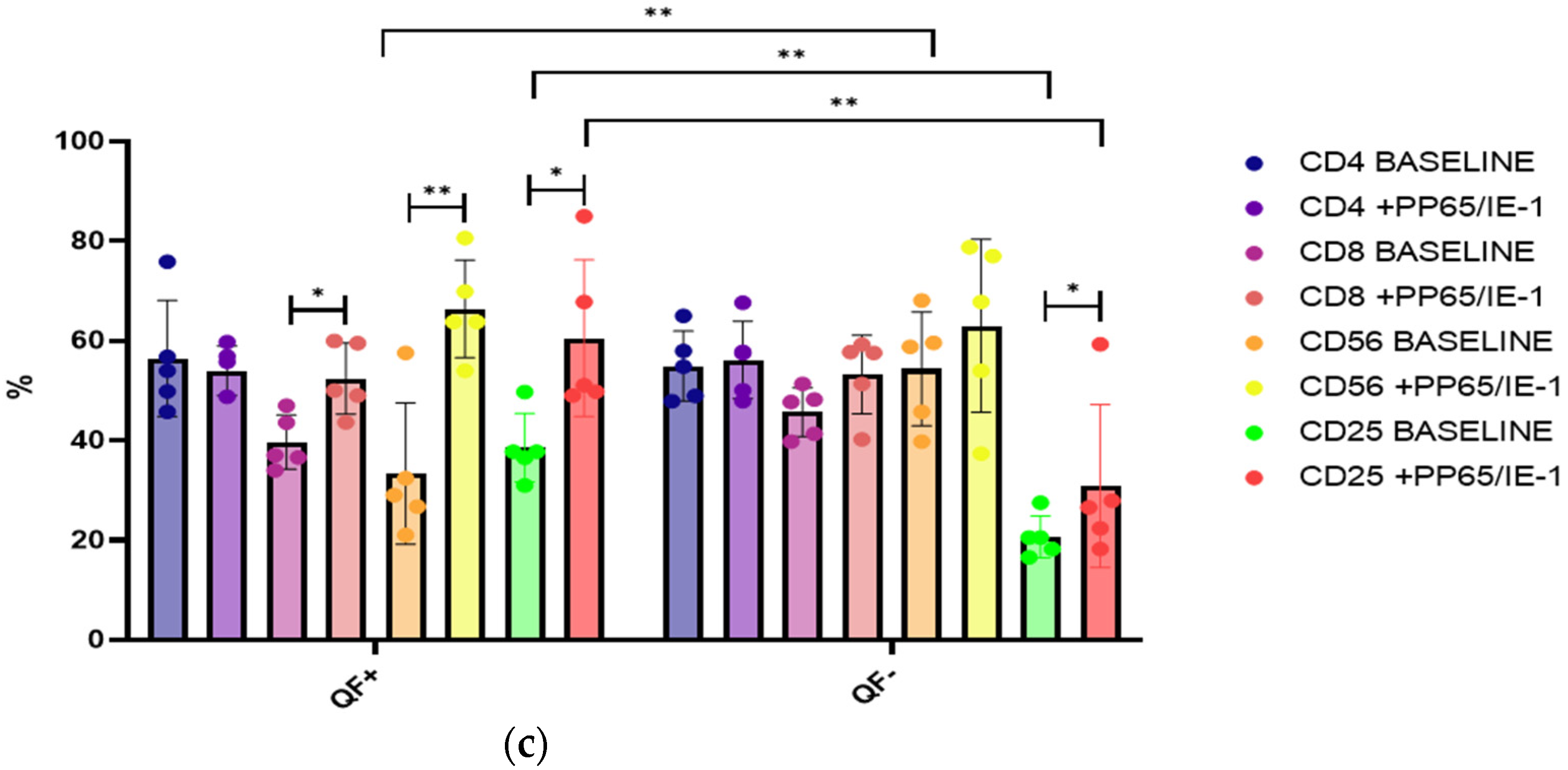
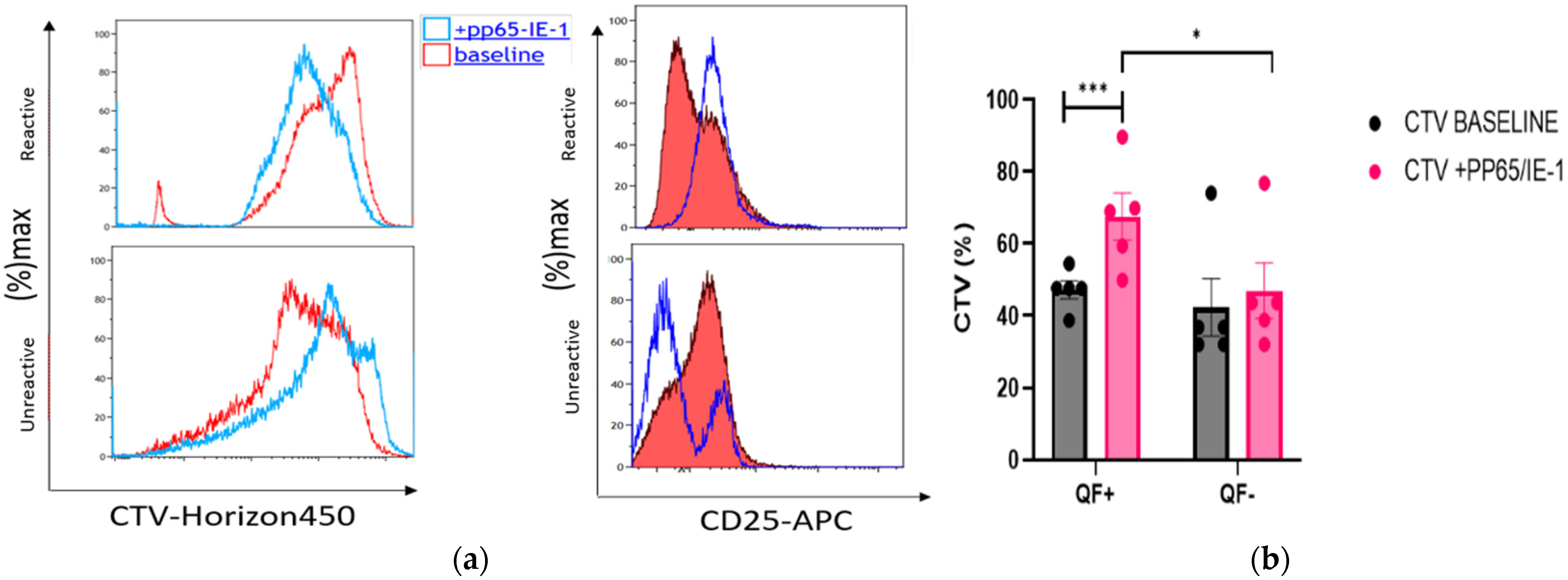
3.6. Rate of Cell Proliferation before and after Stimulation with pp65 and IE-1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bergantini, L.; d’Alessandro, M.; De Vita, E.; Perillo, F.; Fossi, A.; Luzzi, L.; Paladini, P.; Perrone, A.; Rottoli, P.; Sestini, P.; et al. Regulatory and Effector Cell Disequilibrium in Patients with Acute Cellular Rejection and Chronic Lung Allograft Dysfunction after Lung Transplantation: Comparison of Peripheral and Alveolar Distribution. Cells 2021, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Veit, T.; Munker, D.; Barton, J.; Milger, K.; Kauke, T.; Meiser, B.; Michel, S.; Zoller, M.; Nitschko, H.; Keppler, O.T.; et al. Letermovir in lung transplant recipients with cytomegalovirus infection: A retrospective observational study. Am. J. Transplant 2021, 21, 3449–3455. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.J.; Razonable, R.R. New developments in the management of cytomegalovirus infection after solid organ transplantation. Drugs 2010, 70, 965–981. [Google Scholar] [CrossRef] [PubMed]
- Marsico, C.; Kimberlin, D.W. Congenital Cytomegalovirus infection: Advances and challenges in diagnosis, prevention and treatment. Ital. J. Pediatr. 2017, 43, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girmenia, C.; Lazzarotto, T.; Bonifazi, F.; Patriarca, F.; Irrera, G.; Ciceri, F.; Aversa, F.; Citterio, F.; Cillo, U.; Cozzi, E.; et al. Assessment and prevention of cytomegalovirus infection in allogeneic hematopoietic stem cell transplant and in solid organ transplant: A multidisciplinary consensus conference by the Italian GITMO, SITO, and AMCLI societies. Clin. Transpl. 2019, 33, e13666. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, D.R.; Lanier, L.L.; Greenland, J.R. Natural killer cells in lung transplantation. Thorax 2019, 74, 397–404. [Google Scholar] [CrossRef] [Green Version]
- La Rosa, C.; Diamond, D.J. The immune response to human CMV. Future Virol. 2012, 7, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Bergantini, L.; Cameli, P.; d’Alessandro, M.; Vagaggini, C.; Refini, R.M.; Landi, C.; Pieroni, M.G.; Spalletti, M.; Sestini, P.; Bargagli, E. NK and NKT-like cells in granulomatous and fibrotic lung diseases. Clin. Exp. Med. 2019, 19, 487–494. [Google Scholar] [CrossRef]
- Castriconi, R.; Carrega, P.; Dondero, A.; Bellora, F.; Casu, B.; Regis, S.; Ferlazzo, G.; Bottino, C. Molecular Mechanisms Directing Migration and Retention of Natural Killer Cells in Human Tissues. Front. Immunol. 2018, 9, 2324. [Google Scholar] [CrossRef]
- Carrega, P.; Ferlazzo, G. Natural killer cell distribution and trafficking in human tissues. Front. Immunol. 2012, 3, 347. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Vergès, S.; Milush, J.M.; Pandey, S.; York, V.A.; Arakawa-Hoyt, J.; Pircher, H.; Norris, P.J.; Nixon, D.F.; Lanier, L.L. CD57 defines a functionally distinct population of mature NK cells in the human CD56dimCD16+ NK-cell subset. Blood 2010, 116, 3865–3874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- NKG2C + CD57 + Natural Killer Cell Expansion Parallels Cytomegalovirus-Specific CD8+ T Cell Evolution towards Senescence. Available online: https://www.hindawi.com/journals/jir/2016/7470124/ (accessed on 9 February 2022).
- Rousselière, A.; Delbos, L.; Bressollette, C.; Berthaume, M.; Charreau, B. Mapping and Characterization of HCMV-Specific Unconventional HLA-E-Restricted CD8 T Cell Populations and Associated NK and T Cell Responses Using HLA/Peptide Tetramers and Spectral Flow Cytometry. Int. J. Mol. Sci. 2021, 23, 263. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vergès, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.M.; Norris, P.J.; et al. Expansion of a unique CD57+NKG2Chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef] [Green Version]
- Mitrović, M.; Arapović, J.; Jordan, S.; Fodil-Cornu, N.; Ebert, S.; Vidal, S.M.; Krmpotić, A.; Reddehase, M.J.; Jonjić, S. The NK Cell Response to Mouse Cytomegalovirus Infection Affects the Level and Kinetics of the Early CD8+ T-Cell Response. J. Virol. 2012, 86, 2165–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousset, C.M.; Hobo, W.; Woestenenk, R.; Preijers, F.; Dolstra, H.; van der Waart, A.B. Comprehensive Phenotyping of T Cells Using Flow Cytometry. Cytom. A 2019, 95, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Jameson, S.C.; Masopust, D. Understanding Subset Diversity in T Cell Memory. Immunity 2018, 48, 214–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durek, P.; Nordström, K.; Gasparoni, G.; Salhab, A.; Kressler, C.; de Almeida, M.; Bassler, K.; Ulas, T.; Schmidt, F.; Xiong, J.; et al. Epigenomic Profiling of Human CD4+ T Cells Supports a Linear Differentiation Model and Highlights Molecular Regulators of Memory Development. Immunity 2016, 45, 1148–1161. [Google Scholar] [CrossRef] [Green Version]
- van Aalderen, M.C.; van den Biggelaar, M.; Remmerswaal, E.B.M.; van Alphen, F.P.J.; Meijer, A.B.; Ten Berge, I.J.M.; van Lier, R.A.W. Label-free Analysis of CD8+ T Cell Subset Proteomes Supports a Progressive Differentiation Model of Human-Virus-Specific T Cells. Cell Rep. 2017, 19, 1068–1079. [Google Scholar] [CrossRef] [Green Version]
- Aris, R.; Barbers, R.G.; Barst, R.; Baz, M.A.; de Boer, W.; Corris, P.A.; Dark, J.; Davis, R.D.; Eagan, J.J.; Egan, T.M.; et al. International Guidelines for the Selection of Lung Transplant Candidates. Am. J. Respir. Crit. Care Med. 1998, 158, 335–339. [Google Scholar]
- Del Zotto, G.; Marcenaro, E.; Vacca, P.; Sivori, S.; Pende, D.; Della Chiesa, M.; Moretta, F.; Ingegnere, T.; Mingari, M.C.; Moretta, A.; et al. Markers and function of human NK cells in normal and pathological conditions. Cytom. B Clin. Cytom. 2017, 92, 100–114. [Google Scholar] [CrossRef] [Green Version]
- Tzannou, I.; Watanabe, A.; Naik, S.; Daum, R.; Kuvalekar, M.; Leung, K.S.; Martinez, C.; Sasa, G.; Wu, M.; Gee, A.P.; et al. “Mini” bank of only 8 donors supplies CMV-directed T cells to diverse recipients. Blood Adv. 2019, 3, 2571–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehne, G.; Hasan, A.; Doubrovina, E.; Prockop, S.; Tyler, E.; Wasilewski, G.; O’Reilly, R.J. Immunotherapy with Donor T Cells Sensitized with Overlapping Pentadecapeptides for Treatment of Persistent Cytomegalovirus Infection or Viremia. Biol. Blood Marrow Transpl. 2015, 21, 1663–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunde, T.; Kirchner, A.; Hoffmeister, B.; Habedank, D.; Hetzer, R.; Cherepnev, G.; Proesch, S.; Reinke, P.; Volk, H.D.; Lehmkuhl, H.; et al. Protection from cytomegalovirus after transplantation is correlated with immediate early 1–specific CD8 T cells. J. Exp. Med. 2005, 201, 1031–1036. [Google Scholar] [CrossRef]
- Lugli, E.; Dominguez, M.H.; Gattinoni, L.; Chattopadhyay, P.K.; Bolton, D.L.; Song, K.; Klatt, N.R.; Brenchley, J.M.; Vaccari, M.; Gostick, E.; et al. Superior T memory stem cell persistence supports long-lived T cell memory. J. Clin. Investig. 2013, 123, 594–599. [Google Scholar] [CrossRef] [Green Version]
- Opata, M.M.; Carpio, V.H.; Ibitokou, S.A.; Dillon, B.E.; Obiero, J.M.; Stephens, R. Early effector cells survive the contraction phase in malaria infection and generate both central and effector memory T cells. J. Immunol. 2015, 194, 5346–5354. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central memory and effector memory T cell subsets: Function, generation, and maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef]
- Opata, M.M.; Ibitokou, S.A.; Carpio, V.H.; Marshall, K.M.; Dillon, B.E.; Carl, J.C.; Wilson, K.D.; Arcari, C.M.; Stephens, R. Protection by and maintenance of CD4 effector memory and effector T cell subsets in persistent malaria infection. PLoS Pathog. 2018, 14, e1006960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valle-Arroyo, J.; Aguado, R.; Páez-Vega, A.; Pérez, A.B.; González, R.; Fornés, G.; Torre-Cisneros, J.; Cantisán, S. Lack of cytomegalovirus (CMV)-specific cell-mediated immune response using QuantiFERON-CMV assay in CMV-seropositive healthy volunteers: Fact not artifact. Sci. Rep. 2020, 10, 7194. [Google Scholar] [CrossRef]
- Krensky, A.M.; Clayberger, C. Granulysin: A novel host defense molecule. Am. J. Transplant 2005, 5, 1789–1792. [Google Scholar] [CrossRef]
- Sarwal, M.M.; Jani, A.; Chang, S.; Huie, P.; Wang, Z.; Salvatierra, O.; Clayberger, C.; Sibley, R.; Krensky, A.M.; Pavlakis, M. Granulysin expression is a marker for acute rejection and steroid resistance in human renal transplantation. Hum. Immunol. 2001, 62, 21–31. [Google Scholar] [CrossRef]
- van Daalen, K.R.; Reijneveld, J.F.; Bovenschen, N. Modulation of Inflammation by Extracellular Granzyme A. Front. Immunol. 2020, 11, 931. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.G.; Fleck, M.; Kern, E.R.; Liu, D.; Wang, Y.; Hsu, H.C.; Yang, P.; Wang, Z.; Curiel, D.T.; Zhou, T.; et al. Antigen presenting cells expressing Fas ligand down-modulate chronic inflammatory disease in Fas ligand-deficient mice. J. Clin. Investig. 2000, 105, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Béziat, V.; Descours, B.; Parizot, C.; Debré, P.; Vieillard, V. NK Cell Terminal Differentiation: Correlated Stepwise Decrease of NKG2A and Acquisition of KIRs. PLoS ONE 2010, 5, e11966. [Google Scholar] [CrossRef] [PubMed]
- Foley, B.; Cooley, S.; Verneris, M.R.; Curtsinger, J.; Luo, X.; Waller, E.K.; Anasetti, C.; Weisdorf, D.; Miller, J.S. Human cytomegalovirus (CMV)-induced memory-like NKG2C(+) NK cells are transplantable and expand in vivo in response to recipient CMV antigen. J. Immunol. 2012, 189, 5082–5088. [Google Scholar] [CrossRef] [Green Version]
- Redondo-Pachón, D.; Crespo, M.; Yélamos, J.; Muntasell, A.; Pérez-Sáez, M.J.; Pérez-Fernández, S.; Vila, J.; Vilches, C.; Pascual, J.; López-Botet, M. Adaptive NKG2C+ NK Cell Response and the Risk of Cytomegalovirus Infection in Kidney Transplant Recipients. J. Immunol. 2017, 198, 94–101. [Google Scholar] [CrossRef] [Green Version]
- Della Chiesa, M.; Falco, M.; Podestà, M.; Locatelli, F.; Moretta, L.; Frassoni, F.; Moretta, A. Phenotypic and functional heterogeneity of human NK cells developing after umbilical cord blood transplantation: A role for human cytomegalovirus? Blood 2012, 119, 399–410. [Google Scholar] [CrossRef]
- Hendricks, D.W.; Balfour, H.H.; Dunmire, S.K.; Schmeling, D.O.; Hogquist, K.A.; Lanier, L.L. NKG2ChiCD57+ Natural Killer cells respond specifically to acute infection with cytomegalovirus and not Epstein-Barr virus. J. Immunol. 2014, 192, 4492–4496. [Google Scholar] [CrossRef] [Green Version]
- Bozzano, F.; Della Chiesa, M.; Pelosi, A.; Antonini, F.; Ascierto, M.L.; Del Zotto, G.; Moretta, F.; Muccio, L.; Luganini, A.; Gribaudo, G.; et al. HCMV-controlling NKG2C+ NK cells originate from novel circulating inflammatory precursors. J. Allergy Clin. Immunol. 2021, 147, 2343–2357. [Google Scholar] [CrossRef]
- Bayard, C.; Lepetitcorps, H.; Roux, A.; Larsen, M.; Fastenackels, S.; Salle, V.; Vieillard, V.; Marchant, A.; Stern, M.; Boddaert, J.; et al. Coordinated expansion of both memory T cells and NK cells in response to CMV infection in humans. Eur. J. Immunol. 2016, 46, 1168–1179. [Google Scholar] [CrossRef] [Green Version]
- Kared, H.; Martelli, S.; Tan, S.W.; Simoni, Y.; Chong, M.L.; Yap, S.H.; Newell, E.W.; Pender, S.L.F.; Kamarulzaman, A.; Rajasuriar, R.; et al. Adaptive NKG2C+CD57+ Natural Killer Cell and Tim-3 Expression during Viral Infections. Front. Immunol. 2018, 9, 686. Available online: https://www.frontiersin.org/article/10.3389/fimmu.2018.00686 (accessed on 9 February 2022). [CrossRef]
- Sottile, R.; Panjwani, M.K.; Lau, C.M.; Daniyan, A.F.; Tanaka, K.; Barker, J.N.; Brentjens, R.J.; Sun, J.C.; Le Luduec, J.B.; Hsu, K.C. Human cytomegalovirus expands a CD8+ T cell population with loss of BCL11B expression and gain of NK cell identity. Sci. Immunol. 2021, 6, eabe6968. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-LTX (n = 17) | HC (n = 15) | p Values | |
|---|---|---|---|
| Age (m ± SD) | 58 ± 7.3 | 52 ± 12 | ns |
| Gender (M:F) | 10:7 | 8:7 | ns |
| Smoking Habits (never/current/former) | 5/0/12 | 5/2/7 | ns |
| Tx indication: | |||
| - IPF | 7 | ||
| - Emphysema | 5 | ||
| - DIP | 1 | ||
| - FIBROSIS + PH | 1 | ||
| - HP | 2 | ||
| - PLCH | 1 | ||
| Therapy (steroids: antifibrotic) | 17:7 | ||
| Laboratory Findings | |||
| - CRP | 1.4 ± 3.7 | ||
| - Neutrophils (%) | 66.3 ± 15.4 | ||
| - Lymphocytes (%) | 22.9 ± 12.4 | ||
| - Monocytes (%) | 7.9 ± 2.7 | ||
| - Eosinophils (%) | 0.8 ± 0.7 | ||
| - Basophils (%) | 1.7 ± 1.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergantini, L.; d’Alessandro, M.; Otranto, A.; Cavallaro, D.; Gangi, S.; Fossi, A.; Perillo, F.; Luzzi, L.; Zanfrini, E.; Paladini, P.; et al. Characterization of NKG2-A/-C, Kir and CD57 on NK Cells Stimulated with pp65 and IE-1 Antigens in Patients Awaiting Lung Transplant. Life 2022, 12, 1081. https://doi.org/10.3390/life12071081
Bergantini L, d’Alessandro M, Otranto A, Cavallaro D, Gangi S, Fossi A, Perillo F, Luzzi L, Zanfrini E, Paladini P, et al. Characterization of NKG2-A/-C, Kir and CD57 on NK Cells Stimulated with pp65 and IE-1 Antigens in Patients Awaiting Lung Transplant. Life. 2022; 12(7):1081. https://doi.org/10.3390/life12071081
Chicago/Turabian StyleBergantini, Laura, Miriana d’Alessandro, Ambra Otranto, Dalila Cavallaro, Sara Gangi, Antonella Fossi, Felice Perillo, Luca Luzzi, Edoardo Zanfrini, Piero Paladini, and et al. 2022. "Characterization of NKG2-A/-C, Kir and CD57 on NK Cells Stimulated with pp65 and IE-1 Antigens in Patients Awaiting Lung Transplant" Life 12, no. 7: 1081. https://doi.org/10.3390/life12071081
APA StyleBergantini, L., d’Alessandro, M., Otranto, A., Cavallaro, D., Gangi, S., Fossi, A., Perillo, F., Luzzi, L., Zanfrini, E., Paladini, P., Sestini, P., Rottoli, P., Bargagli, E., & Bennett, D. (2022). Characterization of NKG2-A/-C, Kir and CD57 on NK Cells Stimulated with pp65 and IE-1 Antigens in Patients Awaiting Lung Transplant. Life, 12(7), 1081. https://doi.org/10.3390/life12071081