
Comparative Transcriptome Analysis Reveals Differential Gene Expression in Resistant and Susceptible Watermelon Varieties in Response to Meloidogyne incognita

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Nematodes Material, Growth Conditions
2.2. Extraction, Hatching and Collection of Root-Knot Nematode Eggs
2.3. Inoculation and Developmental Status Analysis of M. incognita
2.4. Total RNA Extraction and CDNA Library Preparation
2.5. RNA-Seq Data Sequencing, Assembly and Annotation
2.6. Quantitative Real-Time (qRT)-PCR Validation Analysis of DEGs
3. Results
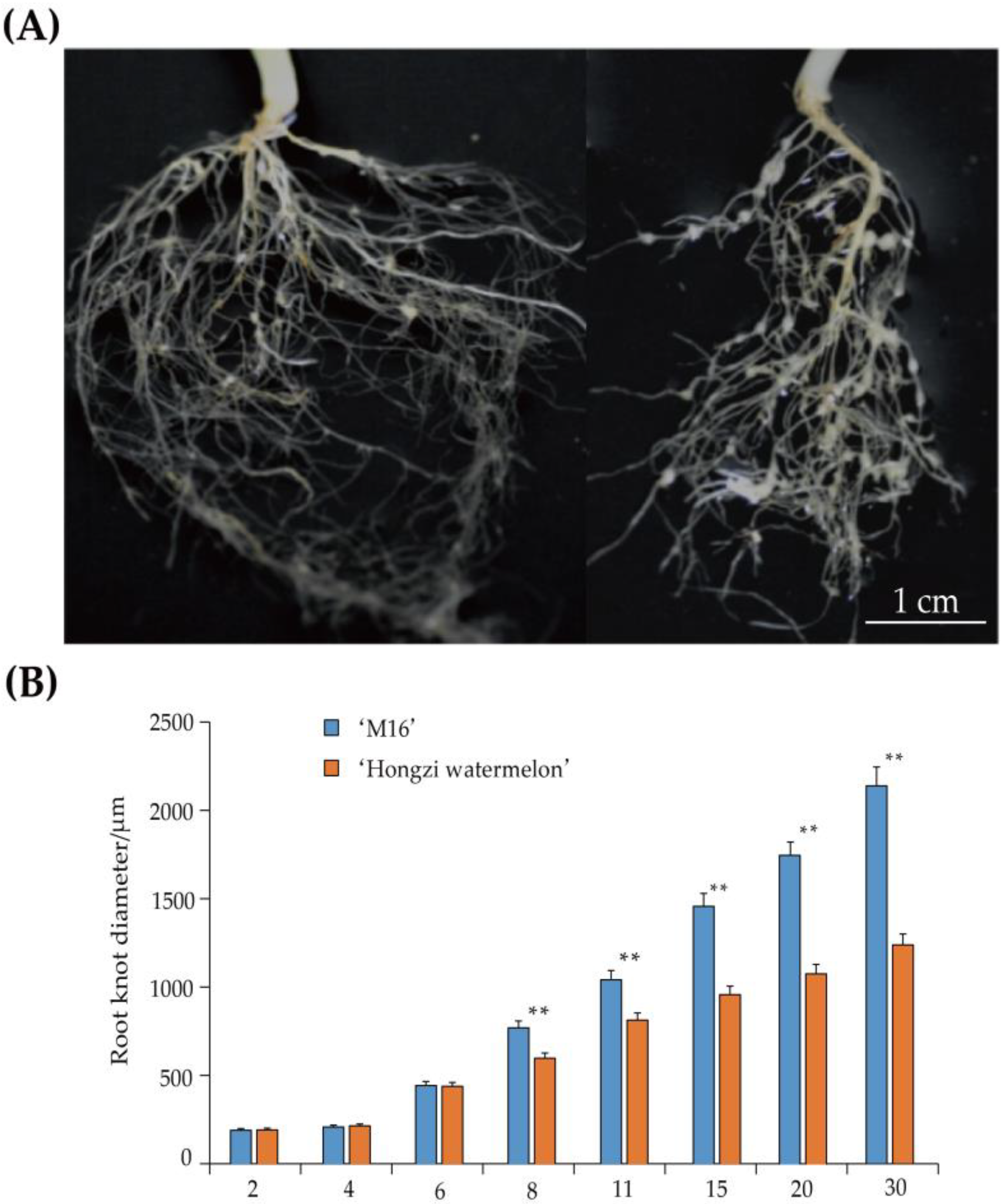
3.1. Analysis of Root-Knot Development
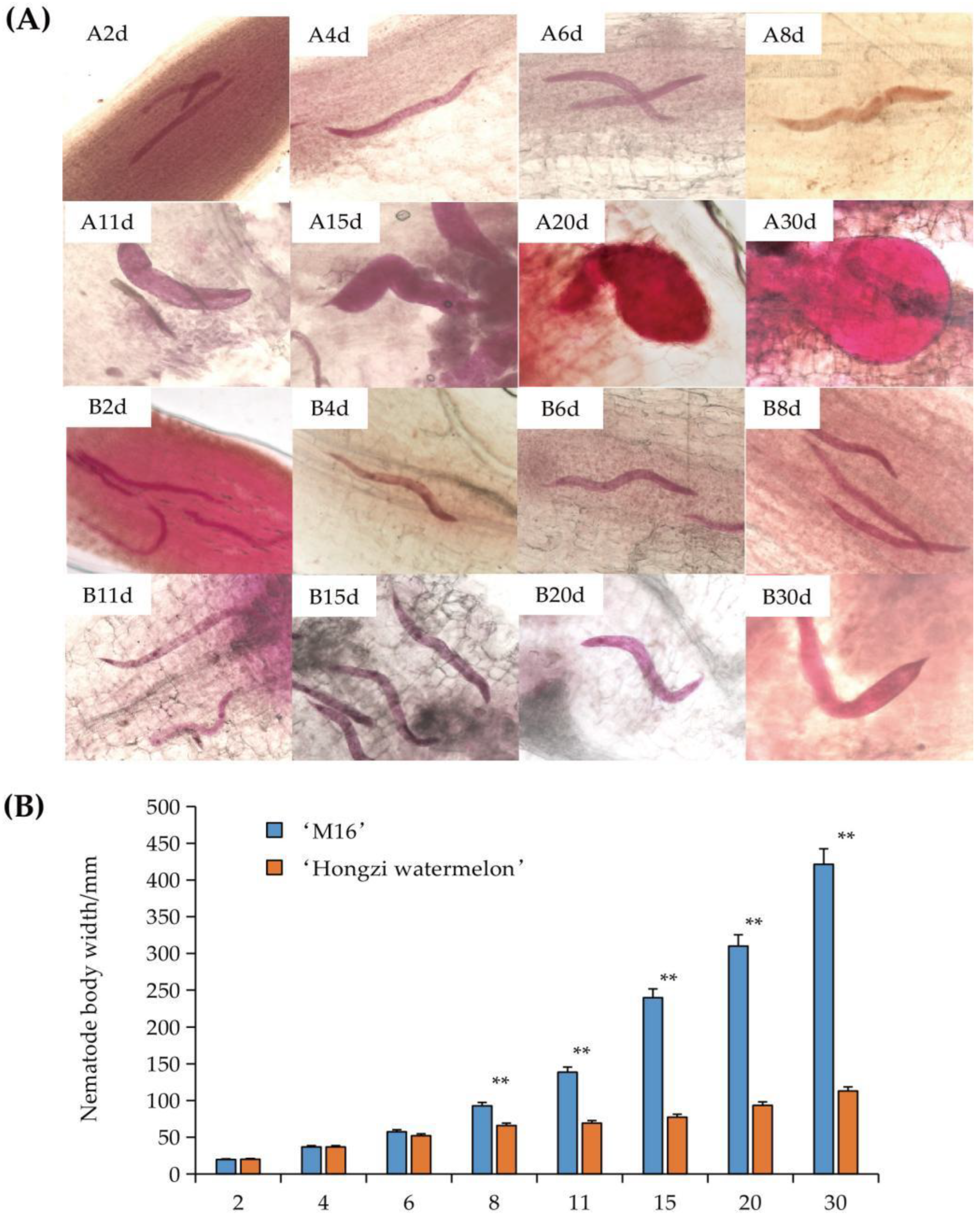
3.2. Analysis of Root-Knot Nematode Development
3.3. Transcriptome Sequencing and Read Mapping to Watermelon Genome
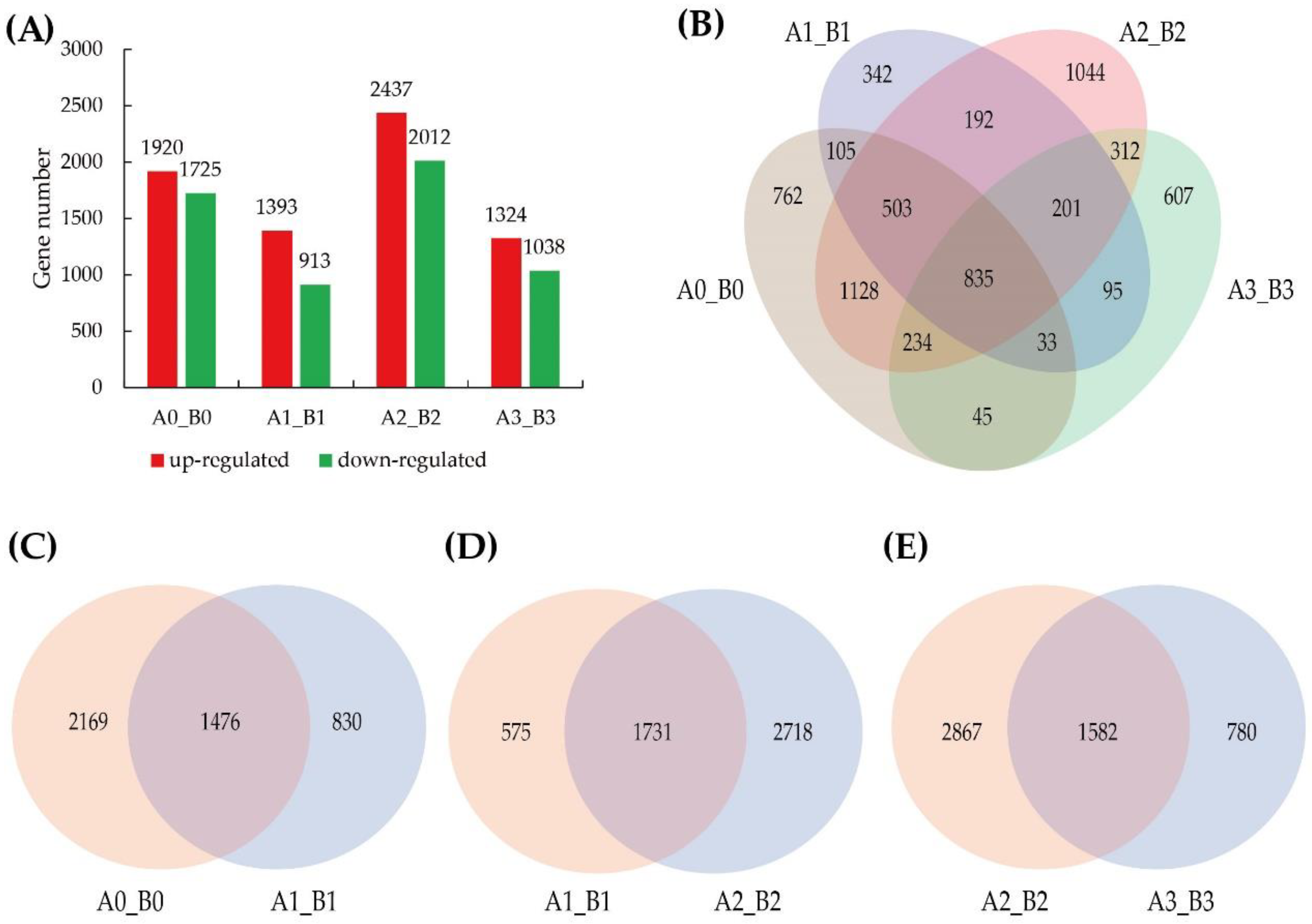
3.4. Analysis of DEGs
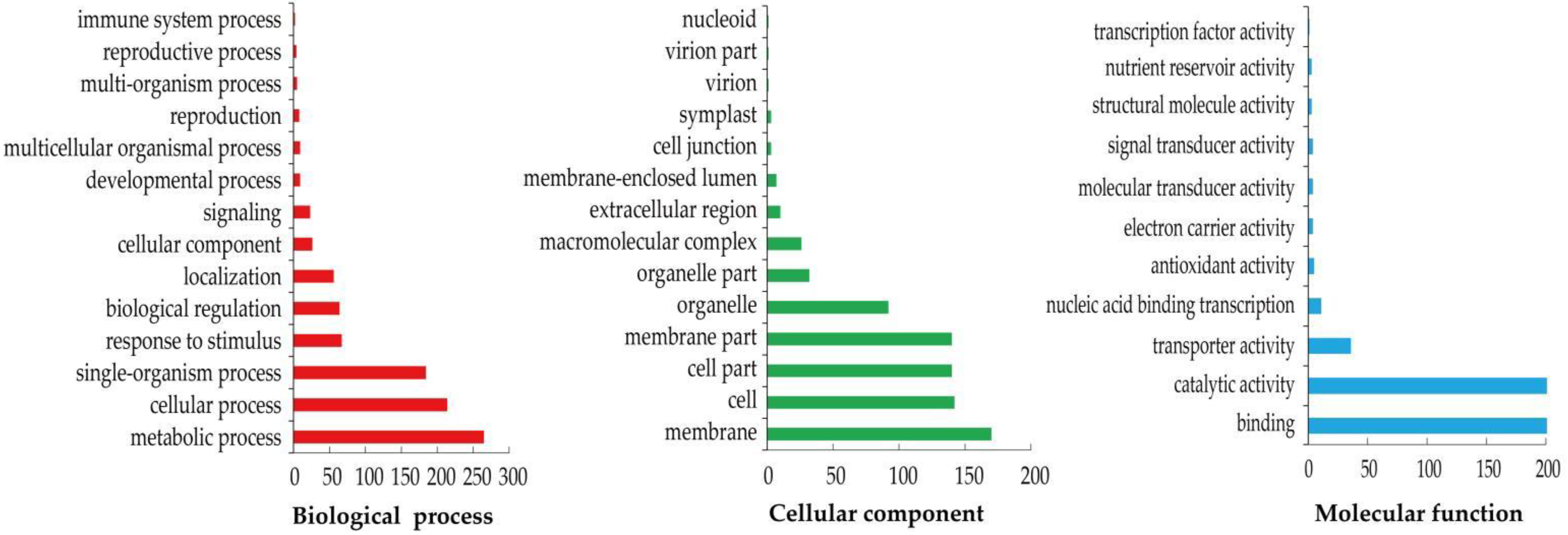
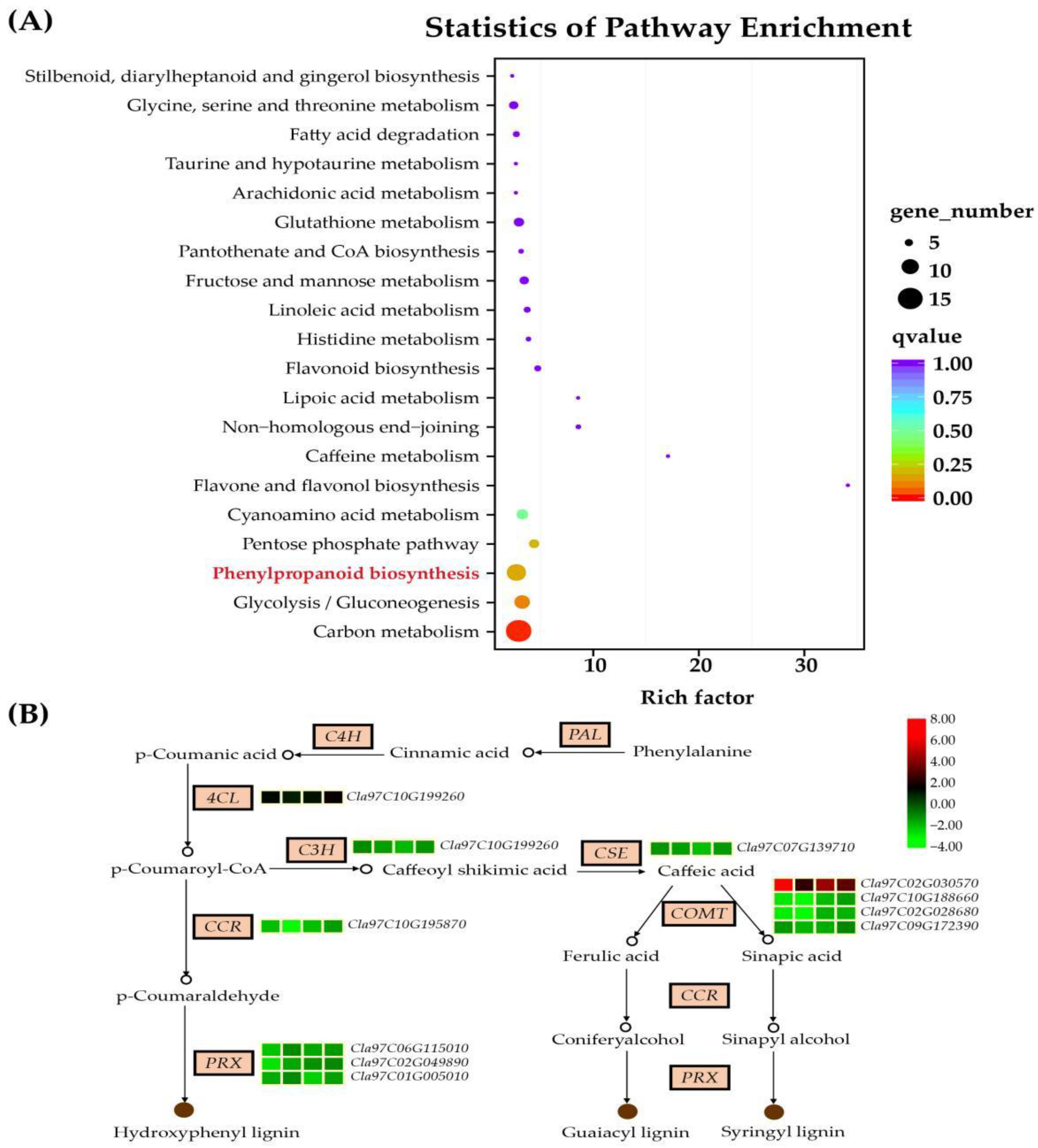
3.5. Functional Annotation and Classification of the DEGs
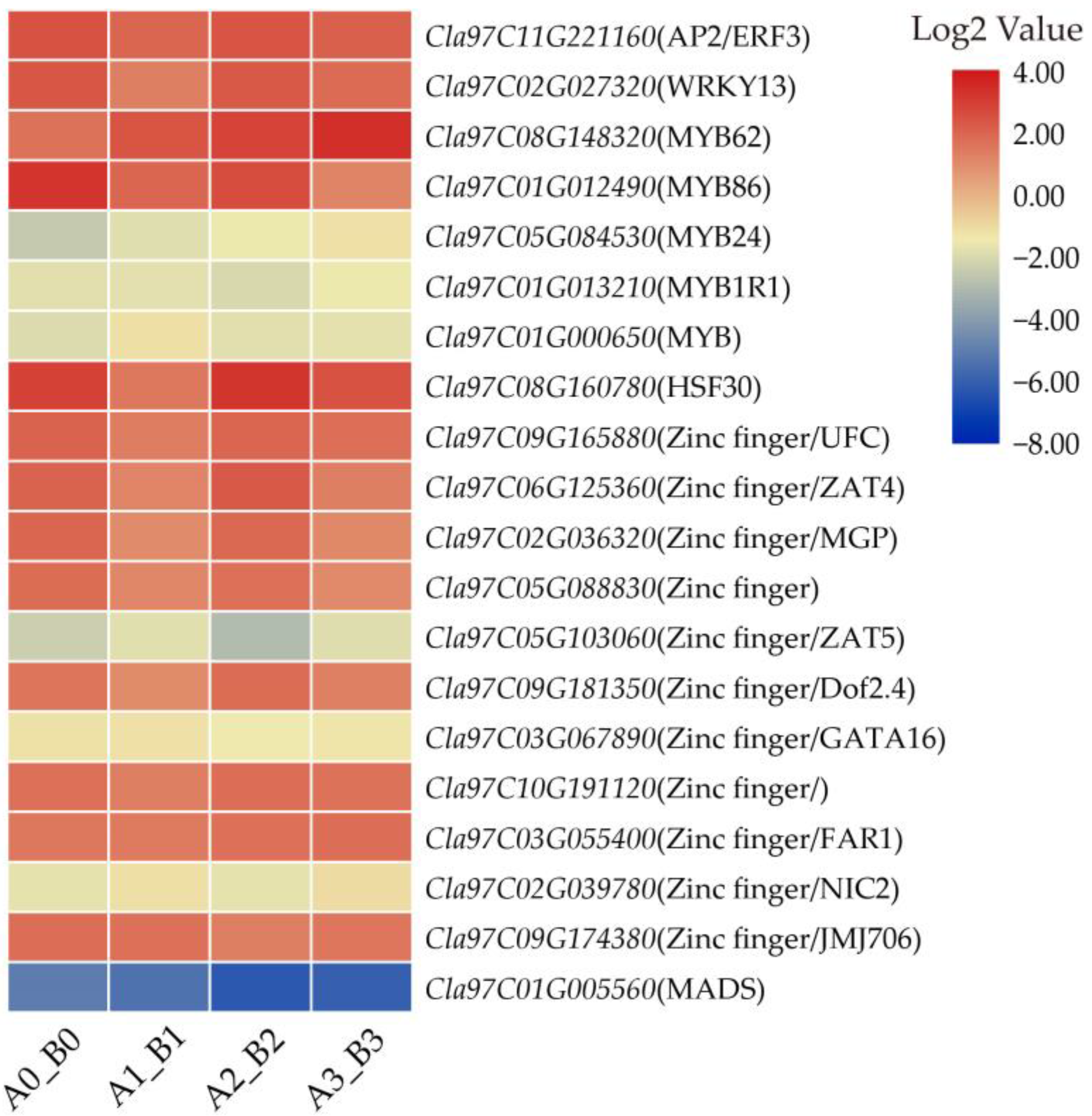
3.6. DEGs Related to Transcriptional Factors during M. incognita Infection
3.7. DEGs Related to Phytohormones
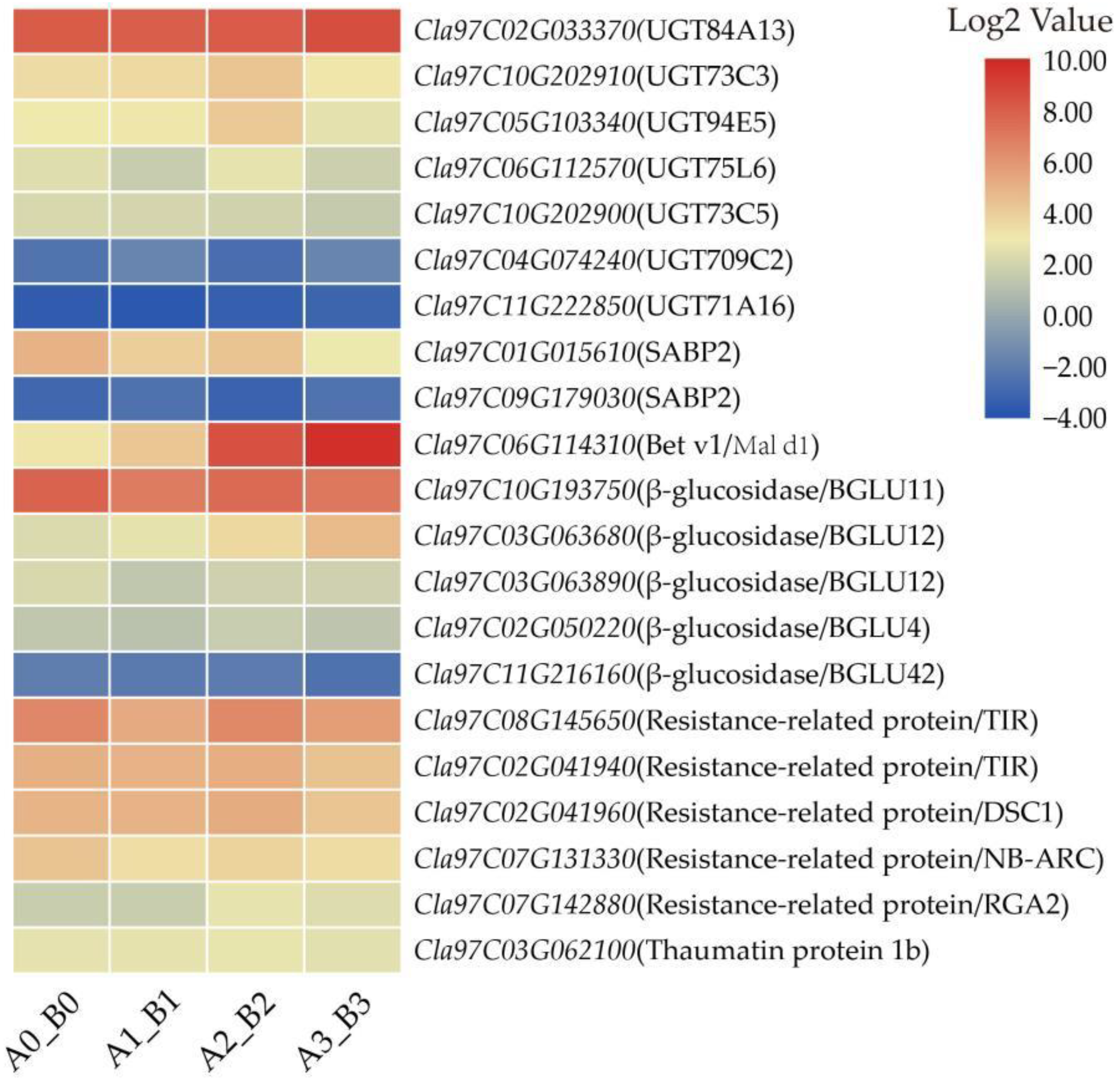
3.8. DEGs Related to Defense-Related Proteins
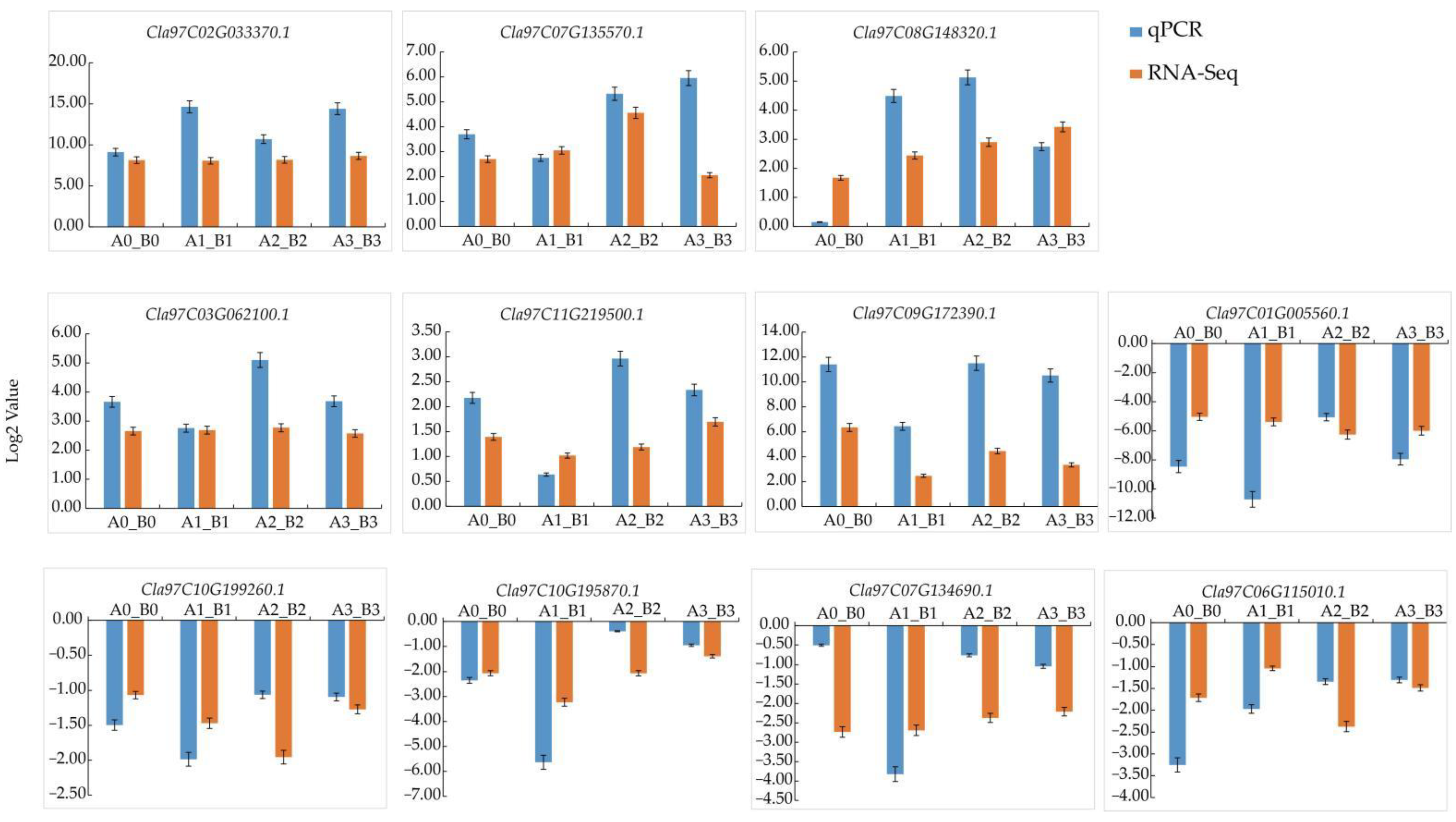
3.9. QRT-PCR Validation of Transcriptome Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DEG | differential expression gene |
| ABA | abscisic acid |
| SA | salicylic acid |
| GA | gibberellin |
| FDR | false discovery rate |
| ROS | reactive oxygen species |
| BP | biological process |
| MF | molecular function |
| CC | cell component |
| CPC | coding potential calculato |
| ORF | open reading frame |
| FPKM | fragments per kilobase of transcript per million fragments |
| qRT-PCR | quantitative real time PCR |
| ABC transporters | ATP binding cassette transporters |
| LTPs | lipid transfer proteins |
| SABP | salicylic acid-binding protein |
| 4CL | 4-coumaric acid-CoA ligase |
| C3H | coumaric acid 3-hydroxylase |
| CSE | caffeoyl shikimate esterase |
| COMT | caffeic acid-O-methyltransferase |
| CCR | cinnamyl CoA reductase |
| PRX | peroxidase |
| DPI | days post-infection |
| TF | transcription factor |
References
- Krishnaveni, M.; Subramanian, S. Root-knot nematodes of cucurbits and their management—A review. Agric. Rev. 2005, 26, 103–113. [Google Scholar]
- Thies, J.A.; Buckner, S.; Horry, M.; Hassell, R.; Levi, A. Influence of Citrullus lanatus var. citroides rootstocks and their F1 hybrids on yield and response to root-knot nematode, Meloidogyne incognita, in grafted watermelon. Hortscience 2015, 50, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Netscher, C.; Sikora, R. Nematode Parasites of Vegetables; CAB International: Wallingford, UK, 1990; pp. 237–283. [Google Scholar]
- Lu, Z.; Chen, M.; Hu, X.; Huang, J.; Xi, X.; Wang, Y. A study on the sensitivity of main vegetables produced in Beijing greenhouse to root knot nematode. In Proceedings of the 13th National Symposium on Plant Nematodes, Coral Gables, FL, USA, 13–15 December 2016; Volume 1, p. 29. [Google Scholar]
- Zhao, H.; Peng, D.L.; Zhu, J.L. Reviews on the root-knot nematodes. Plant Prot. 2003, 6, 6–9. [Google Scholar]
- Zhang, X.W.; Qian, X.L.; Liu, J.W. Evaluation on the resistance to root-knot nematode of watermelon germplasm and control for it. J. Fruit Sci. 1989, 1, 33–38. [Google Scholar]
- Vela, M.D.; Giné, A.; López-Gómez, M.; Sorribas, F.J.; Ornat, C.; Verdejo-Lucas, S.; Talavera, M. Thermal time requirements of root-knot nematodes on zucchini-squash and population dynamics with associated yield losses on spring and autumn cropping cycles. Eur. J. Plant. Pathol. 2014, 140, 481–490. [Google Scholar] [CrossRef]
- Verdejo-Lucasa, S.; Talavera, M. Root-knot nematodes on zucchini (Cucurbita pepo subsp. pepo): Pathogenicity and management. Crop. Prot. 2019, 12, 104943. [Google Scholar] [CrossRef]
- Shahbaz, M.U.; Mukhtar, T.; Irfanulhaque, M.; Begum, N. Biochemical and serological characterization of Ralstonia solanacearum associated with chilli seeds from Pakistan. Int. J. Agric. Biol. 2015, 17, 31–40. [Google Scholar]
- Fu, L. Preliminary study on the occurrence and control technology of plant pathogenic nematode. Agric. Technol. 2015, 35, 40–41. [Google Scholar]
- Morris, K.A.; Langston, D.B.; Davis, R.F.; Noe, J.; Dickson, D.; Timper, P. Efficacy of various application methods of fluensulfone for managing root-knot nematodes in vegetables. J. Nematol. 2016, 48, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thies, J.A.; Levi, A. Characterization of watermelon (Citrullus lanatus var. citroides) germplasm for resistance to root-knot nematodes. Hortscience 2007, 42, 188. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Li, X.; Feng, L.; Wang, H.; Song, J.; Yang, C.; Gong, H. Evaluation on resistance of Cucurbitaceae germplasm resources to root-knot nematode. J. Plant Genet. Resour. 2007, 3, 340–342. [Google Scholar]
- Thies, J.A.; Levi, A.; Ariss, J.J.; Hassell, R. RKVL-318, a root-knot nematode-resistant watermelon line as rootstock for grafted watermelon. Hortscience 2015, 50, 141–142. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, Z.; Sun, D.; Lu, J.; Ma, S.; Zhu, Y.; Liu, J. Screening and evaluation of watermelon germplasm for resistance to Meloidogyne incognita. J. Fruit Sci. 2012, 29, 1103–1109. [Google Scholar]
- Punithaveni, V.; Jansirani, P.; Sivakumar, M. Screening of cucurbitaceous rootstocks and cucumber scions for root knot nematode resistance (Meloidogyne incognita Kofoid and White). Electron. J. Plant Breed. 2015, 6, 486–492. [Google Scholar]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.G.; Huang, Y.H.; Lu, H.B.; Tian, D.Y.; Zhang, Y.W. The application perspective of transcriptome sequencing on discovering the genes and variety breeding of bioenergy grass. Acta Agrestia Sin. 2016, 24, 731–737. [Google Scholar]
- Kyndt, T.; Denil, S.; Haegeman, A.; Geert Trooskens, G.; Bauters, L.; Criekinge, W.V.; Meyer, T.D.; Gheysen, G. Transcriptional reprogramming by root knot and migratory nematode infection in rice. New Phytol. 2012, 196, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Olga, A.; Maria, H.; Jonathan, S.; Andrea, S.; Lev, G. Transcriptome analysis of resistant and susceptible alfalfa cultivars infected with root-knot nematode Meloidogyne incognita. PLoS ONE 2015, 10, e0118269. [Google Scholar]
- Xue, Y.Y.; Deng, Y.; Liu, J.P.; Xu, X.J.; Zhu, Y.C.; An, G.L.; Li, W.H.; Sun, D.X. Molecular cloning and expression analysis of root-knot nematode resistance related aene in watermelon. J. Fruit Sci. 2015, 32, 1107–1113. [Google Scholar]
- Li, K. Screening of Watermelon and Melon Rootstock Resources Resistant to Meloidogyne Incognita and Transcriptomic Analysis. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijng, China, 2016. [Google Scholar]
- Ling, J.; Mao, Z.; Zhai, M.; Zeng, F.; Yang, Y.H.; Xie, B.Y. Transariptome profiling of Cucumis metuliferus infected by Meloidogyne incognita provides new insights into putative defense regulatory network in Cucurbitaceae. Sci. Rep. 2017, 7, 3544. [Google Scholar] [CrossRef] [PubMed]
- Li, S. Study on Transcriptome and Protein Interaction Network of “Huipizhi Heidou” Resistance to Soybean Cyst Nematode. Ph.D. Thesis, Shenyang Agricultural University, Shenyang, China, 2018. [Google Scholar]
- Wang, X.W. Identification of Root-Knot Nematodesfrom Protected Cultivation and Resistance of Vegetable Cultivars. Master’s Thesis, Zhejiang University, Hangzhou, China, 2013. [Google Scholar]
- Guo, S.G.; Zhao, S.J.; Sun, H.; Wang, X.; Wu, S.; Lin, T.; Ren, Y.; Gao, L.; Deng, Y.; Zhang, J.; et al. Resequencing of 414 cultivated and wild watermelon accessions identifies selection for fruit quality traits. Nat. Genet. 2019, 51, 1616–1623. [Google Scholar] [CrossRef]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2014, 7, 562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, G.P.; Liu, J.P.; An, G.L.; Li, W.H.; Si, W.J.; Sun, D.X.; Zhu, Y. Genome-wide identification and characterization of the treha-lose-6-phosphate synthetase (TPS) gene family in watermelon (Citrullus lanatus) and their transcriptional responses to salt stress. Int. J. Mol. Sci. 2021, 23, 276. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.; Frank, M.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, J.; Cheng, Y.; Wei, W.; Li, L.; Chen, Y. Identification and analysis of WRKY transcription factors in Siraitia grosvenorii under infection of Meloidogyne incognita. Plant Protoc. 2020, 46, 169–174. [Google Scholar]
- He, S.; Yuan, G.; Bian, S.; Han, X.; Liu, K.; Cong, P.; Zhang, C. Major latex protein MdMLP423 negatively regulates defense against fungal infections in apple. Int. J. Mol. Sci. 2020, 21, 1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, J.; Baltz, R.; Schmitt, C.; Beffa, R.; Fritig, B.; Sain-Drenan, P. Downregulation of a pathogen-responsive tobacco UDP-Glc: Phenylpropanold glucosyltran sferase reduces scopoletin glucoside accumulation, enhances oxidative stress, and weakens virus resistance. Plant Cell 2002, 14, 1093–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gachon, C.; Baltz, R.; Saindrenan, P. Over-expression of a scopoletin glucosyltransferase in Nicotiana tabacum leads to precocious lesion formation during the hypersensitive response to tobacco mosaic virus but does not affect virus resistance. Plant Mol. Biol. 2004, 54, 137–146. [Google Scholar] [CrossRef]
- Sepulveda-Jimenez, G.; Rueda-Benitez, P.; Porta, H.; Ro-cha-Sosa, M. A red beet (Beta vulgaris) UDP-glucosyltrans-ferase gene induced by wounding, bacterial infiltration and oxidative stress. J. Exp. Bot. 2005, 56, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Mao, Z.; Li, H.; Xie, B. Resistance identification of Cucumis metuliferus to Meloidogyne incognita and characteristic analysis. Acta Hortic. Sin. 2014, 41, 73–79. [Google Scholar]
- Wei, C.; Shi, Q.Q.; Ma, Y.Q.; Ma, J.H.; Mao, Z.C.; Ling, J.; Yang, Y.H.; Xie, B.Y. Effects of peroxidase gene to the resistance of Cucumis metuliferus against Meloidogyne incognita in different temperature. Acta Hortic. Sin. 2016, 43, 1537–1544. [Google Scholar]
- Wang, Z. The Screening and Evaluation of Watermelon Rootstock Resources for Resistance to Meloidogyne Incognita. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2011. [Google Scholar]
- Zhang, Z. Phenolic Compositions and Antioxidant Capacity of the Fruits of Chinese Local Pummelos. Master’s Thesis, Southwest University, Chongqing, China, 2015. [Google Scholar]
- Dou, L. The Negative Regulational Molecular Mechanism of the Poplar Transcription Factor MYB93 Involved in Flavonoids and Lignin Biosynthesis. Master’s Thesis, Southwest University, Chongqing, China, 2019. [Google Scholar]
- Fu, M.; Chen, J.; Xiao, T.; Rui, K. Advances in studies on interaction mechanism between root-knot nematodes and host plants. Chin. J. Trop. Agric. 2008, 3, 73–77. [Google Scholar]
- Xu, X. Study on Evaluation for Resistance to Meloidogyne Incognita and the Resistant Mechanism of Eggplant Rootstocks. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2008. [Google Scholar]
- Zhao, R.Z. Studies on the Transcriptional Analysis of Resistance Response of Watermelon to Meloidogyne Incognita and Induced Control by Red-Light. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2018. [Google Scholar]
- Lin, M.; He, L.; Wen, L.; Fang, Z. Mechanism of morphological structure of sweet potato resistance to potato rot nematode (Ditylenchus destructor). Sci. Agric. Sin. 1996, 29, 8–12. [Google Scholar]
- Ye, D.Y.; Wang, X.; Zhang, Y.X.; Qian, C.; Chen, J.F. Anatomy and cytology of sour cucumber for its resistance to the root-knot nematode Meloidogyne incognita. Acta Phytopathol. Sin. 2010, 3, 73–77. [Google Scholar]
- Bendezu, I.F.; Starr, J.L. Mechanism of resistance to Meloidogyne arenaria in the peanut cultivar COAN. J. Nematol. 2003, 35, 151. [Google Scholar]
- Li, X.; Wang, W.; Wu, Y. Physiological function and economic value of plant tannin. J. West China For. Sci. 2005, 1, 66–69. [Google Scholar]
- Chen, R.; Gao, H.; Zhang, G.; Zhu, K.; Cheng, W. Effects of secondary metabolites in wheat kernels on activities of three detoxifying enzymes and related gene expression in Sitodiplosis mosellena. Sci. Agric. Sin. 2020, 53, 4204–4214. [Google Scholar]
- Wu, Y.Q.; Guo, Y.Y. Potential resistance of tannins-flavonoids in upland cottonagainst Helicpverpa armigera. Acta Ecol. Sin. 2001, 21, 286–289. [Google Scholar]
- Dong, W.; Chen, C.; Ma, H. Analysis of insect resistance and herbicide resistance in transgenic alfalfa plants over-expressing the OvBAN/bar gene. Acta Prataculturae Sin. 2019, 28, 159–167. [Google Scholar]
- Lees, G.L.; Hinks, C.F.; Suttill, N.H. Condensed tannins in some forage legumes their rolein the improvement of ruminant pasture bloat. Basic Life Sci. 1992, 59, 915–934. [Google Scholar] [PubMed]
- Shen, F.; Yu, Y.J.; Zhang, X.; Liu, F.; Yin, C. Insect resistant stock selected byintroducing dogbane DNA into cotton. Acta Agric. Boreali-Sin. 1999, 4, 34–38. [Google Scholar]
- Mittasch, J.; Böttcher, C.; Frolova, N.; Bönn, M.; Milkowski, C. Identification of UGT84A13 as a candidate enzyme for the first committed step of gallotannin biosynthesis in pedunculate oak (Quercus robur). Phytochemistry 2014, 99, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Jammes, F.; Lecomte, P.; de Almeida, E.J.; Bitton, F.; Martin-Magniette, M.; Renou, J.; Abad, P.; Favery, B. Genome-wide expression profiling of the host response to root-knot nematode infection in Arabidopsis. Plant J. 2005, 44, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Mao, Z.; Li, L.; Feng, D.; Yang, Y.; Xie, B. Isolation of WRKY genes in the incompatible interaction between Meloidogyne incognita and capsicum annuum. Acta Hortic. Sin. 2008, 10, 1467–1472. [Google Scholar]
- Zheng, J. Isolation, Expression and Function Analysis of the WRKY Transcription Factor Gene CaWRKY6 and CaWRKY30 in Pepper. Ph.D. Thesis, Central South University, Changsha, China, 2011. [Google Scholar]
- Chinnapandi, B.; Bucki, P.; Miyara, S.B. SIWRKY45, nematode-responsive tomato WRKY gene enhances susceptibility to the M. javanica infection. Plant Signal. Behav. 2017, 12, e1356530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X. Cloning and Localization of Root-Knot Nematode Resistance Relative MYB Transcription Factor Genes in ‘Honggengansutao Peach’ (Prunus kansuensis L.). Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2010. [Google Scholar]
- Warmcrdam, S.; Sterken, M.G.; Schaik, C.V.; Oortwijn, M.E.P.; Lozano-Torre, J.L.; Bakker, J.; Goverse, A.; Smant, G. Mediator of tolerance to abiotic stress ERF6 regulates susceptibility of Arabidopsis to Meloidogyne incognita. Plant Pathol. 2019, 20, 137–152. [Google Scholar]
- Li, L. Transcriptomic and metabolomics study of the early interactions between Siraitia grosvenorii roots and Meloidogyne incognita. Master’s Thesis, Guangxi Normal University, Guilin, China, 2019. [Google Scholar]
- Nahar, K.; Kyndt, T.; de Vleesschauwer, D.; Hofte, M.; Gheysen, G. The jasmonate pathway is a key player in systemically induced defense against root-knot nematodes in rice. Plant Physiol. 2011, 157, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbeek, R.; van Buyten, E.; Alam, M.; de Vleesschauwer, D.; van Bockhaven, J.; Asano, T.; Kikuchi, S.; Haeck, A. Jasmonate-induced defense mechanisms in the belowground antagonistic interaction between Pythium arrhenomanes and Meloidogyne graminicola in rice. Front. Plant Sci. 2019, 10, 1515. [Google Scholar] [CrossRef] [PubMed]
- Nahar, K.; Kyndt, T.; Nzogela, Y.B.; Gheysen, G. Abscisic acid interacts antagonistically with classical defense pathways in rice-migratory nematode interaction. New Phytol. 2012, 196, 901–913. [Google Scholar] [CrossRef]
- Nahar, K.; Kyndt, T.; Hause, B.; Hofte, M.; Godelieve, G. Brassinosteroids suppress rice defense against root-knot nematodes through antagonism with the jasmonate pathway. Mol. Plant Microbe Interact. 2013, 26, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yimer, H.; Nahar, K.; Kyndt, T.; Haeck, A.; van Meulebroek, L.; Vanhaecke, L.; Demeestere, K.; Hofte, M.; Gheysen, G. Gibberellin antagonizes jasmonate-induced defense against Meloidogyne graminicola in rice. New Phytol. 2018, 218, 646–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godelieve, G.; Melissa, G. Phytoparasitic nematode control of plant hormone pathways. Plant Physiol. 2018, 18, 01067. [Google Scholar]
- Baker, E.; Dees, R.; Bakker, J. Mechanisms Involved in Plant Resistance to Nematodes; Springer: Berlin, Germany, 2005. [Google Scholar]
- Vos, C.; Schouteden, N.; Tuinen, D.V. Mycorrhiza-induced resistance against the root-knot nematode Meloidogyne incognita involves priming of defense gene responses in tomato. Soil Biol. Biochem. 2013, 60, 45–54. [Google Scholar] [CrossRef]
- Holajjer, P.; Chakraborty, K.; Nataraja, M.V. Differential reactive oxygen species modulation in leaves of groundnut due to root-knot nematodes, Meloido gnearenaria and M. incognita. J. Nematol. 2016, 46, 102–106. [Google Scholar]
- Jia, S. Study on Evaluation and Mechanism of Tomato Rootstocks for Resistance to Meloidogyne Incognita. Ph.D. Thesis, Shandong Agricultural University, Tai’an, China, 2012. [Google Scholar]
- Guo, Y.; Wang, X.; Xu, K.; Zhang, G. Effect of Meloidogyne incognita on the physiological and chemical changes in ginger. Acta Phytopathol. Sin. 2005, 1, 49–54. [Google Scholar]
- Park, S.; Liu, P.; Forouhar, F.; Vlot, A.; Tong, L.; Tietjen, K.; Klessig, D. Use of a synthetic salicylic acid analog to investigate the roles of methyl salicylate and its esterases in plant disease resistance. J. Biol. Chem. 2009, 284, 7307–7317. [Google Scholar] [CrossRef] [Green Version]
- Forouhar, F.; Yang, Y.; Kumar, D.; Chen, Y.; Fridman, E.; Park, S.; Chiang, Y.; Acton, T.; Montelione, G.; Pichersky, E.; et al. Structural and biochemical studies identify tobacco SABP2 as a methyl salicylate esterase and implicate it in plant innate immunity. Proc. Natl. Acad. Sci. USA 2005, 102, 1773–1778. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Yuan, G.; Zhao, R.; An, G.; Li, W.; Si, W.; Liu, J.; Sun, D. Comparative Transcriptome Analysis Reveals Differential Gene Expression in Resistant and Susceptible Watermelon Varieties in Response to Meloidogyne incognita. Life 2022, 12, 1003. https://doi.org/10.3390/life12071003
Zhu Y, Yuan G, Zhao R, An G, Li W, Si W, Liu J, Sun D. Comparative Transcriptome Analysis Reveals Differential Gene Expression in Resistant and Susceptible Watermelon Varieties in Response to Meloidogyne incognita. Life. 2022; 12(7):1003. https://doi.org/10.3390/life12071003
Chicago/Turabian StyleZhu, Yingchun, Gaopeng Yuan, Renzong Zhao, Guolin An, Weihua Li, Wenjing Si, Junpu Liu, and Dexi Sun. 2022. "Comparative Transcriptome Analysis Reveals Differential Gene Expression in Resistant and Susceptible Watermelon Varieties in Response to Meloidogyne incognita" Life 12, no. 7: 1003. https://doi.org/10.3390/life12071003
APA StyleZhu, Y., Yuan, G., Zhao, R., An, G., Li, W., Si, W., Liu, J., & Sun, D. (2022). Comparative Transcriptome Analysis Reveals Differential Gene Expression in Resistant and Susceptible Watermelon Varieties in Response to Meloidogyne incognita. Life, 12(7), 1003. https://doi.org/10.3390/life12071003