
P62/SQSTM1 beyond Autophagy: Physiological Role and Therapeutic Applications in Laboratory and Domestic Animals

 ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
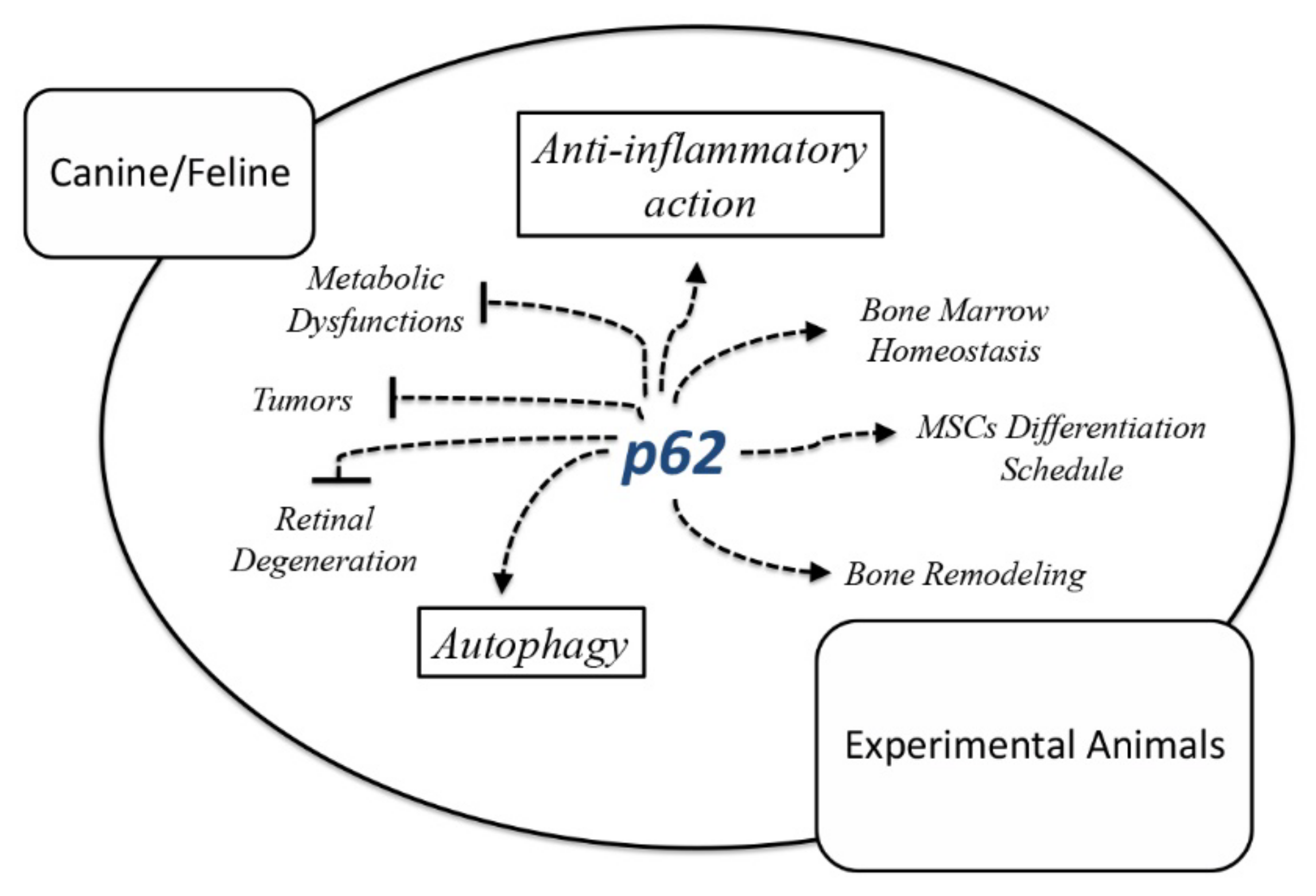
2. P62 as a Multitask Protein for Tissue Homeostasis
2.1. P62 in Rodent’s Bone and Bone Marrow Pathophysiology
2.2. P62 in Metabolic Dysfunctions in Rodents
2.3. P62 Protective Effects in Neurodegenerative Ocular Diseases
2.4. P62 Preventive and Therapeutic Effects against Tumors in Mice
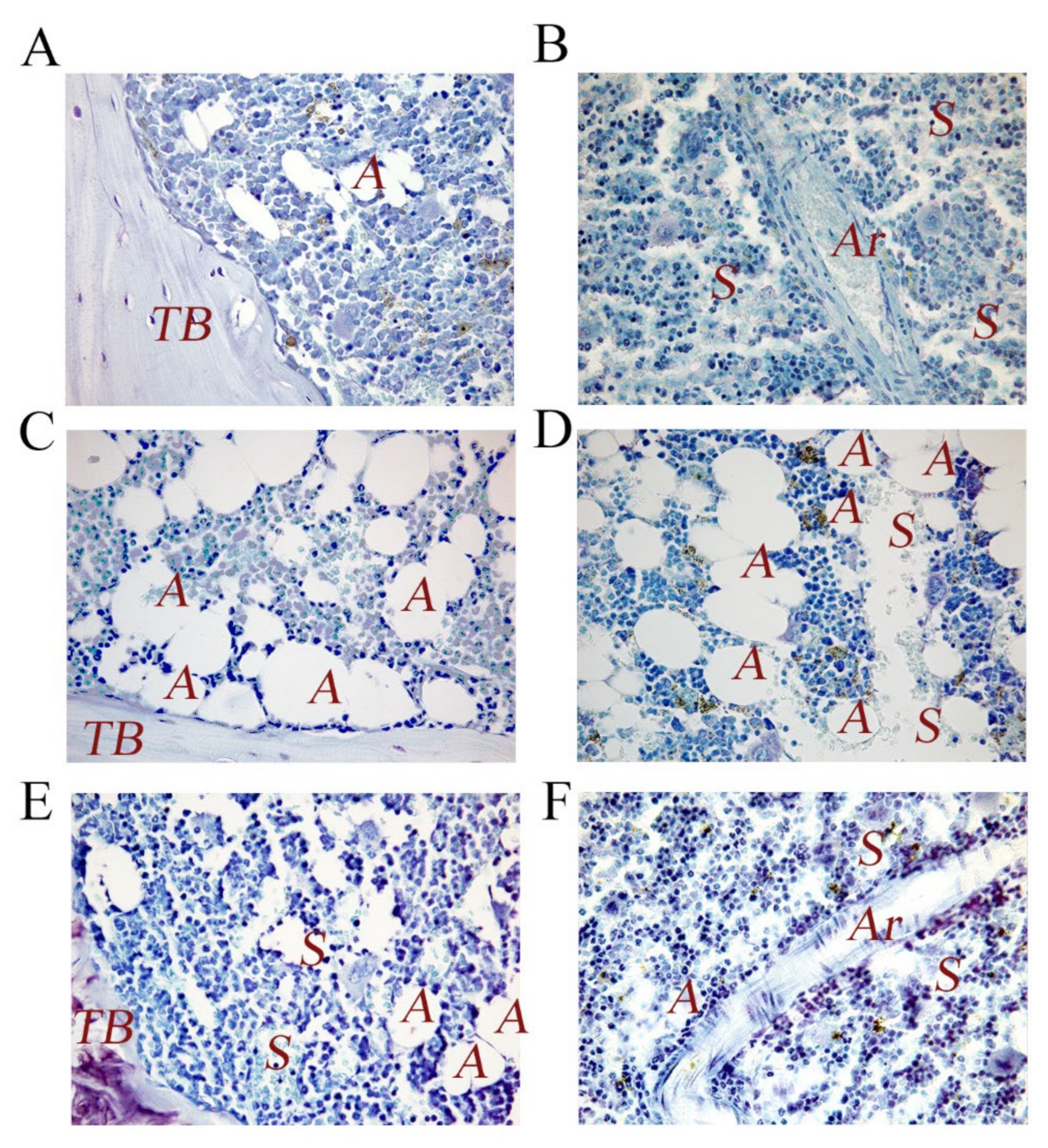
2.5. P62-Encoding Plasmid as Therapeutic Agent in Dogs with Mammary Tumors
3. Can a Human Anticancer Experimental Drug, p62-Encoding Plasmid, Become a Broad-Spectrum Anticancer Therapy for Companion Animals?
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Animal Welfare Statement
References
- Kumar, V.; Abbas, A.K.; Fausto, N.; Robbins, S.L.; Cotran, R.S. Pathologic Basis of Disease, 7th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2005. [Google Scholar]
- Okin, D.; Medzhitov, R. Evolution of inflammatory diseases. Curr. Biol. 2012, 22, R733–R740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, K.R.; Mahajan, U.B.; Unger, B.S.; Goyal, S.N.; Belemkar, S.; Surana, S.J.; Ojha, S.; Patil, C.R. Animal Models of Inflammation for Screening of Anti-inflammatory Drugs: Implications for the Discovery and Development of Phytopharmaceuticals. Int. J. Mol. Sci. 2019, 20, 4367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolz, A.; Ernst, A.; Dikic, I. Cargo recognition and trafficking in selective autophagy. Nat. Cell. Biol. 2014, 16, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zhou, C.; Li, J.; Liu, R.; Shi, B.; Yuan, Q.; Zou, S. Autophagy in bone homeostasis and the onset of osteoporosis. Bone Res. 2019, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Puissant, A.; Fenouille, N.; Auberger, P. When autophagy meets cancer through p62/SQSTM1. Am. J. Cancer Res. 2012, 2, 397–413. [Google Scholar]
- Qiang, L.; Zhao, B.; Ming, M.; Wang, N.; He, T.-H.; Hwang, S.; Thorburn, A.; He, Y.-Y. Regulation of cell proliferation and migration by p62 through stabilization of Twist1. Proc. Natl. Acad. Sci. USA 2014, 111, 9241–9246. [Google Scholar] [CrossRef] [Green Version]
- Agas, D.; Sabbieti, M.G. Autophagic Mediators in Bone Marrow Niche Homeostasis. Adv. Exp. Med. Biol. 2021, 1–15. [Google Scholar] [CrossRef]
- Luo, R.Z.; Yuan, Z.Y.; Li, M.; Xi, S.Y.; Fu, J.; He, J. Accumulation of p62 is associated with poor prognosis in patients with triple-negative breast cancer. Onco Targets Ther. 2013, 6, 883–888. [Google Scholar]
- Inoue, D.; Suzuki, T.; Mitsuishi, Y.; Miki, Y.; Suzuki, S.; Sugawara, S.; Watanabe, M.; Sakurada, A.; Endo, C.; Uruno, A.; et al. Accumulation of p62/SQSTM1 is associated with poor prognosis in patients with lung adenocarcinoma. Cancer Sci. 2012, 103, 760–766. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, Q.; Zhou, Y.; Sun, L.; Hu, B.; Xue, H.; Li, M.; Zhang, K.; Ren, C.; Duan, N.; et al. Overexpression of p62 is associated with poor prognosis and aggressive phenotypes in osteosarcoma. Oncol. Lett. 2018, 15, 9889–9895. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.H.; Sengupta, A.; Nayak, R.C.; Duran, A.; Lee, S.J.; Pratt, R.G.; Cancelas, J.A. p62 is required for stem cell/progenitor retention through inhibition of IKK/NF-κB/Ccl4 signaling at the bone marrow macrophage-osteoblast niche. Cell Rep. 2014, 9, 2084–2097. [Google Scholar] [CrossRef] [Green Version]
- Lacava, G.; Laus, F.; Amaroli, A.; Marchegiani, A.; Censi, R.; Di Martino, P.; Yanagawa, T.; Sabbieti, M.G.; Agas, D. P62 deficiency shifts mesenchymal/stromal stem cell commitment toward adipogenesis and disrupts bone marrow homeostasis in aged mice. J. Cell. Physiol. 2019, 234, 16338–16347. [Google Scholar] [CrossRef]
- Sabbieti, M.G.; Agas, D.; Capitani, M.; Marchetti, L.; Concetti, A.; Vullo, C.; Catone, G.; Gabai, V.; Shifrin, V.; Sherman, M.Y.; et al. Plasmid DNA-coding p62 as a bone effective anti-inflammatory/anabolic agent. Oncotarget 2015, 6, 3590–3599. [Google Scholar] [CrossRef] [Green Version]
- Agas, D.; Amaroli, A.; Lacava, G.; Yanagawa, T.; Sabbieti, M.G. Loss of p62 impairs bone turnover and inhibits PTH-induced osteogenesis. J. Cell. Physiol. 2020, 235, 7516–7529. [Google Scholar] [CrossRef]
- Tipold, A.; Stein, V.M. Inflammatory Diseases of the Spine in Small Animals. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 871–879. [Google Scholar] [CrossRef]
- Diehl, L.; Meyerholz, D.K.; Day, M.J.; Affolter, V.K. Pathology and Pathogenesis of Immune-Mediated Diseases of Animals. Vet. Pathol. 2018, 55, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Al-Hizab, F.; Clegg, P.D.; Thompson, C.C.; Carter, S.D. Microscopic localization of active gelatinases in equine osteochondritis dissecans (OCD) cartilage. Osteoarthr. Cartil. 2002, 10, 653–661. [Google Scholar] [CrossRef] [Green Version]
- Boskey, A.L.; Imbert, L. Bone quality changes associated with aging and disease: A review. Ann. N. Y. Acad. Sci. 2017, 1410, 93–106. [Google Scholar] [CrossRef]
- El Demellawy, D.; Davila, J.; Shaw, A.; Nasr, Y. Brief Review on Metabolic Bone Disease. Acad. Forensic Pathol. 2018, 8, 611–640. [Google Scholar] [CrossRef]
- Bruyette, D.S.; Bexfield, N.; Chretin, J.D.; Kidd, L.; Kube, S.; Langston, C.; Owen, T.J.; Oyama, M.A.; Peterson, N.; Reiter, L.V.; et al. Canine Hypothyroidism. In Clinical Small Animal Internal Medicine; Wiley Online Library: Hoboken, NJ, USA, 2020; Chapter 9; pp. 1002–1097. [Google Scholar]
- Jee, W.S.; Yao, W. Overview: Animal models of osteopenia and osteoporosis. J. Musculoskelet. Neuronal Interact. 2001, 3, 193–207. [Google Scholar]
- Duran, A.; Serrano, M.; Leitges, M.; Flores, J.M.; Picard, S.; Brown, J.P.; Moscat, J.; Diaz-Meco, M.T. The atypical PKC-interacting protein p62 is an important mediator of RANK-activated osteoclastogenesis. Dev. Cell 2004, 6, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Kurihara, N.; Hiruma, Y.; Zhou, H.; Subler, M.A.; Dempster, D.W.; Singer, F.R.; Reddy, S.V.; Gruber, H.E.; Windle, J.J.; Roodman, G.D. Mutation of the sequestosome 1 (p62) gene increases osteoclasto-genesis but does not induce Paget disease. J. Clin. Investig. 2007, 117, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Hiruma, Y.; Kurihara, N.; Subler, M.A.; Zhou, H.; Boykin, C.S.; Zhang, H.; Ishizuka, S.; Dempster, D.W.; Roodman, G.D.; Windle, J.J. A SQSTM1/p62 mutation linked to Paget’s disease increases the osteoclastogenic potential of the bone microenvironment. Hum. Mol. Genet. 2008, 17, 3708–3719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daroszewska, A.; van’t Hof, R.J.; Rojas, J.A.; Layfield, R.; Landao-Basonga, E.; Rose, L.; Rose, K.; Ralston, S.H. A point mutation in the ubiquitin-associated domain of SQSMT1 is sufficient to cause a Paget’s disease-like disorder in mice. Hum. Mol. Genet. 2011, 20, 2734–2744. [Google Scholar] [CrossRef] [Green Version]
- Zach, F.; Polzer, F.; Mueller, A.; Gessner, A. p62/sequestosome 1 deficiency accelerates osteoclastogenesis in vitro and leads to Paget’s disease–like bone phenotypes in mice. J. Biol. Chem. 2018, 293, 9530–9541. [Google Scholar] [CrossRef] [Green Version]
- Agas, D.; Sabbieti, M.G. Archetypal autophagic players through new lenses for bone marrow stem/mature cells regulation. J. Cell. Physiol. 2021, 236, 6101–6114. [Google Scholar] [CrossRef]
- Nelson, A.E.; DeVellis, R.F.; Renner, J.B.; Schwartz, T.A.; Conaghan, P.G.; Kraus, V.B.; Jordan, J.M. Quantification of the whole-body burden of radiographic osteoarthritis using factor analysis. Arthritis Res. Ther. 2011, 3, R176. [Google Scholar] [CrossRef] [Green Version]
- Gallaher, H.M.; Butler, J.R.; Wills, R.W.; Priddy, L.B.; Elder, S.H.; Heller, S.M.; Brinkman, E.; Baumgartner, W. Effects of short- and long-term administration of nonsteroidal anti-inflammatory drugs on osteotomy healing in dogs. Vet. Surg. 2019, 48, 1318–1329. [Google Scholar] [CrossRef]
- Vandeweerd, J.M.; Zhao, Y.; Nisolle, J.F.; Zhang, W.; Zhihong, L.; Clegg, P.; Gustin, P. Effect of corticosteroids on articular cartilage: Have animal studies said everything? Fundam. Clin. Pharmacol. 2015, 29, 427–438. [Google Scholar] [CrossRef]
- Nganvongpanit, K.; Pothacharoen, P.; Suwankong, N. The effect of doxycycline on canine hip osteoarthritis: Design of a 6-month clinical trial. J. Vet. Sci. 2009, 10, 239–247. [Google Scholar] [CrossRef]
- Rogachefsky, R.A.; Dean, D.D.; Howell, D.S.; Altman, R.D. Treatment of canine osteoarthritis with sodium pentosan polysulfate and insulin-like growth factor-1. Ann. N. Y. Acad. Sci. 1994, 732, 392–394. [Google Scholar] [CrossRef]
- Johnson, K.A.; Lee, A.H.; Swanson, K.S. Nutrition and nutraceuticals in the changing management of osteoarthritis for dogs and cats. J. Am. Vet. Med. Assoc. 2020, 256, 1335–1341. [Google Scholar] [CrossRef]
- Sanghani-Kerai, A.; Black, C.; Cheng, S.O.; Collins, L.; Schneider, N.; Blunn, G.; Watson, F.; Fitzpatrick, N. Clinical outcomes following intra-articular injection of autologous adipose-derived mesenchymal stem cells for the treatment of osteoarthritis in dogs characterized by weight-bearing asymmetry. Bone Jt. Res. 2021, 10, 650–658. [Google Scholar] [CrossRef]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [Green Version]
- Wagner, M.; Samdal Steinskog, E.S.; Wiig, H. Adipose tissue macrophages: The inflammatory link between obesity and cancer? Expert Opin. Ther. Targets 2015, 19, 527–538. [Google Scholar] [CrossRef]
- Dantzer, R.; O’Connor, J.C.; Freund, G.G.; Johnson, R.W.; Kelley, K.W. From inflammation to sickness and depression: When the immune system subjugates the brain. Nat. Rev. Neurosci. 2008, 9, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Voigt, J.P.; Fink, H. Serotonin controlling feeding and satiety. Behav. Brain Res. 2015, 277, 14–31. [Google Scholar] [CrossRef]
- Miller, A.H.; Haroon, E.; Raison, C.L.; Felger, J.C. Cytokine targets in the brain: Impact on neurotransmitters and neurocircuits. Depress. Anxiety 2013, 30, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Olivier, B. Serotonin: A never-ending story. Eur. J. Pharmacol. 2015, 753, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Planchez, B.; Surget, A.; Belzung, C. Animal models of major depression: Drawbacks and challenges. J. Neural Transm. 2019, 126, 1383–1408. [Google Scholar] [CrossRef] [Green Version]
- Singh, M. Mood, food, and obesity. Front. Psychol. 2014, 5, 925. [Google Scholar] [CrossRef] [Green Version]
- Halenova, T.; Savchuk, O.; Ostapchenko, L.; Chursov, A.; Fridlyand, N.; Komissarov, A.B.; Venanzi, F.; Kolesnikov, S.I.; Sufianov, A.A.; Sherman, M.Y.; et al. P62 plasmid can alleviate diet-induced obesity and metabolic dysfunctions. Oncotarget 2017, 8, 56030–56040. [Google Scholar] [CrossRef]
- Rodriguez, A.; Durán, A.; Selloum, M.; Champy, M.F.; Diez-Guerra, F.J.; Flores, J.M.; Serrano, M.; Auwerx, J.; Diaz-Meco, M.T.; Moscat, J. Mature-onset obesity and insulin resistance in mice deficient in the signaling adapter p62. Cell Metab. 2006, 3, 211–222. [Google Scholar] [CrossRef]
- Zhong, Z.; Umemura, A.; Sanchez-Lopez, E.; Liang, S.; Shalapour, S.; Wong, J.; He, F.; Boassa, D.; Perkins, G.; Ali, S.R.; et al. NF-kB restricts inflammasome activation via elimination of damaged mitochondria. Cell 2016, 164, 896–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veleri, S.; Lazar, C.H.; Chang, B.; Sieving, P.A.; Banin, E.; Swaroop, A. Biology and therapy of inherited retinal degenerative disease: Insights from mouse models. Dis. Model Mech. 2015, 8, 109–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltran, W.A. The use of canine models of inherited retinal degeneration to test novel therapeutic approaches. Vet. Ophthalmol. 2009, 12, 192–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and its role in agerelated macular degeneration. Cell. Mol. Life Sci. 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ebrahimi, K.B.; Chyn, M.; Cano, M.; Handa, J.T. Biology of p62/sequestosome-1 in Age-Related Macular Degeneration (AMD). Adv. Exp. Med. Biol. 2016, 854, 17–22. [Google Scholar]
- Telegina, D.V.; Kozhevnikova, O.S.; Bayborodin, S.I.; Kolosova, N.G. Contributions of age-related alterations of the retinal pigment epithelium and of glia to the AMD-like pathology in OXYS rats. Sci. Rep. 2017, 7, 41533. [Google Scholar] [CrossRef]
- Kolosova, N.G.; Kozhevnikova, O.S.; Telegina, D.V.; Fursova, A.Z.; Stefanova, N.A.; Muraleva, N.A.; Venanzi, F.; Sherman, M.Y.; Kolesnikov, S.I.; Sufianov, A.A.; et al. p62/SQSTM1 coding plasmid prevents age related macular degeneration in a rat model. Aging 2018, 10, 2136–2147. [Google Scholar] [CrossRef]
- Moscat, J.; Diaz-Meco, M.T.; Wooten, M.W. Signal integration and diversification through the p62 scaffold protein. Trends Biochem. Sci. 2007, 32, 95–100. [Google Scholar] [CrossRef]
- Kitamura, H.; Torigoe, T.; Asanuma, H.; Hisasue, S.I.; Suzuki, K.; Tsukamoto, T.; Satoh, M.; Sato, N. Cytosolic overexpression of p62 sequestosome 1 in neoplastic prostate tissue. Histopathology 2006, 48, 157–161. [Google Scholar] [CrossRef]
- Rolland, P.; Madjd, Z.; Durrant, L.; Ellis, I.O.; Layfield, R.; Spendlove, I. The ubiquitin-binding protein p62 is expressed in breast cancers showing features of aggressive disease. Endocr. Relat. Cancer 2007, 14, 73–80. [Google Scholar] [CrossRef]
- Venanzi, F.; Shifrin, V.; Sherman, M.; Gabai, V.; Kiselev, O.; Komissarov, A.; Grudinin, M.; Shartukova, M.; Romanovskaya-Romanko, E.A.; Kudryavets, Y.; et al. Broad-spectrum anti-tumor and anti-metastatic DNA vaccine based on p62-encoding vector. Oncotarget 2013, 4, 1829–1835. [Google Scholar] [CrossRef] [Green Version]
- Sleeckx, N.; Rooster, H.D.; Veldhuis Kroeze, E.J.; Van Ginneken, C.; Van Brantegem, L. Canine mammary tumours, an overview. Reprod. Domest. Anim. 2011, 46, 1112–1131. [Google Scholar] [CrossRef]
- Sorenmo, K.; Worley, D.; Zappulli, V. Tumors of the mammary gland. In Withrow and MacEwen’s Small Animal Clinical Oncology, 6th ed.; Vail, D., Thamm, D., Liptack, J., Eds.; Elsevier: St. Louis, MO, USA, 2020; pp. 604–625. [Google Scholar]
- Salas, Y.; Márquez, A.; Diaz, D.; Romero, L. Epidemiological study of mammary tumors in female dogs diagnosed during the period 2002-2012: A growing animal health problem. PLoS ONE 2015, 10, e0127381. [Google Scholar] [CrossRef] [Green Version]
- Arenas, C.; Peña, L.; Granados-Soler, J.L.; Pérez-Alenza, M.D. Adjuvant therapy for highly malignant canine mammary tumours: Cox-2 inhibitor versus chemotherapy: A case–control prospective study. Vet. Rec. 2016, 179, 125. [Google Scholar] [CrossRef] [Green Version]
- Tran, C.M.; Moore, A.S.; Frimberger, A.E. Surgical treatment of mammary carcinomas in dogs with or without postoperative chemotherapy. Vet. Comp. Oncol. 2016, 14, 252–262. [Google Scholar] [CrossRef]
- Valdivia, G.; Alonso-Diez, Á.; Pérez-Alenza, D.; Peña, L. From Conventional to Precision Therapy in Canine Mammary Cancer: A Comprehensive Review. Front. Vet. Sci. 2021, 8, 623800. [Google Scholar] [CrossRef]
- Nguyen, F.; Pena, L.; Ibisch, C.; Loussouarn, D.; Gama, A.; Rieder, N.; Belousov, A.; Campone, M.; Abadie, J. Canine invasive mammary carcinomas as models of human breast cancer. Part 1: Natural history and prognostic factors. Breast Cancer Res.Treat. 2018, 167, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Sinha, G. Companion therapeutics. Nat. Biotechnol. 2014, 32, 12–14. [Google Scholar] [CrossRef]
- Gabai, V.; Venanzi, F.M.; Bagashova, E.; Rud, O.; Mariotti, F.; Vullo, C.; Catone, G.; Sherman, M.Y.; Concetti, A.; Chursov, A.; et al. Pilot study of p62 DNA vaccine in dogs with mammary tumors. Oncotarget 2014, 5, 12803–12810. [Google Scholar] [CrossRef] [Green Version]
- Venanzi, F.M.; Gabai, V.; Mariotti, F.; Magi, G.E.; Vullo, C.; Sufianov, A.A.; Kolesnikov, S.I.; Shneider, A. p62-DNA-encoding plasmid reverts tumor grade, changes tumor stroma, and enhances anticancer immunity. Aging 2019, 11, 10711–10722. [Google Scholar] [CrossRef]
- Ponomarenko, D.M.; Klimova, I.D.; Chapygina, Y.A.; Dvornichenko, V.V.; Zhukova, N.V.; Orlova, R.V.; Manikhas, G.M.; Zyryanov, A.V.; Burkhanova, L.A.; Badrtdinova, I.I.; et al. Safety and efficacy of p62 DNA vaccine ELENAGEN in a first-in-human trial in patients with advanced solid tumors. Oncotarget 2017, 8, 53730–53739. [Google Scholar] [CrossRef] [Green Version]
- Hollingsworth, R.E.; Jansen, K. Turning the corner on therapeutic cancer vaccines. NPJ Vaccines 2019, 8, 4–7. [Google Scholar] [CrossRef]
- Ponomarenko, D.M.; Gabai, V.L.; Sufianov, A.A.; Kolesnikov, S.I.; Shneider, A.M. Response of a chemo-resistant triple-negative breast cancer patient to a combination of p62-encoding plasmid, Elenagen, and CMF chemotherapy. Oncotarget 2020, 11, 294–299. [Google Scholar] [CrossRef]
- Krasny, S.; Polyakov, S.; Zhavrid, E.; Khorau, A.; Baranau, Y.; Kozlovskaya, S.; Sergeeva, O.; Zharkova, K.; Sufianov, A.A.; Gabai, V.; et al. Plasmid encoding p62/SQSTM1 administered in combination with CMF chemotherapy improves outcomes in metastatic triple-negative breast cancer patients. Ann. Oncol. 2021, 32, S457–S515. [Google Scholar] [CrossRef]
- Polyakov, S.; Krasny, S.; Zhavrid, E.; Baranau, Y.; Sergeeva, O.; Zharkova, K.; Khorau, A.; Kozlovskaya, S.; Gabai, V.; Shneider, A. Adding dosing of plasmid encoding p62/SQSTM1 to gemcitabine chemotherapy may provide clinical benefits to patients with platinum-resistant ovarian cancer. Ann. Oncol. 2021, 32, S725–S772. [Google Scholar] [CrossRef]
- Schiffman, J.D.; Breen, M. Comparative oncology: What dogs and other species can teach us about humans with cancer. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140231. [Google Scholar] [CrossRef]
- LeBlanc, A.K.; Mazcko, C.N. Improving human cancer therapy through the evaluation of pet dogs. Nat. Rev. Cancer 2020, 20, 727–742. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, D.L.; Duval, D.L.; Regan, D.P.; Thamm, D.H. Canine sarcomas as a surrogate for the human disease. Pharmacol. Ther. 2018, 188, 80–96. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, N.; Konnai, S.; Takagi, S.; Kagawa, Y.; Okagawa, T.; Nishimori, A.; Ikebuchi, R.; Izumi, Y.; Deguchi, T.; Nakajima, C.; et al. A canine chimeric monoclonal antibody targeting PD-L1 and its clinical efficacy in canine oral malignant melanoma or undifferentiated sarcoma. Sci. Rep. 2017, 7, 8951. [Google Scholar] [CrossRef] [PubMed]
- Bergman, P.J. Cancer immunotherapies. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 881–902. [Google Scholar] [CrossRef]
- Rue, S.M.; Eckelman, B.P.; Efe, J.A.; Bloink, K.; Deveraux, Q.L.; Lowery, D.; Nasoff, M. Identification of a candidate therapeutic antibody for treatment of canine B-cell lymphoma. Vet. Immunol. Immunopathol. 2015, 164, 148–159. [Google Scholar] [CrossRef]
- Mizuno, T.; Kato, Y.; Kaneko, M.K.; Sakai, Y.; Shiga, T.; Kato, M.; Tsukui, T.; Takemoto, H.; Tokimasa, A.; Baba, K.; et al. Generation of a canine anti-canine CD20 antibody for canine lymphoma treatment. Sci. Rep. 2020, 10, 11476. [Google Scholar] [CrossRef]
- Haran, K.P.; Lockhart, A.; Xiong, A.; Radaelli, E.; Savickas, P.J.; Posey, A.; Mason, N.J. Generation and validation of an antibody to canine CD19 for diagnostic and future therapeutic purposes. Vet. Pathol. 2020, 57, 241–252. [Google Scholar] [CrossRef]
- Mariotti, F.; Magi, G.E.; Gavazza, A.; Vincenzetti, S.; Komissarov, A.; Shneider, A.; Venanzi, F.M. p62/SQSTM1 expression in canine mammary tumours: Evolutionary notes. Vet. Comp. Oncol. 2019, 17, 570–577. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabbieti, M.G.; Marchegiani, A.; Sufianov, A.A.; Gabai, V.L.; Shneider, A.; Agas, D. P62/SQSTM1 beyond Autophagy: Physiological Role and Therapeutic Applications in Laboratory and Domestic Animals. Life 2022, 12, 539. https://doi.org/10.3390/life12040539
Sabbieti MG, Marchegiani A, Sufianov AA, Gabai VL, Shneider A, Agas D. P62/SQSTM1 beyond Autophagy: Physiological Role and Therapeutic Applications in Laboratory and Domestic Animals. Life. 2022; 12(4):539. https://doi.org/10.3390/life12040539
Chicago/Turabian StyleSabbieti, Maria Giovanna, Andrea Marchegiani, Albert A. Sufianov, Vladimir L. Gabai, Alexander Shneider, and Dimitrios Agas. 2022. "P62/SQSTM1 beyond Autophagy: Physiological Role and Therapeutic Applications in Laboratory and Domestic Animals" Life 12, no. 4: 539. https://doi.org/10.3390/life12040539
APA StyleSabbieti, M. G., Marchegiani, A., Sufianov, A. A., Gabai, V. L., Shneider, A., & Agas, D. (2022). P62/SQSTM1 beyond Autophagy: Physiological Role and Therapeutic Applications in Laboratory and Domestic Animals. Life, 12(4), 539. https://doi.org/10.3390/life12040539