
Non-Photosynthetic Melainabacteria (Cyanobacteria) in Human Gut: Characteristics and Association with Health



Abstract
:1. Introduction
2. The New Class Melainabacteria of the Phylum Cyanobacteria
3. Ecology of Melainabacteria
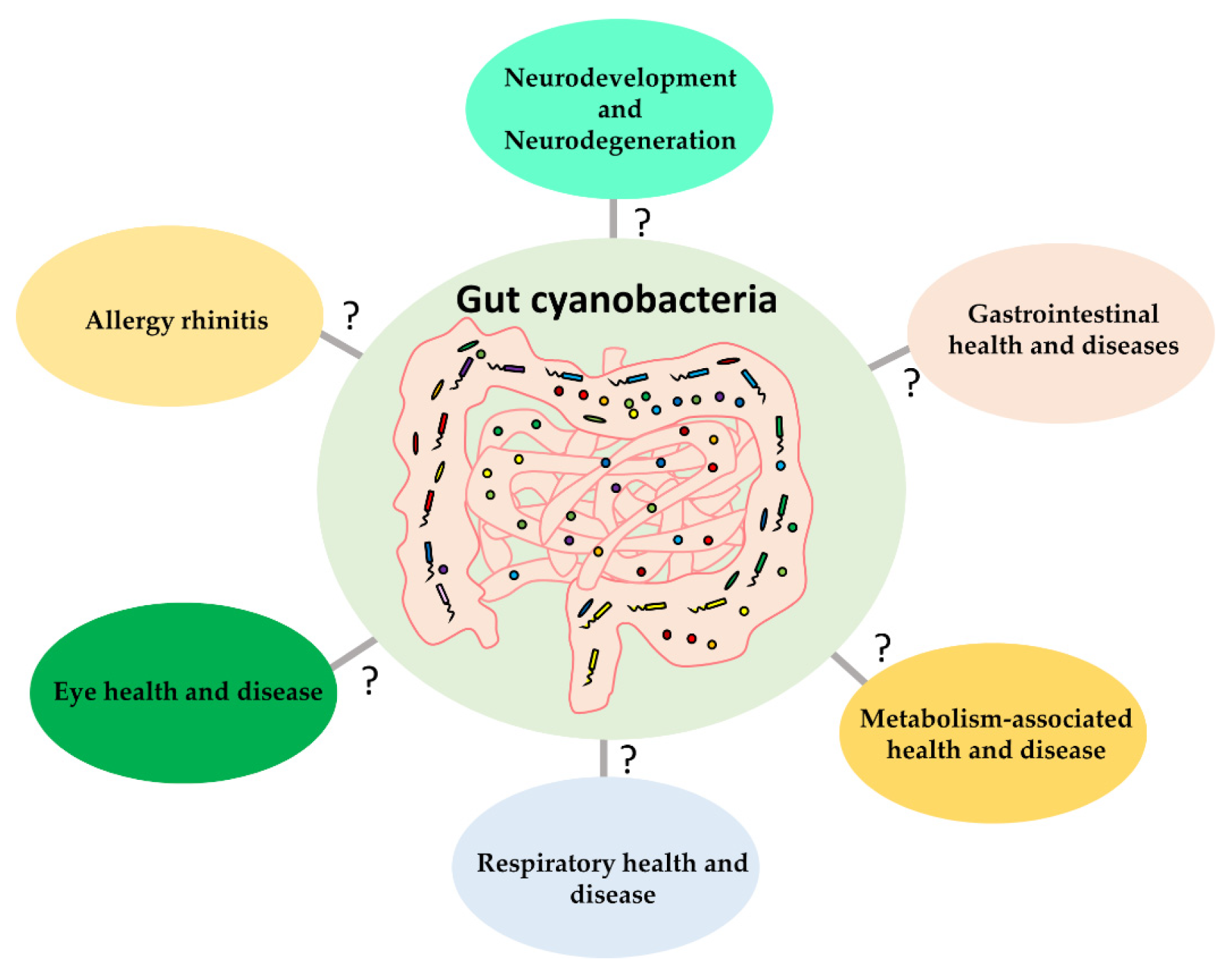
4. The Health Relevance of Melainabacteria in the Human Gut
4.1. Neurodevelopment and Neurodegeneration
4.2. Gastrointestinal and Metabolic Diseases
4.3. Other Diseases
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schopf, J.W.; Packer, B.M. Early Archean (3.3-billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia. Science 1987, 237, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Lyons, T.W.; Reinhard, C.T.; Planavsky, N.J. The rise of oxygen in Earth’s early ocean and atmosphere. Nature 2014, 506, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Diez, B.; Ininbergs, K. Ecological importance of cyanobacteria. In Cyanobacteria: An Economic Perspective; Sharma, N.K., Rai, A.K., Stal, L.J., Eds.; Wiley: Hoboken, NJ, USA, 2014; pp. 43–63. [Google Scholar]
- Hu, C.; Rzymski, P. Programmed Cell Death-Like and Accompanying Release of Microcystin in Freshwater Bloom-Forming Cyanobacterium Microcystis: From Identification to Ecological Relevance. Toxins 2019, 11, 706. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzi, S.C.; Sharon, I.; Wrighton, K.C.; Koren, O.; Hug, L.A.; Thomas, B.C.; Goodrich, J.K.; Bell, J.T.; Spector, T.D.; Banfield, J.F.; et al. The human gut and groundwater harbor non-photosynthetic bacteria belonging to a new candidate phylum sibling to Cyanobacteria. Elife 2013, 2, e01102. [Google Scholar] [CrossRef] [PubMed]
- Utami, Y.D.; Kuwahara, H.; Murakami, T.; Morikawa, T.; Sugaya, K.; Kihara, K.; Yuki, M.; Lo, N.; Deevong, P.; Hasin, S.; et al. Phylogenetic Diversity and Single-Cell Genome Analysis of “Melainabacteria”, a Non-Photosynthetic Cyanobacterial Group, in the Termite Gut. Microbes Environ. 2018, 33, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soo, R.M.; Skennerton, C.T.; Sekiguchi, Y.; Imelfort, M.; Paech, S.J.; Dennis, P.G.; Steen, J.A.; Parks, D.H.; Tyson, G.W.; Hugenholtz, P. An expanded genomic representation of the phylum cyanobacteria. Genome Biol. Evol. 2014, 6, 1031–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monchamp, M.E.; Spaak, P.; Pomati, F. Long Term Diversity and Distribution of Non-photosynthetic Cyanobacteria in Peri-Alpine Lakes. Front. Microbiol. 2018, 9, 3344. [Google Scholar] [CrossRef] [Green Version]
- Soo, R.M.; Hemp, J.; Parks, D.H.; Fischer, W.W.; Hugenholtz, P. On the origins of oxygenic photosynthesis and aerobic respiration in Cyanobacteria. Science 2017, 355, 1436–1440. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Pichel, F.; Zehr, J.P.; Bhattacharya, D.; Pakrasi, H.B. What’s in a name? The case of cyanobacteria. J. Phycol. 2020, 56, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Dextro, R.B.; Delbaje, E.; Cotta, S.R.; Zehr, J.P.; Fiore, M.F. Trends in Free-access Genomic Data Accelerate Advances in Cyanobacteria Taxonomy. J. Phycol. 2021, 57, 1392–1402. [Google Scholar] [CrossRef]
- Dethlefsen, L.; Relman, D.A. Incomplete recovery and individualized responses of the human distal gut microbiota to repeated antibiotic perturbation. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4554–4561. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Tajima, K.; Arai, S.; Ogata, K.; Nagamine, T.; Matsui, H.; Nakamura, M.; Aminov, R.I.; Benno, Y. Rumen bacterial community transition during adaptation to high-grain diet. Anaerobe 2000, 6, 273–284. [Google Scholar] [CrossRef]
- Hongoh, Y.; Ohkuma, M.; Kudo, T. Molecular analysis of bacterial microbiota in the gut of the termite Reticulitermes speratus (Isoptera; Rhinotermitidae). FEMS Microbiol. Ecol. 2003, 44, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Backhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.J.; Ramey, R.R.; Bircher, J.S.; Schlegel, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Soo, R.M.; Woodcroft, B.J.; Parks, D.H.; Tyson, G.W.; Hugenholtz, P. Back from the dead; the curious tale of the predatory cyanobacterium Vampirovibrio chlorellavorus. PeerJ 2015, 3, e968. [Google Scholar] [CrossRef] [Green Version]
- Hovde, B.T.; Steichen, S.A.; Starkenburg, S.R.; Brown, J.K. Vampirovibrio chlorellavorus draft genome sequence, annotation, and preliminary characterization of pathogenicity determinants. Phycol. Res. 2020, 68, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Steichen, S.A.; Li, X.H.; Ogden, K.; Brown, J.K. Association of Vampirovibrio chlorellavorus with decline and death of Chlorella sorokiniana in outdoor reactors. J. Appl. Phycol. 2019, 31, 1131–1142. [Google Scholar] [CrossRef]
- Soo, R.M.; Hemp, J.; Hugenholtz, P. Evolution of photosynthesis and aerobic respiration in the cyanobacteria. Free Radic. Biol. Med. 2019, 140, 200–205. [Google Scholar] [CrossRef]
- Cano-Diaz, C.; Maestre, F.T.; Eldridge, D.J.; Singh, B.K.; Bardgett, R.D.; Fierer, N.; Delgado-Baquerizo, M. Contrasting environmental preferences of photosynthetic and non-photosynthetic soil cyanobacteria across the globe. Glob. Ecol. Biogeogr. 2020, 29, 2025–2038. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Vijay, A.; Valdes, A.M. Role of the gut microbiome in chronic diseases: A narrative review. Eur. J. Clin. Nutr. 2021. [Google Scholar] [CrossRef]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat. Commun. 2017, 8, 1784. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Pramanik, S. Structural diversity, functional aspects and future therapeutic applications of human gut microbiome. Arch. Microbiol. 2021, 203, 5281–5308. [Google Scholar] [CrossRef]
- Di Gioia, D.; Bozzi Cionci, N.; Baffoni, L.; Amoruso, A.; Pane, M.; Mogna, L.; Gaggia, F.; Lucenti, M.A.; Bersano, E.; Cantello, R.; et al. A prospective longitudinal study on the microbiota composition in amyotrophic lateral sclerosis. BMC Med. 2020, 18, 153. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, Z.; Zhang, Q.; Sun, Z.; Su, Y.; Song, J.; Wang, B.; Gao, R. Preliminary evidence for an influence of exposure to polycyclic aromatic hydrocarbons on the composition of the gut microbiota and neurodevelopment in three-year-old healthy children. BMC Pediatr. 2021, 21, 86. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, J.; Zheng, J.; Hu, G.; Wang, J.; Huang, C.; Lou, L.; Wang, X.; Zeng, Y. Mucosal adherent bacterial dysbiosis in patients with colorectal adenomas. Sci. Rep. 2016, 6, 26337. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Li, Y.; Li, J.; Yang, J.; Shang, L.; He, X.; Liu, L.; Luo, Y.; Xie, X. Intestinal microbiota profiles in infants with acute gastroenteritis caused by rotavirus and norovirus infection: A prospective cohort study. Int. J. Infect. Dis. 2021, 111, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Sarangi, A.N.; Goel, A.; Singh, A.; Sasi, A.; Aggarwal, R. Faecal bacterial microbiota in patients with cirrhosis and the effect of lactulose administration. BMC Gastroenterol. 2017, 17, 125. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, R.C.; Wang, Z.; Usyk, M.; Sotres-Alvarez, D.; Daviglus, M.L.; Schneiderman, N.; Talavera, G.A.; Gellman, M.D.; Thyagarajan, B.; Moon, J.Y.; et al. Gut microbiome composition in the Hispanic Community Health Study/Study of Latinos is shaped by geographic relocation, environmental factors, and obesity. Genome Biol. 2019, 20, 219. [Google Scholar] [CrossRef]
- Oduaran, O.H.; Tamburini, F.B.; Sahibdeen, V.; Brewster, R.; Gomez-Olive, F.X.; Kahn, K.; Norris, S.A.; Tollman, S.M.; Twine, R.; Wade, A.N.; et al. Gut microbiome profiling of a rural and urban South African cohort reveals biomarkers of a population in lifestyle transition. BMC Microbiol. 2020, 20, 330. [Google Scholar] [CrossRef]
- Chumpitazi, B.P.; Hoffman, K.L.; Smith, D.P.; McMeans, A.R.; Musaad, S.; Versalovic, J.; Petrosino, J.F.; Shulman, R.J. Fructan-sensitive children with irritable bowel syndrome have distinct gut microbiome signatures. Aliment. Pharmacol. Ther. 2021, 53, 499–509. [Google Scholar] [CrossRef]
- Cai, X.S.; Deng, L.; Ma, X.G.; Guo, Y.S.; Feng, Z.T.; Liu, M.Q.; Guan, Y.B.; Huang, Y.T.; Deng, J.X.; Li, H.W.; et al. Altered diversity and composition of gut microbiota in Wilson’s disease. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Zhu, L.; Xu, F.; Wan, W.; Yu, B.; Tang, L.; Yang, Y.; Du, Y.; Chen, Z.; Xu, H. Gut microbial characteristics of adult patients with allergy rhinitis. Microb. Cell Factories 2020, 19, 171. [Google Scholar] [CrossRef]
- Zhang, W.Q.; Zhao, S.K.; Luo, J.W.; Dong, X.P.; Hao, Y.T.; Li, H.; Shan, L.; Zhou, Y.; Shi, H.B.; Zhang, Z.Y.; et al. Alterations of fecal bacterial communities in patients with lung cancer. Am. J. Transl. Res. 2018, 10, 3171–3185. [Google Scholar]
- Sublette, M.G.; Cross, T.L.; Korcarz, C.E.; Hansen, K.M.; Murga-Garrido, S.M.; Hazen, S.L.; Wang, Z.; Oguss, M.K.; Rey, F.E.; Stein, J.H. Effects of Smoking and Smoking Cessation on the Intestinal Microbiota. J. Clin. Med. 2020, 9, 2963. [Google Scholar] [CrossRef]
- Shi, T.T.; Xin, Z.; Hua, L.; Wang, H.; Zhao, R.X.; Yang, Y.L.; Xie, R.R.; Liu, H.Y.; Yang, J.K. Comparative assessment of gut microbial composition and function in patients with Graves’ disease and Graves’ orbitopathy. J. Endocrinol. Investig. 2021, 44, 297–310. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce beta-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S.R. Blue-green algae or cyanobacteria in the intestinal micro-flora may produce neurotoxins such as Beta-N-Methylamino-L-Alanine (BMAA) which may be related to development of amyotrophic lateral sclerosis, Alzheimer’s disease and Parkinson-Dementia-Complex in humans and Equine Motor Neuron Disease in horses. Med. Hypotheses 2013, 80, 103. [Google Scholar] [CrossRef]


{kind=link}
| Strain | Environment | Genome Size (Mbp) | Completeness | GC (%) | Anaerobic | Flagella | Shape | Reference |
|---|---|---|---|---|---|---|---|---|
| Order Gastranaerophilales | ||||||||
| ACD20 | aquifer | 3.0 | Near Complete | 33.5 | anaerobic | + | N.A. | [5] |
| MEL.A1 | human gut | 1.9 | Complete | 32.9 | anaerobic | − | N.A. | [5] |
| MEL.A2 | human gut | 1.2 | Partial | 30.6 | anaerobic | − | N.A. | [5] |
| MEL.B1 | human gut | 2.3 | Complete | 35.3 | anaerobic | + | N.A. | [5] |
| MEL.B2 | human gut | 2.3 | Complete | 36.3 | anaerobic | + | N.A. | [5] |
| MEL.C1 | human gut | 2.1 | Complete | 34.1 | anaerobic | − | N.A. | [5] |
| MEL.C2 | human gut | 2.2 | Near Complete | 35.3 | anaerobic | − | N.A. | [5] |
| MEL.C3 | human gut | 1.3 | Partial | 29.9 | anaerobic | − | N.A. | [5] |
| Tpq-Mel-01 | termite gut | 0.96 | Partial | 42.5 | anaerobic | + | Rod | [6] |
| Zag_221 | koala gut | 1.8 | complete | 38.5 | anaerobic | − | N.A. | [7] |
| Zag_1 | koala gut | 2 a | Near Complete | 34.9 | anaerobic | − | N.A. | [7] |
| Zag_111 | koala gut | 2.2 a | Near Complete | 36.7 | anaerobic | − | N.A. | [7] |
| MH_37 | human gut | 2.2 | complete | 34.1 | anaerobic | − | N.A. | [7] |
| Order Caenarcaniphilales | ||||||||
| UASB_169 | bioreactor | 1.8 | complete | 27.5 | anaerobic | − | N.A. | [7] |
| Order Obscuribacterales | ||||||||
| EBPR_351 | bioreactor | 5.5 | near complete | 49.4 | microaerophilic | − | N.A. | [7] |
| Order Vampirovibrionales | ||||||||
| NCIB 11383 | coculture with Chlorella vulgaris | 3.0 | complete | 51.4 | microaerophilic | + | N.A. | [20] |
| AZ_1 | algal cultivation ponds | 2.8 | near complete | 54.8 | microaerophilic | + | Sphere | [21,22] |
| AZ_2 | algal cultivation ponds | 3.0 | near complete | 53.0 | microaerophilic | + | Sphere | [21,22] |
| Country | Year | Subject | Disease | Gut Cyanobacterial Abundance | Remark | Reference |
|---|---|---|---|---|---|---|
| Italy | 2020 | Control group (n = 50) vs. diseased group (n = 50) | Amyotrophic lateral sclerosis | Disease group > control group | The finding indicated that cyanobacteria could be involved in the pathogenesis of neurodegenerative diseases | [31] |
| China | 2021 | 3-year old healthy children (n = 38) | Neurodevelopment disorder | Gut cyanobacteria negatively correlated with the neurodevelopment in Adaptation | The finding implied the negative effect of cyanobacteria on neurodevelopment in adaptation | [32] |
| China | 2016 | Control group (n = 20) vs. diseased group (n = 31) | Colon adenomas | Disease group > control | The finding implied the association between the colorectal pre-neoplastic lesion and the increase in gut cyanobacterial abundance. | [33] |
| China | 2021 | Healthy infants (n = 25), infants with acute rotaviral gastroenteritis (n = 18), infants with acute noraviral gastroenteritis (n = 24) | Acute gastroenteritis | Disease group > control | The finding implied the association between human norovirus infection and the increase in gut cyanobacteria. | [34] |
| India | 2017 | Control group (n = 18) vs. diseased group (n = 35) | Cirrhosis | Disease group < control | The finding implied the association between cirrhosis and the reduction in gut cyanobacteria. | [35] |
| USA | 2019 | Hispanic/Latino adults (n = 1647) | Obesity | Gut cyanobacteria negatively correlated with obesity | The finding implied the negative association between obesity and gut cyanobacteria. | [36] |
| South Africa | 2020 | Rural community (n = 119) vs. urban community (n = 51) | - | Rural community > urban community | Gut Melainabacteria can be more abundant in the rural populations | [37] |
| USA | 2020 | Frutcan-sensitive (n = 17) vs. frutcan-tolerant children (n = 21) with irritable bowel syndrome | Irritable bowel syndrome | Fructan-sensitive > fructan-tolerant | Fructan-sensitive children were enriched in the gut cyanobacteria during fructan challenge. | [38] |
| China | 2020 | Control group (n = 16) vs. diseased group (n = 14) | Wilson’s Disease | Disease > Control | The finding implied the association between the Wilson’s disease and the higher gut cyanobacterial abundance. | [39] |
| China | 2020 | Control group (n = 31) vs. allergy-rhinitis (AR) group (n = 33) | Allergy Rhinitis | Disease > Control | The finding implied the association between allergy rhinitis and the higher gut cyanobacterial abundance. | [40] |
| China | 2018 | Healthy controls (n = 41) vs. Lung cancer (n = 41) | Lung cancer | Disease group > Control | The lung cancer group had a significantly higher level of gut cyanobacteria compared to the health group. | [41] |
| USA | 2020 | Smokers (n = 36) | - | Gut cyanobacteria positively correlated with exhaled CO levels | The finding implied the positive association between CO level and gut cyanobacteria among the current smokers. | [42] |
| China | 2021 | Patients with Graves’ disease (n = 30), Graves’ orbitopathy (n = 33) vs. healthy subjects (n = 32) | Graves’ disease Graves’ orbitopathy | Significant difference in the gut cyanobacterial abundance among the studied groups. | The finding implied the association of the changing gut cyanobacterial abundance with Graves’ disease and Graves’ orbitopathy. | [43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, C.; Rzymski, P. Non-Photosynthetic Melainabacteria (Cyanobacteria) in Human Gut: Characteristics and Association with Health. Life 2022, 12, 476. https://doi.org/10.3390/life12040476
Hu C, Rzymski P. Non-Photosynthetic Melainabacteria (Cyanobacteria) in Human Gut: Characteristics and Association with Health. Life. 2022; 12(4):476. https://doi.org/10.3390/life12040476
Chicago/Turabian StyleHu, Chenlin, and Piotr Rzymski. 2022. "Non-Photosynthetic Melainabacteria (Cyanobacteria) in Human Gut: Characteristics and Association with Health" Life 12, no. 4: 476. https://doi.org/10.3390/life12040476
APA StyleHu, C., & Rzymski, P. (2022). Non-Photosynthetic Melainabacteria (Cyanobacteria) in Human Gut: Characteristics and Association with Health. Life, 12(4), 476. https://doi.org/10.3390/life12040476