
Novel Lineage of Infectious Bronchitis Virus from Sub-Saharan Africa Identified by Random Amplification and Next-Generation Sequencing of Viral Genome

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus Isolates
2.2. Whole-Genome Sequencing
2.3. Sequence Data and Analysis Tools
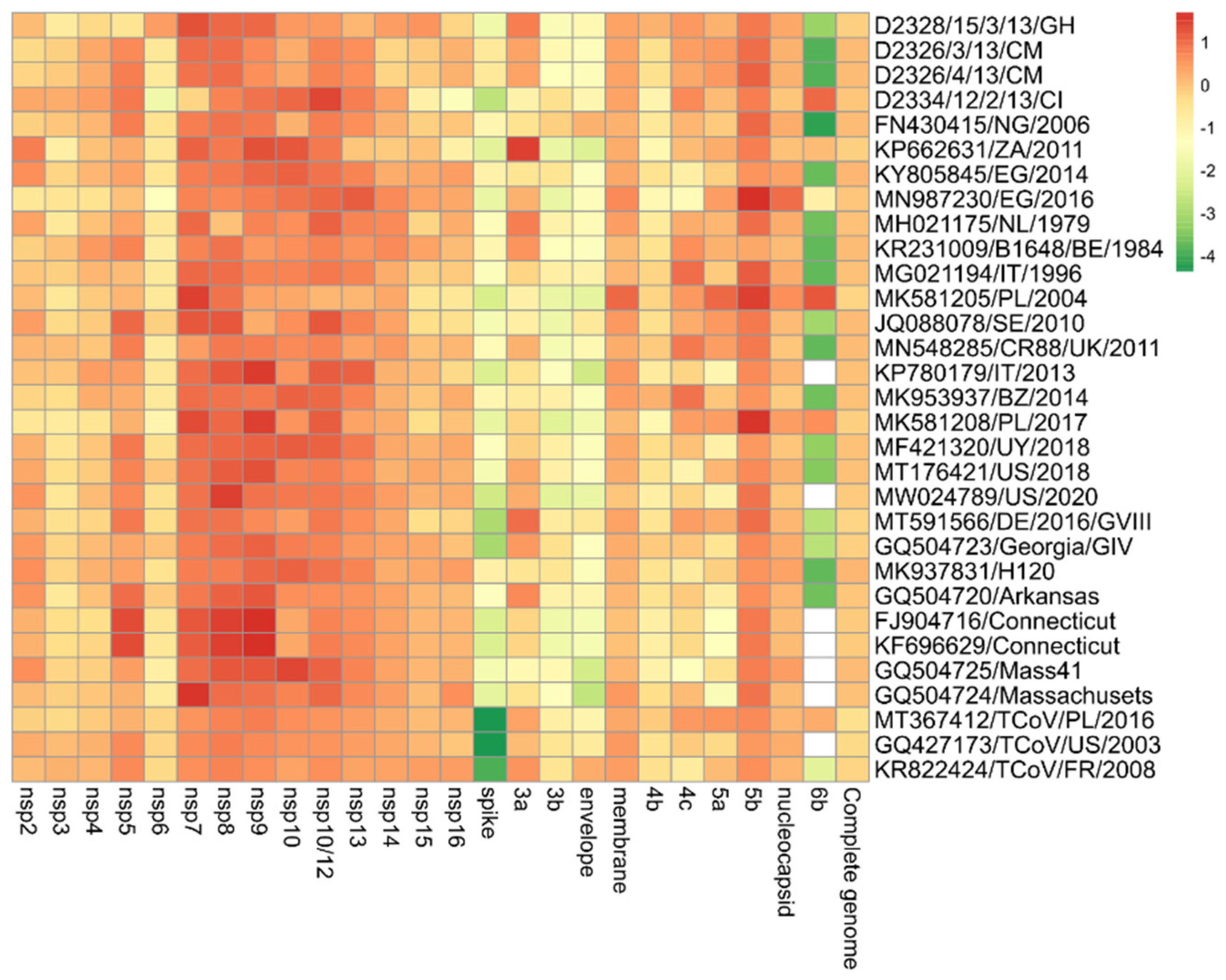
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Wit, J.J.; Cook, J.K.A.; van der Heijden, H.M.J.F. Infectious Bronchitis Virus Variants: A Review of the History, Current Situation and Control Measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Schalk, A.F.; Hawn, M.C. An Apparently New Respiratory Disease of Baby Chicks. J. Am. Vet. Med. Assoc. 1931, 78, 413–423. [Google Scholar]
- Beach, J.R.; Schalm, O.W. A Filterable Virus, Distinct from That of Laryngotracheitis, the Cause of a Respiratory Disease of Chicks. Poult. Sci. 1936, 15, 199–206. [Google Scholar] [CrossRef]
- Johnson, R.B.; Marquardt, W.W. The Neutralizing Characteristics of Strains of Infectious Bronchitis Virus as Measured by the Constant-Virus Variable-Serum Method in Chicken Tracheal Cultures. Avian Dis. 1975, 19, 82–90. [Google Scholar] [CrossRef]
- Lohr, J.E. Serologic Differences between Strains of Infectious Bronchitis Virus from New Zealand, Australia, and the United States. Avian Dis. 1976, 20, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Ignjatovic, J.; McWaters, P.G.Y. 1991 Monoclonal Antibodies to Three Structural Proteins of Avian Infectious Bronchitis Virus: Characterization of Epitopes and Antigenic Differentiation of Australian Strains. J. Gen. Virol. 1991, 72, 2915–2922. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Davis, P.J. Sequence Analysis of Strains of Avian Infectious Bronchitis Coronavirus Isolated during the 1960s in the U.K. Arch. Virol. 1993, 130, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Junker, D.; Collisson, E.W. Evidence of Natural Recombination within the S1 Gens of Infectious Bronchitis Virus. Virology 1993, 192, 710–716. [Google Scholar] [CrossRef]
- Capua, I.; Gough, R.E.; Mancini, M.; Casaccia, C.; Weiss, C. A ‘Novel’ Infectious Bronchitis Strain Infecting Broiler Chickens in Italy. J. Vet. Med. Ser. B 1994, 41, 83–89. [Google Scholar] [CrossRef]
- Lai, M.M.C.; Cavanagh, D. The Molecular Biology of Coronaviruses. In Advances in Virus Research; Maramorosch, K., Murphy, F.A., Shatkin, A.J., Eds.; Academic Press: Amsterdam, The Netherlands, 1997; Volume 48, pp. 1–100. [Google Scholar]
- Cao, J.; Wu, C.-C.; Lin, T.L. Complete Nucleotide Sequence of Polyprotein Gene 1 and Genome Organization of Turkey Coronavirus. Virus Res. 2008, 136, 43–49. [Google Scholar] [CrossRef]
- Cavanagh, D.; Elus, M.M.; Cook, J.K. Relationship between Sequence Variation in the S1 Spike Protein of Infectious Bronchitis Virus and the Extent of Cross-Protection in Vivo. Avian Pathol. 1997, 26, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 Gene-Based Phylogeny of Infectious Bronchitis Virus: An Attempt to Harmonize Virus Classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, L.; Zhao, W.; Liu, L.; Zhao, Y.; Shao, Y.; Li, H.; Han, Z.; Liu, S. Identification and Molecular Characterization of a Novel Serotype Infectious Bronchitis Virus (GI-28) in China. Vet. Microbiol. 2017, 198, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhao, W.; Han, Z.; Chen, Y.; Zhao, Y.; Sun, J.; Li, H.; Shao, Y.; Liu, L.; Liu, S. Genome Characterization, Antigenicity and Pathogenicity of a Novel Infectious Bronchitis Virus Type Isolated from South China. Infect. Genet. Evol. 2017, 54, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Xu, L.; Ren, M.; Shen, J.; Han, Z.; Sun, J.; Zhao, Y.; Liu, S. Novel Genotype of Infectious Bronchitis Virus Isolated in China. Vet. Microbiol. 2019, 230, 178–186. [Google Scholar] [CrossRef]
- Domanska-Blicharz, K.; Sajewicz-Krukowska, J.; Lisowska, A. New PA/1220/98-like Variant of Infectious Bronchitis Virus in Poland. Avian Pathol. 2020, 49, 380–388. [Google Scholar] [CrossRef]
- Brown Jordan, A.; Fusaro, A.; Blake, L.; Milani, A.; Zamperin, G.; Brown, G.; Carrington, C.V.F.; Monne, I.; Oura, C.A.L. Characterization of Novel, Pathogenic Field Strains of Infectious Bronchitis Virus (IBV) in Poultry in Trinidad and Tobago. Transbound. Emerg. Dis. 2020, 67, 2775–2788. [Google Scholar] [CrossRef]
- Kouakou, A.V.; Kouakou, V.; Kouakou, C.; Godji, P.; Kouassi, A.L.; Krou, H.A.; Langeois, Q.; Webby, R.J.; Ducatez, M.F.; Couacy-Hymann, E. Prevalence of Newcastle Disease Virus and Infectious Bronchitis Virus in Avian Influenza Negative Birds from Live Bird Markets and Backyard and Commercial Farms in Ivory-Coast. Res. Vet. Sci. 2015, 102, 83–88. [Google Scholar] [CrossRef]
- Shittu, I.; Gado, D.A.; Meseko, C.A.; Nyam, D.C.; Olawuyi, K.A.; Moses, G.D.; Chinyere, C.N.; Joannis, T.M. Occurrence of Infectious Bronchitis in Layer Birds in Plateau State, North Central Nigeria. Open Vet. J. 2019, 9, 74–80. [Google Scholar] [CrossRef]
- Khataby, K.; Souiri, A.; Kasmi, Y.; Loutfi, C.; Ennaji, M.M. Current Situation, Genetic Relationship and Control Measures of Infectious Bronchitis Virus Variants Circulating in African Regions. J. Basic Appl. Zool. 2016, 76, 20–30. [Google Scholar] [CrossRef]
- Oyejide, A.; Demangam, V.; Akinyemi, J. Serological Survey of Antibodies to Infectious Bronchitis in Commercial and Indigenous Nigerian Chickens Using ELISA. Bull. Anim. Health Prod. Afr. 1988, 3, 259–262. [Google Scholar]
- Komolafe, O.; Ozeigbe, P.; Anene, B. A Survey of Avian Infectious Bronchitis Antibodies in Nsukka, Nigeria. Bull. Anim. Health Prod. Afr. 1990, 38, 471–472. [Google Scholar]
- Owoade, A.A.; Ducatez, M.F.; Muller, C.P. Seroprevalence of Avian Influenza Virus, Infectious Bronchitis Virus, Reovirus, Avian Pneumovirus, Infectious Laryngotracheitis Virus, and Avian Leukosis Virus in Nigerian Poultry. Avian Dis. 2006, 50, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Emikpe, B.O.; Ohore, O.G.; Olujonwo, M.; Akpavie, S.O. Prevalence of Antibodies to Infectious Bronchitis Virus (IBV) in Chickens in Southwestern Nigeria. Afr. J. Microbiol. Res. 2010, 4, 92–95. [Google Scholar]
- Musa, W.I.; Abdullahi, B.Y.; Lawal, S.; Bello, M.; Abdu, P.A. Detection of Antibodies to Avian Influenza, Infectious Bronchitis and Newcastle Disease Viruses in Wild Birds in Three States of Nigeria. Sokoto J. Vet. Sci. 2017, 15, 28–35. [Google Scholar] [CrossRef][Green Version]
- Ayim-Akonor, M.; Owusu-Ntumy, D.D.; Ohene-Asa, H.E.; Oduro-Abrokwa, A.; Hammond, P.; Appenteng, M.; Annan, D. Serological and Molecular Surveillance of Infectious Bronchitis Virus Infection in Free-Range Chickens and Guinea Fowls in the Ga-East District of Ghana. J. Vet. Med. 2018, 2018, e4949580. [Google Scholar] [CrossRef]
- Daodu, O.B.; Jokotola, P.T.; Omowon, A.A.; Olorunshola, I.D.; Ahmed, O.A.; Raufu, I.A.; Raji, M.A.; Daodu, O.C. Cross-Species Surveillance and Risk Factors Associated with Avian Coronavirus in North-Central and South West Regions of Nigeria. Trop. Biomed. 2021, 38, 28–32. [Google Scholar] [CrossRef]
- Tarnagda, Z.; Yougbare, I.; Kam, A.; Tahita, M.C.; Ouedraogo, J.B. Prevalence of Infectious Bronchitis and Newcastle Disease Virus among Domestic and Wild Birds in H5N1 Outbreaks Areas. J. Infect. Dev. Ctries. 2011, 5, 565–570. [Google Scholar] [CrossRef]
- Ayim-Akonor, M.; Baryeh, K.; Asante, I.A. Molecular Based Survey of Pathogens Associated with Respiratory Disease Outbreaks in Broiler Chickens in Accra. J. Nat. Sci. Res. 2013, 3, 25. [Google Scholar]
- Ayim-Akonor, M.; Cornelius, A.; Ohene, A.; Kwaku, B. Infectious Bronchitis Virus: A Major Cause of Respiratory Disease Outbreaks in Chickens in Ghana. J. Biol. Agric. Healthc. 2013, 3, 56. [Google Scholar]
- Ducatez, M.F.; Martin, A.M.; Owoade, A.A.; Olatoye, I.O.; Alkali, B.R.; Maikano, I.; Snoeck, C.J.; Sausy, A.; Cordioli, P.; Muller, C.P. Characterization of a New Genotype and Serotype of Infectious Bronchitis Virus in Western Africa. J. Gen. Virol. 2009, 90, 2679–2685. [Google Scholar] [CrossRef] [PubMed]
- Ganapathy, K.; Ball, C.; Forrester, A. Genotypes of Infectious Bronchitis Viruses Circulating in the Middle East between 2009 and 2014. Virus Res. 2015, 210, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Domanska-Blicharz, K.; Lisowska, A.; Sajewicz-Krukowska, J. Molecular Epidemiology of Infectious Bronchitis Virus in Poland from 1980 to 2017. Infect. Genet. Evol. 2020, 80, 104177. [Google Scholar] [CrossRef] [PubMed]
- Bányai, K.; Palya, V.; Dénes, B.; Glávits, R.; Ivanics, É.; Horváth, B.; Farkas, S.L.; Marton, S.; Bálint, Á.; Gyuranecz, M.; et al. Unique Genomic Organization of a Novel Avipoxvirus Detected in Turkey (Meleagris Gallopavo). Infect. Genet. Evol. 2015, 35, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Bali, K.; Bálint, Á.; Farsang, A.; Marton, S.; Nagy, B.; Kaszab, E.; Belák, S.; Palya, V.; Bányai, K. Recombination Events Shape the Genomic Evolution of Infectious Bronchitis Virus in Europe. Viruses 2021, 13, 535. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and Analysis of Recombination Patterns in Virus Genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-Length Human Immunodeficiency Virus Type 1 Genomes from Subtype C-Infected Seroconverters in India, with Evidence of Intersubtype Recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef]
- Casais, R.; Dove, B.; Cavanagh, D.; Britton, P. Recombinant Avian Infectious Bronchitis Virus Expressing a Heterologous Spike Gene Demonstrates That the Spike Protein Is a Determinant of Cell Tropism. J. Virol. 2003, 77, 9084–9089. [Google Scholar] [CrossRef]
- Cavanagh, D. Coronaviruses in Poultry and Other Birds. Avian Pathol. 2005, 34, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Coronavirus Avian Infectious Bronchitis Virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.; Madbouly, H.; Gelb, J.; Ladman, B. Isolation and Identification of Egypt/Beni-Suef/01 a Novel Genotype of Infectious Bronchitis Virus. Vet. Med. J. Giza. 2002, 50, 1065–1078. [Google Scholar]
- Jones, R.C.; Savage, C.E.; Naylor, C.J.; Cook, J.K.A.; El-Houadfi, M. A Possible North African Progenitor of the Major European Infectious Bronchitis Virus Variant (793B, 4/91, CR88, Etc.). In Proceedings of the IV International Symposium on Avian Corona- and Pneumovirus Infections, Rauischholzhausen, Germany, 20–23 June 2004; pp. 105–111. [Google Scholar]
- Abdel-Moneim, A.S.; El-Kady, M.F.; Ladman, B.S.; Gelb, J. S1 Gene Sequence Analysis of a Nephropathogenic Strain of Avian Infectious Bronchitis Virus in Egypt. Virol. J. 2006, 3, 78. [Google Scholar] [CrossRef]
- Bourogâa, H.; Miled, K.; Gribâa, L.; El Behi, I.; Ghram, A. Characterization of New Variants of Avian Infectious Bronchitis Virus in Tunisia. Avian Dis. 2009, 53, 426–433. [Google Scholar] [CrossRef]
- Toffan, A.; Monne, I.; Terregino, C.; Cattoli, G.; Hodobo, C.T.; Gadaga, B.; Makaya, P.V.; Mdlongwa, E.; Swiswa, S. QX-like Infectious Bronchitis Virus in Africa. Vet. Rec. 2011, 169, 589. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.S.; Afifi, M.A.; El-Kady, M.F. Emergence of a Novel Genotype of Avian Infectious Bronchitis Virus in Egypt. Arch. Virol. 2012, 157, 2453–2457. [Google Scholar] [CrossRef]
- Fellahi, S.; Ducatez, M.; El Harrak, M.; Guérin, J.-L.; Touil, N.; Sebbar, G.; Bouaiti, E.A.; Khataby, K.; Ennaji, M.M.; El-Houadfi, M. Prevalence and Molecular Characterization of Avian Infectious Bronchitis Virus in Poultry Flocks in Morocco from 2010 to 2014 and First Detection of Italy 02 in Africa. Avian Pathol. 2015, 44, 287–295. [Google Scholar] [CrossRef]
- Fellahi, S.; El Harrak, M.; Khayi, S.; Guerin, J.-L.; Kuhn, J.H.; El Houadfi, M.; Ennaji, M.M.; Ducatez, M. Phylogenetic Analysis of Avian Infectious Bronchitis Virus Isolates from Morocco: A Retrospective Study (1983 to 2014). Virol. Sin. 2017, 32, 155–158. [Google Scholar] [CrossRef]
- Awad, F.; Baylis, M.; Ganapathy, K. Detection of Variant Infectious Bronchitis Viruses in Broiler Flocks in Libya. Int. J. Vet. Sci. Med. 2014, 2, 78–82. [Google Scholar] [CrossRef]
- Sid, H.; Benachour, K.; Rautenschlein, S. Co-Infection with Multiple Respiratory Pathogens Contributes to Increased Mortality Rates in Algerian Poultry Flocks. Avian Dis. 2015, 59, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Hutton, S.; Bettridge, J.; Christley, R.; Habte, T.; Ganapathy, K. Detection of Infectious Bronchitis Virus 793B, Avian Metapneumovirus, Mycoplasma Gallisepticum and Mycoplasma Synoviae in Poultry in Ethiopia. Trop. Anim. Health Prod. 2017, 49, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Lounas, A.; Oumouna-Benachour, K.; Medkour, H.; Oumouna, M. The First Evidence of a New Genotype of Nephropathogenic Infectious Bronchitis Virus Circulating in Vaccinated and Unvaccinated Broiler Flocks in Algeria. Vet. World 2018, 11, 1630–1636. [Google Scholar] [CrossRef] [PubMed]
- Awuni, J.A.; Bianco, A.; Dogbey, O.J.; Fusaro, A.; Yingar, D.T.; Salviato, A.; Ababio, P.T.; Milani, A.; Bonfante, F.; Monne, I. Avian Influenza H9N2 Subtype in Ghana: Virus Characterization and Evidence of Co-Infection. Avian Pathol. 2019, 48, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Knoetze, A.D.; Moodley, N.; Abolnik, C. Two Genotypes of Infectious Bronchitis Virus Are Responsible for Serological Variation in KwaZulu-Natal Poultry Flocks Prior to 2012. Onderstepoort J. Vet. Res. 2014, 81, 1–10. [Google Scholar] [CrossRef]
- Jackwood, M.W.; Hilt, D.A.; Callison, S.A.; Lee, C.W.; Plaza, H.; Wade, E. Spike Glycoprotein Cleavage Recognition Site Analysis of Infectious Bronchitis Virus. Avian Dis. 2001, 45, 366–372. [Google Scholar] [CrossRef]
- Cavanagh, D. The Coronavirus Surface Glycoprotein. In The Coronaviridae; Siddell, S.G., Ed.; The Viruses; Springer US: Boston, MA, USA, 1995; pp. 73–113. ISBN 978-1-4899-1531-3. [Google Scholar]
- Abozeid, H.H.; Paldurai, A.; Khattar, S.K.; Afifi, M.A.; El-Kady, M.F.; El-Deeb, A.H.; Samal, S.K. Complete Genome Sequences of Two Avian Infectious Bronchitis Viruses Isolated in Egypt: Evidence for Genetic Drift and Genetic Recombination in the Circulating Viruses. Infect. Genet. Evol. 2017, 53, 7–14. [Google Scholar] [CrossRef]
- Mousavi, F.S.; Ghalyanchilangeroudi, A.; Hosseini, H.; Nayeri Fasaei, B.; Ghafouri, S.A.; Abdollahi, H.; Fallah-Mehrabadi, M.H.; Sadri, N. Complete Genome Analysis of Iranian IS-1494 like Avian Infectious Bronchitis Virus. Virusdisease 2018, 29, 390–394. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Q.; Zhang, Z. Global Diversification and Distribution of Coronaviruses with Furin Cleavage Sites. Front. Microbiol. 2021, 12, 649314. [Google Scholar] [CrossRef]
- Franzo, G.; Listorti, V.; Naylor, C.J.; Lupini, C.; Laconi, A.; Felice, V.; Drigo, M.; Catelli, E.; Cecchinato, M. Molecular Investigation of a Full-Length Genome of a Q1-like IBV Strain Isolated in Italy in 2013. Virus Res. 2015, 210, 77–80. [Google Scholar] [CrossRef]
- Marandino, A.; Tomás, G.; Panzera, Y.; Greif, G.; Parodi-Talice, A.; Hernández, M.; Techera, C.; Hernández, D.; Pérez, R. Whole-Genome Characterization of Uruguayan Strains of Avian Infectious Bronchitis Virus Reveals Extensive Recombination between the Two Major South American Lineages. Infect. Genet. Evol. 2017, 54, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Xue, Y.; Wang, F.; Chen, F.; Shu, D.; Xie, Q. Analysis of S1 Gene of Avian Infectious Bronchitis Virus Isolated in Southern China during 2011–2012. Virus Genes 2014, 49, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Capua, I.; Minta, Z.; Karpinska, E.; Mawditt, K.; Britton, P.; Cavanagh, D.; Gough, R.E. Co-Circulation of Four Types of Infectious Bronchitis Virus (793/B, 624/I, B1648 and Massachusetts). Avian Pathol. 1999, 28, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Ababneh, M.; Dalab, A.E.; Alsaad, S.; Al-Zghoul, M. Presence of Infectious Bronchitis Virus Strain CK/CH/LDL/97I in the Middle East. ISRN Vet. Sci. 2012, 2012, 201721. [Google Scholar] [CrossRef] [PubMed]
- Toffan, A.; Bonci, M.; Bano, L.; Bano, L.; Valastro, V.; Vascellari, M.; Capua, I.; Terregino, C. Diagnostic and Clinical Observation on the Infectious Bronchitis Virus Strain Q1 in Italy. Vet. Ital. 2013, 49, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Sabra, M.; Abdellatif, W.; Ahmed, A.; Osman, N. Molecular Characterization and Phylogenetic Analysis of Full-Length S1 Gene of GI-16 and GI-23 Infectious Bronchitis Virus in Qena, Egypt. J. World Poult. Res. 2020, 10, 71–80. [Google Scholar] [CrossRef]
- Liu, S.; Kong, X. A New Genotype of Nephropathogenic Infectious Bronchitis Virus Circulating in Vaccinated and Non-Vaccinated Flocks in China. Avian Pathol. 2004, 33, 321–327. [Google Scholar] [CrossRef]
- Terregino, C.; Toffan, A.; Beato, M.S.; De Nardi, R.; Vascellari, M.; Meini, A.; Ortali, G.; Mancin, M.; Capua, I. Pathogenicity of a QX Strain of Infectious Bronchitis Virus in Specific Pathogen Free and Commercial Broiler Chickens, and Evaluation of Protection Induced by a Vaccination Programme Based on the Ma5 and 4/91 Serotypes. Avian Pathol. 2008, 37, 487–493. [Google Scholar] [CrossRef]
- Benyeda, Z.; Mató, T.; Süveges, T.; Szabó, E.; Kardi, V.; Abonyi-Tóth, Z.; Rusvai, M.; Palya, V. Comparison of the Pathogenicity of QX-like, M41 and 793/B Infectious Bronchitis Strains from Different Pathological Conditions. Avian Pathol. 2009, 38, 449–456. [Google Scholar] [CrossRef]
- Khanh, N.P.; Tan, S.W.; Yeap, S.K.; Lee, H.J.; Choi, K.S.; Hair-Bejo, M.; Bich, T.N.; Omar, A.R. Comparative Pathogenicity of Malaysian QX-like and Variant Infectious Bronchitis Virus Strains in Chickens at Different Age of Exposure to the Viruses. J. Comp. Pathol. 2018, 161, 43–54. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Age (d) | Organ of Isolation | Country of Origin | Clinical Features/Pathology | Vaccination History against IBV | Genotype of Study Strain |
|---|---|---|---|---|---|---|
| D2326/3/13/CM/2013 | 42 | I | Cameroon | respiratory signs, diarrhea | not vaccinated | GI-14 (B1648-like) |
| D2326/4/13/CM/2013 | 60 | I | Cameroon | respiratory signs: groans, snoring, mucoid nasal discharge | not vaccinated | GI-14 (B1648-like) |
| D2328/15/3/13/GH/2013 | 27 | CT | Ghana | respiratory signs: hard breathing, groans, nasal discharges, digestive signs: green feces | not vaccinated | GI-19 (QX-like) |
| D2334/11/2/13/CI/2013 | 31 | CT | Ivory Coast | nasal discharge, groans, diarrhea, growth problem | D1: H120 | unique variant (GI-31) |
| D2334/12/2/13/CI/2013 | 45 | CT | Ivory Coast | respiratory signs, diarrhea | D1: H120, D28: Mass | GI-16 (Q1-like) |
| Study Strains | All Read Output | Assembled Reads | Assembled Nucleotides | Mean Sequencing Depth (X) | Genome Size * |
|---|---|---|---|---|---|
| D2326/3/13/CM/2013 | 40,932 | 33,342 | 5,960,515 | 205 | 27,641 |
| D2326/4/13/CM/2013 | 40,949 | 31,821 | 5,818,590 | 180 | 27,671 |
| D2328/15/3/13/GH/2013 | 77,472 | 62,435 | 12,043,754 | 391 | 27,616 |
| D2334/11/2/13/CI/2013 | 97,955 | 80,170 | 17,037,995 | 559 | 27,640 |
| D2334/12/2/13/CI/2013 | 74,763 | 56,316 | 11,871,797 | 456 | 27,646 |
| ORF Length (nt) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Study Strains | 1a | 1ab | Spike | 3a | 3b | Envelope | Membrane | 4b | 4c | 5a | 5b | Nucleocapsid | 6b |
| D2326/3/13/CM/2013 | 11,841 | 19,875 | 3498 | 174 | 186 | 324 | 672 | 285 | 171 | 198 | 249 | 1230 | 225 |
| D2326/4/13/CM/2013 | 11,862 | 19,896 | 3501 | 174 | 192 | 324 | 672 | 285 | 171 | 198 | 249 | 1230 | 225 |
| D2328/15/3/13/GH/2013 | 11,829 | 19,863 | 3498 | 174 | 192 | 285 | 678 | 285 | 171 | 198 | 249 | 1224 | 225 |
| D2334/11/2/13/CI/2013 | 11,850 | 19,884 | 3507 | 174 | 195 | 282 | 672 | 201 | 171 | 198 | 249 | 1230 | 222 |
| D2334/12/2/13/CI/2013 | 11,862 | 19,896 | 3501 | 174 | 195 | 282 | 672 | 285 | 171 | 198 | 249 | 1230 | 222 |
| D2334/11/2/13/CI | D2334/12/2/13/CI | D2326/4/13/CM | D2326/3/13/CM | D2328/15/3/13/GH | |
|---|---|---|---|---|---|
| D2334/11/2/13/CI | |||||
| D2334/12/2/13/CI | 94.58 | ||||
| D2326/4/13/CM | 91.42 | 91.62 | |||
| D2326/3/13/CM | 91.35 | 91.55 | 99.37 | ||
| D2328/15/3/13/GH | 90.23 | 90.05 | 89.15 | 89.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bali, K.; Kaszab, E.; Marton, S.; Hamdiou, S.H.; Bentaleb, R.K.; Kiss, I.; Palya, V.; Bányai, K. Novel Lineage of Infectious Bronchitis Virus from Sub-Saharan Africa Identified by Random Amplification and Next-Generation Sequencing of Viral Genome. Life 2022, 12, 475. https://doi.org/10.3390/life12040475
Bali K, Kaszab E, Marton S, Hamdiou SH, Bentaleb RK, Kiss I, Palya V, Bányai K. Novel Lineage of Infectious Bronchitis Virus from Sub-Saharan Africa Identified by Random Amplification and Next-Generation Sequencing of Viral Genome. Life. 2022; 12(4):475. https://doi.org/10.3390/life12040475
Chicago/Turabian StyleBali, Krisztina, Eszter Kaszab, Szilvia Marton, Seydou Hamadou Hamdiou, Reza Karim Bentaleb, István Kiss, Vilmos Palya, and Krisztián Bányai. 2022. "Novel Lineage of Infectious Bronchitis Virus from Sub-Saharan Africa Identified by Random Amplification and Next-Generation Sequencing of Viral Genome" Life 12, no. 4: 475. https://doi.org/10.3390/life12040475
APA StyleBali, K., Kaszab, E., Marton, S., Hamdiou, S. H., Bentaleb, R. K., Kiss, I., Palya, V., & Bányai, K. (2022). Novel Lineage of Infectious Bronchitis Virus from Sub-Saharan Africa Identified by Random Amplification and Next-Generation Sequencing of Viral Genome. Life, 12(4), 475. https://doi.org/10.3390/life12040475