
The Role of Photo-Cycles in the Modulation of Growth and Biochemical Profile of Microalgae: Part I—Food Interest Compounds


 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae and Culture Media
2.2. Photobioreactor Design
2.3. Obtaining the Kinetic Data in an Experimental Photobioreactor
2.4. Total Lipids and Fatty Acids Profile
2.5. Protein Amino Acids Profile
2.6. Sterols Profile
2.7. Statistical Analysis
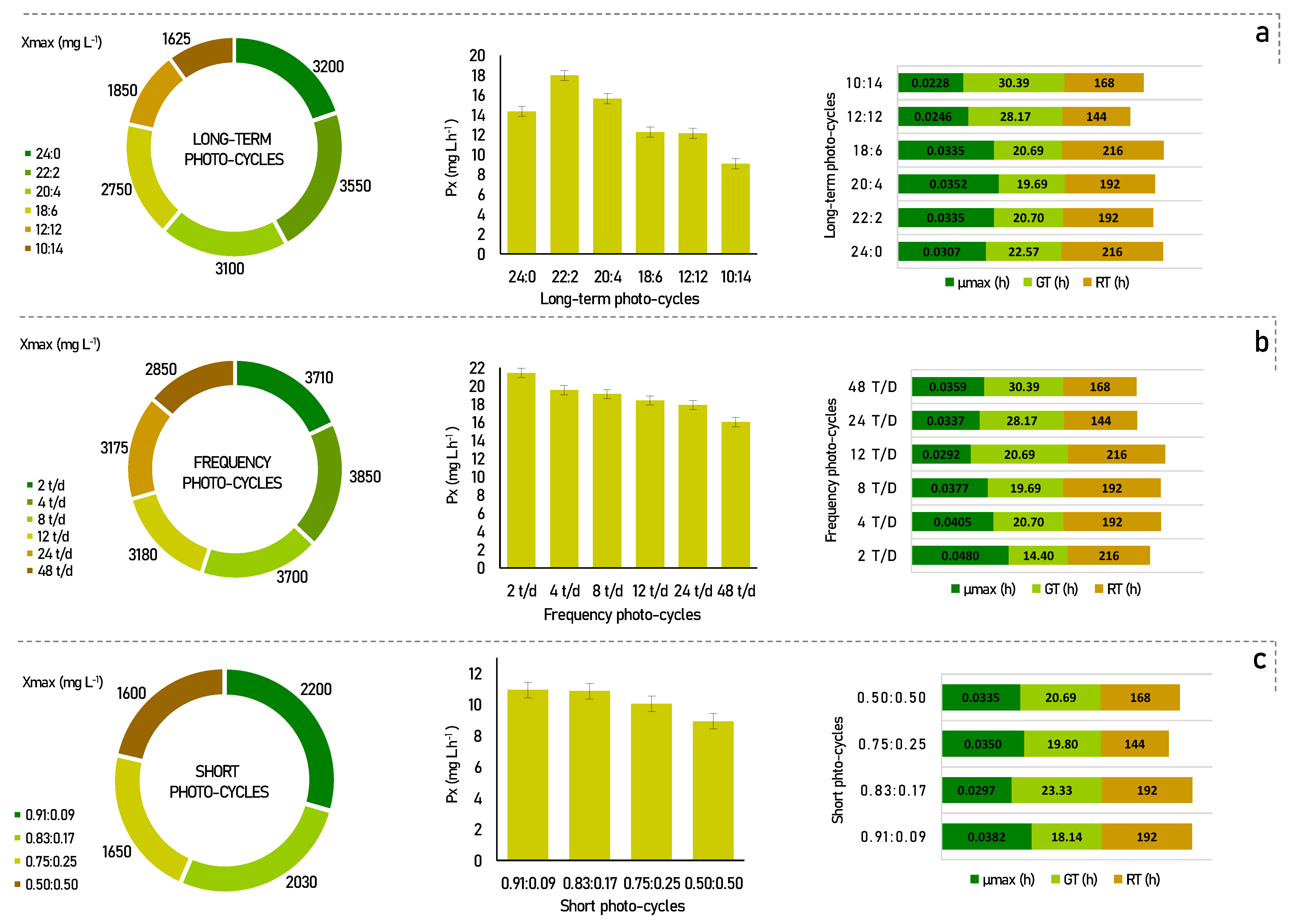
3. Results and Discussion
3.1. Analysis of the Biochemical Profile
3.1.1. Total Lipids and Fatty Acids
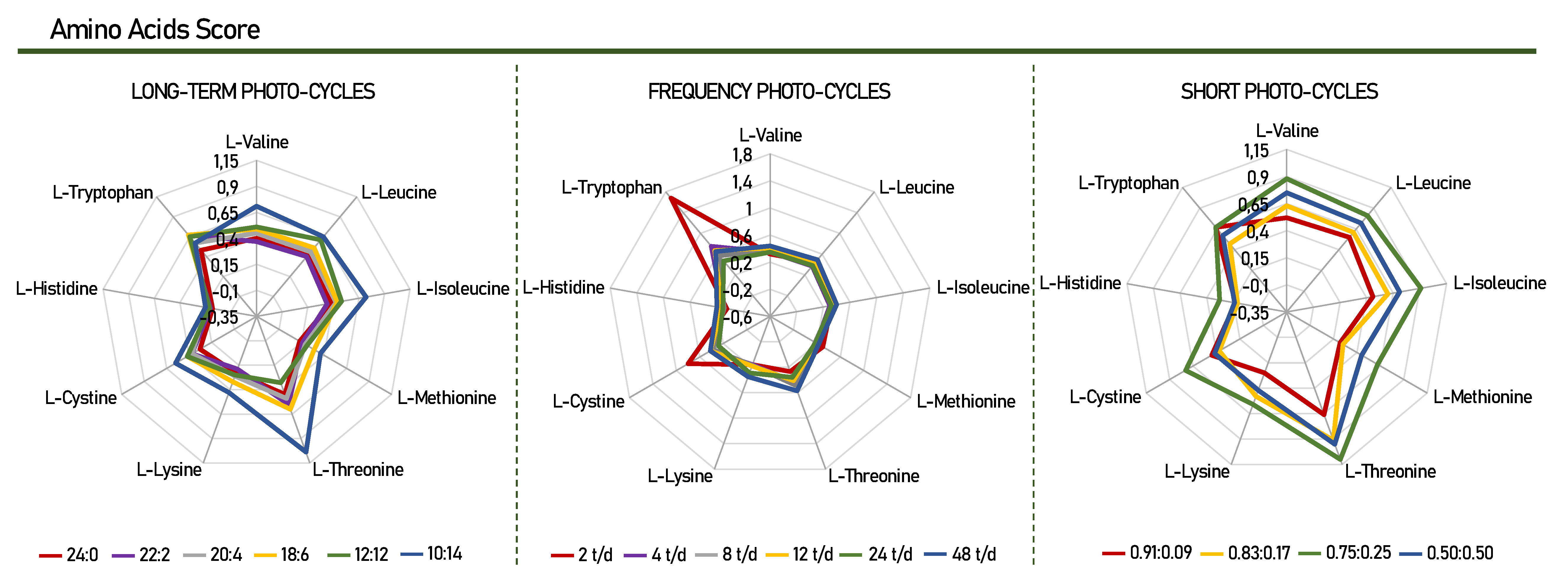
3.1.2. Protein Amino Acid
3.1.3. Total Sterols
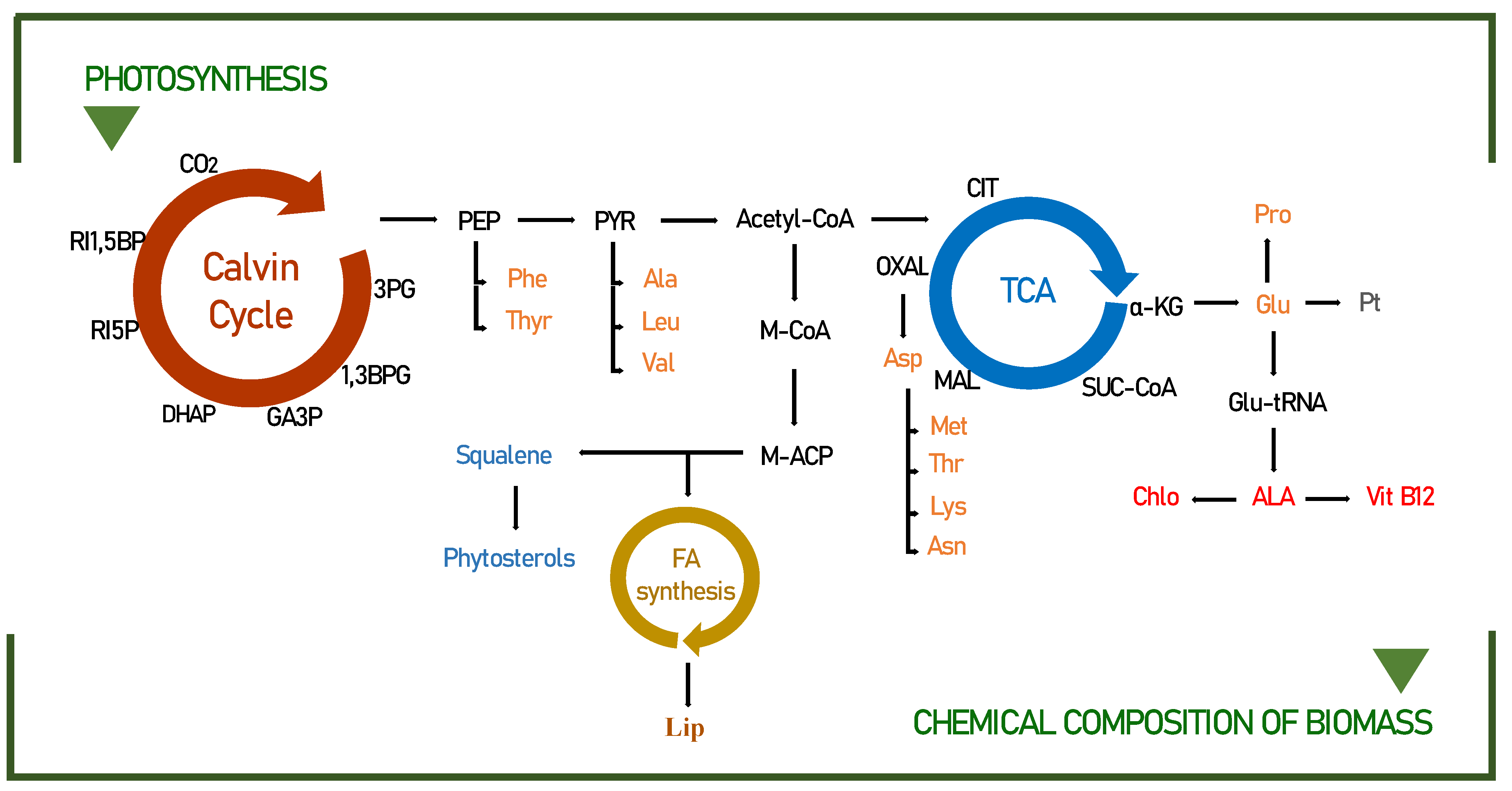
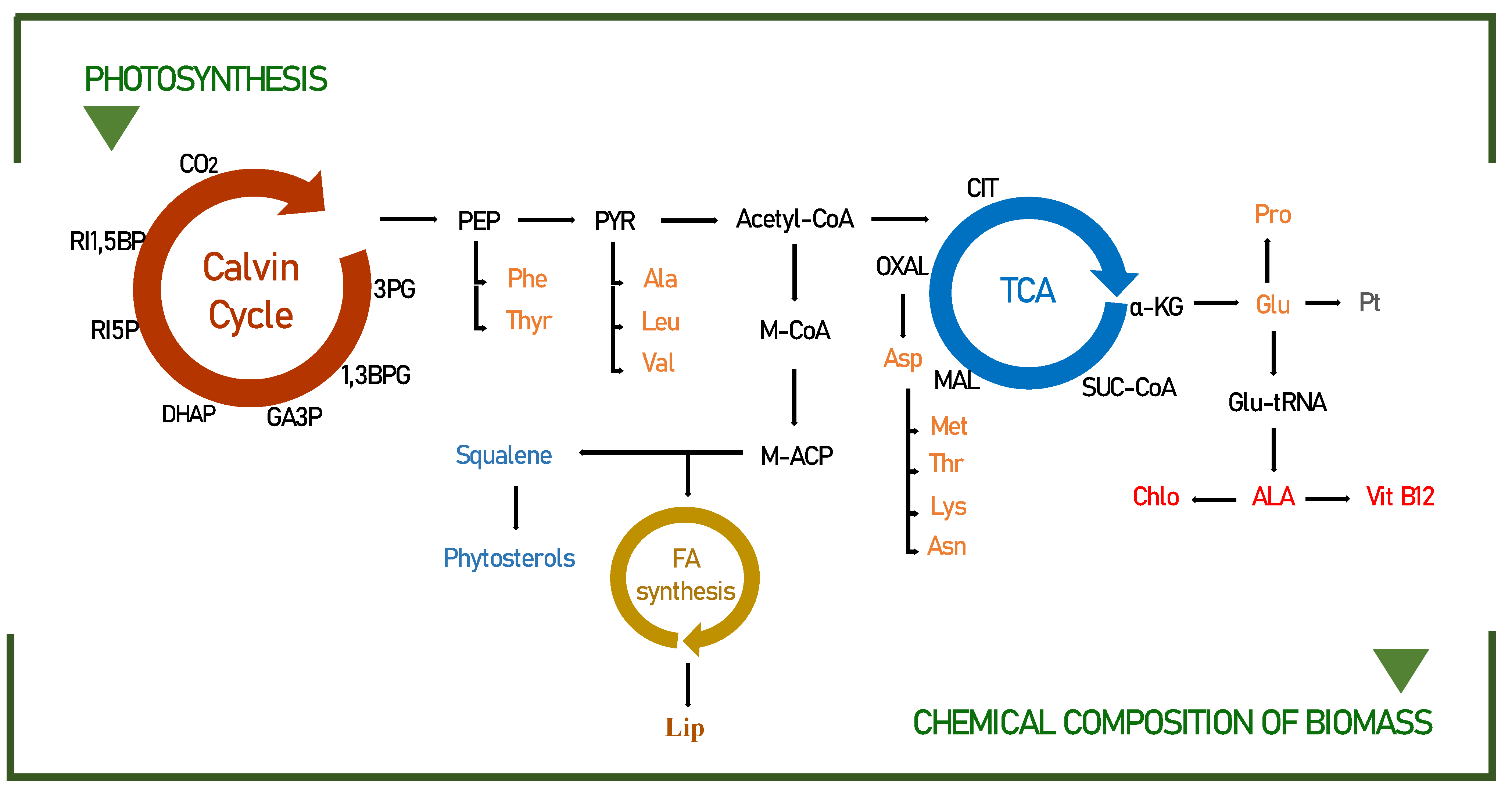
3.2. Biochemical Pathways of Food Interest Compounds
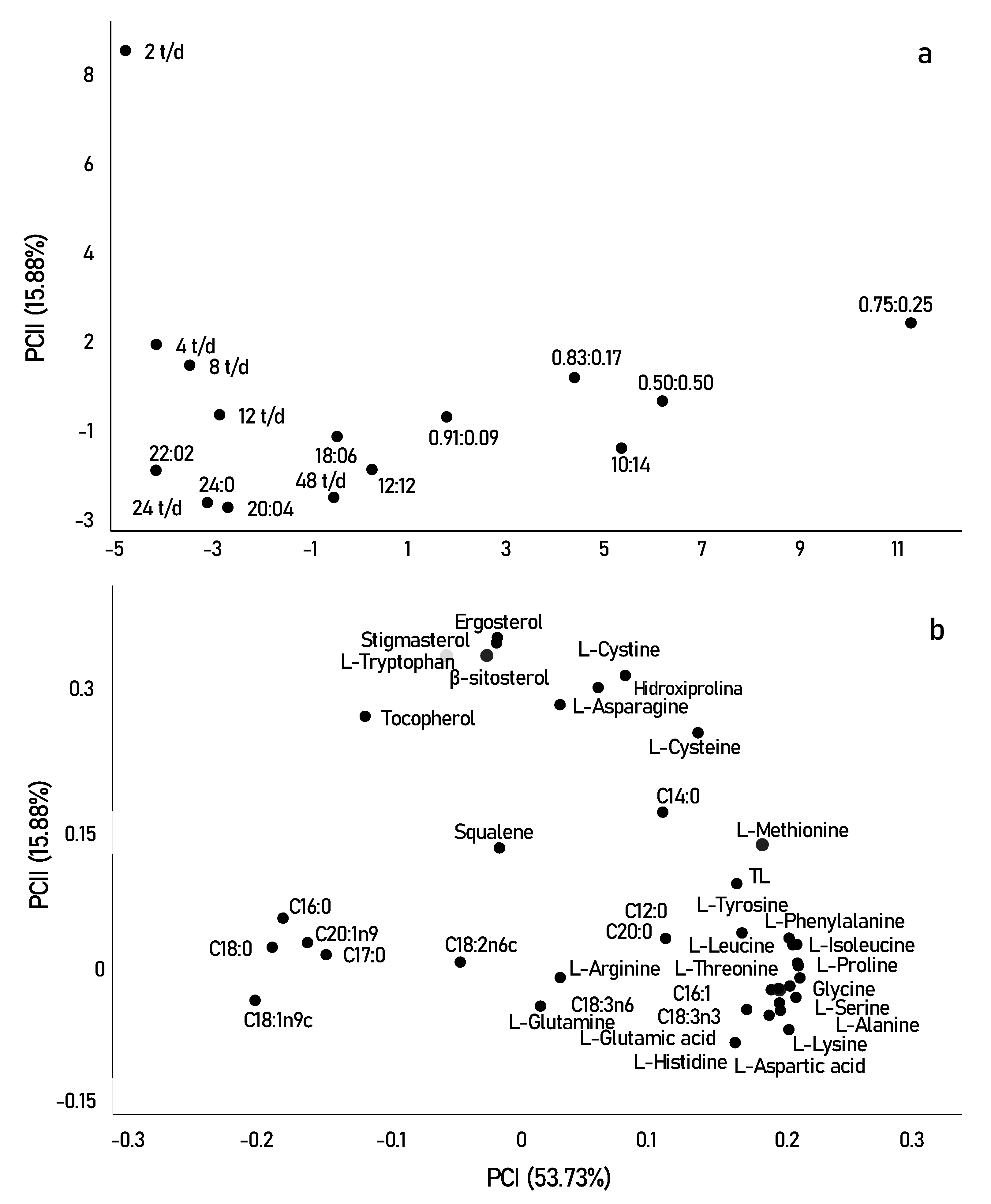
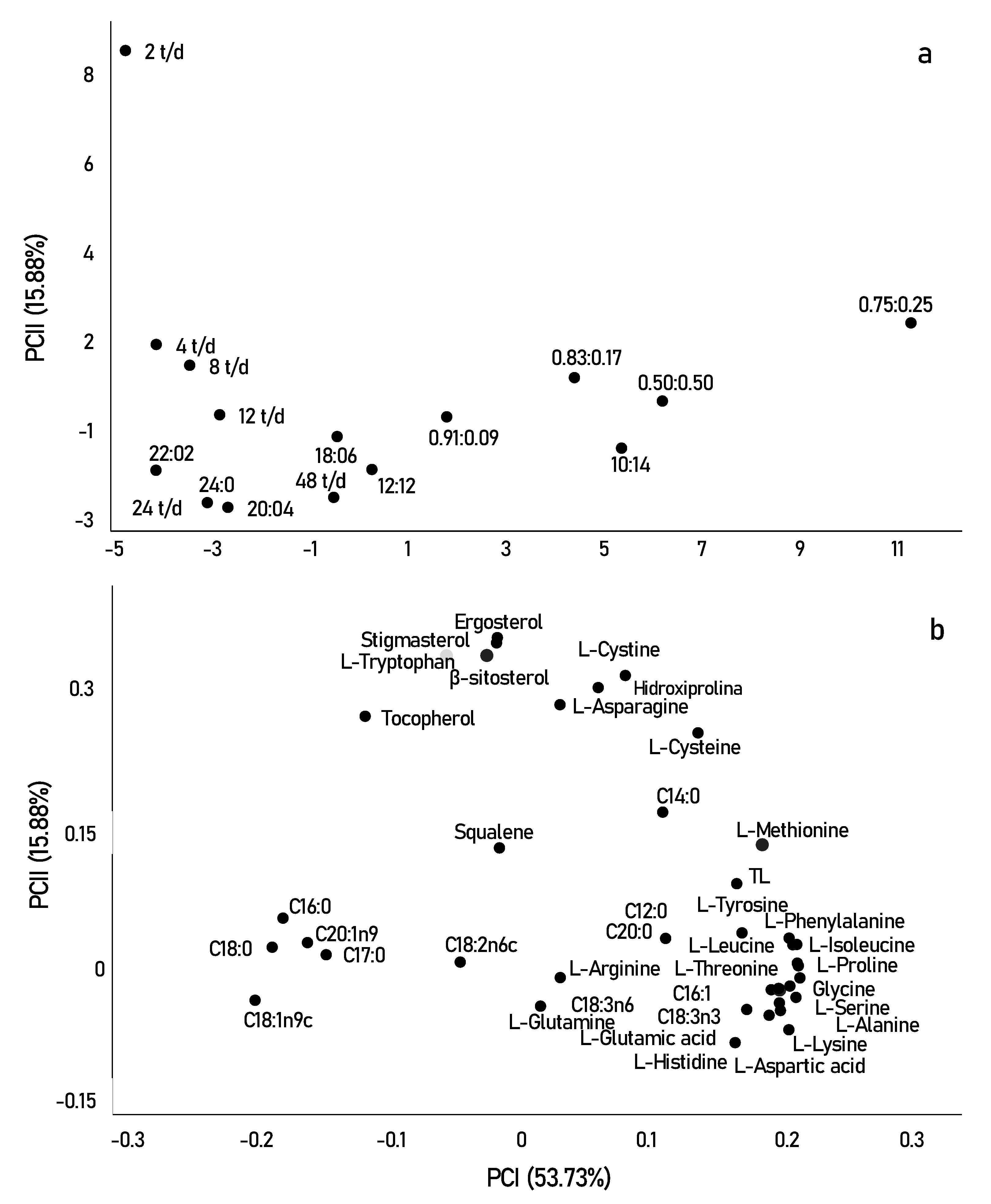
3.3. Exploratory Multivariate Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Fernandes, A.S.; Nascimento, T.C.; Pinheiro, P.N.; De Rosso, V.V.; De Menezes, C.R.; Jacob-Lopes, E.; Zepka, L.Q. Insights on the intestinal absorption of chlorophyll series from microalgae. Food Res. Int. 2021, 140, 110031. [Google Scholar] [CrossRef] [PubMed]
- Pugazhendhi, A.; Nagappan, S.; Bhosale, R.R.; Tsai, P.S.; Natarajan, S.; Devendran, S.; Al-Haj, L.; Ponnusamy, K.; Kumar, G. Various potential techniques to reduce the water footprint of microalgal biomass production for biofuel—A review. Sci. Total Environ. 2020, 749, 142218. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, S.F.; Maroneze, M.M.; Dias, R.R.; Vendruscolo, R.G.; Wagner, R.; De Menezes, C.R.; Zepka, L.Q.; Jacob-Lopes, E. Mapping the performance of photobioreactors for microalgae cultivation: Geographic position and local climate. J. Chem. Technol. Biotechnol. 2020, 95, 2411–2420. [Google Scholar] [CrossRef]
- Mutanda, T.; Naidoo, D.; Bwapwa, J.K.; Anandraj, A. Biotechnological Applications of Microalgal Oleaginous Compounds: Current Trends on Microalgal Bioprocessing of Products. Front. Energy Res. 2020, 8, 598803. [Google Scholar] [CrossRef]
- Pôjo, V.; Tavares, T.; Malcata, F.X. Processing Methodologies of Wet Microalga Biomass toward Oil Separation: An Overview. Molecules 2021, 26, 641. [Google Scholar] [CrossRef]
- Maltsev, Y.; Maltseva, K.; Kulikovskiy, M.; Maltseva, S. Influence of Light Conditions on Microalgae Growth and Content of Lipids, Carotenoids, and Fatty Acid Composition. Biology 2021, 10, 1060. [Google Scholar] [CrossRef]
- Lisondro, I.; Serrano, C.G.; Sepúlveda, C.; Ceballos, A.I.B.; Acién-Fernández, F.G. Infuence of irradiance on the growth and biochemical composition of Nitzschia aff. pelúcida. J. Appl. Phycol. 2022, 34, 19–30. [Google Scholar] [CrossRef]
- Telfer, A. What is β-carotene doing in the photosystem II reaction centre? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2002, 357, 1431–1439. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Thomas-Hall, S.R.; Chua, E.T.; Eltanahy, E.; Netzel, M.E.; Netzel, G.; Lu, Y.; Schenk, P.M. LED power efficiency of biomass, fatty acid, and carotenoid production in Nannochloropsis microalgae. Bioresour. Technol. 2018, 252, 118–126. [Google Scholar] [CrossRef]
- Cezare-Gomes, E.A.; Silva, L.C.M.; Pérez-Mora, L.S.; Matsudo, M.C.; Camargo, L.S.F.; Singh, A.K.; Carvalho, J.C.M. Potential of Microalgae Carotenoids for Industrial Application. Appl. Biochem. Biotechnol. 2019, 188, 602–634. [Google Scholar] [CrossRef]
- Dias, R.R.; Lasta, P.; Vendruscolo, R.G.; Wagner, R.; Zepka, L.Q.; Jacob-Lopes, E. Mapping the Performance of Photobioreactors for Microalgae Cultivation. Part II: Equatorial and Tropical Climate Zone. J. Chem. Technol. Biotechnol. 2020, 96, 613–621. [Google Scholar] [CrossRef]
- Deprá, M.C.; Dias, R.R.; Severo, I.A.; De Menezes, C.R.; Zepka, L.Q.; Jacob-Lopes, E. Carbon dioxide capture and use in photobioreactors: The role of the carbon dioxide loads in the carbon footprint. Bioresour. Technol. 2020, 314, 123745. [Google Scholar] [CrossRef] [PubMed]
- Jacob-Lopes, E.; Lacerda, L.M.C.F.; Scoparo, C.H.G.; Franco, T.T. Effect of light cycles (night/day) on CO2 fixation and biomass production by microalgae in photobioreactors. Chem. Eng. Process. 2009, 48, 306–310. [Google Scholar] [CrossRef]
- Liu, S.; Daiggerd, G.T.; Kanga, J.; Zhange, G. Effects of light intensity and photoperiod on pigments production and corresponding key gene expression of Rhodopseudomonas palustris in a photobioreactor system. Bioresour. Technol. 2019, 294, 122172. [Google Scholar] [CrossRef]
- Chiarini, A.; Quadrio, M. The light/dark cycle of microalgae in a thin-layer photobioreactor. J. Appl. Phycol. 2021, 33, 183–195. [Google Scholar] [CrossRef]
- Urbina-Suarez, N.A.; Ayala-González, D.D.; Rivera-Amaya, J.D.; Barajas-Solano, A.F.; Machuca-Martínez, A.F. Evaluation of the Light/Dark Cycle and Concentration of Tannery Wastewater in the Production of Biomass and Metabolites of Industrial Interest from Microalgae and Cyanobacteria. Water 2022, 14, 346. [Google Scholar] [CrossRef]
- Rippka, R.; Derulles, J.; Watrerbury, J.B.; Herdman, M.; STainer, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Vendruscolo, R.G.; Facchi, M.M.X.; Maroneze, M.M.; Fagundes, M.B.; Cichoski, A.J.; Zepka, L.Q.; Barin, J.S.; Jacob–Lopes, E.; Wagner, R. Polar and non–polar intracellular compounds from microalgae: Methods of simultaneous extraction, gas chromatography determination and comparative analysis. Food Res. Int. 2018, 109, 204–212. [Google Scholar] [CrossRef]
- Visentainer, J.V. Aspectos analíticos da resposta do detector de ionização em chama para ésteres de ácidos graxos em biodiesel e alimentos. Quim. Nova 2012, 19, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Do Nascimento, T.C.; Cazarin, C.B.; Maróstica, M.R., Jr.; Risso, É.M.; Amaya-Farfan, J.; Grimaldi, R.; Mercadante, A.Z.; Jacob-Lopes, E.; Zepka, L.Q. Microalgae biomass intake positively modulates serum lipid profile and antioxidant status. J. Funct. Foods 2019, 58, 11–20. [Google Scholar] [CrossRef]
- Fagundes, M.B.; Falk, R.B.; Facchi, M.M.X.; Vendruscolo, R.G.; Maroneze, M.M.; Zepka, L.Q.; Jacob-Lopes, E.; Wagner, R. Insights in cyanobacteria lipidomics: A sterols characterization from Phormidium autumnale biomass in heterotrophic cultivation. Food Res. Int. 2019, 119, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Achyuthan, K.; Harper, J.; ManginelL, R.; Moorman, M. Volatile Metabolites Emission by In Vivo Microalgae—An Overlooked Opportunity? Metabolites 2017, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Simionato, D.; Sforza, E.; Carpinelli, E.C.; Bertucco, A.; Giacometti, G.M.; Morosinotto, T. Acclimation of Nannochloropsis gaditana to different illumination regimes: Effects on lipids accumulation. Bioresour. Technol. 2011, 102, 6026–6032. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, P.; Zhang, G. Biomass and carotenoid production in photosynthetic bacteria wastewater treatment: Effects of light intensity. Bioresour. Technol. 2014, 171, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Jalalizadeh, M.; Zhang, Q. Growth kinetic models for microalgae cultivation: A review. Algal Res. 2015, 12, 497–512. [Google Scholar] [CrossRef]
- González-Camejo, J.; Viruela, A.; Ruano, M.V.; Barat, R.; Seco, A.; Ferrer, J. Effect of light intensity, light duration and photoperiods in the performance of an outdoor photobioreactor for urban wastewater treatment. Algal Res. 2019, 40, 101511. [Google Scholar] [CrossRef]
- Murata, N.; Takahashi, S.; Nishiyama, Y.; Allakhverdiev, S.I. Photoinhibition of photosystem II under environmental stress. Biochim. Biophys. Acta Bioenerg. 2007, 1767, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Liao, Q.; Li, L.; Chen, R.; Zhu, X. A novel photobioreactor generating the light/dark cycle to improve microalgae cultivation. Bioresour. Technol. 2014, 161, 186–191. [Google Scholar] [CrossRef]
- Jia, T.; Ito, H.; Tanaka, A. Simultaneous regulation of antenna size and photosystem I/II stoichiometry in Arabidopsis thaliana. Planta 2016, 244, 1041–1053. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Fan, J.; Xu, C.C. Analysis of Oil Droplets in Microalgae. Methods Cell Biol. 2013, 116, 71–82. [Google Scholar] [CrossRef]
- Morales, M.; Aflalo, C.; Bernard, O. Microalgal lipids: A review of lipids potential and quantification for 95 phytoplankton species. Biomass Bioenergy 2021, 150, 106108. [Google Scholar] [CrossRef]
- Behera, B.; Unpaprom, Y.; Ramaraj, R.; Maniam, G.P.; Govindan, N.; Paramasivan, B. Integrated biomolecular and bioprocess engineering strategies for enhancing the lipid yield from microalgae. Renew. Sustain. Energy Rev. 2021, 148, 111270. [Google Scholar] [CrossRef]
- Tang, H.; Chen, M.; Garcia, M.E.D.; Abunasser, N. Culture of Microalgae Chlorella minutissima for Biodiesel Feedstock Production. Biotechnol. Bioeng. 2011, 108, 2280–2287. [Google Scholar] [CrossRef]
- Sassi, K.K.B.; Silva, J.A.; Calixto, C.D.; Sassi, R.; Sassi, C.F.C. Metabolites of interest for food technology produced by microalgae from the Northeast Brazil. Rev. Cien. Agron. 2019, 50, 54–65. [Google Scholar] [CrossRef]
- Sahu, A.; Pancha, I.; Jain, D.; Paliwal, C.; Ghosh, T.; Patidar, S.; Bhattacharya, S.; Mishra, S. Fatty acids as biomarkers of microalgae. Phytochemistry 2013, 89, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.J.; Wu, K.C.; Chan, S.C.Y.; Yau, Y.H.; Chan, K.K.; Lee, F.W.F. Investigation of Growth, Lipid Productivity, and Fatty Acid Profiles in Marine Bloom-Forming Dinoflagellates as Potential Feedstock for Biodiesel. J. Mar. Sci. Eng. 2020, 8, 381. [Google Scholar] [CrossRef]
- FAO. Dietary Protein Quality Evaluation in Human Nutrition: Report of an FAO Expert Consultation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; p. 92. [Google Scholar]
- Prates, D.F.; Duarte, J.H.; Vendruscolob, R.G.; Wagner, R.; Ballus, C.A.; Oliveira, W.S.; Godoyc, H.T.; Barcia, M.T.; Morais, M.G.; Radmanna, E.M.; et al. Role of light emitting diode (LED) wavelengths on increase of protein productivity and free amino acid profile of Spirulina sp. cultures. Bioresour. Technol. 2020, 306, 123184. [Google Scholar] [CrossRef]
- Wang, Y.; Tibbetts, S.M.; McGinn, P.J. Microalgae as Sources of High-Quality Protein for Human Food and Protein Supplements. Foods 2021, 10, 3002. [Google Scholar] [CrossRef]
- Cerri, A.; Niccolai, A.; Cardinaletti, G.; Tulli, F.; Mina, F.; Daniso, E.; Bongiorno, T.; Zittelli, G.C.; Biondi, N.; Tredici, M.R.; et al. Chemical composition and apparent digestibility of a panel of dried microalgae and cyanobacteria biomasses in rainbow trout (Oncorhynchus mykiss). Aquaculture 2021, 544, 737075. [Google Scholar] [CrossRef]
- Becker, E. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Tibbetts, S.M.; Milley, J.E.; Lall, S.P. Chemical composition and nutritional properties of freshwater and marine microalgal biomass cultured in photobioreactors. J. Appl. Phycol. 2015, 27, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Andreeva, A.; Budenkova, E.; Babich, O.; Sukhikh, S.; Ulrikh, E.; Ivanova, S.; Prosekov, A.; Dolganyuk, V. Production, Purification, and Study of the Amino Acid Composition of Microalgae Proteins. Molecules 2021, 26, 2767. [Google Scholar] [CrossRef] [PubMed]
- Bamary, Z.; Einali, A. Changes in Carbon Partitioning and Pattern of Antioxidant Enzyme Activity Induced by Arginine Treatment in the Green Microalga Dunaliella salina under Long-Term Salinity. Microb. Ecol. 2021, 103, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nehmé, R.; Atieh, C.; Fayad, S.; Claude, B.; Chartier, A.; Tannoury, M.; Elleuch, F.; Abdelkafi, S.; Pichon, C.; Morin, P. Microalgae amino acid extraction and analysis at nanomolar level using electroporation and capillary electrophoresis with laser-induced fluorescence detection. J. Sep. Sci. 2017, 40, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Carlson, R.M.; Tarchini, C.; Djerassi, C. Biological implications of recent discoveries in the marine sterol field. In Frontiers of Bioorganic Chemistry and Molecular Biology; Perg. Press Ltd.: Oxford, UK, 1980; pp. 211–224. [Google Scholar]
- Volkman, J. Sterols in microorganisms. Appl. Microbiol. Biotechnol. 2003, 60, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Jong, D.L.C.; Timmermans, K.R.; Winter, J.M.; Derksen, G.C.H. Effects of nutrient availability and light intensity on the sterol content of Saccharina latissima (Laminariales, Phaeophyceae). J. Appl. Phycol. 2021, 33, 1101–1113. [Google Scholar] [CrossRef]
- Lu, Y.; Zhou, W.; Wei, L.; Li, J.; Jia, J.; Li, F.; Smith, S.M.; Xu, J. Regulation of the cholesterol biosynthetic pathway and its integration with fatty acid biosynthesis in the oleaginous microalga Nannochloropsis oceânica. Biotechnol. Biofuels 2014, 7, 81. [Google Scholar] [CrossRef] [Green Version]
- Randhir, A.; Laird, D.W.; Maker, G.; Trengove, R.; Moheimani, N.R. Microalgae: A potential sustainable commercial source of sterols. Algal Res. 2020, 46, 101772. [Google Scholar] [CrossRef]
- Piepho, M.; Martin-Creuzburg, D.; Wacker, A. Simultaneous effects of light intensity and phosphorus supply on the sterol content of phytoplankton. PLoS ONE 2010, 5, 15828. [Google Scholar] [CrossRef]
- Fernandes, T.; Cordeiro, N. Microalgae as Sustainable Biofactories to Produce High-Value Lipids: Biodiversity, Exploitation, and Biotechnological Applications. Mar. Drugs 2021, 19, 573. [Google Scholar] [CrossRef]
- Fagundes, M.B.; Wagner, R. Biosynthesis of Sterols in Algae; IntechOpen: London, UK, 2021; Volume 9, p. 230. [Google Scholar] [CrossRef]
- Morançais, M.; Mouget, J.L.; Dumay, J. Proteins and Pigments. In Microalgae in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2018; Volume 7, pp. 145–175. [Google Scholar] [CrossRef]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Sartori, R.B.; Siqueira, S.F.; Maroneze, M.M.; Fagundes, M.B.; Wagner, R.; Zepka, L.Q.; Jacob-Lopes, E. Microalgal secondary metabolites: Effect of climatic variables, seasons, and photocycles on the biogeneration of volatile organic compounds (VOCs). J. Appl. Phycol. 2021, 33, 1457–1472. [Google Scholar] [CrossRef]
- Vélez-Landa, L.; De León, H.R.H.; Pérez-Luna, Y.C.; Velázquez-Trujillo, S.; Moreira-Acosta, J.; Berrones-Hernández, R.; Sánchez-Roque, Y. Influence of Light Intensity and Photoperiod on the Photoautotrophic Growth and Lipid Content of the Microalgae Verrucodesmus verrucosus in a Photobioreactor. Sustainability 2021, 13, 6606. [Google Scholar] [CrossRef]
- Kato, Y.; Fujihara, Y.; Vavricka, C.J.; Chang, J.S.; Hasunuma, T.; Kondo, A. Light/dark cycling causes delayed lipid accumulation and increased photoperiod-based biomass yield by altering metabolic fux in oleaginous Chlamydomonas sp. Biotechnol. Biofuels 2019, 12, 39. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, Y.B.; Ho, S.H.; Xing, F.; Ren, N.Q.; Liu, B.F. Cell growth and lipid accumulation of a microalgal mutant Scenedesmus sp. Z-4 by combining light/dark cycle with temperature variation. Biotechnol. Biofuels 2017, 10, 260. [Google Scholar] [CrossRef] [Green Version]
- Jacob-Lopes, E.; Maroneze, M.M.; Depra, M.C.; Sartori, R.B.; Dias, R.R.; Zepka, L.Q. Bioactive food compounds from microalgae: An innovative framework on industrial biorefineries. Curr. Opin. Food Sci. 2019, 25, 1–7. [Google Scholar] [CrossRef]
- Galili, G.; Avin-Wittenberg, T.; Angelovici, R.; Fernie, A.R. The role of photosynthesis and amino acid metabolism in the energy state during seed development. Front. Plant Sci. 2014, 5, 447. [Google Scholar] [CrossRef]
- Ballesteros-Torres, J.M.; Samaniego-Moreno, L.; Gomez-Flores, R.; Tamez-Guerra, R.S.; Rodríguez-Padilla, C.; Tamez-Guerra, P. Amino acids and acylcarnitine production by Chlorella vulgaris and Chlorella sorokiniana microalgae from wastewater culture. PeerJ 2019, 7, 7977. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.M.P.; Nunes, C.F.P.; Zambotti-Villela, L.; Streit, N.M.; Dias, D.; Pinto, E.; Colepicolo, P. Extraction of sterols in brown macroalgae from Antarctica and their identification by liquid chromatography coupled with tandem mass spectrometry. J. Appl. Phycol. 2017, 29, 751–757. [Google Scholar] [CrossRef]
- Strauch, S.M.; Coutinho, P.B.N. Bioactive molecules from microalgae. Nat. Bioact. Compd. 2021, 23, 453–470. [Google Scholar] [CrossRef]
- Lin, H.; Li, Y.; Hill, R.T. Microalgal and bacterial auxin biosynthesis: Implications for algal biotechnology. Curr. Opin. Biotechnol. 2022, 73, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocr. 2019, 10, 249. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Long-Term Photo-Cycle | Frequency Photo-Cycle | Short Photo-Cycle | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 24:0 | 22:2 | 20:4 | 18:6 | 12:12 | 10:14 | 2 t/d | 4 t/d | 8 t/d | 12 t/d | 24 t/d | 48 t/d | 0.91:0.09 | 0.83:0.17 | 0.75:0.25 | 0.50:0.50 | |
| C12:0 | 0.83 | 0.43 | 0.90 | 1.10 | 2.42 | 1.68 | 0.90 | 0.82 | 0.96 | 1.27 | 1.13 | 1.90 | 2.73 | 3.03 | 5.04 | 0.97 |
| C14:0 | 0.38 | 0.27 | 0.39 | 0.32 | 0.56 | 0.41 | 0.53 | 0.44 | 0.50 | 0.45 | 0.44 | 0.48 a | 0.50 | 0.47 | 0.61 | 0.57 |
| C16:0 | 45.66 | 38.18 | 44.71 | 36.72 | 39.83 | 36.87 | 47.57 | 43.95 | 45.86 | 47.02 | 44.61 | 40.41 | 37.37 | 32.75 | 32.87 | 35.04 |
| C16:1 | 0.60 | 0.38 | 0.43 | 0.51 | 0.60 | 0.73 | 0.34 | 0.29 | 0.30 | 0.55 | 0.34 | 0.83 | 0.98 | 1.00 | 1.17 | 1.16 |
| C17:0 | 0.26 | 0.34 | 0.26 | 0.32 | 0.30 | 0.27 | 0.32 | 0.33 | 0.27 | 0.30 | 0.31 | 0.32 | 0.26 | 0.25 | 0.23 | 0.26 |
| C18:0 | 3.09 | 5.04 | 5.06 | 3.36 | 2.75 | 1.67 | 4.52 | 4.73 | 5.55 | 3.75 | 4.33 | 3.47 | 2.10 | 1.74 | 1.48 | 1.82 |
| C18:1n9c | 16.10 | 18.45 | 18.86 | 14.23 | 13.37 | 11.45 | 16.39 | 19.50 | 20.62 | 16.19 | 18.30 | 14.58 | 11.25 | 11.03 | 5.80 | 6.52 |
| C18:2n6c | 11.59 | 15.90 | 11.93 | 12.54 | 9.57 | 12.23 | 13.24 | 13.81 | 11.73 | 14.32 | 13.29 | 16.12 | 13.67 | 16.54 | 11.77 | 10.47 |
| C20:0S | 0.21 | 0.25 | 0.22 | 0.22 | 0.25 | 0.24 | 0.20 | 0.22 | 0.27 | 0.16 | 0.17 | 0.16 | 0.28 | 0.22 | 0.27 | 0.31 |
| C18:3n6 | 0.71 | 0.72 | 0.59 | 0.81 | 0.87 | 1.21 | 0.66 | 0.66 | 0.64 | 0.72 | 0.74 | 1.29 | 0.85 | 1.17 | 1.16 | 1.13 |
| C18:3n3 | 20.47 | 19.81 | 16.44 | 29.67 | 29.41 | 33.16 | 15.13 | 15.04 | 13.10 | 15.16 | 16.18 | 20.25 | 29.92 | 31.77 | 39.56 | 41.67 |
| C20:1n9 | 0.10 | 0.23 | 0.20 | 0.21 | 0.06 | 0.10 | 0.20 | 0.21 | 0.19 | 0.12 | 0.15 | 0.18 | 0.07 | 0.05 | 0.05 | 0.07 |
| ∑SFA | 50.43 | 44.51 | 51.54 | 42.05 | 46.12 | 41.13 | 54.04 | 50.49 | 53.41 | 52.94 | 51.00 | 46.74 | 43.24 | 38.45 | 40.51 | 38.97 |
| ∑MUFA | 16.80 | 19.06 | 19.49 | 14.94 | 14.04 | 12.27 | 16.93 | 20.01 | 21.11 | 16.86 | 18.79 | 15.60 | 12.31 | 12.08 | 7.01 | 7.75 |
| ∑PUFA | 32.76 | 36.43 | 28.97 | 43.01 | 39.85 | 46.60 | 29.03 | 29.50 | 25.47 | 30.20 | 30.21 | 37.66 | 44.45 | 49.47 | 53.28 | 52.49 |
| TL | 11.89 n | 11.41 o | 13.74 k | 16.28 g | 15.30 i | 16.60 e | 13.33 l | 15.81 h | 16.47 f | 13.29 m | 14.82 j | 9.30 p | 16.66 d | 19.69 b | 23.26 a | 18.21 c |
| Compounds | Long-Term Photo-Cycle | Frequency Photo-Cycle | Short Photo-Cycle | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 24:0 | 22:2 | 20:4 | 18:6 | 12:12 | 10:14 | 2 t/d | 4 t/d | 8 t/d | 12 t/d | 24 t/d | 48 t/d | 0.91:0.09 | 0.83:0.17 | 0.75:0.25 | 0.50:0.50 | |
| L-Alanine | 24.10 | 27.70 | 27.40 | 39.30 | 32.90 | 42.50 | 23.30 | 27.10 | 26.50 | 25.40 | 27.30 | 32.30 | 33.80 | 34.90 | 45.50 | 40.50 |
| Glycine | 15.40 | 13.80 | 14.40 | 20.40 | 18.90 | 27.40 | 12.30 | 13.10 | 13.70 | 14.60 | 16.40 | 22.90 | 21.60 | 25.00 | 33.40 | 28.90 |
| L-Valine | 15.60 | 14.60 | 17.60 | 19.10 | 19.90 | 27.50 | 12.50 | 13.60 | 14.70 | 16.10 | 13.80 | 17.00 | 20.40 | 24.70 | 34.40 | 29.10 |
| L-Leucine | 24.40 | 23.50 | 27.00 | 29.80 | 36.10 | 38.40 | 25.90 | 21.70 | 22.90 | 25.20 | 22.20 | 29.10 | 32.30 | 35.90 | 47.50 | 42.50 |
| L-Isoleucine | 11.30 | 10.10 | 13.00 | 13.20 | 14.50 | 21.70 | 9.60 | 8.90 | 10.00 | 11.60 | 9.70 | 12.10 | 13.90 | 18.10 | 27.20 | 21.30 |
| L-Proline | 26.50 | 22.50 | 26.90 | 33.30 | 33.70 | 49.70 | 22.00 | 27.90 | 29.20 | 31.10 | 28.80 | 38.70 | 41.00 | 44.90 | 69.00 | 54.30 |
| L-Methionine | 2.10 | 2.50 | 3.10 | 4.70 | 3.40 | 5.80 | 4.8 | 3.00 | 2.70 | 3.00 | 2.90 | 3.70 | 3.50 | 4.00 | 9.90 | 7.20 |
| L-Serine | 13.90 | 12.80 | 13.90 | 16.90 | 15.00 | 26.40 | 9.00 | 15.30 | 16.30 | 14.60 | 13.50 | 17.00 | 16.90 | 22.80 | 32.40 | 24.20 |
| L-Threonine | 10.40 | 12.60 | 11.20 | 13.90 | 7.60 | 23.90 | 6.30 | 9.90 | 11.00 | 9.40 | 8.30 | 13.10 | 15.20 | 20.90 | 25.20 | 21.90 |
| L-Phenylalanine | 9.20 | 9.90 | 12.10 | 12.40 | 15.50 | 25.30 | 8.90 | 9.40 | 11.00 | 12.40 | 9.80 | 12.60 | 14.90 | 21.4 | 35.40 | 24.00 |
| L-Asparticacid | 23.10 | 24.50 | 24.40 | 29.60 | 32.00 | 45.30 | 12.70 | 25.70 | 29.70 | 29.40 | 27.60 | 32.30 | 35.00 | 44.90 | 54.60 | 45.90 |
| Hidroxiprolina | 1.20 | 1.70 | 1.40 | 2.00 | 1.50 | 2.40 | 3.40 | 1.70 | 1.70 | 2.00 | 1.40 | 2.20 | 2.20 | 1.60 | 2.70 | 2.00 |
| L-Cysteine | 1.40 | 1.70 | 1.40 | 2.20 | 1.70 | 2.40 | 2.80 | 1.70 | 1.60 | 1.50 | 1.40 | 1.50 | 2.00 | 2.90 | 3.00 | 2.10 |
| L-Glutamicacid | 25.30 | 25.20 | 29.50 | 27.60 | 25.60 | 40.90 | 16.60 | 29.80 | 30.60 | 29.90 | 26.70 | 26.40 | 30.30 | 35.40 | 48.70 | 39.10 |
| L-Asparagine | 0.50 | 0.70 | 0.60 | 0.50 | 0.50 | 0.60 | 1.10 | 0.50 | 0.40 | 0.50 | 0.30 | 0.50 | 0.50 | 0.50 | 1.00 | 0.40 |
| L-Lysine | 10.40 | 8.40 | 11.60 | 14.40 | 11.30 | 19.20 | 7.10 | 7.20 | 7.60 | 8.20 | 13.10 | 15.80 | 11.40 | 21.70 | 25.20 | 19.00 |
| L-Glutamine | 1.00 | 1.20 | 3.40 | 0.50 | 0.70 | 1.60 | 0.90 | nd | 1.00 | 1.50 | nd | nd | 0.50 | 0.30 | 1.50 | 0.90 |
| L-Cystine | 1.70 | 2.20 | 2.20 | 2.50 | 2.50 | 3.30 | 4.80 | 2.50 | 2.30 | 2.50 | 1.70 | 2.50 | 2.70 | 2.20 | 4.40 | 2.50 |
| L-Arginine | 34.00 | 36.60 | 73.00 | 36.00 | 21.10 | 57.60 | 30.10 | 85.00 | 86.70 | 94.40 | 72.40 | 57.90 | 60.10 | 46.30 | 93.60 | 56.60 |
| L-Histidine | 1.20 | 1.50 | 2.10 | 1.70 | 1.70 | 2.30 | 0.70 | 1.50 | 1.30 | 2.00 | 1.60 | 3.00 | 2.00 | 1.70 | 4.20 | 2.10 |
| L-Tyrosine | 5.50 | 5.20 | 7.50 | 6.20 | 7.60 | 16.80 | 5.40 | 6.40 | 7.00 | 8.60 | 6.90 | 6.70 | 9.90 | 15.00 | 20.80 | 15.40 |
| L-Tryptophan | 2.90 | 4.00 | 3.40 | 4.00 | 3.90 | 3.40 | 10.10 | 3.90 | 3.40 | 4.00 | 2.80 | 3.90 | 4.00 | 2.80 | 4.00 | 3.40 |
| ΣEssential AA | 87.50 k | 87.10 l | 101.10 i | 113.20 g | 113.90 f | 167.50 c | 85.90 m | 79.10 p | 84.60 n | 91.90 j | 84.20 o | 110.30 h | 117.60 e | 151.20 d | 213.00 a | 170.50 b |
| ΣNon-essential AA | 173.60 o | 175.80 n | 226.00 j | 217.00 l | 193.70 m | 316.90 b | 144.40 p | 236.70 i | 246.70 g | 256.00 f | 224.40 k | 240.90 h | 256.50 e | 276.70 d | 410.60 a | 312.80 c |
| Compounds | Long-Term Photo-Cycle | Frequency Photo-Cycle | Short Photo-Cycle | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 24:0 | 22:2 | 20:4 | 18:6 | 12:12 | 10:14 | 2 t/d | 4 t/d | 8 t/d | 12 t/d | 24 t/d | 48 t/d | 0.91:0.09 | 0.83:0.17 | 0.75:0.25 | 0.50:0.50 | |
| Squalene | 6.77 | 4.12 | 1.59 | 2.79 | 3.30 | 2.46 | 5.66 | 7.89 | 5.02 | 3.92 | 2.32 | 1.01 | 3.93 | 10.70 | 1.64 | 6.40 |
| Tocopherol | nd | 12.76 | nd | nd | nd | nd | 36.95 | 22.96 | 21.84 | 28.68 | nd | nd | nd | nd | nd | nd |
| Ergosterol | 32.86 | 10.26 | 14.43 | 27.70 | 14.81 | 23.74 | 142.52 | 79.49 | 92.15 | 28.15 | 15.92 | 6.80 | 33.53 | 79.78 | 43.32 | 66.13 |
| Stigmasterol | 83.56 | 32.99 | 35.35 | 118.33 | 49.99 | 66.18 | 450.16 | 265.91 | 239.90 | 115.68 | 54.70 | 22.06 | 112.74 | 276.55 | 130.98 | 196.31 |
| β-sitosterol | nd | nd | nd | 37.05 | nd | nd | 99.98 | 68.62 | 63.04 | 37.03 | 30.37 | nd | 36.60 | 52.28 | 36.43 | 40.35 |
| ∑Total sterol | 123.19 j | 60.13 n | 51.37 o | 185.87 i | 68.10 m | 92.38 l | 735.2 a | 444.87 b | 421.95 c | 213.46 f | 103.31 k | 29.87 p | 186.80 h | 419.31 d | 212.37 g | 309.19 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sartori, R.B.; Vendruscolo, R.G.; Ribeiro, S.R.; Furlan, V.J.M.; Wagner, R.; Zepka, L.Q.; Jacob-Lopes, E. The Role of Photo-Cycles in the Modulation of Growth and Biochemical Profile of Microalgae: Part I—Food Interest Compounds. Life 2022, 12, 462. https://doi.org/10.3390/life12030462
Sartori RB, Vendruscolo RG, Ribeiro SR, Furlan VJM, Wagner R, Zepka LQ, Jacob-Lopes E. The Role of Photo-Cycles in the Modulation of Growth and Biochemical Profile of Microalgae: Part I—Food Interest Compounds. Life. 2022; 12(3):462. https://doi.org/10.3390/life12030462
Chicago/Turabian StyleSartori, Rafaela Basso, Raquel Guidetti Vendruscolo, Stephanie Reis Ribeiro, Valcenir Júnior Mendes Furlan, Roger Wagner, Leila Queiroz Zepka, and Eduardo Jacob-Lopes. 2022. "The Role of Photo-Cycles in the Modulation of Growth and Biochemical Profile of Microalgae: Part I—Food Interest Compounds" Life 12, no. 3: 462. https://doi.org/10.3390/life12030462
APA StyleSartori, R. B., Vendruscolo, R. G., Ribeiro, S. R., Furlan, V. J. M., Wagner, R., Zepka, L. Q., & Jacob-Lopes, E. (2022). The Role of Photo-Cycles in the Modulation of Growth and Biochemical Profile of Microalgae: Part I—Food Interest Compounds. Life, 12(3), 462. https://doi.org/10.3390/life12030462