

Mortality in Pedigrees with Acute Intermittent Porphyria
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Family Tree Mortality Ratio (FTMR) Method
3. Results
3.1. Genealogical Searches
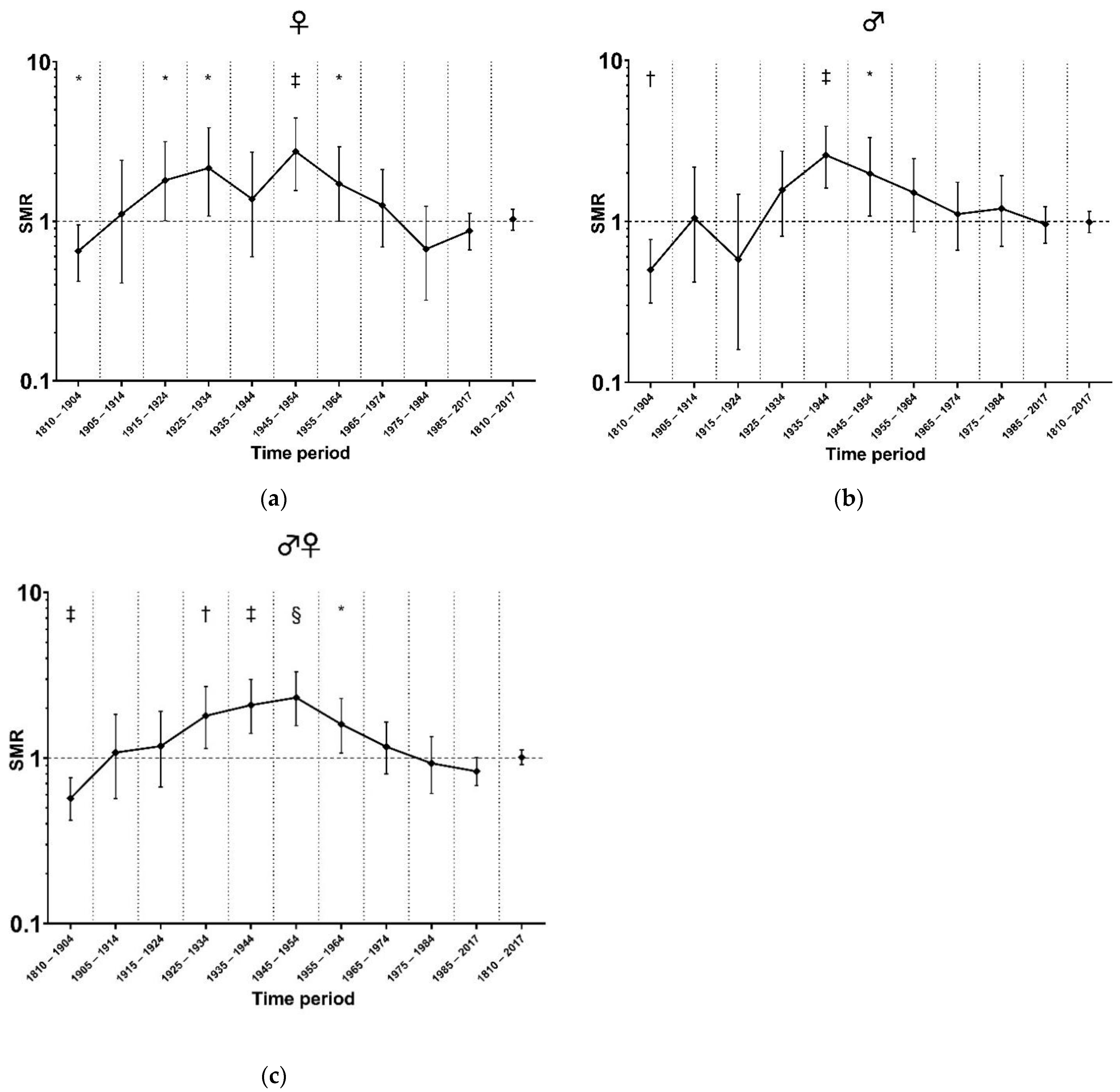
3.2. Standardized Mortality in Different Time Periods
3.3. Mortality Risk According to Age Category
4. Discussions
Strengths and Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Puy, H.; Gouya, L.; Deybach, J.C. Porphyrias. Lancet 2010, 375, 924–937. [Google Scholar] [CrossRef] [PubMed]
- Bissell, D.M.; Lai, J.C.; Meister, R.K.; Blanc, P.D. Role of delta-aminolevulinic acid in the symptoms of acute porphyria. Am. J. Med. 2015, 128, 313–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.E.; Desnick, R.J.; Stewart, M.F.; Ventura, P.; Bonkovsky, H.L. Acute Hepatic Porphyrias: “Purple Flags”-Clinical Features That Should Prompt Specific Diagnostic Testing. Am. J. Med. Sci. 2022, 363, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Naik, H.; Stoecker, M.; Sanderson, S.C.; Balwani, M.; Desnick, R.J. Experiences and concerns of patients with recurrent attacks of acute hepatic porphyria: A qualitative study. Mol. Genet. Metab. 2016, 119, 278–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neeleman, R.A.; Wagenmakers, M.A.; Koole-Lesuis, R.H.; Mijnhout, G.S.; Wilson, J.P.; Friesema, E.C.; Langendonk, J.G. Medical and financial burden of acute intermittent porphyria. J. Inherit. Metab. Dis. 2018, 41, 809–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, Y.; Puy, H.; Da Silva, V.; Simonin, S.; Robreau, A.M.; Bonaiti, C.; Phung, L.N.; Deybach, J.C. Acute intermittent porphyria: Prevalence of mutations in the porphobilinogen deaminase gene in blood donors in France. J. Intern. Med. 1997, 242, 213–217. [Google Scholar] [CrossRef]
- Chen, B.; Solis-Villa, C.; Hakenberg, J.; Qiao, W.; Srinivasan, R.R.; Yasuda, M.; Balwani, M.; Doheny, D.; Peter, I.; Chen, R.; et al. Acute Intermittent Porphyria: Predicted Pathogenicity of HMBS Variants Indicates Extremely Low Penetrance of the Autosomal Dominant Disease. Hum. Mutat. 2016, 37, 1215–1222. [Google Scholar] [CrossRef] [Green Version]
- Lenglet, H.; Schmitt, C.; Grange, T.; Manceau, H.; Karboul, N.; Bouchet-Crivat, F.; Robreau, A.-M.; Nicolas, G.; Lamoril, J.; Simonin, S.; et al. From a dominant to an oligogenic model of inheritance with environmental modifiers in acute intermittent porphyria. Hum. Mol. Genet. 2018, 27, 1164–1173. [Google Scholar] [CrossRef] [Green Version]
- Pischik, E.; Kauppinen, R. An update of clinical management of acute intermittent porphyria. Appl. Clin. Genet. 2015, 8, 201–214. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.J.; Heck, F.J. The Porphyrins and Porphyria A Review of Eighty-one Cases. Am. J. Med. 1956, 20, 239–250. [Google Scholar] [CrossRef]
- Canivet, J.; Fallot, P. The porphyrias; observations on 43 cases. Dtsch. Med. Wochenschr. 1959, 84, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A. Acute Intermittent Porphyria A Study of 50 Cases. Q. J. Med. 1959, 28, 183–209. [Google Scholar] [PubMed]
- Saint, E.G.; Curnow, D.H. Porphyria in Western Australia. Lancet 1962, 1, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Jeans, J.B.; Savik, K.; Gross, C.R.; Weimer, M.K.; Bossenmaier, I.C.; Pierach, C.A.; Bloomer, J.R. Mortality in patients with acute intermittent porphyria requiring hospitalization: A United States case series. Am. J. Med. Genet. 1996, 65, 269–273. [Google Scholar] [CrossRef]
- Sijbrands, E.J.; Westendorp, R.G.; Defesche, J.C.; de Meier, P.H.; Smelt, A.H.; Kastelein, J.J. Mortality over two centuries in large pedigree with familial hypercholesterolaemia: Family tree mortality study. BMJ 2001, 322, 1019–1023. [Google Scholar] [CrossRef] [Green Version]
- Coleman, M.P.; Herman, C.; Douglas, A. Person-Years (PYRS): A Further Program for Cohort Study Analysis; WHO: Lyons, Cook Islands, 1989. [Google Scholar]
- Roveri, G.; Nascimbeni, F.; Rocchi, E.; Ventura, P. Drugs and acute porphyrias: Reasons for a hazardous relationship. Postgrad. Med. 2014, 126, 108–120. [Google Scholar] [CrossRef]
- Handschin, C.; Lin, J.; Rhee, J.; Peyer, A.-K.; Chin, S.; Wu, P.-H.; Meyer, U.A.; Spiegelman, B.M. Nutritional regulation of hepatic heme biosynthesis and porphyria through PGC-1alpha. Cell 2005, 122, 505–515. [Google Scholar] [CrossRef] [Green Version]
- Andersson, C.; Innala, E.; Backstrom, T. Acute intermittent porphyria in women: Clinical expression, use and experience of exogenous sex hormones. A population-based study in northern Sweden. J. Intern. Med. 2003, 254, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Baravelli, C.M.; Aarsand, A.K.; Sandberg, S.; Tollanes, M.C. Sick leave, disability, and mortality in acute hepatic porphyria: A nationwide cohort study. Orphanet. J. Rare Dis. 2020, 15, 56. [Google Scholar] [CrossRef] [Green Version]
- Baumann, K.; Kauppinen, R. Long-term follow-up of acute porphyria in female patients: Update of clinical outcome and life expectancy. Mol. Genet. Metab. Rep. 2022, 30, 100842. [Google Scholar] [CrossRef]
- Ramai, D.; Deliwala, S.S.; Chandan, S.; Lester, J.; Singh, J.; Samanta, J.; di Nunzio, S.; Perversi, F.; Cappellini, F.; Shah, A.; et al. Risk of Hepatocellular Carcinoma in Patients with Porphyria: A Systematic Review. Cancers 2022, 14, 2947. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Number (%) | |
|---|---|
| Pedigrees/families included | 46 |
| Known HMBS variants * | 17 |
| Unknown HMBS variants | 3 |
| Subjects | 523 |
| Females | 270 (51.6%) |
| 1.0 probability of heterozygosity † | 264 (50.5%) |
| 0.5 probability of heterozygosity † | 259 (49.5%) |
| Follow-up time (in PYRS) ‡ | 29,679 |
| # | (NM_000190.4) | Number (%) | Number of Individuals | Number of Pedigrees |
|---|---|---|---|---|
| 1 | c.346C > T | Missense | 125 | 13 |
| 2 | c.91G > A | Missense | 74 | 6 |
| 3 | c.33 + 1G > A | Splicing | 68 | 4 |
| 4 | c.825 + 1G > A | Splicing | 39 | 1 |
| 5 | c.181dupG | Frameshift | 32 | 1 |
| 6 | c.287C > T | Missense | 24 | 2 |
| 7 | c.500G > A | Missense | 23 | 3 |
| 8 | c.991G > A | Missense | 23 | 1 |
| 9 | c.87 + 5G > A | Splicing | 21 | 1 |
| 10 | c.716A > G | Missense | 17 | 1 |
| 11 | c.709delG | Frameshift | 13 | 1 |
| 12 | c.625G > A | Missense | 13 | 1 |
| 13 | c.219_220delGA | Frameshift | 11 | 1 |
| 14 | c.499C > T | Missense | 11 | 1 |
| 15 | c.739T > C | Missense | 7 | 1 |
| 16 | c.230T > C | Missense | 5 | 1 |
| 17 | c.275T > C | Missense | 5 | 1 |
| Unknown † | Missense | 16 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neeleman, R.; Musters, K.; Wagenmakers, M.; Mijnhout, S.; Friesema, E.; Sijbrands, E.; Langendonk, J. Mortality in Pedigrees with Acute Intermittent Porphyria. Life 2022, 12, 2059. https://doi.org/10.3390/life12122059
Neeleman R, Musters K, Wagenmakers M, Mijnhout S, Friesema E, Sijbrands E, Langendonk J. Mortality in Pedigrees with Acute Intermittent Porphyria. Life. 2022; 12(12):2059. https://doi.org/10.3390/life12122059
Chicago/Turabian StyleNeeleman, Rochus, Kyra Musters, Margreet Wagenmakers, Sophie Mijnhout, Edith Friesema, Eric Sijbrands, and Janneke Langendonk. 2022. "Mortality in Pedigrees with Acute Intermittent Porphyria" Life 12, no. 12: 2059. https://doi.org/10.3390/life12122059
APA StyleNeeleman, R., Musters, K., Wagenmakers, M., Mijnhout, S., Friesema, E., Sijbrands, E., & Langendonk, J. (2022). Mortality in Pedigrees with Acute Intermittent Porphyria. Life, 12(12), 2059. https://doi.org/10.3390/life12122059