
Single Cell Cryo-Soft X-ray Tomography Shows That Each Chlamydia Trachomatis Inclusion Is a Unique Community of Bacteria

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
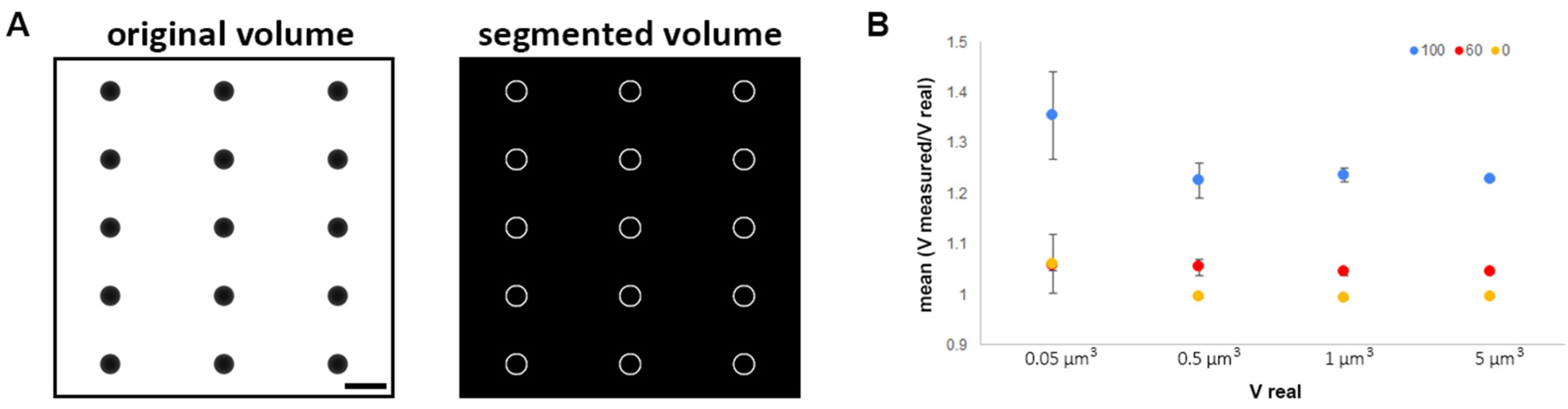
2.1. Imaging Chlamydia Inclusion with Cryo-SXT: Advantages and Limitations
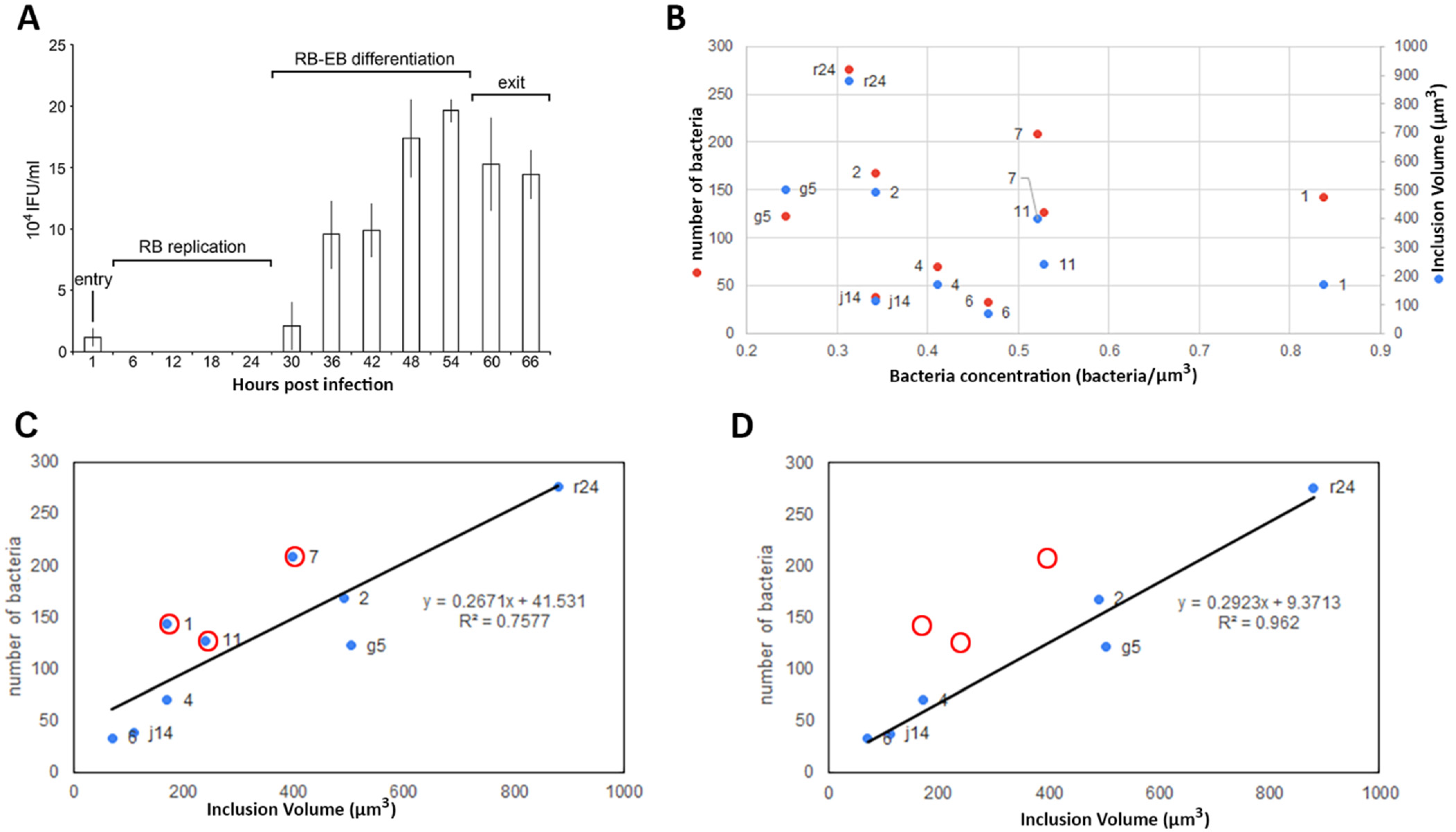
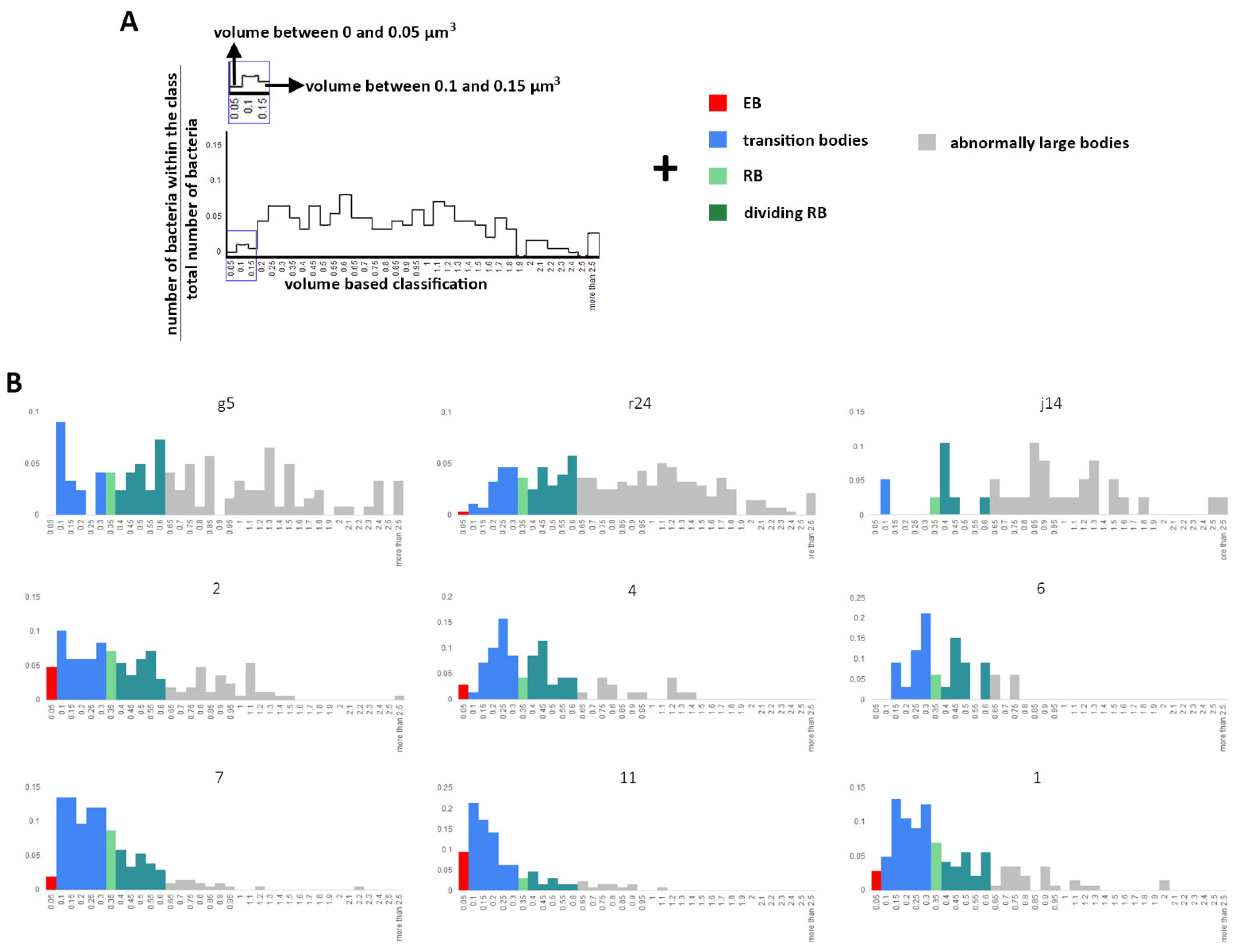
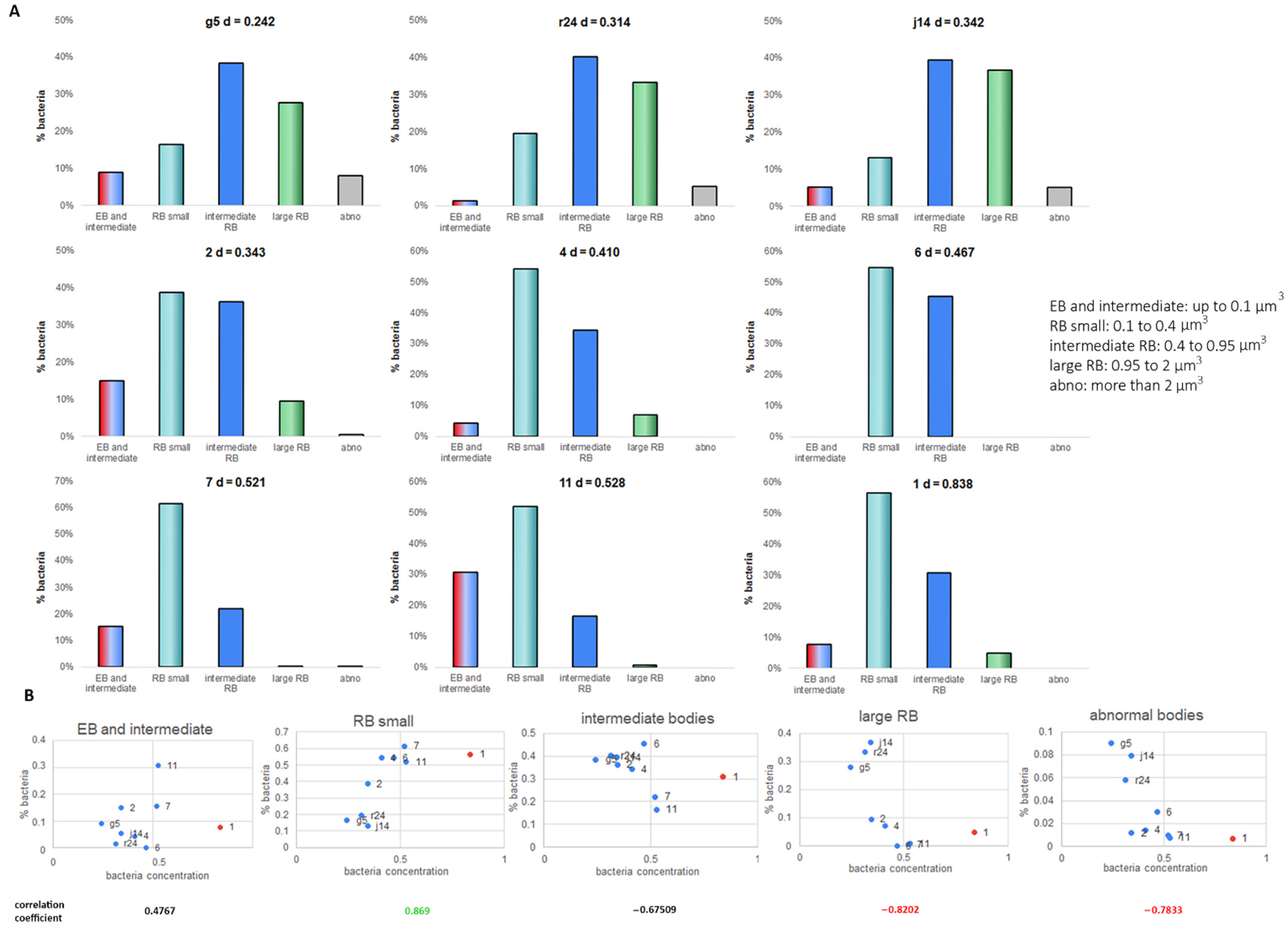
2.2. Volume Calculation and Classification of Bacteria within the Chlamydia Inclusion
2.3. Bacteria Concentration Is Not Correlated with Transition to EB but Does Correlate with the RB Volume
3. Discussion
4. Material and Methods
4.1. Sample Preparation
4.2. Soft X-ray Tomography
4.3. Tomogram Reconstruction
4.4. Simulation Data
4.5. Segmentation and Classification
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Borel, N.; Polkinghorne, A.; Pospischil, A. A Review on Chlamydial Diseases in Animals: Still a Challenge for Pathologists? Vet. Pathol. 2018, 55, 374–390. [Google Scholar] [CrossRef] [PubMed]
- Waugh, C.; Hanger, J.; Loader, J.; King, A.; Hobbs, M.; Johnson, R.; Timms, P. Infection with koala retrovirus subgroup B (KoRV-B), but not KoRV-A, is associated with chlamydial disease in free-ranging koalas (Phascolarctos cinereus). Sci. Rep. 2017, 7, 134. [Google Scholar] [CrossRef]
- Taylor, H.R.; Burton, M.J.; Haddad, D.; West, S.; Wright, H. Trachoma. Lancet 2014, 384, 2142–2152. [Google Scholar] [CrossRef]
- O’Connell, C.M.; Ferone, M.E. Chlamydia trachomatis Genital Infections. Microb. Cell 2016, 3, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Cunha, B. The atypical pneumonias: Clinical diagnosis and importance. Clin. Microbiol. Infect. 2006, 12, 12–24. [Google Scholar] [CrossRef]
- Amor, B. Chlamydia and Reiter’s syndrome. Rheumatology 1983, 22, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Boland, J.P. Chlamydia pneumoniae and cardiovascular disease: Current status. West Va. Med. J. 2003, 96, 458. [Google Scholar]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia cell biology and pathogenesis. Nat. Rev. Genet. 2016, 14, 385–400. [Google Scholar] [CrossRef]
- Engström, P.; Bergström, M.; Alfaro, A.C.; Krishnan, K.S.; Bahnan, W.; Almqvist, F.; Bergström, S. Expansion of the Chlamydia trachomatis inclusion does not require bacterial replication. Int. J. Med. Microbiol. 2015, 305, 378–382. [Google Scholar] [CrossRef]
- Higashi, N. Electron microscopic studies on the mode of reproduction of trachoma virus and psittacosis virus in cell cultures. Exp. Mol. Pathol. 1965, 4, 24–39. [Google Scholar] [CrossRef]
- Bastidas, R.J.; Valdivia, R.H. Emancipating Chlamydia: Advances in the Genetic Manipulation of a Recalcitrant Intracellular Pathogen. Microbiol. Mol. Biol. Rev. 2016, 80, 411–427. [Google Scholar] [CrossRef]
- Cottet-Rousselle, C.; Ronot, X.; Leverve, X.; Mayol, J.-F. Cytometric assessment of mitochondria using fluorescent probes. Cytom. Part A 2011, 79, 405–425. [Google Scholar] [CrossRef]
- Pattison, D.I.; Davies, M. Actions of ultraviolet light on cellular structures. In Cancer: Cell Structures, Carcinogens and Genomic Instability; Birkhäuser: Basel, Switzerland, 2006; pp. 131–157. [Google Scholar] [CrossRef]
- Zhang, P. Advances in cryo-electron tomography and subtomogram averaging and classification. Curr. Opin. Struct. Biol. 2019, 58, 249–258. [Google Scholar] [CrossRef]
- Tegunov, D.; Xue, L.; Dienemann, C.; Cramer, P.; Mahamid, J. Multi-particle cryo-EM refinement with M visualizes ribosome-antibiotic complex at 3.5 Å in cells. Nat. Methods 2021, 18, 186–193. [Google Scholar] [CrossRef]
- Egerton, R.F. Outrun radiation damage with electrons? Adv. Struct. Chem. Imaging 2015, 1, 5. [Google Scholar] [CrossRef]
- Duke, E.; Dent, K.; Razi, M.; Collinson, L.M. Biological applications of cryo-soft X-ray tomography. J. Microsc. 2014, 255, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Ekman, A.A.; Chen, J.H.; Guo, J.; McDermott, G.; Le Gros, M.A.; Larabell, C.A. Mesoscale imaging with cryo-light and X-rays: Larger than molecular machines, smaller than a cell. Biol. Cell 2017, 109, 24–38. [Google Scholar] [CrossRef]
- Lee, J.K.; Enciso, G.A.; Boassa, D.; Chander, C.N.; Lou, T.H.; Pairawan, S.S.; Guo, M.C.; Wan, F.Y.M.; Ellisman, M.H.; Sütterlin, C.; et al. Replication-dependent size reduction precedes differentiation in Chlamydia trachomatis. Nat. Commun. 2018, 9, 45. [Google Scholar] [CrossRef]
- Shelton, E.; Mowczko, W.E. Membrane blisters: A fixation artifact a study in fixation for scanning electron microscopy. Scanning 1978, 1, 166–173. [Google Scholar] [CrossRef]
- Cheville, N.F.; Stasko, J. Techniques in Electron Microscopy of Animal Tissue. Vet. Pathol. 2013, 51, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Richter, K. Cutting artefacts on ultrathin cryosections of biological bulk specimens. Micron 1994, 25, 297–308. [Google Scholar] [CrossRef]
- Monikandan, V.V.; Joseph, M.A.; Rajendrakumar, P.K. Studies on artefacts induced in the specimen preparation routines of electron microscopy characterization. IOP Conf. Ser. Mater. Sci. Eng. 2016, 149, 012016. [Google Scholar] [CrossRef]
- Kounatidis, I.; Stanifer, M.L.; Philips, M.A.; Paul-Gilloteaux, P.; Helligenstein, X.; Wang, H.; Okolo, C.A.; Fish, M.T.; Spink, M.C.; Stuard, D.I. 3D Correlative Cryo-Structured Illumination Fluorescence and Soft X-ray Microscopy Elucidates Reovirus Intra-cellular Release Pathway. Cell 2020, 182, 515–530. [Google Scholar] [CrossRef] [PubMed]
- Mellwig, R.S.; Franke, J.; Jaedicke, A.; Gorjanacz, M.; Bauer, U.; Budd, A.; Mattaj, I.W.; Devos, D.P. The compartmentalized bacteria of the planctomycetes-verrucomicrobia-chlamydiae superphylum have membrane coat-like proteins. PLoS Biol. 2010, 8, e1000281. [Google Scholar] [CrossRef] [PubMed]
- Gordon, F.B.; Quan, A.L. Occurrence of Glycogen in Inclusions of the Psittacosis-Lymphogranuloma Venereum-Trachoma Agents. J. Infect. Dis. 1965, 115, 186–196. [Google Scholar] [CrossRef]
- Dumoux, M.; Clare, D.K.; Saibil, H.R.; Hayward, R.D. Chlamydiae assemble a pathogen synapse to hijack the host endoplasmic reticulum. Traffic 2012, 13, 1612–1627. [Google Scholar] [CrossRef]
- Kremer, J.R.; Mastronarde, D.N.; Mc Intosh, J.R. Computer Visualization of Three-Dimensional Image Data Using IMOD. J. Struct. Biol. 1996, 116, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Luengo, I.; Darrow, M.C.; Spink, M.C.; Sun, Y.; Dai, W.; He, C.; Chiu, W.; Pridmore, T.; Ashton, A.W.; Duke, E.M.H.; et al. SuRVoS: Super-Region Volume Segmentation workbench. J. Struct. Biol. 2017, 198, 43–53. [Google Scholar] [CrossRef]
- Schindelin, J.; Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Radon, J. Über die Bestimmung von Funktionen durch ihre Integralwerte längs gewisser Mannigfaltigkeiten, Berichte über die Verhandlungen der Königlich-Sächsischen Akademie der Wissenschaften zu Leipzig. Math. Phys. Kl. 1917, 5, 21. [Google Scholar]
- Palmer, C.M.; Lowe, J. A cylindrical specimen holder for electron cryo-tomography. Ultramicroscopy 2014, 137, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Hayward, R.J.; Marsh, J.W.; Humphrys, M.S.; Huston, W.M.; Myers, G.S. Early Transcriptional Landscapes of Chlamydia trachomatis-Infected Epithelial Cells at Single Cell Resolution. Front Cell Infect. Microbiol. 2019, 9, 392. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A. Structural Characteristics of Chlamydial Bodies. Microbiol. Chlamydia 1988, 21–45. [Google Scholar] [CrossRef]
- Jones, F.M.; Arteta, C.; Zisserman, A.; Lempitsky, V.; Lintott, C.J.; Hart, T. Processing citizen science- and machine-annotated time-lapse imagery for biologically meaningful metrics. Sci. Data 2020, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Chiarelli, T.J.; Grieshaber, N.A.; Omsland, A.; Remien, C.H.; Grieshaber, S.S. Cell Type Development in Chlamydia trachomatis Follows a Program Intrinsic to the Reticulate Body, under revision mSystems. bioRxiv 2020. [Google Scholar] [CrossRef]
- Andrew, S.C.; Dumoux, M.; Hayward, R.D. Chlamydia Uses K+ Electrical Signalling to Orchestrate Host Sensing, Inter-Bacterial Communication and Differentiation. Microorganism 2021, 9, 173. [Google Scholar] [CrossRef]
- Prindle, A.; Liu, J.; Asally, M.; Ly, S.; Garcia-Ojalvo, J.; Süel, G.M. Ion channels enable electrical communication in bacterial communities. Nat. Cell Biol. 2015, 527, 59–63. [Google Scholar] [CrossRef]
- Bolte, S.; Cordelies, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phillips, P.; Parkhurst, J.M.; Kounatidis, I.; Okolo, C.; Fish, T.M.; Naismith, J.H.; Walsh, M.A.; Harkiolaki, M.; Dumoux, M. Single Cell Cryo-Soft X-ray Tomography Shows That Each Chlamydia Trachomatis Inclusion Is a Unique Community of Bacteria. Life 2021, 11, 842. https://doi.org/10.3390/life11080842
Phillips P, Parkhurst JM, Kounatidis I, Okolo C, Fish TM, Naismith JH, Walsh MA, Harkiolaki M, Dumoux M. Single Cell Cryo-Soft X-ray Tomography Shows That Each Chlamydia Trachomatis Inclusion Is a Unique Community of Bacteria. Life. 2021; 11(8):842. https://doi.org/10.3390/life11080842
Chicago/Turabian StylePhillips, Patrick, James M. Parkhurst, Ilias Kounatidis, Chidinma Okolo, Thomas M. Fish, James H. Naismith, Martin A. Walsh, Maria Harkiolaki, and Maud Dumoux. 2021. "Single Cell Cryo-Soft X-ray Tomography Shows That Each Chlamydia Trachomatis Inclusion Is a Unique Community of Bacteria" Life 11, no. 8: 842. https://doi.org/10.3390/life11080842
APA StylePhillips, P., Parkhurst, J. M., Kounatidis, I., Okolo, C., Fish, T. M., Naismith, J. H., Walsh, M. A., Harkiolaki, M., & Dumoux, M. (2021). Single Cell Cryo-Soft X-ray Tomography Shows That Each Chlamydia Trachomatis Inclusion Is a Unique Community of Bacteria. Life, 11(8), 842. https://doi.org/10.3390/life11080842