
Response of Controlled Cell Load Biofilms to Cold Atmospheric Plasma Jet: Evidence of Extracellular Matrix Contribution

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Cultivation Method
2.2. Biofilm Formation
2.3. Plasma Source and Exposure
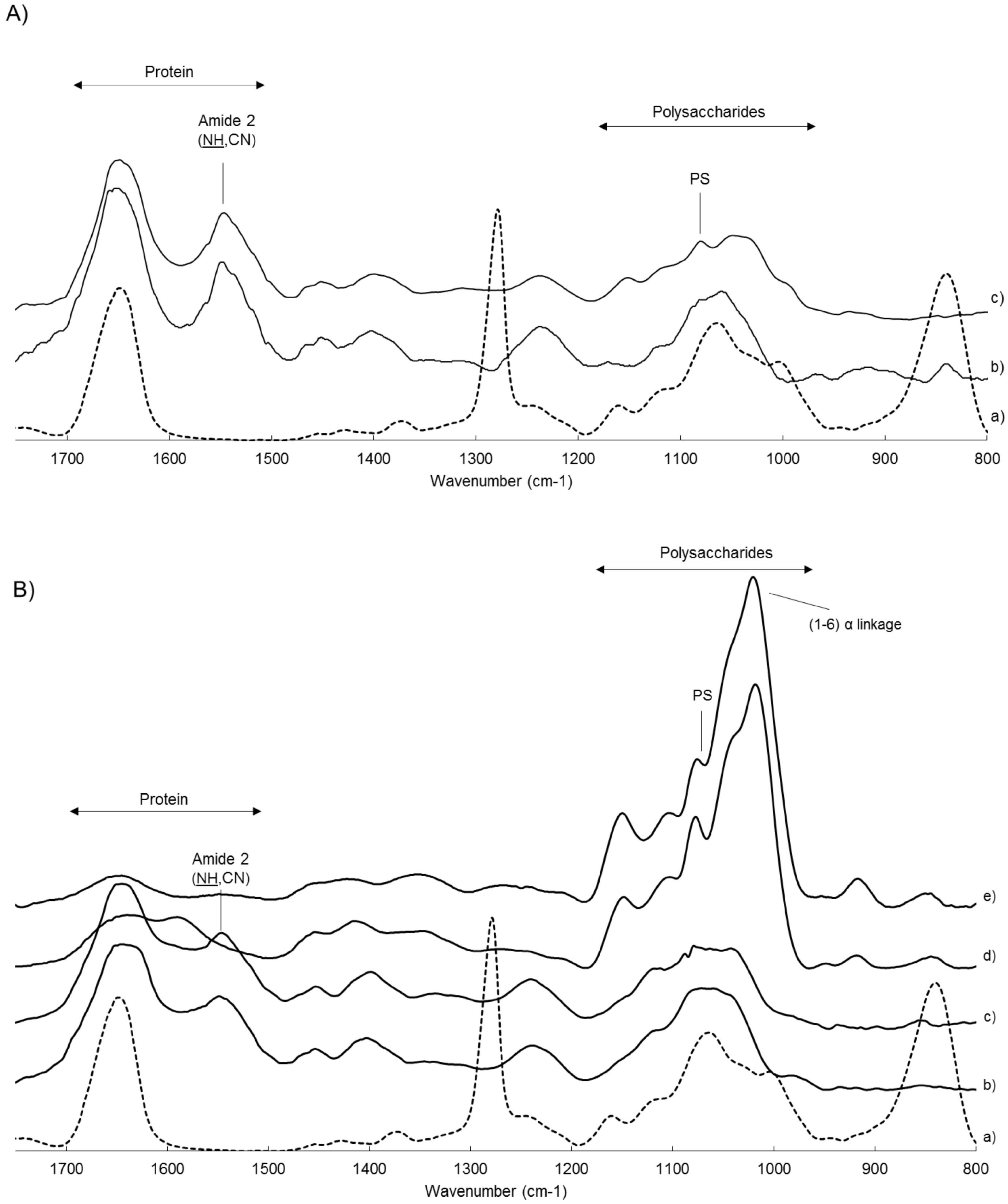
2.4. ATR-FTIR Spectroscopy
2.5. Scanning Electron Microscopy
2.6. Bacterial Viability
2.7. Statistical Analysis
3. Results
3.1. Standardization and Characterization of the Bacterial Model Biofilms
3.2. Characteristics of Corona Plasma Jet
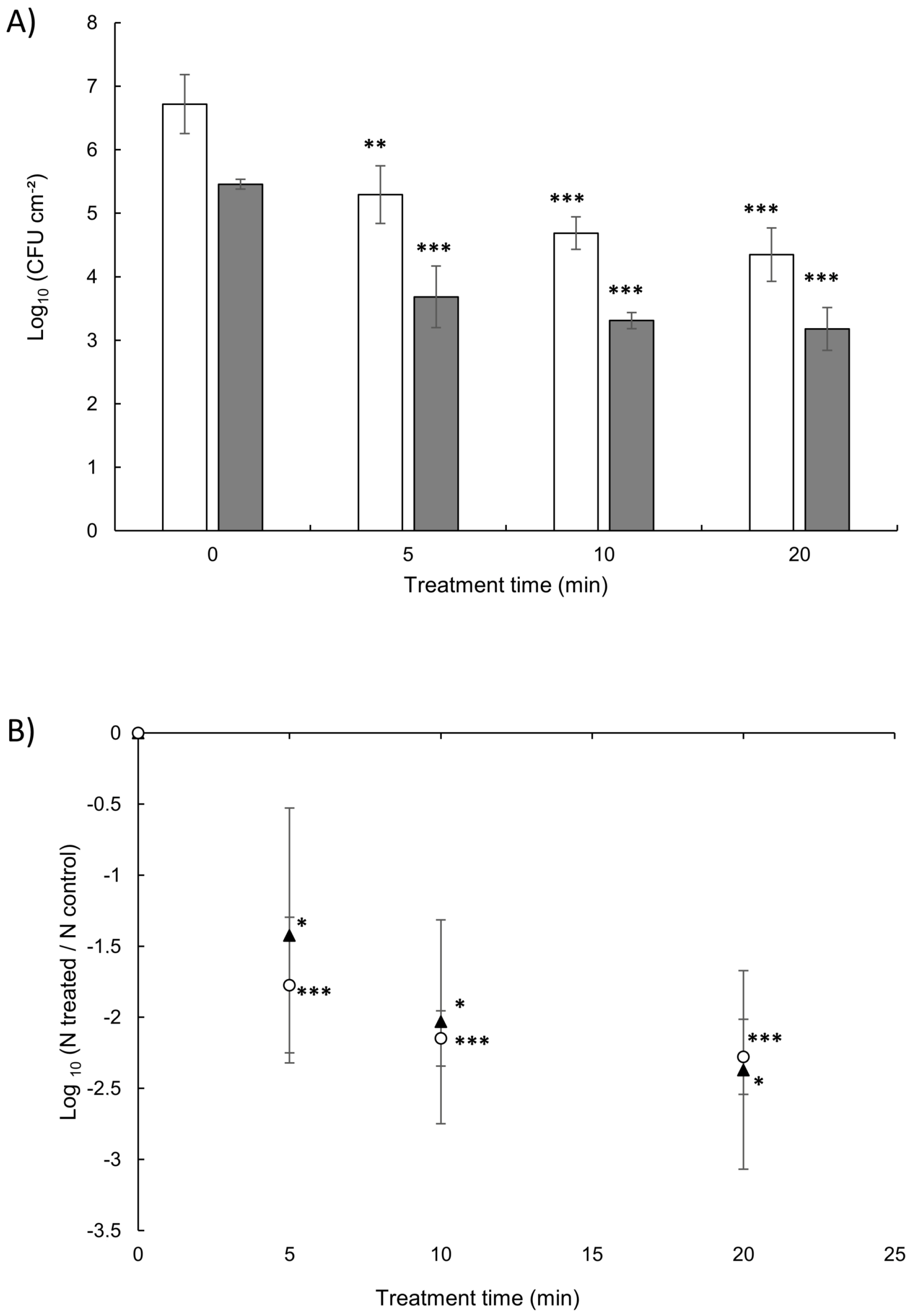
3.3. Cold Plasma Jet Treatment of Surface-Spread Cells
3.4. Viability of Biofilm Cells after Plasma Exposure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Branda, S.S.; Vik, Å.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef]
- Gilmore, B.F.; Flynn, P.B.; O’Brien, S.; Hickok, N.; Freeman, T.; Bourke, P. Cold Plasmas for Biofilm Control: Opportunities and Challenges. Trends Biotechnol. 2018, 36, 627–638. [Google Scholar] [CrossRef]
- Moreau, M.; Orange, M.; Feuilloley, M.G.J. Non-thermal plasma technologies: New tools for bio-decontamination. Biotechnol. Adv. 2008, 26, 610–617. [Google Scholar] [CrossRef]
- Lackmann, J.W.; Bandow, J.E. Inactivation of microbes and macromolecules by atmospheric-pressure plasma jets. Appl. Microbiol. Biotechnol. 2014, 98, 6205–6213. [Google Scholar] [CrossRef]
- Mai-Prochnow, A.; Murphy, A.B.; McLean, K.M.; Kong, M.G.; Ostrikov, K.K. Atmospheric pressure plasmas: Infection control and bacterial responses. Int. J. Antimicrob. Agents 2014, 43, 508–517. [Google Scholar] [CrossRef]
- Liao, X.; Liu, D.; Xiang, Q.; Ahn, J.; Chen, S.; Ye, X.; Ding, T. Inactivation mechanisms of non-thermal plasma on microbes: A review. Food Control 2017, 75, 83–91. [Google Scholar] [CrossRef]
- Flynn, P.B.; Gilmore, B.F. Understanding plasma biofilm interactions for controlling infection and virulence. J. Phys. Appl. Phys. 2018, 51, 263001. [Google Scholar] [CrossRef]
- Yousfi, M.; Merbahi, N.; Pathak, A.; Eichwald, O. Low-temperature plasmas at atmospheric pressure: Toward new pharmaceutical treatments in medicine. Fundam. Clin. Pharmacol. 2014, 28, 123–135. [Google Scholar] [CrossRef]
- Pankaj, S.K.; Wan, Z.; Keener, K.M. Effects of Cold Plasma on Food Quality: A Review. Foods 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Bourke, P.; Ziuzina, D.; Han, L.; Cullen, P.J.; Gilmore, B.F. Microbiological interactions with cold plasma. J. Appl. Microbiol. 2017, 123, 308–324. [Google Scholar] [CrossRef]
- Kamgang, J.; Briandet, R.; Herry, J.; Brisset, J.; Naïtali, M. Destruction of planktonic, adherent and biofilm cells of Staphylococcus epidermidis using a gliding discharge in humid air. J. Appl. Microbiol. 2007, 103, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Salamitou, S.; Kirkpatrick, M.J.; Ly, H.M.; Leblon, G.; Odic, E.; DuBow, M.S. Augmented Survival of Bacteria Within Biofilms to Exposure to an Atmospheric Pressure Non-Thermal Plasma Source. Biotechnology 2009, 8, 228–234. [Google Scholar] [CrossRef][Green Version]
- Marchal, F.; Robert, H.; Merbahi, N.; Fontagné-Faucher, C.; Yousfi, M.; Romain, C.E.; Eichwald, O.; Rondel, C.; Gabriel, B. Inactivation of Gram-positive biofilms by low-temperature plasma jet at atmospheric pressure. J. Phys. Appl. Phys. 2012, 45, 345202. [Google Scholar] [CrossRef]
- Puligundla, P.; Mok, C. Potential applications of nonthermal plasmas against biofilm-associated micro-organisms in vitro. J. Appl. Microbiol. 2017, 122, 1134–1148. [Google Scholar] [CrossRef]
- Yu, H.; Perni, S.; Shi, J.J.; Wang, D.Z.; Kong, M.G.; Shama, G. Effects of cell surface loading and phase of growth in cold atmospheric gas plasma inactivation of Escherichia coli K12. J. Appl. Microbiol. 2006, 101, 1323–1330. [Google Scholar] [CrossRef]
- Joshi, S.G.; Cooper, M.; Yost, A.; Paff, M.; Ercan, U.K.; Fridman, G.; Friedman, G.; Fridman, A.; Brooks, A.D. Nonthermal Dielectric-Barrier Discharge Plasma-Induced Inactivation Involves Oxidative DNA Damage and Membrane Lipid Peroxidation in Escherichia coli. Antimicrob. Agents Chemother. 2011, 55, 1053–1062. [Google Scholar] [CrossRef]
- Judée, F.; Wattieaux, G.; Merbahi, N.; Mansour, M.; Castanié-Cornet, M.P. The antibacterial activity of a microwave argon plasma jet at atmospheric pressure relies mainly on UV-C radiations. J. Phys. Appl. Phys. 2014, 47, 405201. [Google Scholar] [CrossRef]
- Fernàndez, A.; Shearer, N.; Wilson, D.; Thompson, A. Effect of microbial loading on the efficiency of cold atmospheric gas plasma inactivation of Salmonella enterica serovar Typhimurium. Int. J. Food Microbiol. 2012, 152, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Bounaix, M.S.; Gabriel, V.; Morel, S.; Robert, H.; Rabier, P.; Remaud-Siméon, M.; Gabriel, B.; Fontagné-Faucher, C.M.S.B.; Gabriel, V.; Robert, H.; et al. Biodiversity of Exopolysaccharides Produced from Sucrose by Sourdough Lactic Acid Bacteria. J. Agric. Food Chem. 2009, 57, 10889–10897. [Google Scholar] [CrossRef]
- Passerini, D.; Vuillemin, M.; Ufarté, L.; Morel, S.; Loux, V.; Fontagné-Faucher, C.; Monsan, P.; Remaud-Siméon, M.; Moulis, C. Inventory of the GH70 enzymes encoded by Leuconostoc citreum NRRL B-1299–identification of three novel α-transglucosylases. FEBS J. 2015, 282, 2115–2130. [Google Scholar] [CrossRef]
- Wang, Y.; Li, C.; Gao, C.; Ma, C.; Xu, P. Genome Sequence of the Nonpathogenic Pseudomonas aeruginosa Strain ATCC 15442. Microbiol. Resour. Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Merbahi, N.; Yousfi, M.; Eichwald, O. Device for Emitting a Plasma Jet from the Atmospheric Pressure Air at Ambient Temperature and Pressure, and Use of Said Device. Patent WO 2011/00170 A1, 1 June 2011. [Google Scholar]
- Marchal, F.; Robert, H.; Sejer, D.; Merbahi, N.; Randrianjatovo, I.; Fontagné-Faucher, C.; Di Martino, P.; Yousfi, M.; Neuhauser, E. Atmospheric cold plasmas for biofilm inactivation: Does biofilm extracellular matrix limit the bactericidal process? J. Eng. Technol. Res. 2013, 1, 123–135. [Google Scholar]
- Quilès, F.; Humbert, F.; Delille, A. Analysis of changes in attenuated total reflection FTIR fingerprints of Pseudomonas fluorescens from planktonic state to nascent biofilm state. Spectrochim. Acta Part Mol. Biomol. Spectrosc. 2010, 75, 610–616. [Google Scholar] [CrossRef]
- Khan, M.S.I.; Lee, E.J.; Kim, Y.J. A submerged dielectric barrier discharge plasma inactivation mechanism of biofilms produced by Escherichia coli O157:H7, Cronobacter sakazakii, and Staphylococcus aureus. Sci. Rep. 2016, 6, 37072. [Google Scholar] [CrossRef]
- Lackmann, J.W.; Schneider, S.; Edengeiser, E.; Jarzina, F.; Brinckmann, S.; Steinborn, E.; Havenith, M.; Benedikt, J.; Bandow, J.E. Photons and particles emitted from cold atmospheric-pressure plasma inactivate bacteria and biomolecules independently and synergistically. J. R. Soc. Interface 2013, 10, 20130591. [Google Scholar] [CrossRef]
- Soler-Arango, J.; Figoli, C.; Muraca, G.; Bosch, A.; Brelles-Mariño, G. The Pseudomonas aeruginosa biofilm matrix and cells are drastically impacted by gas discharge plasma treatment: A comprehensive model explaining plasma-mediated biofilm eradication. PLoS ONE 2019, 14, e0216817. [Google Scholar] [CrossRef]
- Bayliss, D.L.; Walsh, J.L.; Iza, F.; Shama, G.; Holah, J.; Kong, M.G. Complex Responses of Microorganisms as a Community to a Flowing Atmospheric Plasma. Plasma Process. Polym. 2012, 9, 597–611. [Google Scholar] [CrossRef]
- Flynn, P.B.; Higginbotham, S.; Alshraiedeh, N.H.; Gorman, S.P.; Graham, W.G.; Gilmore, B.F. Bactericidal efficacy of atmospheric pressure non-thermal plasma (APNTP) against the ESKAPE pathogens. Int. J. Antimicrob. Agents 2015, 46, 101–107. [Google Scholar] [CrossRef]
- Mai-Prochnow, A.; Clauson, M.; Hong, J.; Murphy, A.B. Gram positive and Gram negative bacteria differ in their sensitivity to cold plasma. Sci. Rep. 2016, 6, 38610. [Google Scholar] [CrossRef] [PubMed]
- Helgadóttir, S.; Pandit, S.; Mokkapati, V.R.S.S.; Westerlund, F.; Apell, P.; Mijakovic, I. Vitamin C Pretreatment Enhances the Antibacterial Effect of Cold Atmospheric Plasma. Front. Cell. Infect. Microbiol. 2017, 7, 43. [Google Scholar] [CrossRef]
- Humbert, F.; Quilès, F. Science against Microbial Pathogens: Communicating Current Research and Technological Advances, formatex microbiology ed.; Antonio Mendez-Vilas: Badajoz, Spain, 2011; chapter In-situ study of early stages of biofilm formation under different environmental stresses by ATR-FTIR spectroscopy; pp. 889–895. [Google Scholar]
- Randrianjatovo, I.; Girbal-Neuhauser, E.; Marcato-Romain, C.E. Epicocconone, a sensitive and specific fluorescent dye for in situ quantification of extracellular proteins within bacterial biofilms. Appl. Microbiol. Biotechnol. 2015, 99, 4835–4844. [Google Scholar] [CrossRef] [PubMed]
- Randrianjatovo-Gbalou, I.; Girbal-Neuhauser, E.; Marcato-Romain, C.E. Quantification of biofilm exopolysaccharides using an in situ assay with periodic acid–Schiff reagent. Anal. Biochem. 2016, 500, 12–14. [Google Scholar] [CrossRef]
- Mann, E.E.; Wozniak, D.J. Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Ma, L.Z. Biofilm Matrix and Its Regulation in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2013, 14, 20983–21005. [Google Scholar] [CrossRef]
- Badel, S.; Laroche, C.; Gardarin, C.; Bernardi, T.; Michaud, P. New Method Showing the Influence of Matrix Components in Leuconostoc mesenteroides Biofilm Formation. Appl. Biochem. Biotechnol. 2008, 151, 364–370. [Google Scholar] [CrossRef]
- Leathers, T.D.; Bischoff, K.M. Biofilm formation by strains of Leuconostoc citreum and L. mesenteroides. Biotechnol. Lett. 2011, 33, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Bounaix, M.S.; Robert, H.; Gabriel, V.; Morel, S.; Remaud-Siméon, M.; Gabriel, B.; Fontagné-Faucher, C. Characterization of dextran-producing Weissella strains isolated from sourdoughs and evidence of constitutive dextransucrase expression. FEMS Microbiol. Lett. 2010, 311, 18–26. [Google Scholar] [CrossRef]
- Purama, R.K.; Goswami, P.; Khan, A.T.; Goyal, A. Structural analysis and properties of dextran produced by Leuconostoc mesenteroides NRRL B-640. Carbohydr. Polym. 2009, 76, 30–35. [Google Scholar] [CrossRef]
- Han, L.; Patil, S.; Boehm, D.; Milosavljević, V.; Cullen, P.J.; Bourke, P. Mechanisms of Inactivation by High-Voltage Atmospheric Cold Plasma Differ for Escherichia coli and Staphylococcus aureus. Appl. Environ. Microbiol. 2016, 82, 450–458. [Google Scholar] [CrossRef]
- Duan, J.; Kasper, D.L. Oxidative depolymerization of polysaccharides by reactive oxygen/nitrogen species. Glycobiology 2010, 21, 401–409. [Google Scholar] [CrossRef]
- Laroussi, M. Low-Temperature Plasmas for Medicine? IEEE Trans. Plasma Sci. 2009, 37, 714–725. [Google Scholar] [CrossRef]
- Pei, X.; Lu, X.; Liu, J.; Liu, D.; Yang, Y.; Ostrikov, K.; Chu, P.K.; Pan, Y. Inactivation of a 25.5 μm Enterococcus faecalisbiofilm by a room-temperature, battery-operated, handheld air plasma jet. J. Phys. Appl. Phys. 2012, 45, 165205. [Google Scholar] [CrossRef]
- Alkawareek, M.Y.; Algwari, Q.T.; Gorman, S.P.; Graham, W.G.; O’Connell, D.; Gilmore, B.F. Application of atmospheric pressure nonthermal plasma for the in vitro eradication of bacterial biofilms. Pathog. Dis. 2012, 65, 381–384. [Google Scholar] [CrossRef]
- Yan, M.; Han, J.; Xu, X.; Liu, L.; Gao, C.; Zheng, H.; Chen, Y.; Tao, Y.; Zhou, H.; Li, Y.; et al. Gsy, a novel glucansucrase from Leuconostoc mesenteroides, mediates the formation of cell aggregates in response to oxidative stress. Sci. Rep. 2016, 6, 38122. [Google Scholar] [CrossRef] [PubMed]
- Gambino, M.; Cappitelli, F. Mini-review: Biofilm responses to oxidative stress. Biofouling 2016, 32, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, B.F. The matrix revisited: Understanding the role of extracellular matrix components in biofilm tolerance to atmospheric pressure non-thermal plasma exposure. In Proceedings of the 22nd International Symposium on Plasma Chemistry, Antwerp, Belgium, 5–10 July 2015. [Google Scholar]
- Alshraiedeh, N.H.; Kelly, S.A.; Thompson, T.P.; Flynn, P.B.; Tunney, M.M.; Gilmore, B.F. Extracellular polymeric substance-mediated tolerance of Pseudomonas aeruginosa biofilms to atmospheric pressure nonthermal plasma treatment. Plasma Process. Polym. 2020, 17, 2000108. [Google Scholar] [CrossRef]
- Alshraiedeh, N.H.; Higginbotham, S.; Flynn, P.B.; Alkawareek, M.Y.; Tunney, M.M.; Gorman, S.P.; Graham, W.G.; Gilmore, B.F. Eradication and phenotypic tolerance of Burkholderia cenocepacia biofilms exposed to atmospheric pressure non-thermal plasma. Int. J. Antimicrob. Agents 2016, 47, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Flynn, P.; Graham, W.; Gilmore, B. Acinetobacter baumannii biofilm biomass mediates tolerance to cold plasma. Lett. Appl. Microbiol. 2019, 68, 344–349. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. aeruginosa | L. citreum | ||
|---|---|---|---|
| without Sucrose | with Sucrose | ||
| Cells density (log CFU cm ) | () | () | () |
| Wet mass (mg) | () | () | () |
| Dry mass (mg) | () | () | () |
| P. aeruginosa | L. citreum | ||
|---|---|---|---|
| without Sucrose | with Sucrose | ||
| Cells | |||
| 24 h biofilm-untreated | |||
| 24 h biofilm-treated 20 min | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labadie, M.; Marchal, F.; Merbahi, N.; Girbal-Neuhauser, E.; Fontagné-Faucher, C.; Marcato-Romain, C.-E. Response of Controlled Cell Load Biofilms to Cold Atmospheric Plasma Jet: Evidence of Extracellular Matrix Contribution. Life 2021, 11, 694. https://doi.org/10.3390/life11070694
Labadie M, Marchal F, Merbahi N, Girbal-Neuhauser E, Fontagné-Faucher C, Marcato-Romain C-E. Response of Controlled Cell Load Biofilms to Cold Atmospheric Plasma Jet: Evidence of Extracellular Matrix Contribution. Life. 2021; 11(7):694. https://doi.org/10.3390/life11070694
Chicago/Turabian StyleLabadie, Maritxu, Frédéric Marchal, Nofel Merbahi, Elisabeth Girbal-Neuhauser, Catherine Fontagné-Faucher, and Claire-Emmanuelle Marcato-Romain. 2021. "Response of Controlled Cell Load Biofilms to Cold Atmospheric Plasma Jet: Evidence of Extracellular Matrix Contribution" Life 11, no. 7: 694. https://doi.org/10.3390/life11070694
APA StyleLabadie, M., Marchal, F., Merbahi, N., Girbal-Neuhauser, E., Fontagné-Faucher, C., & Marcato-Romain, C.-E. (2021). Response of Controlled Cell Load Biofilms to Cold Atmospheric Plasma Jet: Evidence of Extracellular Matrix Contribution. Life, 11(7), 694. https://doi.org/10.3390/life11070694