
Mechanosensitive TRPV4 Channel-Induced Extracellular ATP Accumulation at the Acupoint Mediates Acupuncture Analgesia of Ankle Arthritis in Rats

 and
and 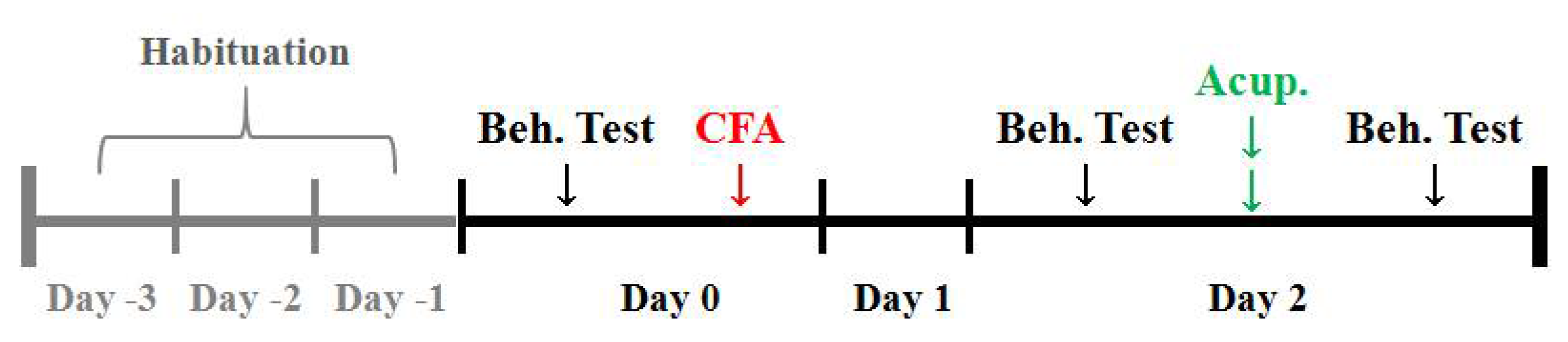{kind=link}
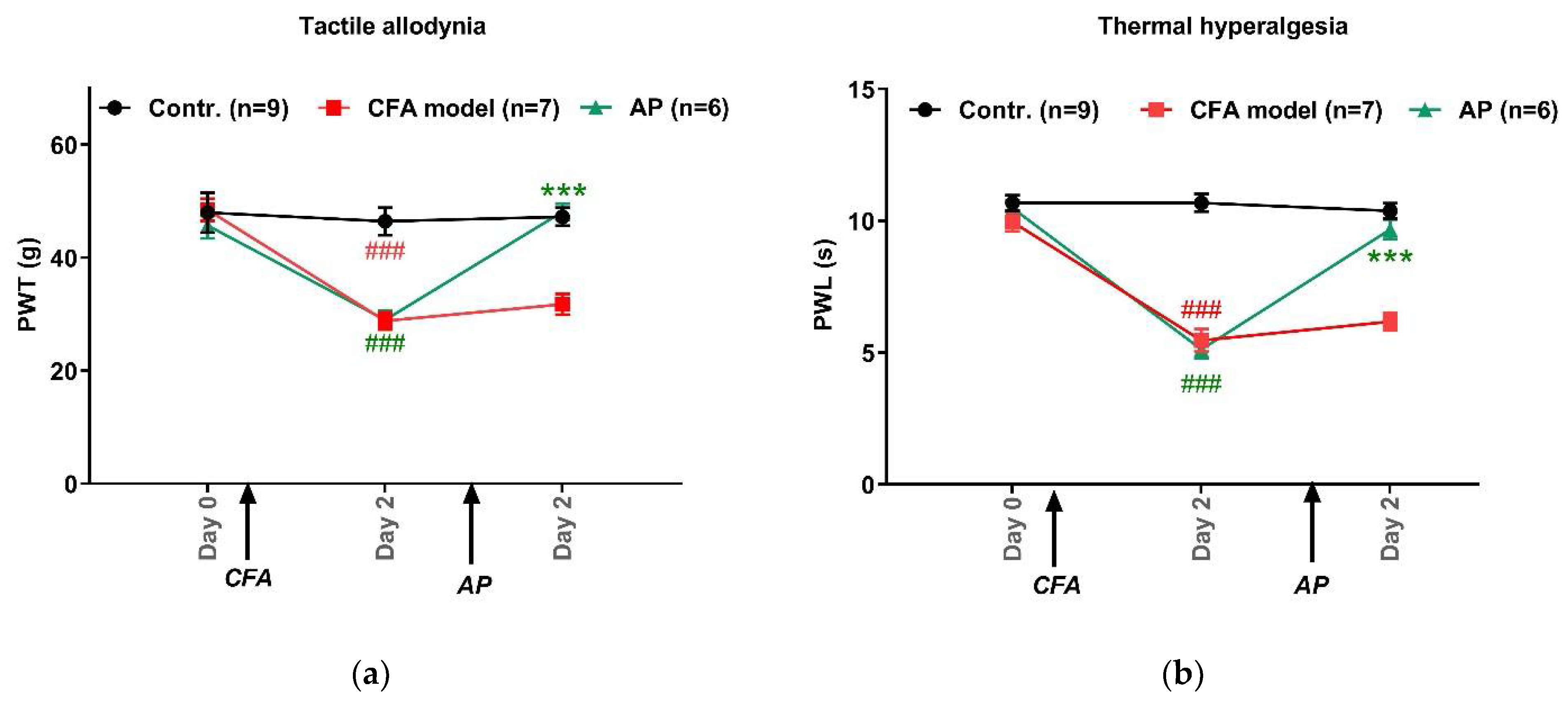{kind=link}
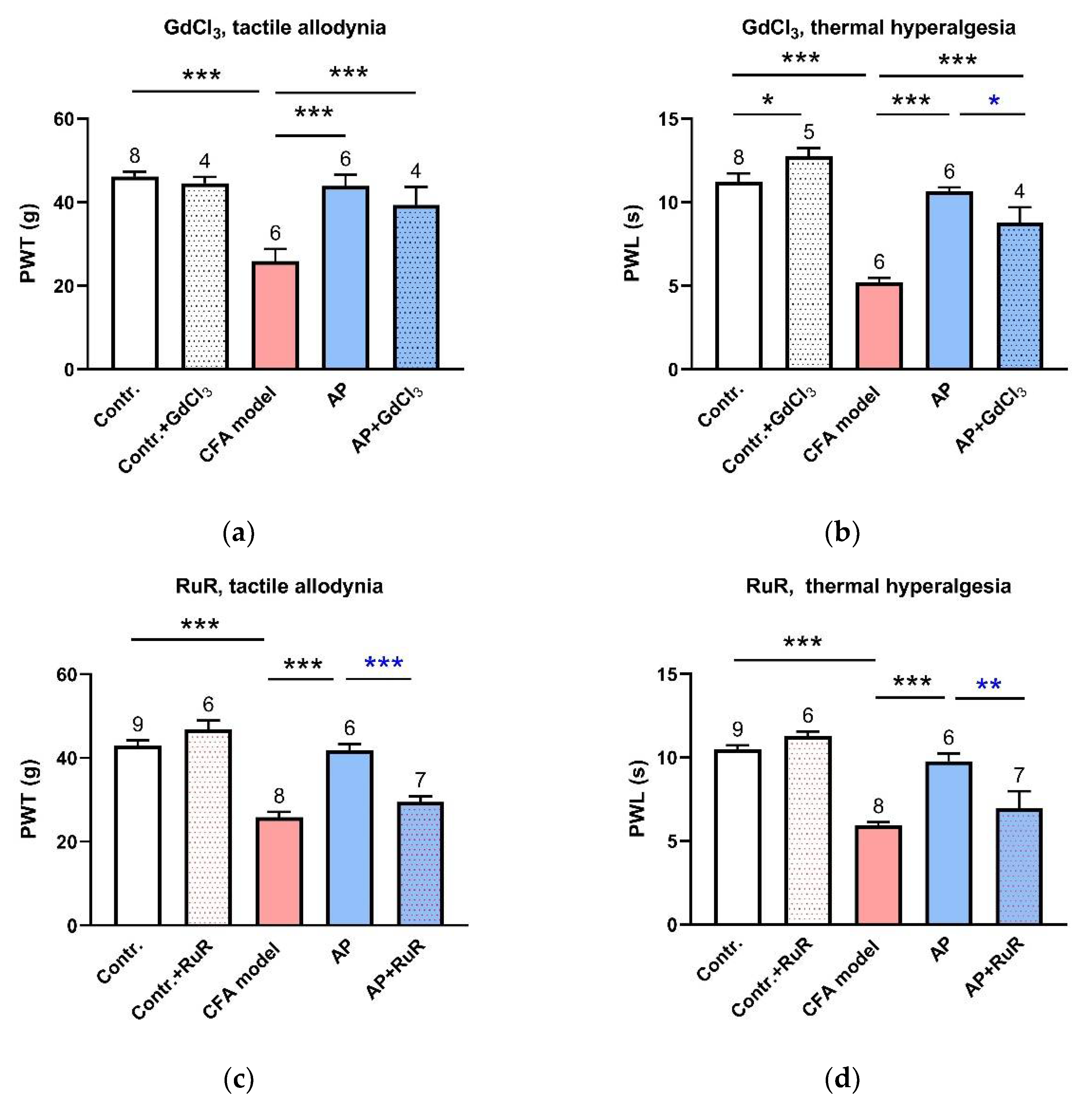{kind=link}
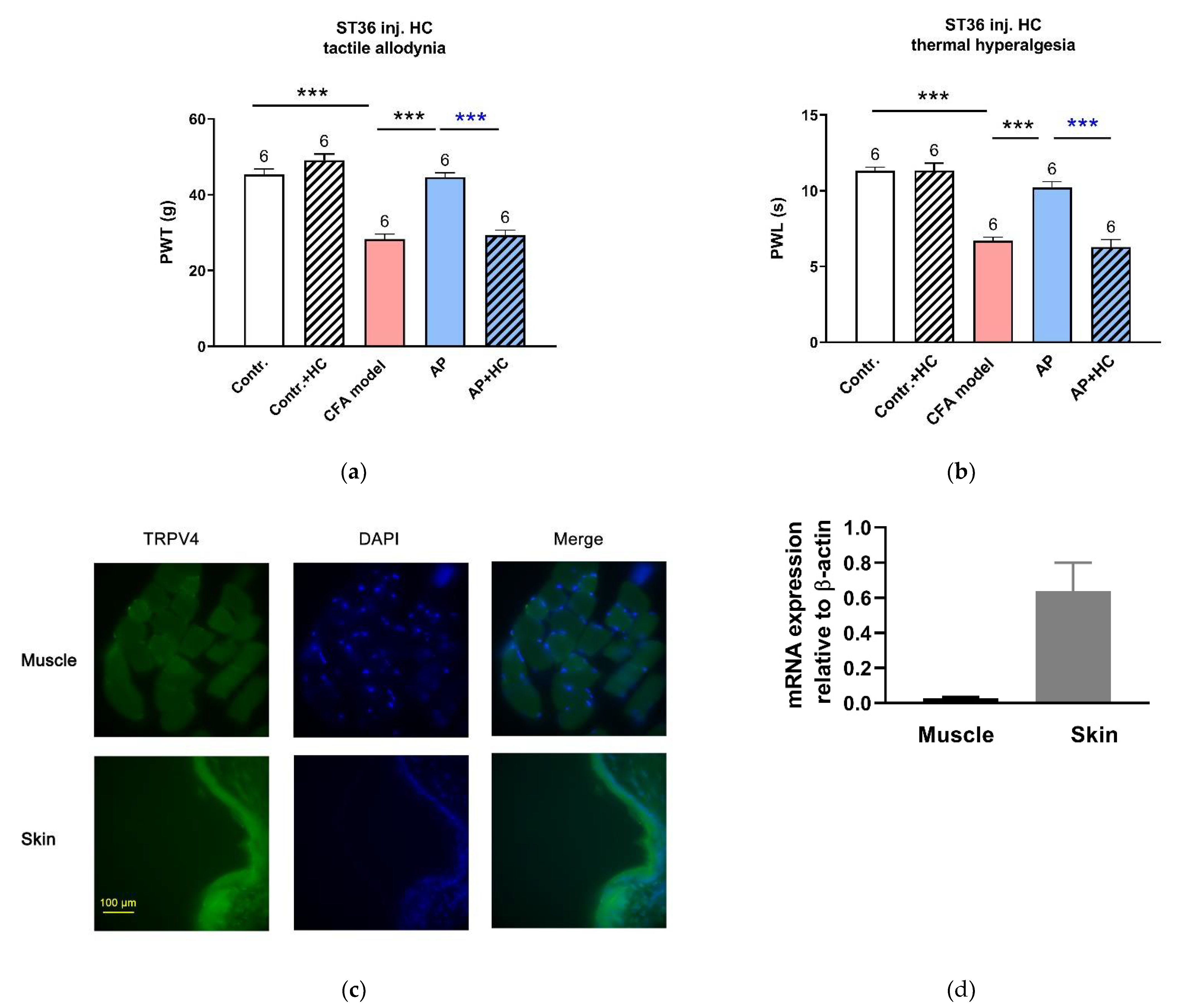{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Reagents
2.2. Animals
2.3. Acute Adjuvant Arthritis Model
2.4. Behavioral Tests
2.5. AP Treatments
2.6. Acupoint/Intraplantar Injection
2.7. ST36 Tissue Preparation
2.8. Immunofluorescence Staining on Cryosections
2.9. Microdialysis and Luminescence Analysis of eATP
2.10. Quantitative RT-PCR
2.11. Statistical Analysis
3. Results
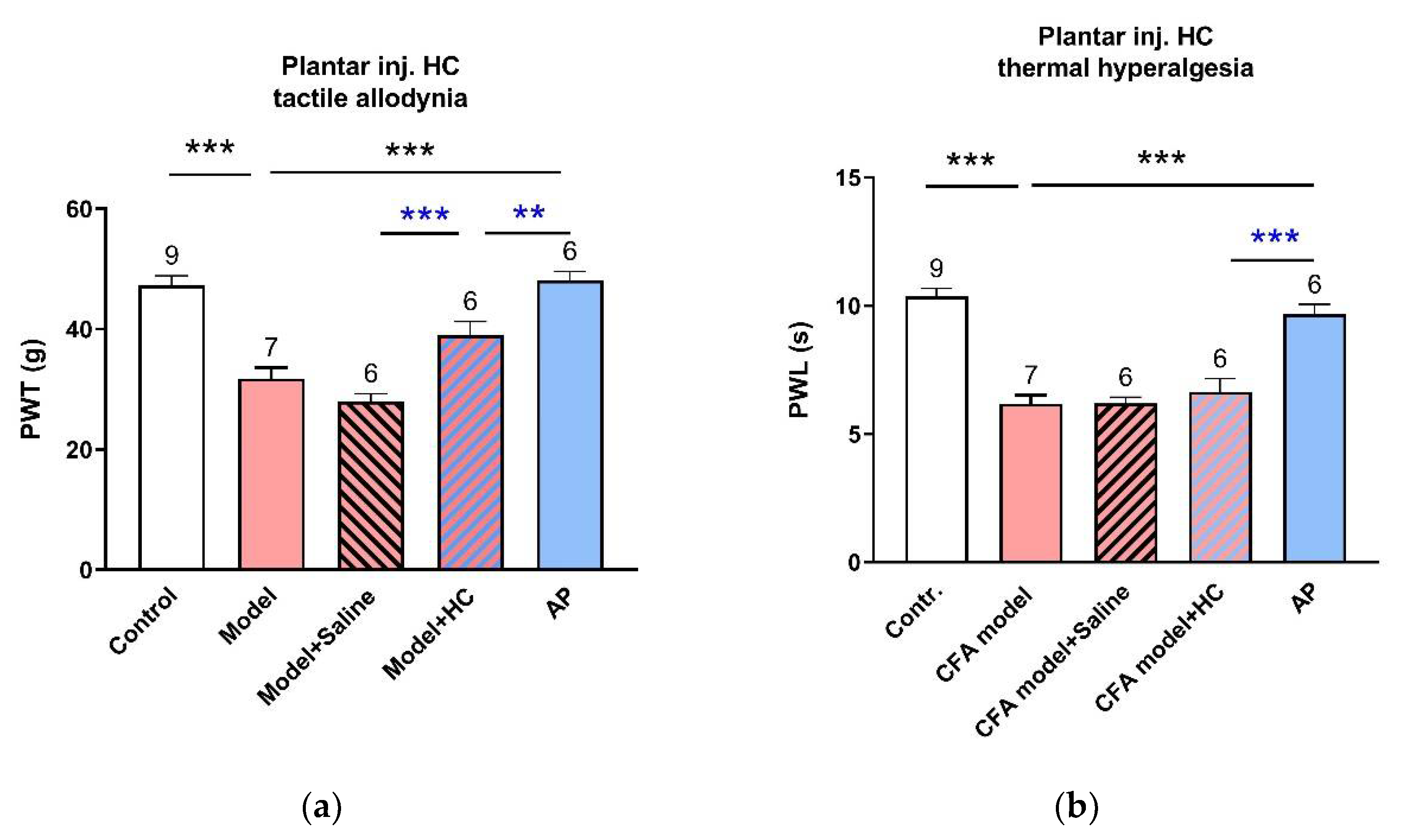
3.1. AP Treatment at ST36 Relieved CFA-Induced Hypersensitivity in Hindpaws
3.2. Inhibition of MS Channels at the Acupoints Partially Restrained AP Analgesic Effect
3.3. TRPV4 Channels at the Acupoints Participated in the AP Analgesic Mechanism as Non-Nociceptive Protein
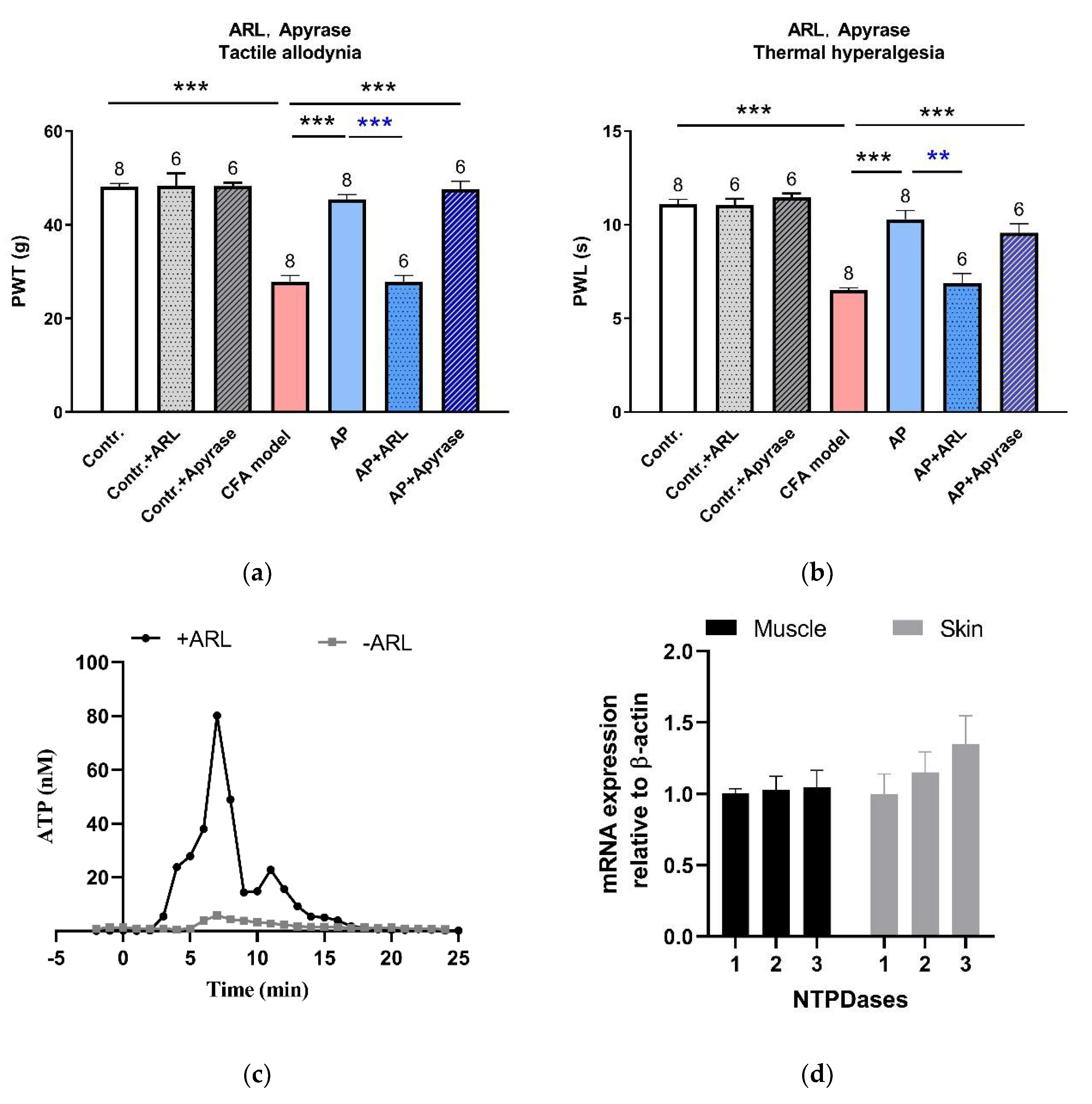
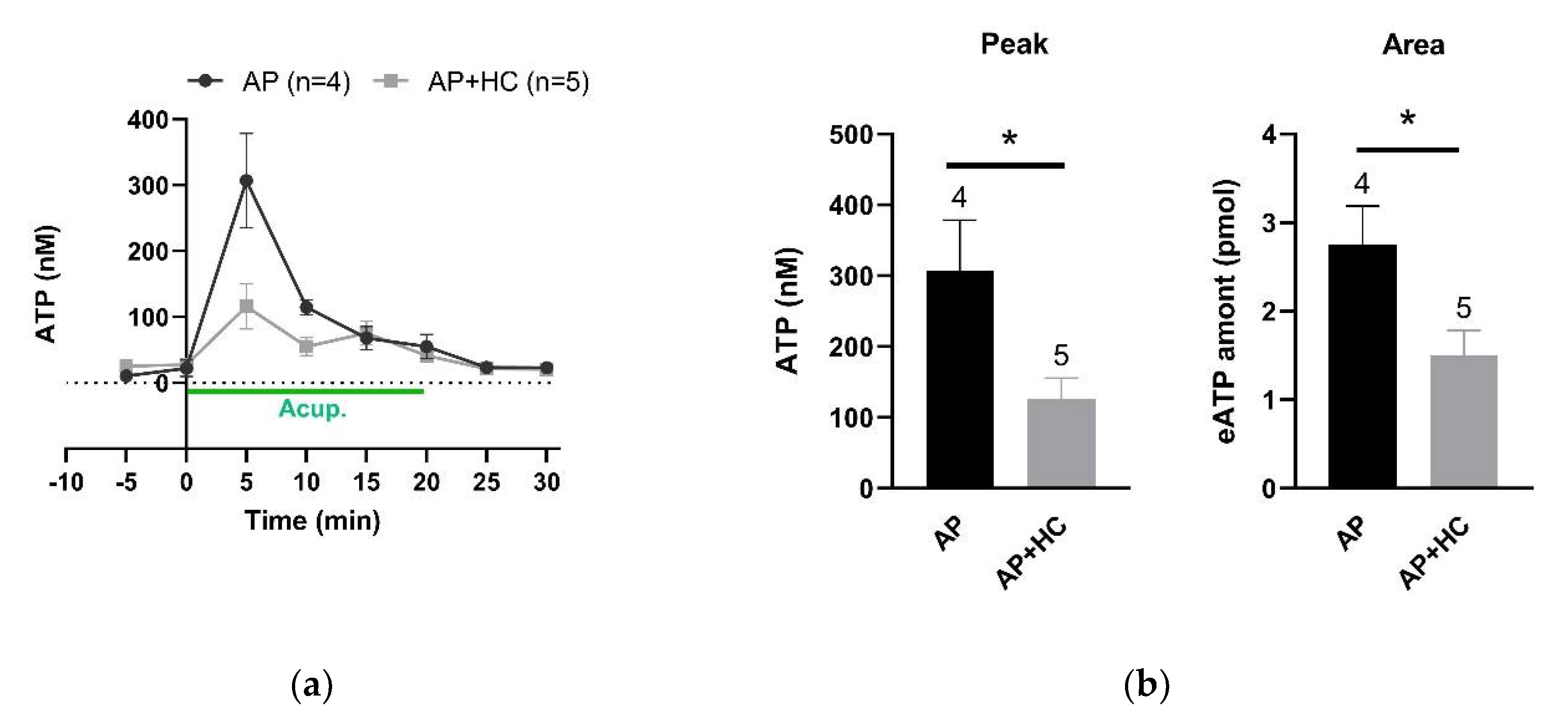
3.4. Needling-Induced eATP Transient Accumulation Mediated AP Analgesic Effect
3.5. TRPV4 Channels Mediated eATP Accumulation at the Acupoints during AP
4. Discussion
4.1. AP Treatments Exerted Immediate Analgesic Effect on Acute Ankle Arthritis-Induced Pain Hypersensitivity
4.2. The Mechanism of AP Analgesia Involved the Activation of Local Mechanosensitive TRPV4 Channels
4.3. Needling-Induced Local eATP Accumulation Via Activation of TRPV4 Channels
4.4. Local eATP Hydrolysis was Necessary for AP Analgesia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ado | Adenosine |
| AP | Acupuncture |
| ARL67156 | 6-N,N-Diethyl-β-γ-dibromomethylene-D-adenosine-5′-triphosphate |
| CFA | Complete Freund’s adjuvant |
| DMSO | Dimethyl sulfoxide |
| eATP | Extracellular ATP |
| HBSS | Hank’s balanced salt solution |
| HC067047 | (2-Methyl-1-[3-(4-morpholinyl)propyl]-5-phenyl-N-[3-(trifluoromethyl)phenyl]-1H-pyrrole-3-carboxamide |
| L-L assay | Luciferin–luciferase assay |
| MS | Mechanosensitive |
| Nt5e | Ecto-5′-nucleotidase |
| NTPDases | Nucleoside triphosphate diphosphohydrolases |
| OCT | Optimal cutting temperature compound |
| PAP | Prostatic acid phosphates |
| PBS | Phosphate-buffered saline |
| PFA | Paraformaldehyde |
| PWL | Paw withdraw latency |
| PWT | Paw withdraw threshold |
| RuR | Ruthenium red |
| TRPA1 | Transient receptor potential ankyrin 1 |
| TRPV4 | Transient receptor potential vanilloid 4 |
References
- Sheng, H.J. Acupuncture analgesia: Areas of consensus and controversy. Pain 2011, 152, 41–48. [Google Scholar]
- Zhao, Z.Q. Neural mechanism underlying acupuncture analgesia. Prog. Neurobiol. 2008, 85, 355–375. [Google Scholar] [CrossRef]
- Ding, G.; Zhang, D.; Huang, M.; Wang, L.; Yao, W. The function of collagen and mast cells at acupoints. In Current Research in Acupuncture, 1st ed.; Xia, Y., Ding, G., Wu, G., Eds.; Springer: New York, NY, USA, 2012; pp. 53–87. [Google Scholar]
- Zhang, D.; Ding, G.; Shen, X.; Yao, W.; Zhang, Z.; Zhang, Y.; Lin, J.; Gu, Q. Role of mast cells in acupuncture effect: A pilot study. Explore 2008, 4, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ding, G.; Huang, H.; Lin, J.; Yao, W.; Zhan, R. Role of collagen fibers in acupuncture analgesia therapy on rats. Connect. Tissue Res. 2009, 50, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Langevin, H.M.; Churchill, D.L.; Wu, J.; Badger, G.J.; Krag, M.H. Evidence of connective tissue involvement in acupuncture. FASEB J. 2002, 16, 872–874. [Google Scholar] [CrossRef]
- Zhang, D.; Spielmann, A.; Wang, L.; Ding, G.; Huang, F.; Gu, Q.; Schwarz, W. Mast-cell degranulation induced by physical stimuli involves the activation of transient-receptor-potential channel TRPV2. Physiol. Res. 2012, 61, 113–124. [Google Scholar] [CrossRef]
- Wang, L.; Sikora, J.; Hu, L.; Shen, X.; Grygorczyk, R.; Schwarz, W. ATP release from mast cells by physical stimulation: A putative early step in activation of acupuncture points. Evid. Based Complement. Alternat. Med. 2013, 2013, 350949. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Wang, X.; Xing, B.; Yang, H.; Sa, Z.; Zhang, D.; Yao, W.; Yin, N.; Xia, Y.; Ding, G. Critical roles of TRPV2 channels, histamine H1 and adenosine A1 receptors in the initiation of acupoint signals for acupuncture analgesia. Sci. Rep. 2018, 8, 6523–6533. [Google Scholar] [CrossRef]
- Ohsaki, A.; Tanuma, S.; Tsukimoto, M. TRPV4 channel-regulated ATP release contributes to gamma-irradiation-induced production of IL-6 and IL-8 in epidermal keratinocytes. Biol. Pharm. Bull. 2018, 41, 1620–1626. [Google Scholar] [CrossRef] [PubMed]
- Pritschow, B.W.; Lange, T.; Kasch, J.; Kunert-Keil, C.; Liedtke, W.; Brinkmeier, H. Functional TRPV4 channels are expressed in mouse skeletal muscle and can modulate resting Ca2+ influx and muscle fatigue. Pflügers Arch. Eur. J. Physiol. 2011, 461, 115–122. [Google Scholar] [CrossRef]
- Ho, T.C.; Horn, N.A.; Tuan, H.; Kelava, L.; Lansman, J.B. Evidence TRPV4 contributes to mechanosensitive ion channels in mouse skeletal muscle fibers. Channels 2012, 6, 246–254. [Google Scholar] [CrossRef]
- Bakri, M.M.; Yahya, F.; Munawar, K.M.M.; Kitagawa, J.; Hossain, M.Z. Transient receptor potential vanilloid 4 (TRPV4) expression on the nerve fibers of human dental pulp is upregulated under inflammatory condition. Arch. Oral Biol. 2018, 89, 94–98. [Google Scholar] [CrossRef]
- Takano, T.; Chen, X.; Luo, F.; Fujita, T.; Ren, Z.; Goldman, N.; Zhao, Y.; Markman, J.D.; Nedergaard, M. Traditional acupuncture triggers a local increase in adenosine in human subjects. J. Pain 2012, 13, 1215–1223. [Google Scholar] [CrossRef]
- Goldman, N.; Chen, M.; Fujita, T.; Xu, Q.; Peng, W.; Liu, W.; Jensen, T.K.; Pei, Y.; Wang, F.; Han, X.; et al. Adenosine A1 receptors mediate local anti-nociceptive effects of acupuncture. Nat. Neurosci. 2010, 13, 883–888. [Google Scholar] [CrossRef]
- Mikolajewicz, N.; Mohammed, A.; Morris, M.; Komarova, S.V. Mechanically stimulated ATP release from mammalian cells: Systematic review and meta-analysis. J. Cell Sci. 2018, 131, 223354. [Google Scholar] [CrossRef]
- Egbuniwe, O.; Grover, S.; Duggal, A.K.; Mavroudis, A.; Yazdi, M.; Renton, T.; Di Silvio, L.; Grant, A.D. TRPA1 and TRPV4 activation in human odontoblasts stimulates ATP release. J. Dent. Res. 2014, 93, 911–917. [Google Scholar] [CrossRef]
- Zheng, Y.W.; Wu, M.Y.; Shen, X.Y.; Wang, L.N. Application of unrestrained conscious rats with acute inflammatory ankle pain to study of acupuncture analgesia. Zhen Ci Yan Jiu 2020, 45, 645–651. [Google Scholar]
- Shen, D.; Zhen, Y.; Ding, G.; Wang, L.; Shen, X. Comparative study on the analgesic effect of acupuncture along meridians or sciatic nerves in rats with acute ankle inflammation. Chin. Arch. Tradit. Chin. Med. 2021, 39, 119–123. [Google Scholar]
- Shen, D.; Shen, X.; Schwarz, W.; Grygorczyk, R.; Wang, L. P2Y13 and P2X7 receptors modulate mechanically induced adenosine triphosphate release from mast cells. Exp. Dermatol. 2020, 29, 499–508. [Google Scholar] [CrossRef]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef]
- Alessandri-Haber, N.; Yeh, J.J.; Boyd, A.E.; Parada, C.A.; Chen, X.; Reichling, D.B.; Levine, J.D. Hypotonicity Induces TRPV4-mediated nociception in rat. Neuron 2003, 39, 497–511. [Google Scholar] [CrossRef]
- Alessandri-Haber, N.; Joseph, E.; Dina, O.A.; Liedtke, W.; Levine, J.D. TRPV4 mediates pain-related behavior induced by mild hypertonic stimuli in the presence of inflammatory mediator. Pain 2005, 118, 70–79. [Google Scholar] [CrossRef]
- Zimmermann, H.; Zebisch, M.; Strater, N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef]
- Tang, Y.; Yin, H.; Rubini, P.; Illes, P. Acupuncture-induced analgesia: A neurobiological basis in purinergic signaling. Neuroscientist 2016, 22, 563–578. [Google Scholar] [CrossRef]
- Shen, D.; Zheng, Y.; Zhang, D.; Shen, X.; Wang, L. Acupuncture modulates extracellular ATP levels in peripheral sensory nervous system during analgesia of ankle arthritis in rats. Purinergic Signal. 2021. [Google Scholar] [CrossRef]
- Wang, L.; Ding, G.; Gu, Q.; Schwarz, W. Single-channel properties of a stretch-sensitive chloride channel in the human mast cell line HMC-1. Eur. Biophys. J. 2010, 39, 757–767. [Google Scholar] [CrossRef][Green Version]
- Gunthorpe, M.J.; Benham, C.D.; Randall, A.; Davis, J.B. The diversity in the vanilloid (TRPV) receptor family of ion channels. Trends Pharmacol. Sci. 2002, 23, 183–191. [Google Scholar] [CrossRef]
- Dias, F.C.; Alves, V.S.; Matias, D.O.; Figueiredo, C.P.; Costa, R. The selective TRPV4 channel antagonist HC-067047 attenuates mechanical allodynia in diabetic mice. Eur. J. Pharmacol. 2019, 856, 172408. [Google Scholar] [CrossRef]
- Wood, J.N.; Eijkelkamp, N. Noxious mechanosensation—molecules and circuits. Curr. Opin. Pharmacol. 2012, 12, 4–8. [Google Scholar] [CrossRef]
- Mochizuki, T.; Sokabe, T.; Araki, I.; Fujishita, K.; Shibasaki, K.; Uchida, K.; Naruse, K.; Koizumi, S.; Takeda, M.; Tominaga, M. The TRPV4 cation channel mediates stretch-evoked Ca2+ influx and ATP release in primary urothelial cell cultures. J. Biol. Chem. 2009, 284, 21257–21264. [Google Scholar] [CrossRef]
- Strotmann, R.; Harteneck, C.; Nunnenmacher, K.; Schultz, G.; Plant, T.D. OTRPC4, a nonselective cation channel that confers sensitivity to extracellular osmolarity. Nat. Cell Biol. 2000, 2, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Tatur, S.; Groulx, N.; Orlov, S.N.; Grygorczyk, R. Ca2+-dependent ATP release from A549 cells involves synergistic autocrine stimulation by coreleased uridine nucleotides. J. Physiol. 2007, 584, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Bonvini, S.J.; Birrell, M.A.; Dubuis, E.; Adcock, J.J.; Wortley, M.A.; Flajolet, P.; Bradding, P.; Belvisi, M.G. Novel airway smooth muscle-mast cell interactions and a role for the TRPV4-ATP axis in non-atopic asthma. Eur. Respir. J. 2020, 56, 11. [Google Scholar] [CrossRef]
- Mihara, H.; Boudaka, A.; Sugiyama, T.; Moriyama, Y.; Tominaga, M. Transient receptor potential vanilloid 4 (TRPV4)-dependent calcium influx and ATP release in mouse oesophageal keratinocytes. J. Physiol. Lond. 2011, 589, 3471–3482. [Google Scholar] [CrossRef]
- Rahman, M.; Sun, R.; Mukherjee, S.; Nilius, B.; Janssen, L.J. TRPV4 Stimulation Releases ATP via Pannexin Channels in Human Pulmonary Fibroblasts. Am. J. Respir. Cell Mol. Biol. 2018, 59, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Levesque, S.A.; Lavoie, E.G.; Lecka, J.; Bigonnesse, F.; Sevigny, J. Specificity of the ecto-ATPase inhibitor ARL 67156 on human and mouse ectonucleotidases. Br. J. Pharmacol. 2007, 152, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Zylka, M.J.; Sowa, N.A.; Taylor-Blake, B.; Twomey, M.A.; Herrala, A.; Voikar, V.; Vihko, P. Prostatic acid phosphatase is an ectonucleotidase and suppresses pain by generating adenosine. Neuron 2008, 60, 111–122. [Google Scholar] [CrossRef]
- Sowa, N.A.; Taylor-Blake, B.; Zylka, M.J. Ecto-5′-nucleotidase (CD73) inhibits nociception by hydrolyzing AMP to adenosine in nociceptive circuits. J. Neurosci. 2010, 30, 2235–2244. [Google Scholar] [CrossRef]
- Sowa, N.A.; Voss, M.K.; Zylka, M.J. Recombinant ecto-5′-nucleotidase (CD73) has long lasting antinociceptive effects that are dependent on adenosine A1 receptor activation. Mol. Pain 2010, 6, 20. [Google Scholar] [CrossRef]
- Hurt, J.K.; Zylka, M.J. PAPupuncture has localized and long-lasting antinociceptive effects in mouse models of acute and chronic pain. Mol. Pain 2012, 8, 28. [Google Scholar] [CrossRef]
- Ma, L.; Thu, T.; Ren, Y.; Dirksen, R.T.; Liu, X. Neuronal NTPDase3 mediates extracellular ATP degradation in trigeminal nociceptive pathway. PLoS ONE 2016, 11, e0164028. [Google Scholar] [CrossRef]
- Vongtau, H.O.; Lavoie, E.G.; Sévigny, J.; Molliver, D.C. Distribution of ecto-nucleotidases in mouse sensory circuits suggests roles for nucleoside triphosphate diphosphohydrolase-3 in nociception and mechanoreception. Neuroscience 2011, 193, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Mizumoto, N.; Kumamoto, T.; Robson, S.C.; Sévigny, J.; Matsue, H.; Enjyoji, K.; Takashima, A. CD39 is the dominant Langerhans cell–associated ecto-NTPDase: Modulatory roles in inflammation and immune responsiveness. Nat. Med. 2002, 8, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Kinoshita, M.; Mizumura, N.; Miyazaki, S.; Aoki, R.; Momosawa, A.; Shimada, S.; Kambe, T.; Kawamura, T. Purinergic molecules in the epidermis. J. Investig. Dermatol. 2018, 138, 2486–2488. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Zuo, W.; Shen, D.; Cui, K.; Huang, M.; Zhang, D.; Shen, X.; Wang, L. Mechanosensitive TRPV4 Channel-Induced Extracellular ATP Accumulation at the Acupoint Mediates Acupuncture Analgesia of Ankle Arthritis in Rats. Life 2021, 11, 513. https://doi.org/10.3390/life11060513
Zheng Y, Zuo W, Shen D, Cui K, Huang M, Zhang D, Shen X, Wang L. Mechanosensitive TRPV4 Channel-Induced Extracellular ATP Accumulation at the Acupoint Mediates Acupuncture Analgesia of Ankle Arthritis in Rats. Life. 2021; 11(6):513. https://doi.org/10.3390/life11060513
Chicago/Turabian StyleZheng, Yawen, Weimin Zuo, Dan Shen, Kaiyu Cui, Meng Huang, Di Zhang, Xueyong Shen, and Lina Wang. 2021. "Mechanosensitive TRPV4 Channel-Induced Extracellular ATP Accumulation at the Acupoint Mediates Acupuncture Analgesia of Ankle Arthritis in Rats" Life 11, no. 6: 513. https://doi.org/10.3390/life11060513
APA StyleZheng, Y., Zuo, W., Shen, D., Cui, K., Huang, M., Zhang, D., Shen, X., & Wang, L. (2021). Mechanosensitive TRPV4 Channel-Induced Extracellular ATP Accumulation at the Acupoint Mediates Acupuncture Analgesia of Ankle Arthritis in Rats. Life, 11(6), 513. https://doi.org/10.3390/life11060513