
The Biomechanics of Cartilage—An Overview





Abstract
1. Introduction
2. Anatomy of Hyaline Articular Cartilage
2.1. General Composition
2.2. Chondrocytes
2.3. Zonal Regions
2.4. Collagens
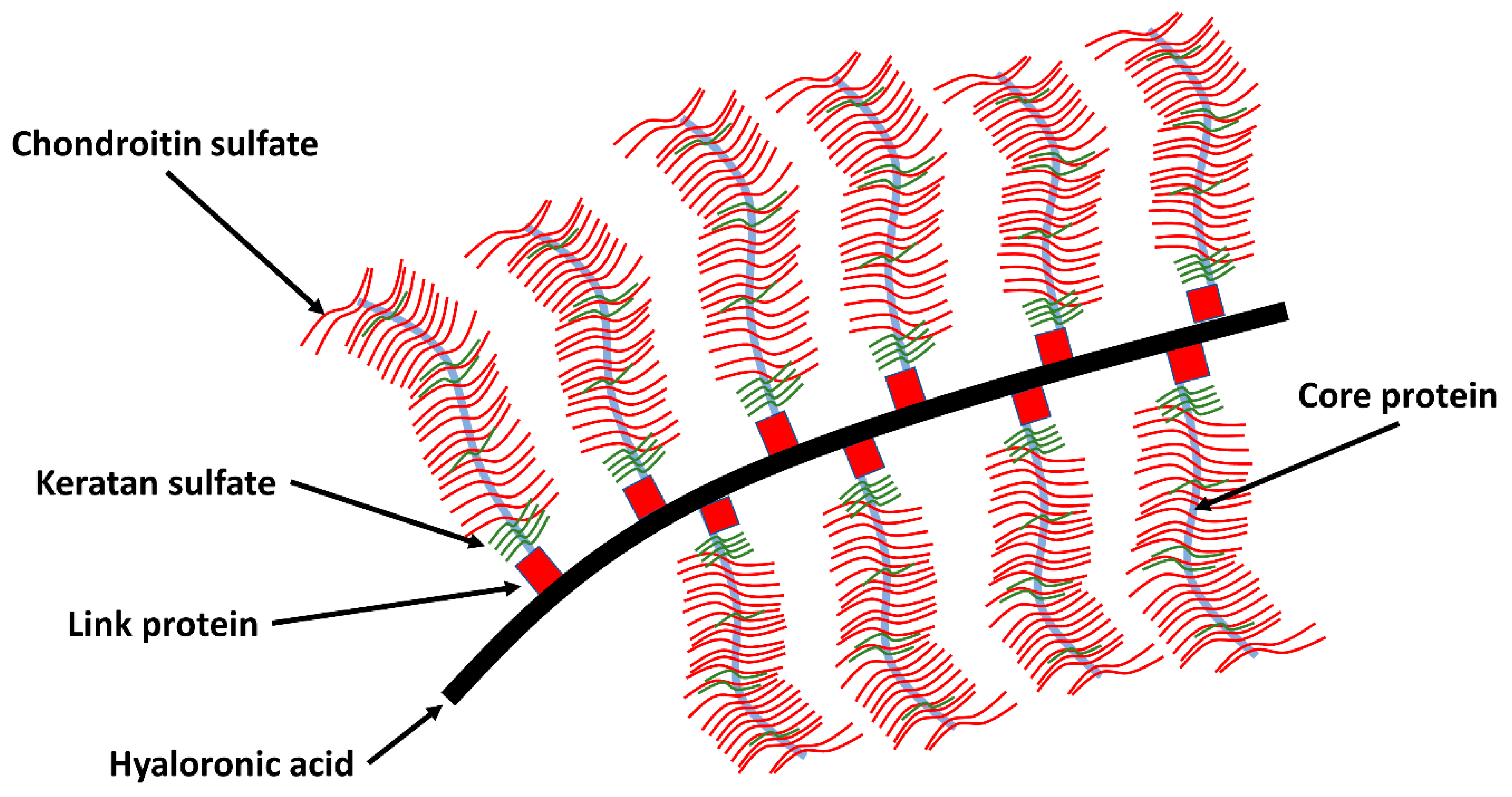
2.5. Protein Network
3. Biomechanics of Articular Cartilage
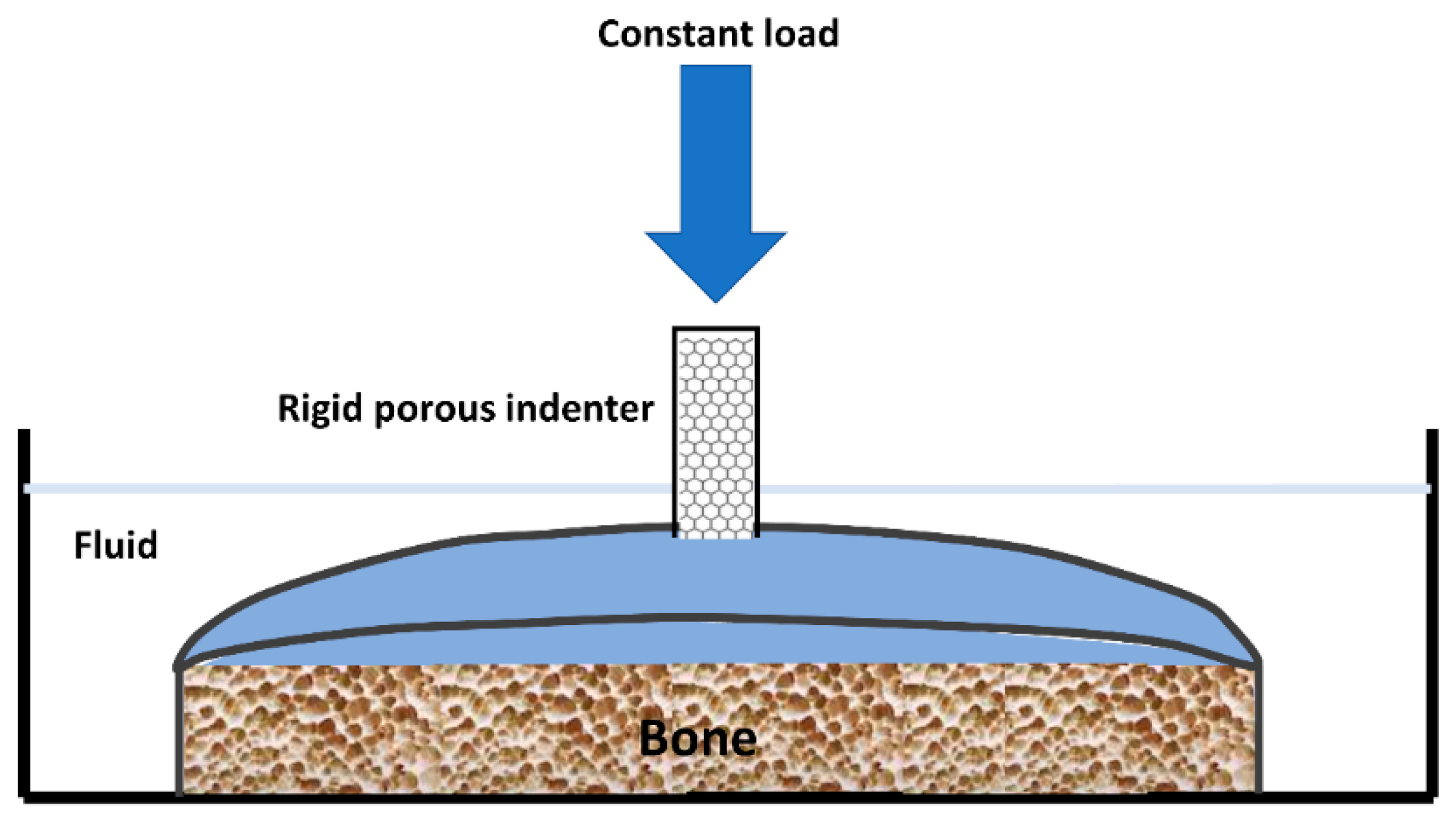
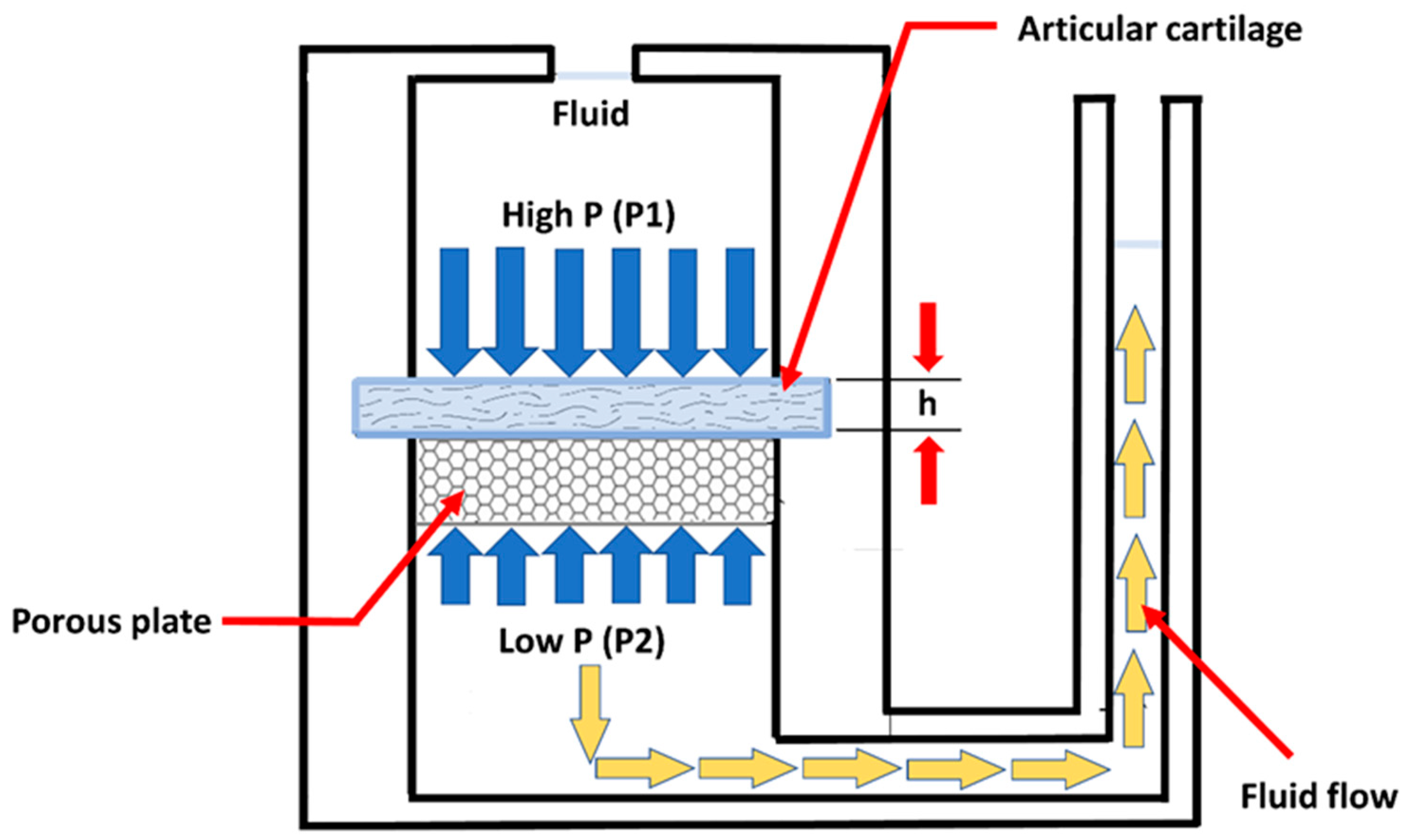
4. Biomechanical Testing of Hyaline Cartilage
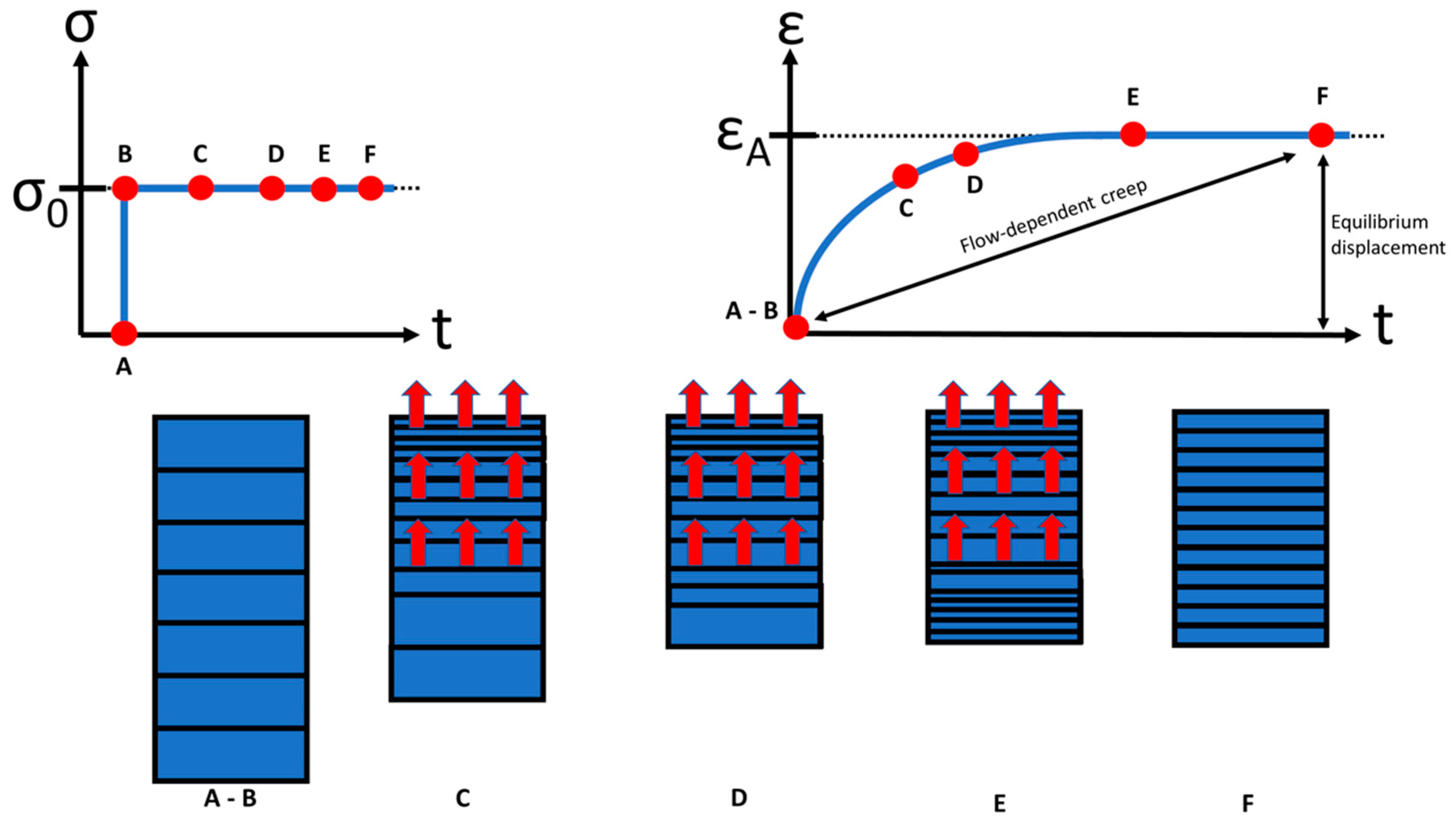
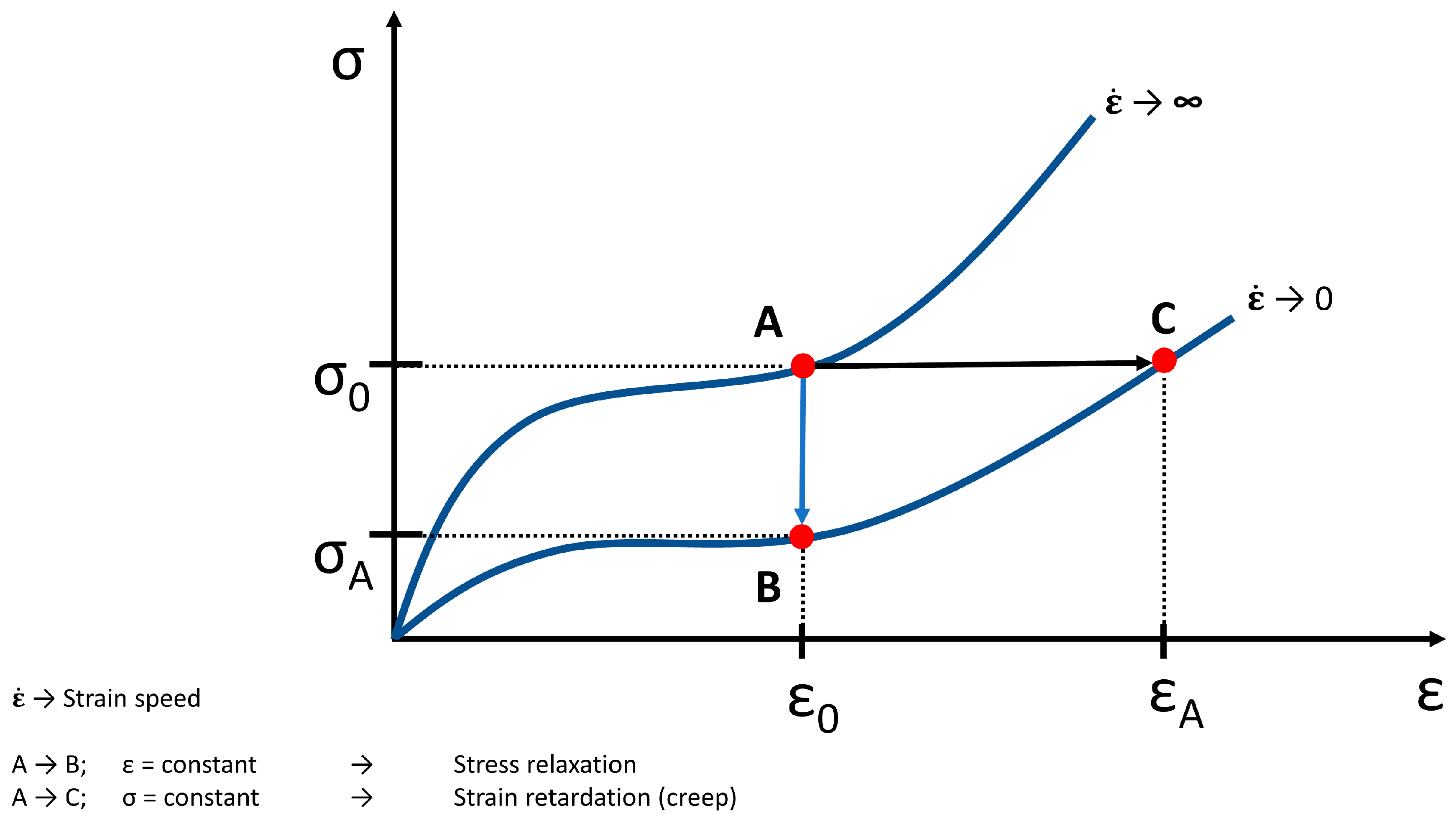
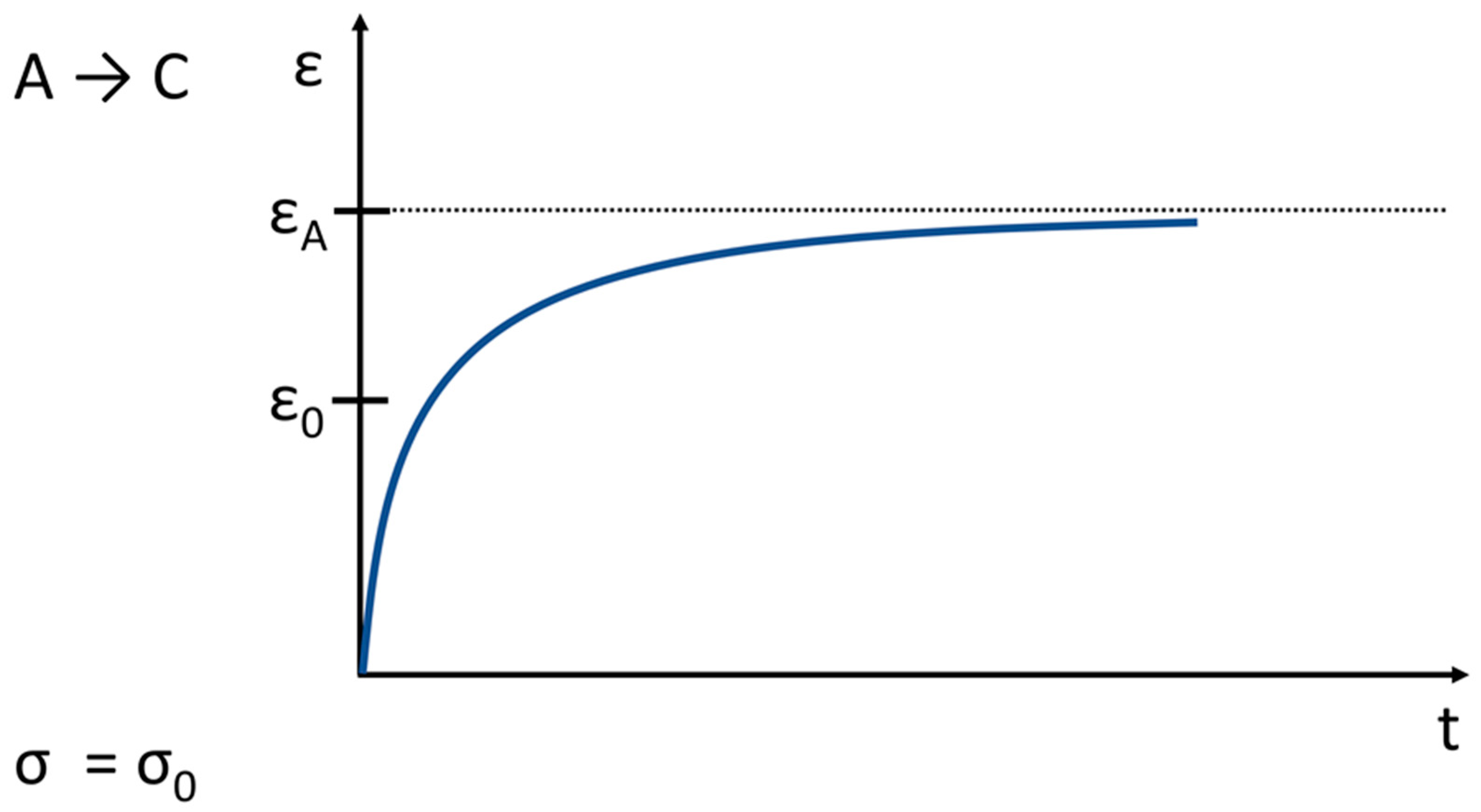
5. Creep and Relaxation of Soft Tissue
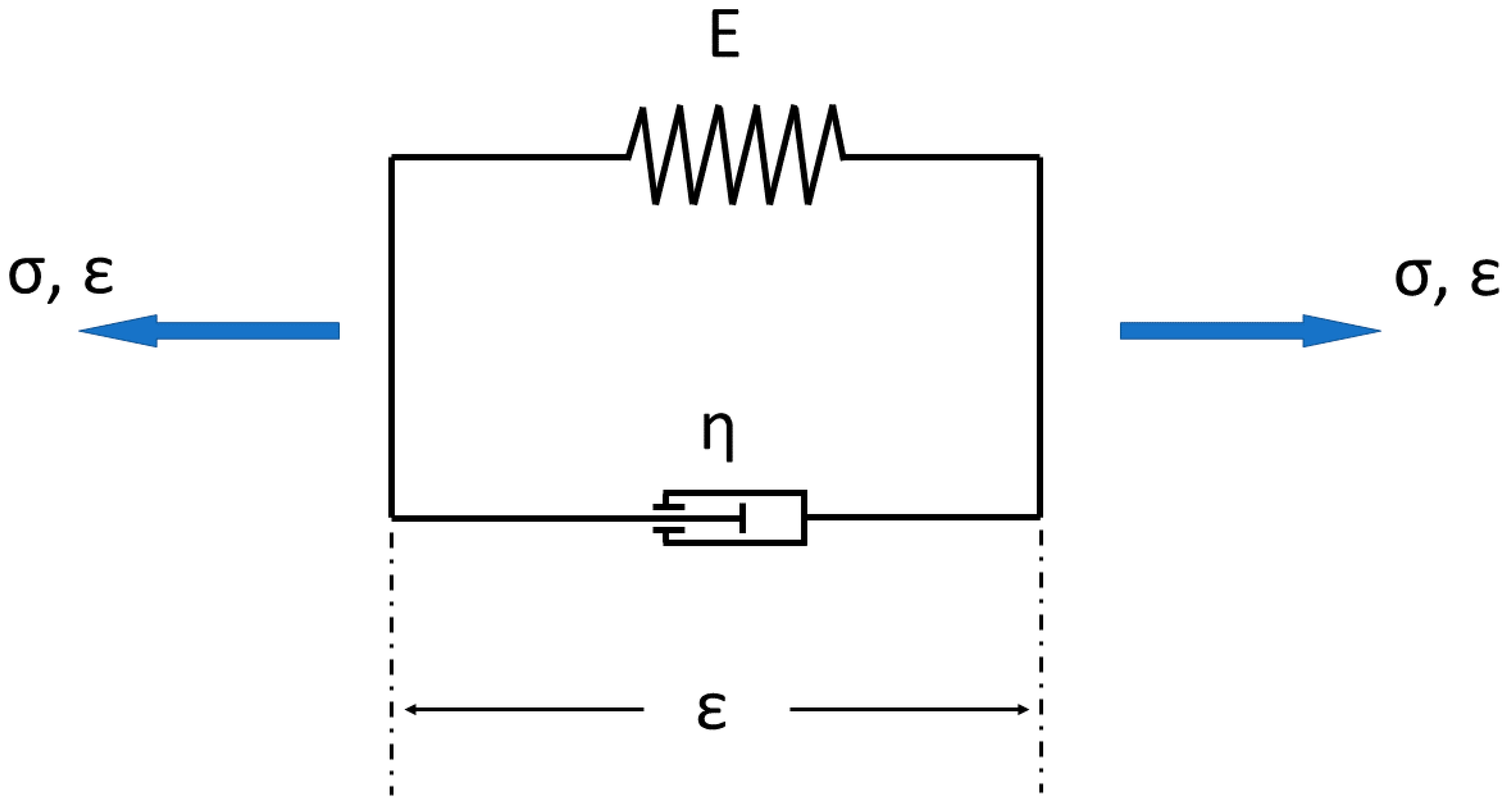
6. Modeling of Hyaline Articular Cartilage
6.1. General Theoretical and Technical Approximation
6.2. Specific Models of Articular Cartilage
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weber, J.F.; Perez, R.; Waldman, S.D. Mechanobioreactors for Cartilage Tissue Engineering. Methods Mol. Biol. 2015, 1340, 203–219. [Google Scholar]
- Buckwalter, J.A.; Mankin, H.J.; Grodzinsky, A.J. Articular cartilage and osteoarthritis. Instr. Course Lect. Am. Acad. Orthop. Surg. 2005, 54, 465. [Google Scholar]
- Nachtsheim, J.; Dursun, G.; Markert, B.; Stoffel, M. Chondrocyte colonisation of a tissue-engineered cartilage substitute under a mechanical stimulus. Med. Eng. Phys. 2019, 74, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Buckwalter, J.A. (Ed.) Orthopaedic Basic Science: Biology and Biomechanics of the Musculoskeletal System, 2nd ed.; American Academy of Orthopaedic Surgeons: Rosemont, IL, USA, 2000. [Google Scholar]
- Lu, X.L.; van Mow, C. Biomechanics of articular cartilage and determination of material properties. Med. Sci. Sports Exerc. 2008, 40, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.P.; Foster, R.J.; Mow, V.C. Composition and Dynamics of Articular Cartilage: Structure, Function, and Maintaining Healthy State. J. Orthop. Sports Phys. Ther. 1998, 28, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Vina, E.R.; Kwoh, C.K. Epidemiology of osteoarthritis: Literature update. Curr. Opin. Rheumatol. 2018, 30, 160–167. [Google Scholar] [CrossRef]
- Syed, M.A.; Azim, S.R.; Baig, M. Frequency of orthopedic problems among patients attending an orthopedic outpatient department: A retrospective analysis of 23,495 cases. Ann. Saudi Med. 2019, 39, 172–177. [Google Scholar] [CrossRef]
- Kingsbury, S.R.; Gross, H.J.; Isherwood, G.; Conaghan, P.G. Osteoarthritis in Europe: Impact on health status, work productivity and use of pharmacotherapies in five European countries. Rheumatology 2014, 53, 937–947. [Google Scholar] [CrossRef]
- Primorac, D.; Molnar, V.; Rod, E.; Jeleč, Ž.; Čukelj, F.; Matišić, V.; Vrdoljak, T.; Hudetz, D.; Hajsok, H.; Borić, I. Knee Osteoarthritis: A Review of Pathogenesis and State-Of-The-Art Non-Operative Therapeutic Considerations. Genes 2020, 11, 854. [Google Scholar] [CrossRef]
- Glyn-Jones, S.; Palmer, A.J.R.; Agricola, R.; Price, A.J.; Vincent, T.L.; Weinans, H.; Carr, A.J. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef]
- Little, C.J.; Bawolin, N.K.; Chen, X. Mechanical Properties of Natural Cartilage and Tissue-Engineered Constructs. Tissue Eng. Part B Rev. 2011, 17, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, M.-F.; Önnerfjord, P.; Bolognesi, M.P.; Easley, M.E.; Kraus, V.B. Analysis of “old” proteins unmasks dynamic gradient of cartilage turnover in human limbs. Sci. Adv. 2019, 5, eaax3203. [Google Scholar] [CrossRef]
- Migliorini, F.; Tingart, M.; Maffulli, N. Progress with stem cell therapies for tendon tissue regeneration. Expert Opin. Biol. Ther. 2020, 20, 1373–1379. [Google Scholar] [CrossRef] [PubMed]
- Mansour, J.M. Biomechanics of cartilage. Kinesiol. Mech. Pathomech. Hum. Mov. 2003, 2, 69–83. [Google Scholar]
- Martinek, V. Anatomie und Pathophysiologie des hyalinen Knorpels. Dtsch. Z. Sportmed. 2003, 54, 166–170. [Google Scholar]
- Nebelung, S.; Post, M.; Raith, S.; Fischer, H.; Knobe, M.; Braun, B.; Prescher, A.; Tingart, M.; Thüring, J.; Bruners, P.; et al. Functional in situ assessment of human articular cartilage using MRI: A whole-knee joint loading device. Biomech. Model. Mechanobiol. 2017, 16, 1971–1986. [Google Scholar] [CrossRef]
- Erggelet, C.; Steinwachs, M. Gelenkknorpeldefekte; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Shepherd, D.E.T.; Seedhom, B.B. Thickness of human articular cartilage in joints of the lower limb. Ann. Rheum. Dis. 1999, 58, 27–34. [Google Scholar] [CrossRef]
- Wayne, J.S.; Brodrick, C.W.; Mukherjee, N. Measurement of articular cartilage thickness in the articulated knee. Ann. Biomed. Eng. 1998, 26, 96–102. [Google Scholar] [CrossRef]
- Cohen, Z.A.; McCarthy, D.M.; Kwak, S.D.; Legrand, P.; Fogarasi, F.; Ciaccio, E.J.; Ateshian, G.A. Knee cartilage topography, thickness, and contact areas from MRI: In-vitro calibration and in-vivo measurements. Osteoarthr. Cartil. 1999, 7, 95–109. [Google Scholar] [CrossRef]
- Grässel, S.; Aszodi, A. (Eds.) Cartilage: Volume 1: Physiology and Development, 1st ed.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar]
- Klika, V.; Gaffney, E.A.; Chen, Y.-C.; Brown, C.P. An overview of multiphase cartilage mechanical modelling and its role in understanding function and pathology. J. Mech. Behav. Biomed. Mater. 2016, 62, 139–157. [Google Scholar] [CrossRef]
- Fox, A.J.S.; Bedi, A.; Rodeo, S.A. The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health A Multidiscip. Approach 2009, 1, 461–468. [Google Scholar]
- Buckwalter, J.A.; Mow, V.C.; Ratcliffe, A. Restoration of Injured or Degenerated Articular Cartilage. J. Am. Acad. Orthop. Surg. 1994, 2, 192–201. [Google Scholar] [CrossRef]
- Doran, P.M. (Ed.) Cartilage Tissue Engineering: Methods and Protocols; Humana Pr: New York, NY, USA, 2015. [Google Scholar]
- O’Connor, K.M. Unweighting Accelerates Tidemark Advancement in Articular Cartilage at the Knee Joint of Rats. J. Bone Miner. Res. 1997, 12, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Mankin, H.J. Articular cartilage, cartilage injury, and osteoarthritis. Patellofemoral Jt. 1993, 1993, 13–45. [Google Scholar]
- Redler, I.; Mow, V.C.; Zimny, M.L.; Mansell, J. The ultrastructure and biomechanical significance of the tidemark of articular cartilage. Clin. Orthop. Relat. Res. 1975, 112, 357–362. [Google Scholar] [CrossRef]
- Jeffery, A.K.; Blunn, G.W.; Archer, C.W.; Bentley, G. Three-dimensional collagen architecture in bovine articular cartilage. J. Bone Jt. Surg. 1991, 73, 795–801. [Google Scholar] [CrossRef]
- Chen, F.H.; Rousche, K.T.; Tuan, R.S. Technology Insight: Adult stem cells in cartilage regeneration and tissue engineering. Nat. Clin. Pr. Rheumatol. 2006, 2, 373–382. [Google Scholar] [CrossRef]
- Muir, H. The chondrocyte, architect of cartilage. Biomechanics, structure, function and molecular biology of cartilage matrix macromolecules. BioEssays 1995, 17, 1039–1048. [Google Scholar] [CrossRef]
- Newman, A.P. Articular Cartilage Repair. Am. J. Sports Med. 1998, 26, 309–324. [Google Scholar] [CrossRef]
- Hurschler, C.; Abedian, R. Möglichkeiten der biomechanischen Charakterisierung von Knorpelgewebe. Der Orthopäde 2013, 42, 232–241. [Google Scholar] [CrossRef]
- Mow, V.C.; Huiskes, R. Basic Orthopaedic Biomechanics & Mechano-Biology, 3rd ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2005. [Google Scholar]
- Lai, W.M.; Hou, J.S.; Mow, V.C. A Triphasic Theory for the Swelling and Deformation Behaviors of Articular Cartilage. J. Biomech. Eng. 1991, 113, 245–258. [Google Scholar] [CrossRef]
- van Mow, C. (Ed.) Biomechanics of Diarthrodial Joints: Volume I; Springer: New York, NY, USA, 1990. [Google Scholar]
- Mow, V.C.; Kuei, S.C.; Lai, W.M.; Armstrong, C.G. Biphasic creep and stress relaxation of articular cartilage in compression? Theory and experiments. J. Biomech. Eng. 1980, 102, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Athanasiou, K.A.; Rosenwasser, M.P.; Buckwalter, J.A.; Malinin, T.I.; Mow, V.C. Interspecies comparisons of in situ intrinsic mechanical properties of distal femoral cartilage. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 1991, 9, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, C.G.; Lai, W.M.; Mow, V.C. An Analysis of the Unconfined Compression of Articular Cartilage. J. Biomech. Eng. 1984, 106, 165–173. [Google Scholar] [CrossRef]
- Li, L.; Shirazi-Adl, A.; Buschmann, M.D. Investigation of mechanical behavior of articular cartilage by fibril reinforced poroelastic models. Biorheology 2003, 40, 227–233. [Google Scholar] [PubMed]
- Berteau, J.-P.; Oyen, M.; Shefelbine, S.J. Permeability and shear modulus of articular cartilage in growing mice. Biomech. Model. Mechanobiol. 2015, 15, 205–212. [Google Scholar] [CrossRef]
- Maroudas, A. Physicochemical Properties of Cartilage in the Light of Ion Exchange Theory. Biophys. J. 1968, 8, 575–595. [Google Scholar] [CrossRef]
- Maroudas, A.; Bullough, P. Permeability of Articular Cartilage. Nat. Cell Biol. 1968, 219, 1260–1261. [Google Scholar]
- Maroudas, A.; Bullough, P.; Swanson, S.A.; Freeman, M.A. The permeability of articular cartilage. J. Bone Jt. Surg. 1968, 50, 166–177. [Google Scholar] [CrossRef]
- Mansour, J.M.; Mow, V.C. The permeability of articular cartilage under compressive strain and at high pressures. J. Bone Jt. Surg. Am. Vol. 1976, 58, 509–516. [Google Scholar] [CrossRef]
- Donnan, F.G. The Theory of Membrane Equilibria. Chem. Rev. 1924, 1, 73–90. [Google Scholar] [CrossRef]
- Lehneis, H.R. Bone and cartilage-structure and biomechanics. Orthot. Prosthet. 1987, 41, 37–40. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chondrocytes | 1–10% |
| Water | 70–80% |
| Collagen | 12–14% |
| - Type II | 10–12% |
| - Type IX | ~1% |
| - Type XI | ~1% |
| Proteoglycans | 7–9% |
| - Hyaluronic acid—proteoglycan—aggregates | 6–8% |
| - Other proteoglycans | ~1% |
| Mineralic materials | <4% |
| Matrix proteins | <1% |
| Zone | Name | Description | Functional Behaviour |
|---|---|---|---|
| Zone I | superficial (tangential) zone | The superficial zone makes up approximately 10% to 20% of AC thickness. The collagen fibers of this zone are packed tightly and aligned parallel to the articular surface. The chondrocytes in the superficial zone are flatter and smaller and generally have a greater density than that of the cells deeper in the matrix. It protects deeper layers from shear stresses. The zona superficialis has most of the water and very little proteoglycans. In general, the cells are not very active, which means that there is little wear and tear. The zone can also be used as a barrier against large molecules, for example antibodies. | The superficial zona is responsible for the behavior of the cartilage under stress. It deforms more strongly and is therefore less rigid than the underlying zones (Guili.k et al., 1995b). If this zone is disturbed tissue permeability increases, leading to greater fluid exchange of the cartilage with its surroundings and during compression, this leads to greater mechanical stress on the macromolecular network. |
| Zone II | middle (transitional) zone, | The middle zone represents 40% to 60% of the total AC volume, and it contains proteoglycans and thicker collagen fibrils. In this layer, the collagen is organized obliquely. Chondrocytes are spherical and at low density. | Functionally, the middle zone is the first line of resistance against compressive forces. |
| Zone III | deep zone | The deep zone represents approximately 30% of the AC volume. The deep zone contains the largest diameter collagen fibrils in a radial disposition, the highest proteoglycan content, and the lowest water concentration. The chondrocytes are typically arranged in a columnar orientation, parallel to the collagen fibers and perpendicular to the joint line. | It is responsible for providing the greatest resistance to compressive forces, given that collagen fibrils are arranged perpendicular to the articular surface. This creates a arcade formation. These arcades are to be created by the attempt to transfer the initial fibril network to a higher order, whereby the arcadic structure supports the overlying load. |
| Interface | tide mark | The tide mark distinguishes the deep zone from the calcified cartilage. This irregularly salty layer lies between the Zonae radiata and calcificata and separates the lime-poor from the lime-rich cartilage. O’Connor describes this area as the mineralization front of the cartilage [27]. According to Mankin, this layer arises by causing the collagen bundles to twist before going deeper to penetrate the zona calcificata and the bone tissue [28]. | It consists of a band of fibrils attached to the collagen fibers which are anchored in the lime-poor layer, and thus prevent them from tearing off cartilage from bone [29]. |
| Zone IV | calcified zone | The calcified zone plays an integral role in securing the cartilage to bone by anchoring the collagen fibrils of the deep zone to subchondral bone. In this zone, the cell population is scarce and chondrocytes are hypertrophic. | The calcified zone has numerous protrusions, hollows, and interlacing, which gives an excellent resistance to shear forces to prevent the cartilage detaching from the underlying bone. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eschweiler, J.; Horn, N.; Rath, B.; Betsch, M.; Baroncini, A.; Tingart, M.; Migliorini, F. The Biomechanics of Cartilage—An Overview. Life 2021, 11, 302. https://doi.org/10.3390/life11040302
Eschweiler J, Horn N, Rath B, Betsch M, Baroncini A, Tingart M, Migliorini F. The Biomechanics of Cartilage—An Overview. Life. 2021; 11(4):302. https://doi.org/10.3390/life11040302
Chicago/Turabian StyleEschweiler, Joerg, Nils Horn, Bjoern Rath, Marcel Betsch, Alice Baroncini, Markus Tingart, and Filippo Migliorini. 2021. "The Biomechanics of Cartilage—An Overview" Life 11, no. 4: 302. https://doi.org/10.3390/life11040302
APA StyleEschweiler, J., Horn, N., Rath, B., Betsch, M., Baroncini, A., Tingart, M., & Migliorini, F. (2021). The Biomechanics of Cartilage—An Overview. Life, 11(4), 302. https://doi.org/10.3390/life11040302