
Usefulness of Microbiome for Forensic Geolocation: A Review

 ,
,
Abstract
:1. Introduction
2. Forensic Microbiome as a Tool for Geolocation
2.1. Soil and Surface Microbiome
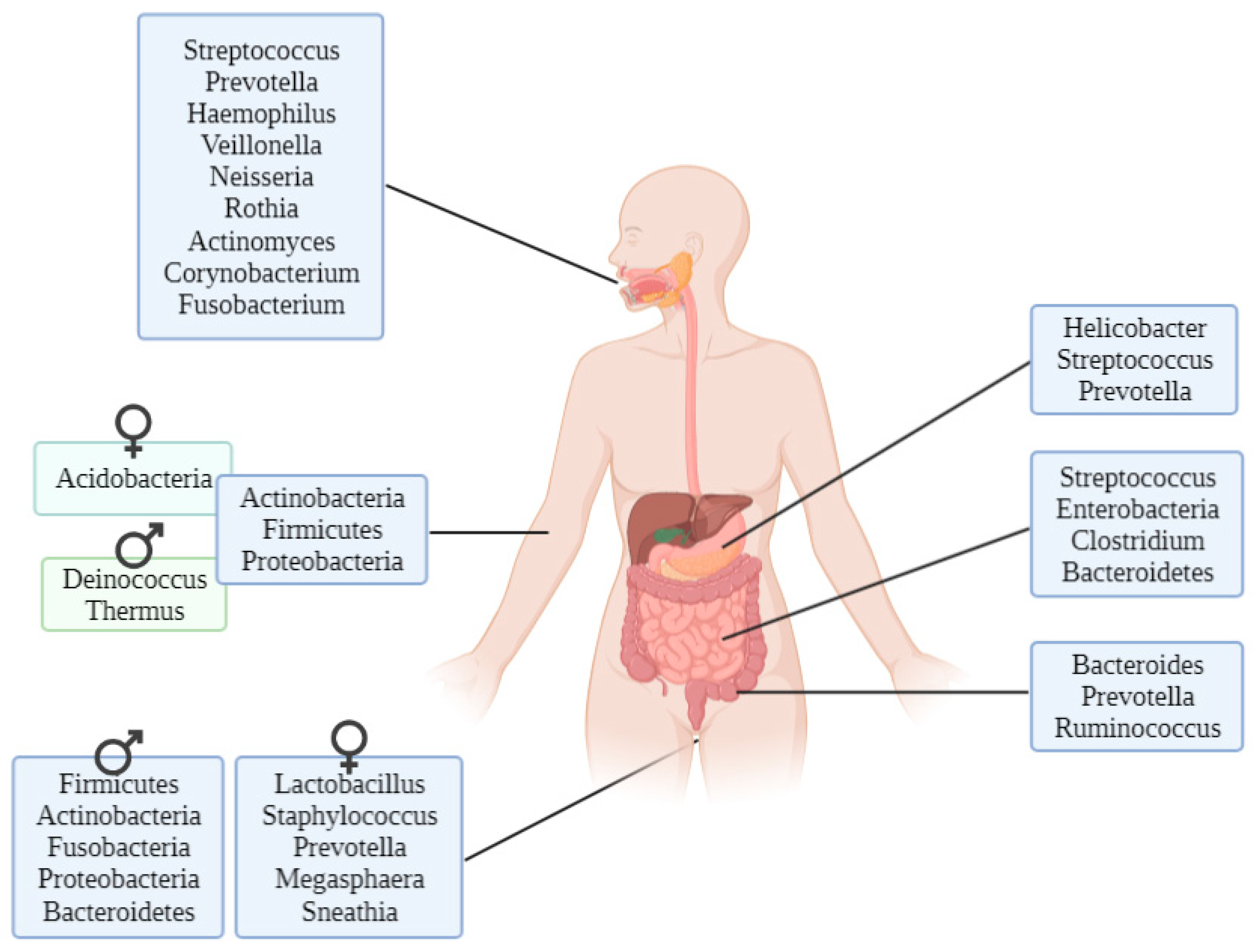
2.2. In Vivo Microbiome
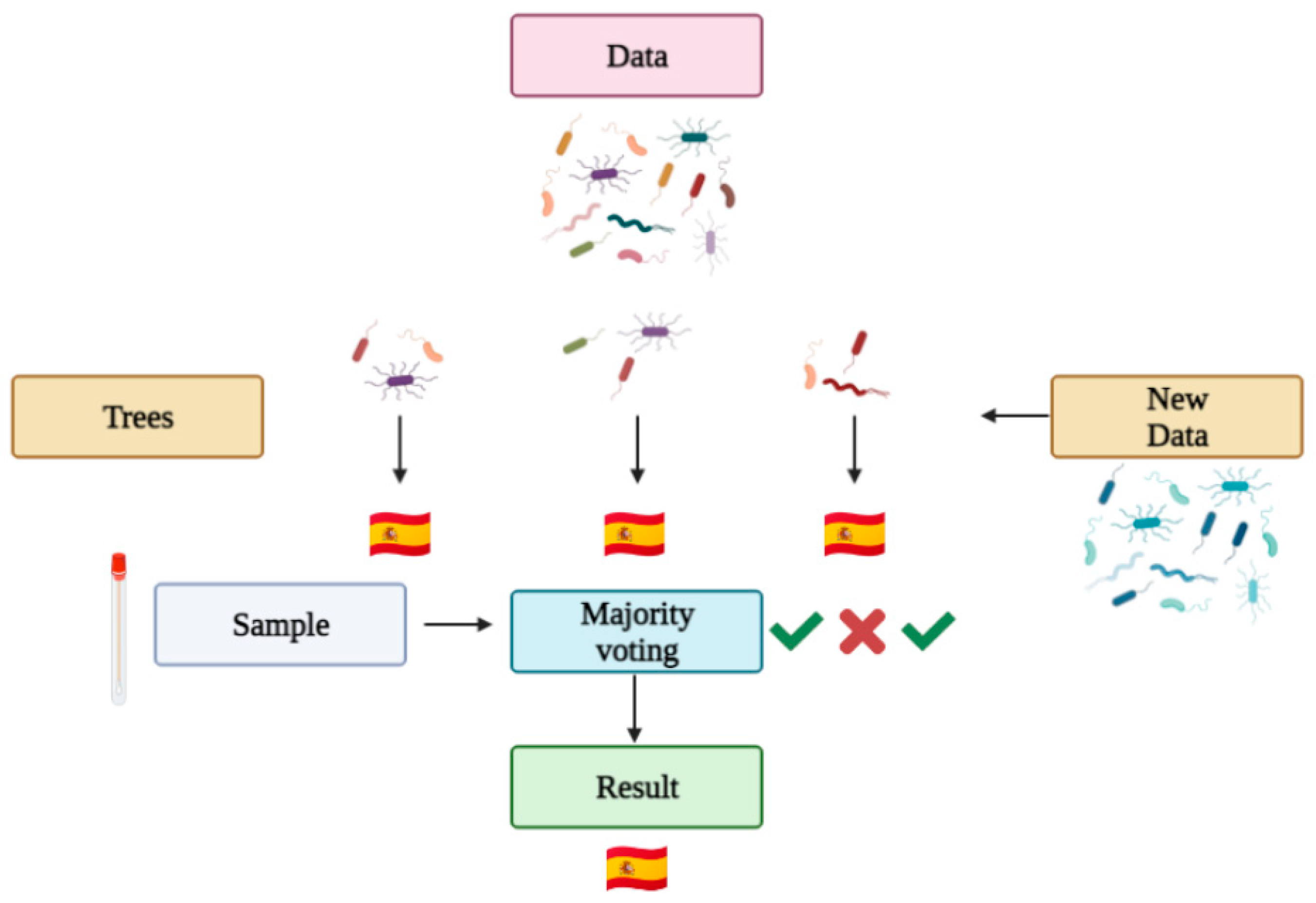
2.3. Machine Learning and Geolocation
2.4. Protocols
2.4.1. Sampling
2.4.2. DNA Extraction
2.4.3. Sequencing
2.5. Current Applications
2.5.1. DNA-Prokids
2.5.2. Skeletal Remains
3. Challenges and Limitations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kambouris, M.E.; Velegraki, A.; Patrinos, G.P.; Zerva, L. Introduction: The Microbiome as a Concept: Vogue or Necessity? In Microbiomics; Kambouris, M.E., Velegraki, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–4. ISBN 978-0-12-816664-2. [Google Scholar]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef] [PubMed]
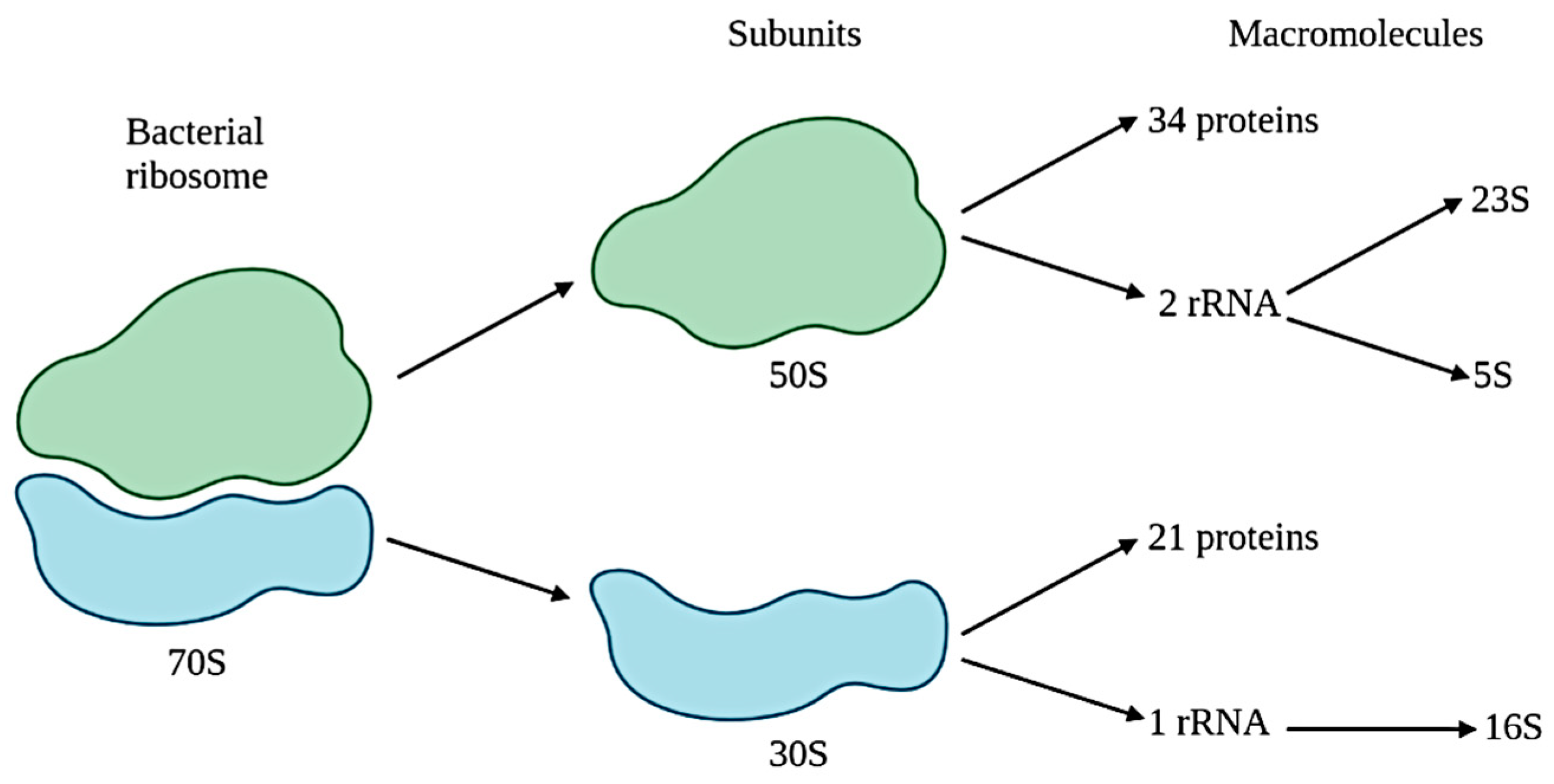
- Fox, G.E.; Magrum, L.J.; Balch, W.E.; Wolfe, R.S.; Woese, C.R. Classification of methanogenic bacteria by 16S ribosomal RNA characterization. Proc. Natl. Acad. Sci. USA 1977, 74, 4537–4541. [Google Scholar] [CrossRef] [Green Version]
- Jan-da, J.M. Taxonomic Classification of Bacteria. In Practical Handbook of Microbiology; Green, L.H., Goldman, E., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 161–167. ISBN 9780367567637. [Google Scholar]
- Schleifer, K.H. Classification of Bacteria and Archaea: Past, present and future. Syst. Appl. Microbiol. 2009, 32, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Willey, J.M.; Sherwood, L.M.; Woolverton, C.J. Prescott’s Principles of Microbiology; McGraw-Hill: New York, NY, USA, 2009; ISBN 978-0-07-337523-6. [Google Scholar]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Marchesi, J.R. The Human Microbiota and Microbiome; Advances in Molecular and Cellular Microbiology, CABI: Wallingford, UK, 2014; ISBN 9781780640495. [Google Scholar]
- Budowle, B.; Schutzer, S.E.; Einseln, A.; Kelley, L.C.; Walsh, A.C.; Smith, J.A.L.; Marrone, B.L.; Robertson, J.; Campos, J. Building Microbial Forensics as a Response to Bioterrorism. Science 2003, 301, 1852–1853. [Google Scholar] [CrossRef] [Green Version]
- Bur-cham, Z.M.; Jordan, H.R. History, current, and future use of microorganisms as physical evidence. In Forensic Microbiology; Carter, D.O., Tomberlin, J.K., Benbow, M.E., Metcalf, J.L., Eds.; Wiley: Chichester, UK, 2017; pp. 25–56. ISBN 9781119062554. [Google Scholar]
- Clarke, T.H.; Gomez, A.; Singh, H.; Nelson, K.E.; Brinkac, L.M. Integrating the microbiome as a resource in the forensics toolkit. Forensic Sci. Int. Genet. 2017, 30, 141–147. [Google Scholar] [CrossRef] [Green Version]
- National Institute of Justice. The Forensic Microbiome: The Invisible Traces We Leave Behind. Available online: https://nij.ojp.gov/topics/articles/forensic-microbiome-invisible-traces-we-leave-behind (accessed on 25 October 2021).
- Robinson, J.M.; Pasternak, Z.; Mason, C.E.; Elhaik, E. Forensic Applications of Microbiomics: A Review. Front. Microbiol. 2021, 11, 3455. [Google Scholar] [CrossRef]
- Thompson, L.R.; Sanders, J.G.; McDonald, D.; Amir, A.; Ladau, J.; Locey, K.J.; Prill, R.J.; Tripathi, A.; Gibbons, S.M.; Ackermann, G.; et al. A communal catalogue reveals Earth’s multiscale microbial diversity. Nature 2017, 551, 457–463. [Google Scholar] [CrossRef] [Green Version]
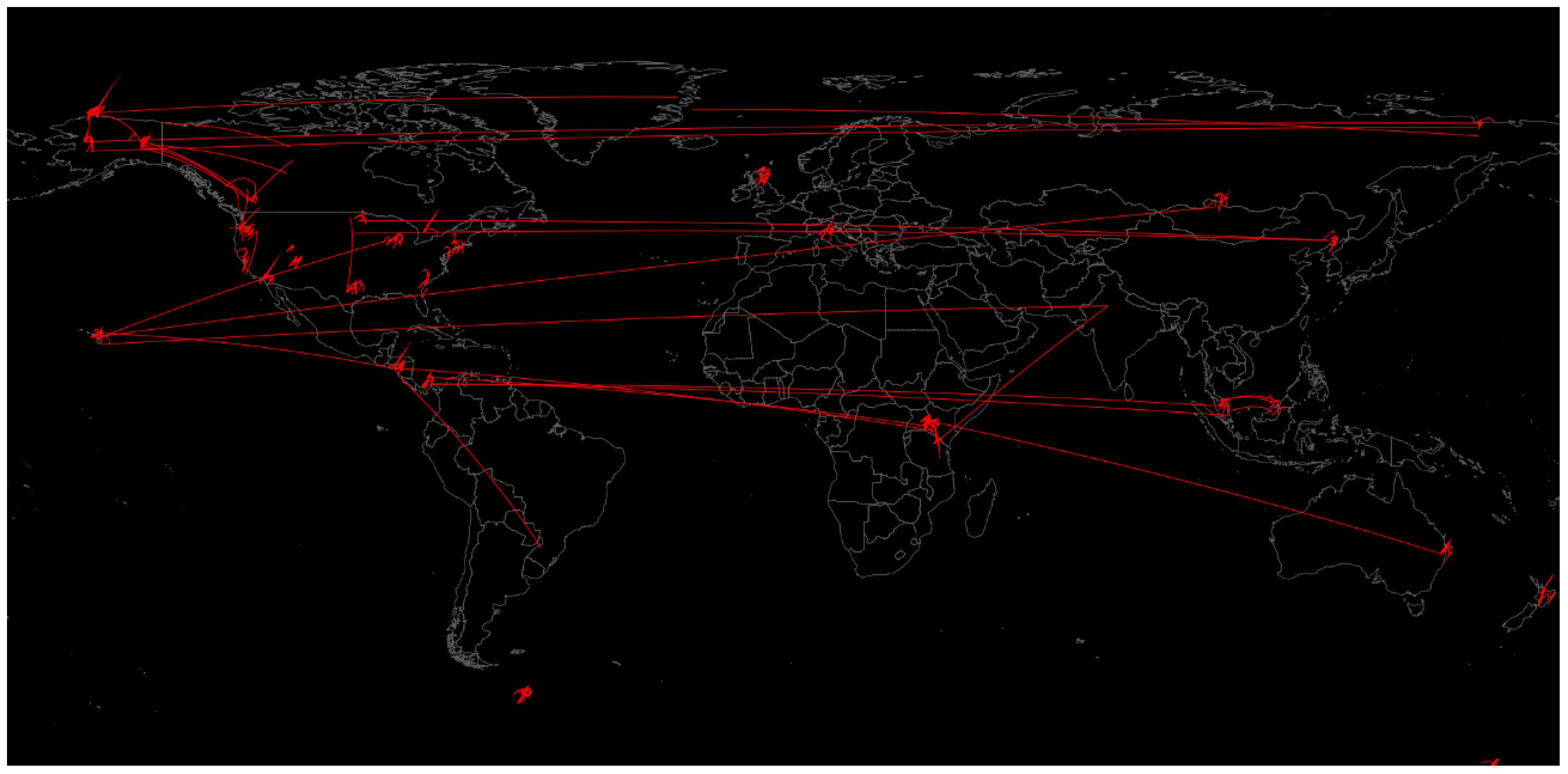
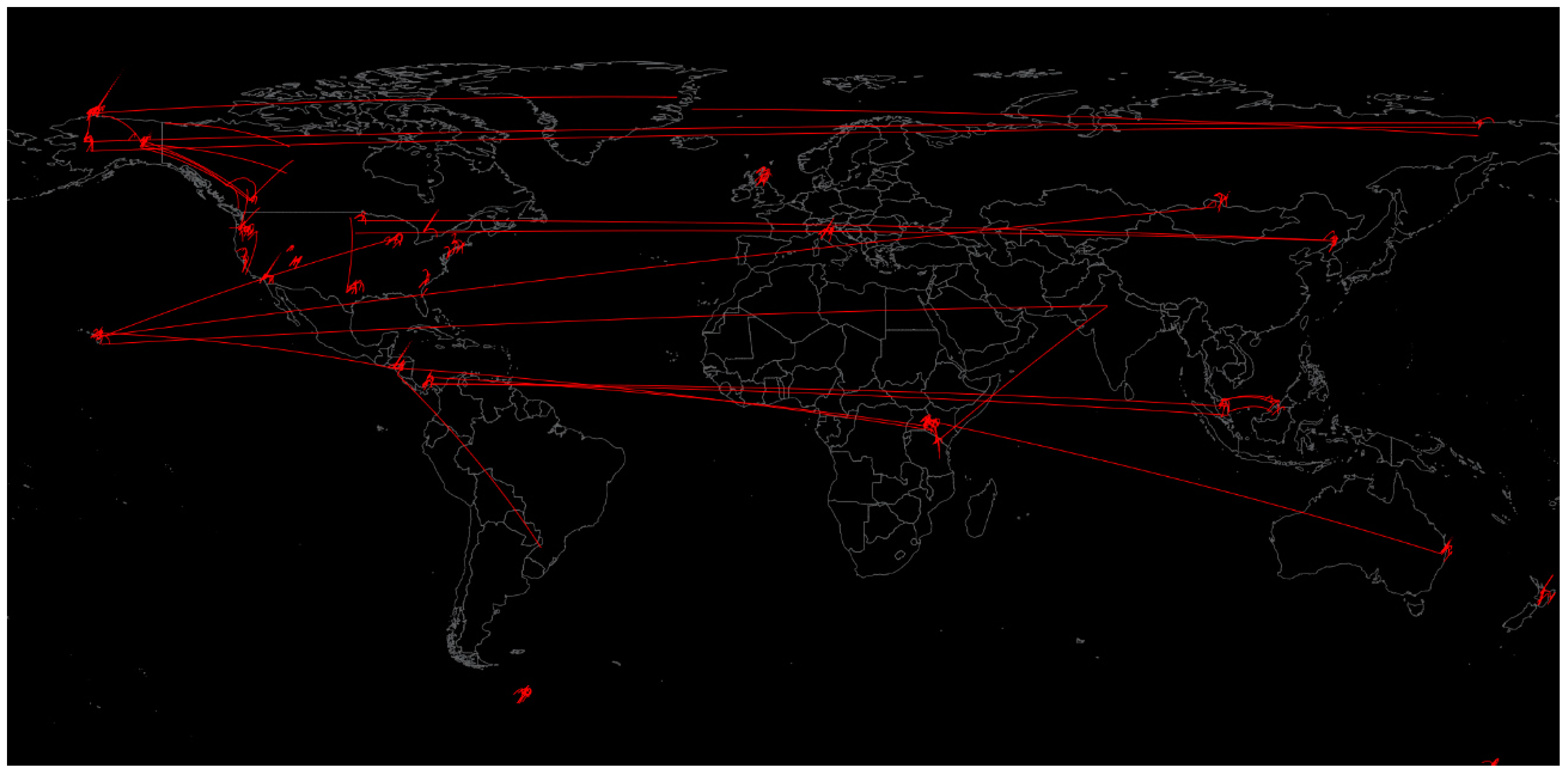
- Afshinnekoo, E.; Meydan, C.; Chowdhury, S.; Jaroudi, D.; Boyer, C.; Bernstein, N.; Maritz, J.M.; Reeves, D.; Gandara, J.; Chhangawala, S.; et al. Geospatial Resolution of Human and Bacterial Diversity with City-Scale Metagenomics. Cell Syst. 2015, 1, 72–87. [Google Scholar] [CrossRef] [Green Version]
- The MetaSUB International Consortium; Mason, C. The Metagenomics and Metadesign of the Subways and Urban Biomes (MetaSUB) International Consortium inaugural meeting report. Microbiome 2016, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Danko, D.; Bezdan, D.; Afshinnekoo, E.; Ahsanuddin, S.; Bhattacharya, C.; Butler, D.J.; Chng, K.R.; DeFilippis, F.; Hecht, J.; Kahles, A.; et al. Global Genetic Cartography of Urban Meta genomes and Anti-Microbial Resistance. bioRxiv 2019, 724526. [Google Scholar] [CrossRef] [Green Version]
- Rosenfeld, J.A.; Reeves, D.; Brugler, M.R.; Narechania, A.; Simon, S.; Durrett, R.; Foox, J.; Shianna, K.; Schatz, M.; Gandara, J.; et al. Genome assembly and geospatial phylogenomics of the bed bug Cimex lectularius. Nat. Commun. 2016, 7, 10164. [Google Scholar] [CrossRef]
- Walker, A.R.; Datta, S. Identification of city specific important bacterial signature for the MetaSUB CAMDA challenge microbiome data. Biol. Direct 2019, 14, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habtom, H.; Pasternak, Z.; Matan, O.; Azulay, C.; Gafny, R.; Jurkevitch, E. Applying microbial biogeography in soil forensics. Forensic Sci. Int. Genet. 2019, 38, 195–203. [Google Scholar] [CrossRef]
- Demanèche, S.; Schauser, L.; Dawson, L.; Franqueville, L.; Simonet, P. Microbial soil community analyses for forensic science: Application to a blind test. Forensic Sci. Int. 2017, 270, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Ryan, F.J. Application of machine learning techniques for creating urban microbial fingerprints. Biol. Direct 2019, 14, 13. [Google Scholar] [CrossRef] [Green Version]
- Young, J.M.; Weyrich, L.S.; Breen, J.; Macdonald, L.; Cooper, A. Predicting the origin of soil evidence: High throughput eukaryote sequencing and MIR spectroscopy applied to a crime scene scenario. Forensic Sci. Int. 2015, 251, 22–31. [Google Scholar] [CrossRef]
- Chase, J.; Fouquier, J.; Zare, M.; Sonderegger, D.; Knight, R.; Kelley, S.T.; Siegel, J.; Caporaso, J.G. Geography and Location Are the Primary Drivers of Office Microbiome Composition. mSystems 2016, 1, e00022-16. [Google Scholar] [CrossRef] [Green Version]
- Lax, S.; Smith, D.P.; Hampton-Marcell, J.; Owens, S.M.; Handley, K.M.; Scott, N.M.; Gibbons, S.M.; Larsen, P.; Shogan, B.D.; Weiss, S.; et al. Longitudinal analysis of microbial interaction between humans and the indoor environment. Science 2014, 345, 1048–1052. [Google Scholar] [CrossRef] [Green Version]
- Zeng, B.; Zhao, J.; Guo, W.; Zhang, S.; Hua, Y.; Tang, J.; Kong, F.; Yang, X.; Fu, L.; Liao, K.; et al. High-Altitude Living Shapes the Skin Microbiome in Humans and Pigs. Front. Microbiol. 2017, 8, 1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, M.G.; Pérez-Cárceles, M.D.; Osuna, E.; Legaz, I. Impact of the Human Microbiome in Forensic Sciences: A Systematic Review. Appl. Environ. Microbiol. 2020, 86, e01451-20. [Google Scholar] [CrossRef]
- Nagasawa, S.; Motani-Saitoh, H.; Inoue, H.; Iwase, H. Geographic diversity of Helicobacter pylori in cadavers: Forensic estimation of geographical origin. Forensic Sci. Int. 2013, 229, 7–12. [Google Scholar] [CrossRef]
- Escobar, J.S.; Klotz, B.; Valdes, B.E.; Agudelo, G.M. The gut microbiota of Colombians differs from that of Americans, Europeans and Asians. BMC Microbiol. 2014, 14, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yooseph, S.; Kirkness, E.F.; Tran, T.M.; Harkins, D.M.; Jones, M.B.; Torralba, M.G.; O’Connell, E.; Nutman, T.B.; Doumbo, S.; Doumbo, O.K.; et al. Stool microbiota composition is associated with the prospective risk of Plasmodium falciparum infection. BMC Genom. 2015, 16, 631. [Google Scholar] [CrossRef] [PubMed]
- Brinkac, L.; Clarke, T.H.; Singh, H.; Greco, C.; Gomez, A.; Torralba, M.G.; Frank, B.; Nelson, K.E. Spatial and Environmental Variation of the Human Hair Microbiota. Sci. Rep. 2018, 8, 9017. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.A.; Worobey, M. Geographical variation of human gut microbial composition. Biol. Lett. 2014, 10, 20131037. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.L.; Xu, Z.Z.; Bouslimani, A.; Dorrestein, P.; Carter, D.O.; Knight, R. Microbiome Tools for Forensic Science. Trends Biotechnol. 2017, 35, 814–823. [Google Scholar] [CrossRef]
- Knights, D.; Costello, E.K.; Knight, R. Supervised classification of human microbiota. FEMS Microbiol. Rev. 2011, 35, 343–359. [Google Scholar] [CrossRef] [Green Version]
- Pavlov, Y.L. Random Forests; De Gruyter: Berlin, Germany, 2019; Volume 45, ISBN 9783110941975. [Google Scholar]
- Hengl, T.; Nussbaum, M.; Wright, M.N.; Heuvelink, G.B.M.; Gräler, B. Random forest as a generic framework for predictive modeling of spatial and spatio-temporal variables. PeerJ 2018, 6, e5518. [Google Scholar] [CrossRef] [Green Version]
- Grantham, N.S.; Reich, B.J.; Laber, E.B.; Pacifici, K.; Dunn, R.R.; Fierer, N.; Gebert, M.; Allwood, J.S.; Faith, S.A. Global forensic geolocation with deep neural networks. J. R. Stat. Soc. Ser. C Appl. Stat. 2020, 69, 909–929. [Google Scholar] [CrossRef]
- Bjerre, R.D.; Hugerth, L.W.; Boulund, F.; Seifert, M.; Johansen, J.D.; Engstrand, L. Effects of sampling strategy and DNA extraction on human skin microbiome investigations. Sci. Rep. 2019, 9, 17287. [Google Scholar] [CrossRef] [Green Version]
- Thompson, L.; Ackermann, G.; Humphrey, G.; Gilbert, J.; Jansson, J.; Knight, R. EMP Sample Submission Guide v1. protocols.io 2018. Available online: https://www.protocols.io/view/emp-sample-submission-guide-pfqdjmw (accessed on 25 October 2021).
- Marotz, C.; Amir, A.; Humphrey, G.; Gaffney, J.; Gogul, G.; Knight, R. DNA extraction for streamlined metagenomics of diverse environmental samples. Biotechniques 2017, 62, 290–293. [Google Scholar] [CrossRef] [Green Version]
- Minich, J.J.; Zhu, Q.; Janssen, S.; Hendrickson, R.; Amir, A.; Vetter, R.; Hyde, J.; Doty, M.M.; Stillwell, K.; Benardini, J.; et al. KatharoSeq Enables High-Throughput Microbiome Analysis from Low-Biomass Samples. mSystems 2018, 3, e00218-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandrini, F.; Brenciani, A.; Fioriti, S.; Melchionda, F.; Mingoia, M.; Morroni, G.; Tagliabracci, A. Validation of a universal DNA extraction method for human and microbiAL DNA analysis. Forensic Sci. Int. Genet. Suppl. Ser. 2019, 7, 256–258. [Google Scholar] [CrossRef]
- Applied Biosystems Mag MAXTM Microbiome Ultra Nucleic Acid Isolation Kit. Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https://assets.thermofisher.com/TFS-Assets%2FLSG%2Fmanuals%2FMAN0018070_MagMAXMicrobiomeNuclAcidIsolatKit_SoilSalivaUrine_Automated_UG.pdf (accessed on 25 October 2021).
- Invitrogen Pure Link TM Microbiome DNA Purification Kit. Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https://assets.thermofisher.com/TFS-Assets%2FLSG%2Fmanuals%2FMAN0014331_PureLinkMicrobiome_Soil_UG.pdf (accessed on 25 October 2021).
- QIAGEN QIAamp DNA Microbiome Kit. Available online: https://www.qiagen.com/us/products/discovery-and-translational-research/dna-rna-purification/dna-purification/microbial-dna/qiaamp-dna-microbiome-kit/ (accessed on 25 October 2021).
- QIAGENMO-BIO’s Power Mag Soil DNA Isolation Kit EPH and Book. Available online: https://www.qiagen.com/us/resources/resourcedetail?id=faaba49d-9d10-48cf-a461-01d5302fea49&lang=en (accessed on 25 October 2021).
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Amaral-Zettler, L.A.; McCliment, E.A.; Ducklow, H.W.; Huse, S.M. A Method for Studying Protistan Diversity Using Massively Parallel Sequencing of V9 Hypervariable Regions of Small-Subunit Ribosomal RNA Genes. PLoS ONE 2009, 4, e6372. [Google Scholar] [CrossRef]
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.M.; Breiner, H.-W.; Richards, T.A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Mol. Ecol. 2010, 19, 21–31. [Google Scholar] [CrossRef]
- Emmons, A.L.; Mundorff, A.Z.; Keenan, S.W.; Davoren, J.; Andronowski, J.; Carter, D.O.; Debruyn, J.M. Characterizing the postmortem human bone microbiome from surface-decomposed remains. PLoS ONE 2020, 15, e0218636. [Google Scholar] [CrossRef] [PubMed]
- Lorente, J.A.; Saiz, M.; Haarkötter, C.; Robles-Fernandez, I.; Alvarez-Cubero, M.J.; Galvez, X.; Martinez-Gonzalez, L.J.; Lorente-Remon, B.; Alvarez, J.C. Genetic identification against traffic in human beings. Wiley Interdiscip. Rev. Forensic Sci. 2021, 3, e1392. [Google Scholar] [CrossRef]
- Turner-Walker, G. Light at the end of the tunnels? The origins of microbial bioerosion in mineralised collagen. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 529, 24–38. [Google Scholar] [CrossRef]
- Damann, F.E.; Jans, M.M.E. Microbes, anthropology, and bones. In Forensic Microbiology; Carter, D.O., Tomberlin, J.K., Benbow, M.E., Metcalf, J.L., Eds.; Wiley: Chichester, UK, 2017; pp. 312–327. ISBN 9781119062554. [Google Scholar]
- Damann, F.E.; Williams, D.E.; Layton, A.C. Potential Use of Bacterial Community Succession in Decaying Human Bone for Estimating Postmortem Interval. J. Forensic Sci. 2015, 60, 844–850. [Google Scholar] [CrossRef]
- Bohan, T. Scientific Evidence and Forensic Science since Daubert: Maine Decidesto Sitouton the Dance. Maine Law Rev. 2004, 56, 101. [Google Scholar]
- European Network of Forensic Science Institutes ENFSI Guideline for Evaluative Reporting in Forensic Science. Available online: https://enfsi.eu/wp-content/uploads/2016/09/m1_guideline.pdf (accessed on 25 October 2021).
- Neckovic, A.; Van Oorschot, R.A.H.; Szkuta, B.; Durdle, A. Challenges in Human Skin Microbial Profiling for Forensic Science: A Review. Genes 2020, 11, 1015. [Google Scholar] [CrossRef]
- Pasternak, Z.; Luchibia, A.O.; Matan, O.; Dawson, L.; Gafny, R.; Shpitzen, M.; Avraham, S.; Jurkevitch, E. Mitigating temporal mismatches in forensic soil microbial profiles. Aust. J. Forensic Sci. 2019, 51, 685–694. [Google Scholar] [CrossRef]
- Pasternak, Z.; Al Ashhab, A.; Gatica, J.; Gafny, R.; Avraham, S.; Minz, D.; Gillor, O.; Jurkevitch, E. Spatial and Temporal Biogeography of Soil Microbial Communities in Arid and Semiarid Regions. PLoS ONE 2013, 8, e69705. [Google Scholar] [CrossRef]
- Carter, D.O.; Metcalf, J.L.; Bibat, A.; Knight, R. Seasonal variation of postmortem microbial communities. Forensic Sci. Med. Pathol. 2015, 11, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Keet, J.-H.; Ellis, A.; Hui, C.; Le Roux, J. Strong spatial and temporal turnover of soil bacterial communities in South Africa’s hyperdiverse fynbos biome. Soil Biol. Biochem. 2019, 136, 107541. [Google Scholar] [CrossRef]
- Stallins, J.A.; Law, D.M.; Strosberg, S.A.; Rossi, J.J. Geography and postgenomics: How space and place are the new DNA. GeoJournal 2018, 83, 153–168. [Google Scholar] [CrossRef]
- David, L.A.; Materna, A.C.; Friedman, J.; Campos-Baptista, M.I.; Blackburn, M.C.; Perrotta, A.; Erdman, S.E.; Alm, E.J. Host lifestyle affects human microbiota on daily timescales. Genome Biol. 2014, 15, R89-15. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Clarke, T.; Brinkac, L.; Greco, C.; Nelson, K.E. Forensic Microbiome Database: A Tool for Forensic Geolocation Meta-Analysis Using Publicly Available 16S rRNA Microbiome Sequencing. Front. Microbiol. 2021, 12, 650. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling | Samples Should Be Collected Fresh and Then Frozen without Using Any Buffer or Solution. | |
|---|---|---|
| Soil | Swabs | |
| Procedure | Split fresh sample into 2 mL tubes (10) with, at least, 200 mg biomass and store at −80 or −20 °C. | Take 10 replicate swabs with no buffers or solutions and store in −80 or −20 °C |
| Shipping | Samples should be shipped with dry ice in an extruded polystyrene foam container or similar. | |
| Commercial Kit | Principle | Format | Time | Automation |
|---|---|---|---|---|
| MagMAX Microbiome Ultra Nucleic Acid Isolation Kit (ThermoFisher Scientific) [44] | Magnetic beads | 100 reactions | ~60 min | KingFisher™ Duo Prime, Flex and Presto |
| Invitrogen PureLink Microbiome DNA Purification Kit (ThermoFisher Scientific) [45] | Spin column | 100 reactions | 120 min | - |
| QIAamp DNA Microbiome Kit (QIAGEN) [46] | Silica columns | 50 reactions | ~180 min | - |
| MO BIO’s PowerMag® Soil DNA Isolation Kit (QIAGEN) [47] | Magnetic beads | 4 × 96 or 32 × 12 | 60–120 min | epMotion® |
| Forensic microbiome geolocation applications | City planning, public health and architectural design [17]. |
| City geolocation [18]. | |
| Soil evidence geolocation [21]. | |
| Room inhabitant determination and home cohabiting establishment [25]. | |
| Altitude estimation [27]. | |
| Latitude assessment [33]. | |
| Human trafficking [12]. | |
| Ancient DNA studies [51]. |
| Challenge | Considerations |
|---|---|
| Microbiome Transfer |
|
| Sample collection |
|
| DNA extraction |
|
| Sequencing and analysis |
|
| Training and interpretation |
|
| Challenges | Possible Solutions | |
|---|---|---|
| Temporal mismatch [59] | Significant differences in bacterial communities can be observed in the same soil sample if it is analysed at different times due to natural (seasonal) or artificial (storage) changes. | Soil sample enrichment so bacterial communities survive for longer periods. |
| Type of environment [60] | Water availability, changes in plant cover, input of fresh organic matter and temperature variations affect microbiota composition, so ecosystems with high variations in these factors can be challenging to analyse. | Sampling at local scales. |
| Post-mortem microbial communities [61] | Decaying body-associated microbiota changes the soil’s original bacterial communities; in addition, it changes during the various stages of decomposition, and there appear to be seasonal variations in the same soil. | Winter and summer characterization of the soil microbiome. |
| Abiotic soil variables [62] | pH and NH4+ fluctuations, as well as interactions between plants and microorganisms, affect soil bacterial communities. | Monitoring of soil microbiome changes. |
| In vivo microbiome bias [63] | In vivo microbiota can be a consequence not of the geographic place, but of certain sociodemographic aspects linked to culture or inequalities. | Continuous remapping. |
| In vivo microbiota lifestyle [64] | Among individuals in the same area with different lifestyles, diets or routines, the microbiome can vary. An individual microbiome changes drastically due to travelling, dietary changes or a recent infectious disease. | Additional studies on how infections affect host microbiota. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haarkötter, C.; Saiz, M.; Gálvez, X.; Medina-Lozano, M.I.; Álvarez, J.C.; Lorente, J.A. Usefulness of Microbiome for Forensic Geolocation: A Review. Life 2021, 11, 1322. https://doi.org/10.3390/life11121322
Haarkötter C, Saiz M, Gálvez X, Medina-Lozano MI, Álvarez JC, Lorente JA. Usefulness of Microbiome for Forensic Geolocation: A Review. Life. 2021; 11(12):1322. https://doi.org/10.3390/life11121322
Chicago/Turabian StyleHaarkötter, Christian, María Saiz, Xiomara Gálvez, María Isabel Medina-Lozano, Juan Carlos Álvarez, and José Antonio Lorente. 2021. "Usefulness of Microbiome for Forensic Geolocation: A Review" Life 11, no. 12: 1322. https://doi.org/10.3390/life11121322
APA StyleHaarkötter, C., Saiz, M., Gálvez, X., Medina-Lozano, M. I., Álvarez, J. C., & Lorente, J. A. (2021). Usefulness of Microbiome for Forensic Geolocation: A Review. Life, 11(12), 1322. https://doi.org/10.3390/life11121322