
Effects of Hydrostatic Pressure Treatment of Newly Fertilized Eggs on the Ploidy Level and Karyotype of Pikeperch Sander lucioperca (Linnaeus, 1758)


 ,
,  , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hydrostatic Pressure Shock Treatment
2.2. Chromosome Analysis of Pikeperch Larvae from the Treated and Control Groups
2.3. Sample Preparation for Laser-Scanning Cytometry from the Hydrostatic-Pressure-Treated and Control Groups
2.4. Laser-Scanning Cytometry Measurements and Analysis of Data
2.5. Statistical Analysis
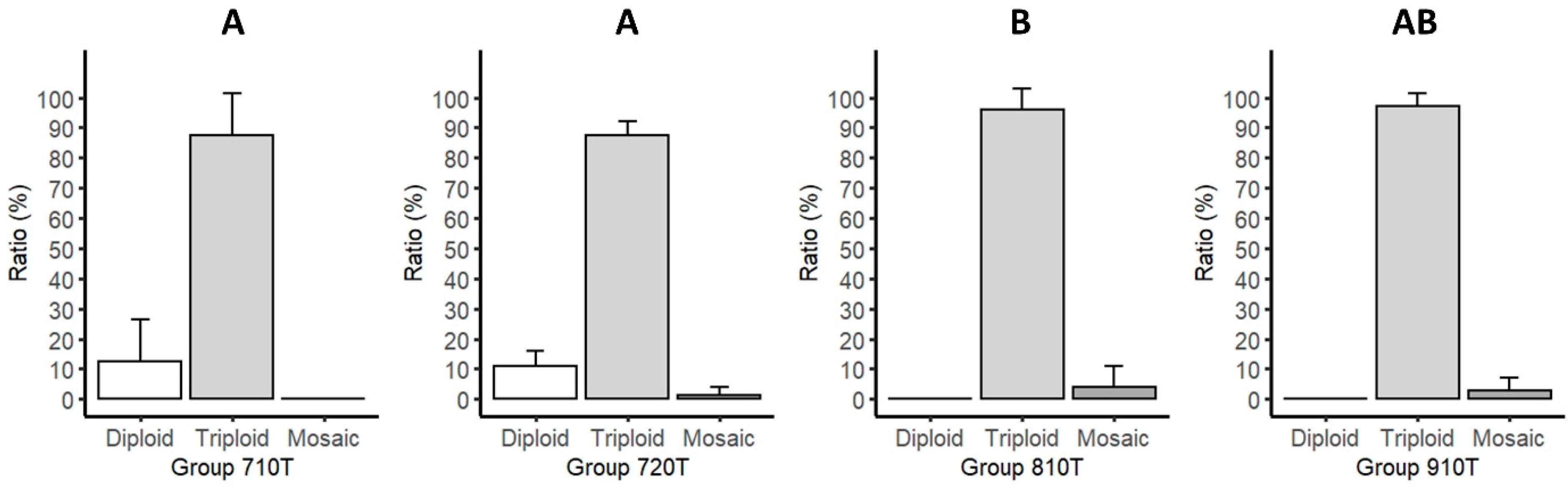
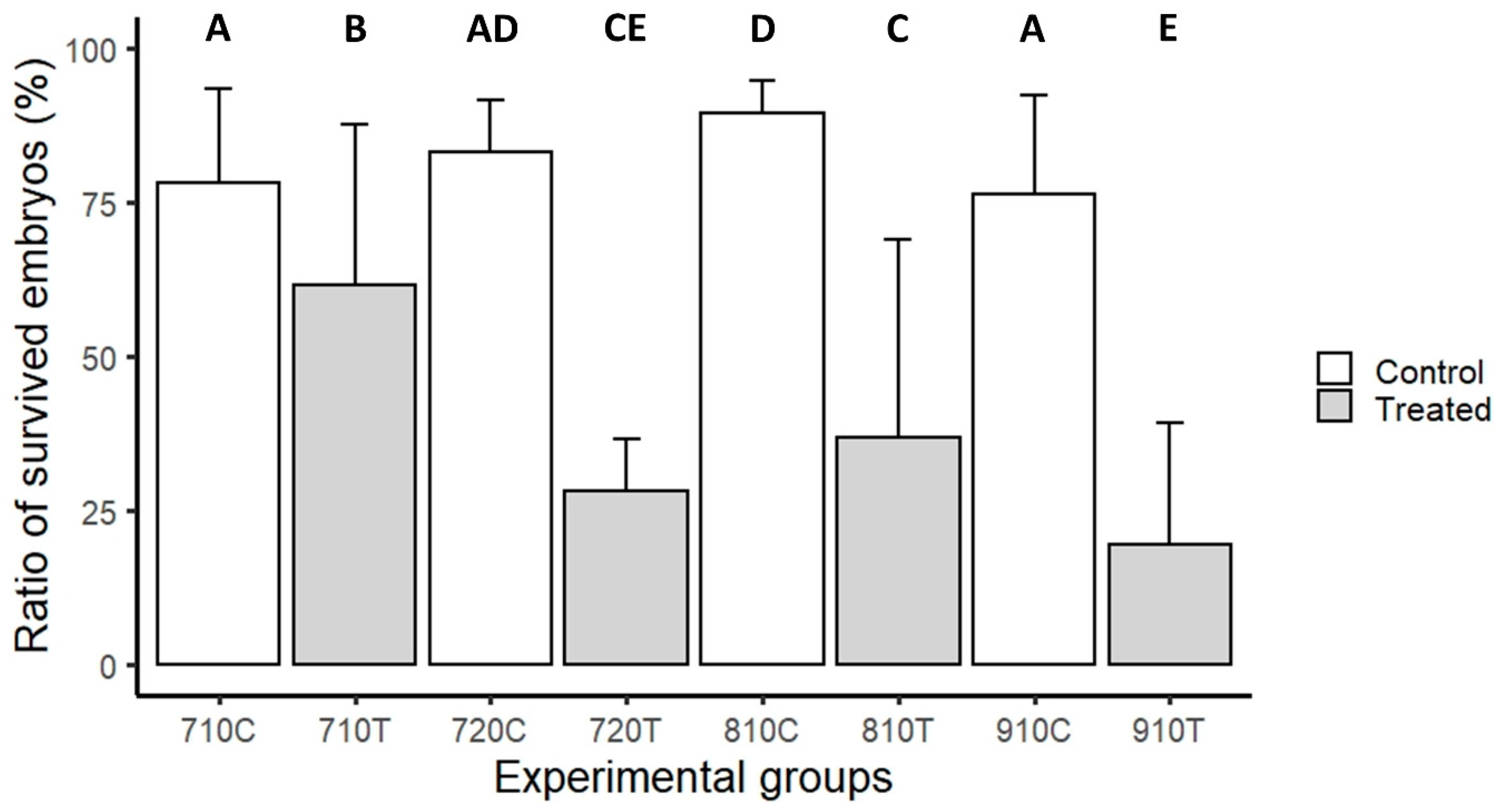
3. Results
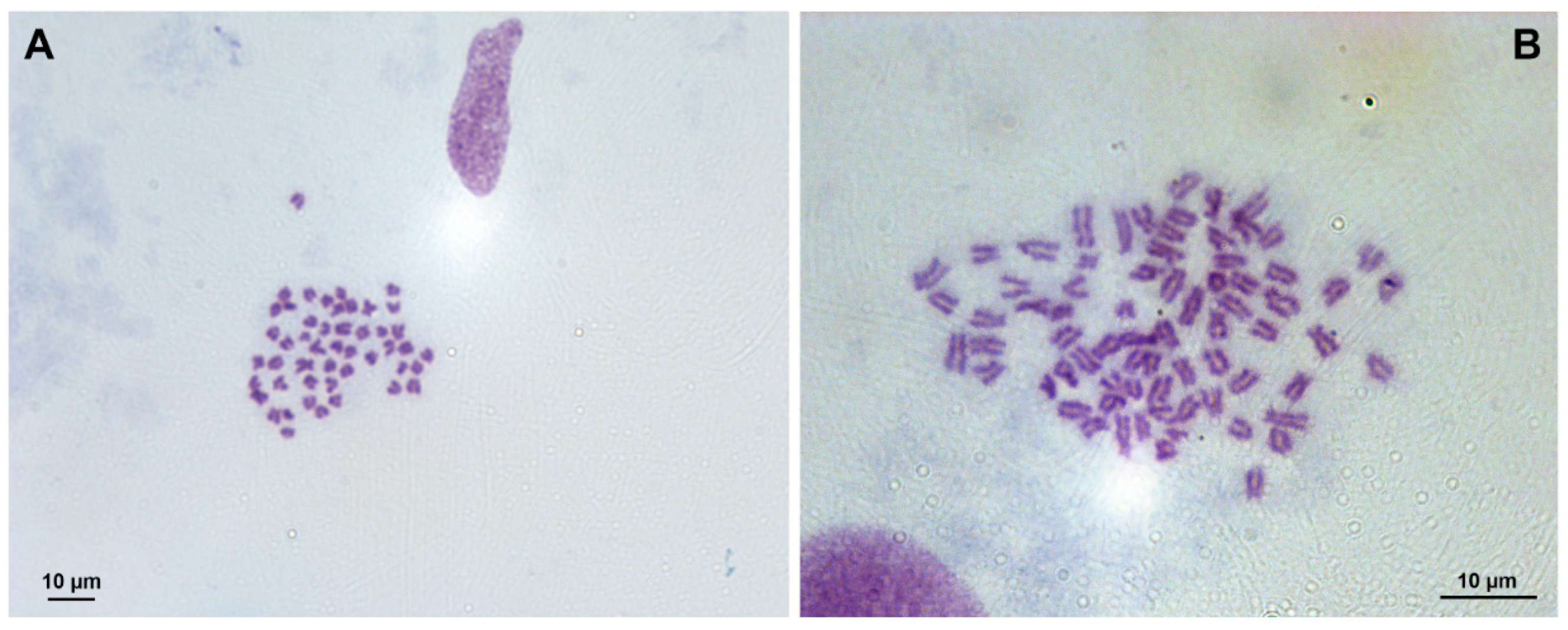
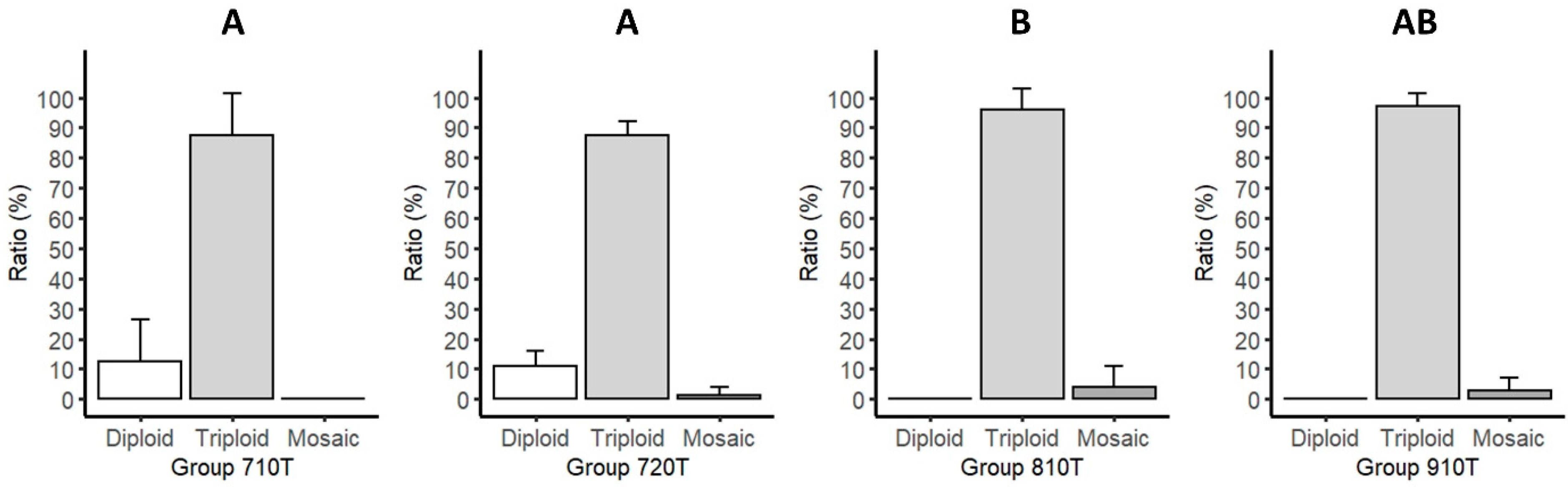
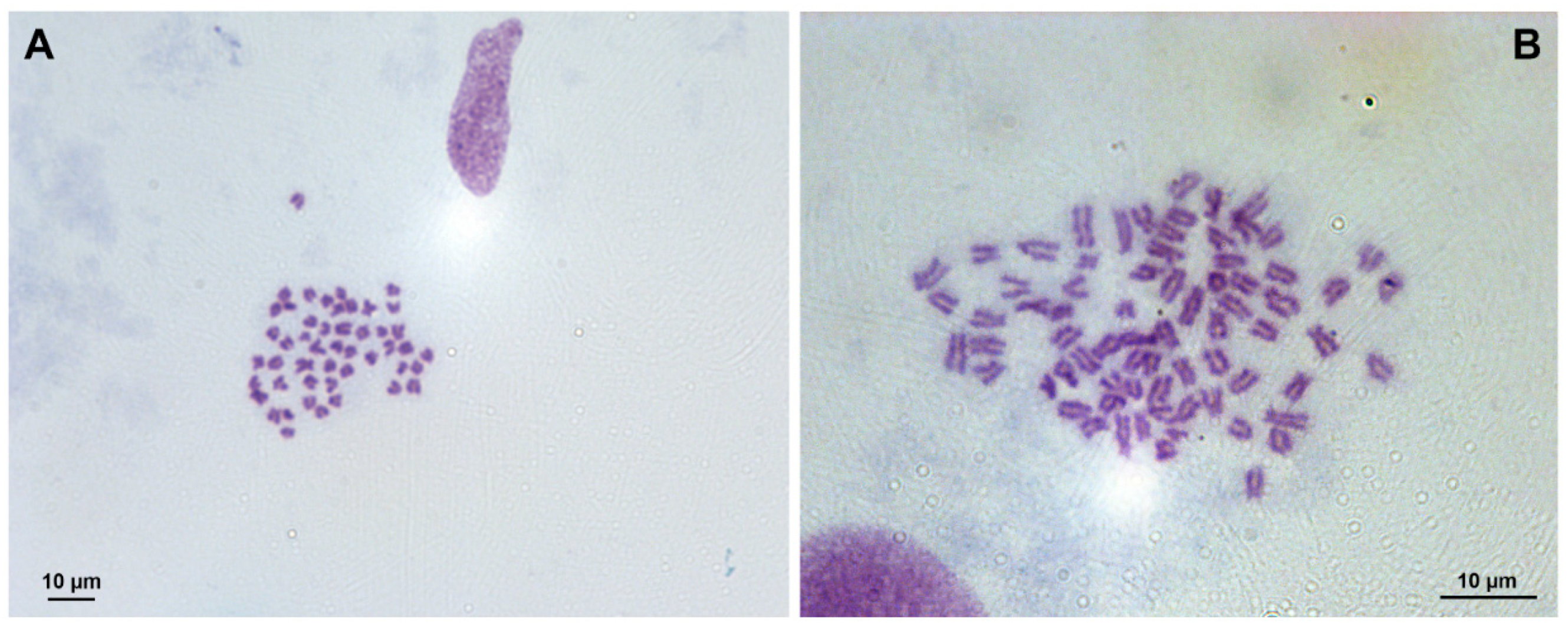
3.1. Chromosome Analysis
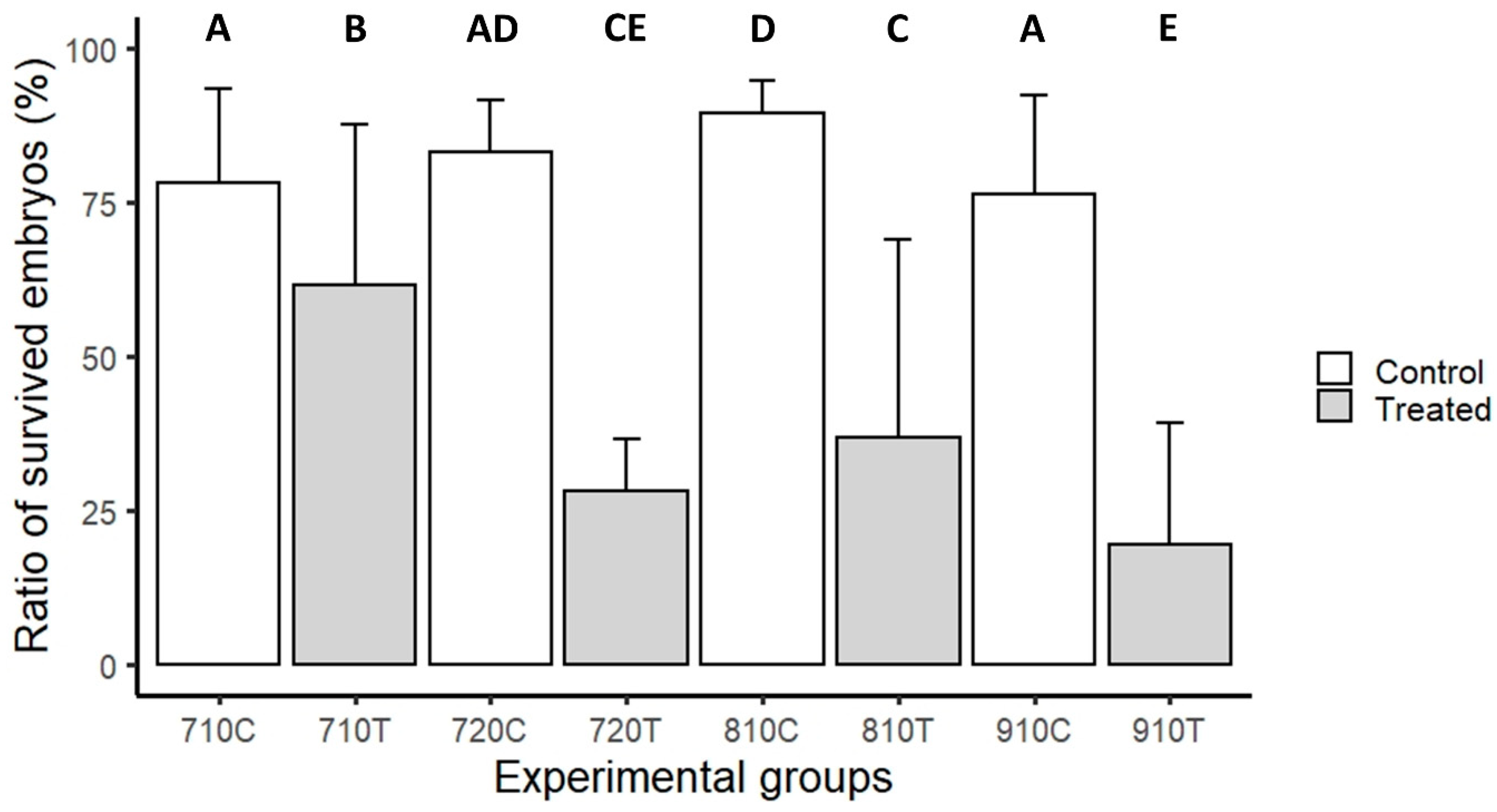
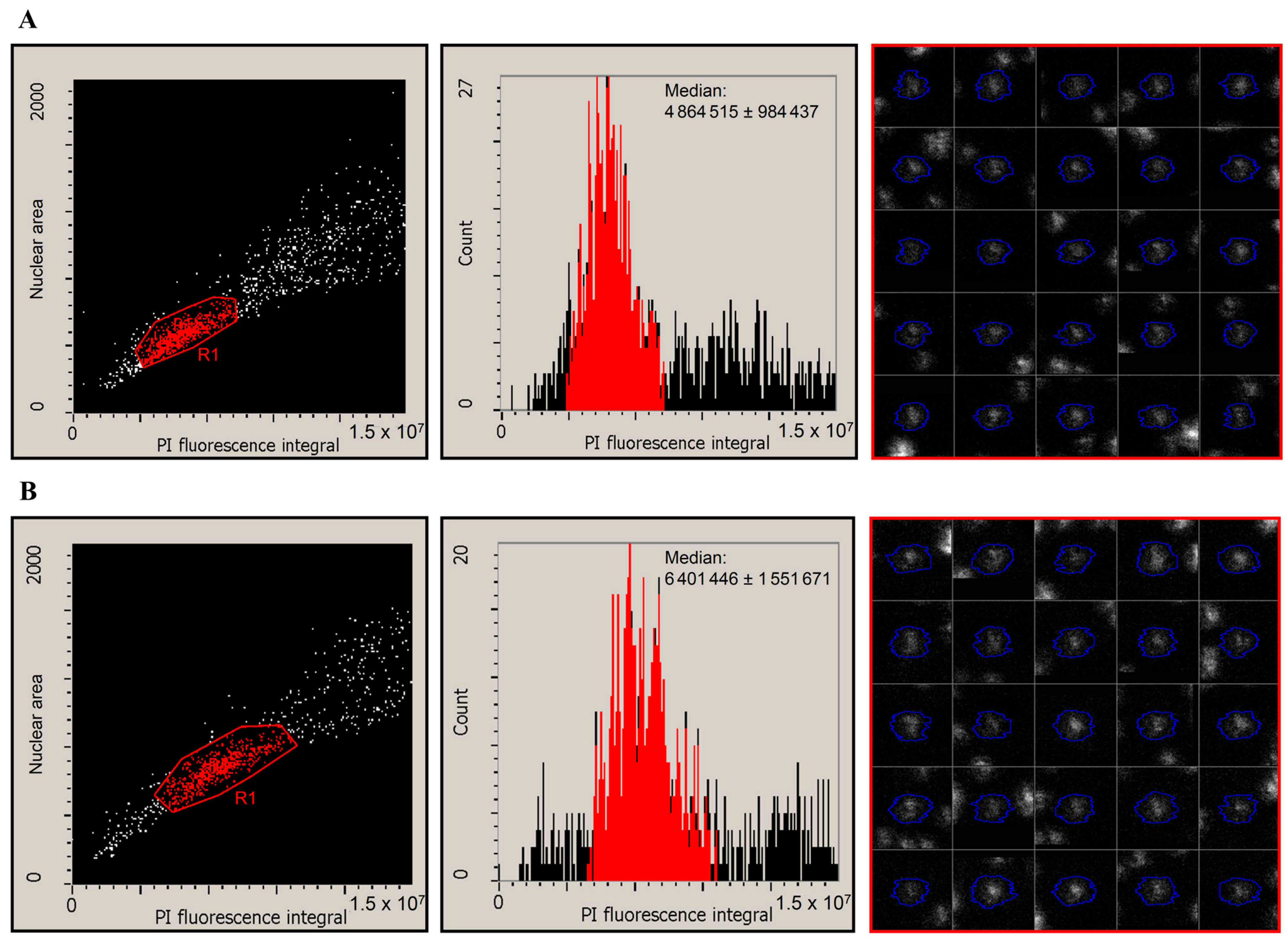
3.2. Laser-Scanning Cytometry Measurement
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tóth, B.; Várkonyi, E.; Hidas, A.; Edviné Meleg, E.; Váradi, L. Genetic analysis of offspring from intra- and interspecific crosses of Carassius auratus gibelio by chromosome and RAPD analysis. J. Fish Biol. 2005, 66, 784–797. [Google Scholar] [CrossRef]
- Zhou, L.; Gui, J. Natural and artificial polyploids in aquaculture. Aquac. Fish. 2017, 2, 103–111. [Google Scholar] [CrossRef]
- Ohno, S. Evolution by Gene Duplication; Springer Science + Business Media: New York, NY, USA, 1970; p. 160. [Google Scholar]
- Ráb, P.; Roth, P.; Mayr, B. Karyotype Study of Eight Species of European Percid Fishes (Pisces, Percidae). Caryologia 1987, 40, 307–318. [Google Scholar] [CrossRef]
- Ihssen, P.E.; McKay, L.R.; McMillan, I.; Phillips, R.B. Ploidy Manipulation and Gynogenesis in Fishes: Cytogenetic and Fisheries Applications. Trans. Am. Fish. Soc. 1990, 119, 698–717. [Google Scholar] [CrossRef]
- Rougeot, C. Sex and Ploidy Manipulation in Percid Fishes. In Biology and Culture of Percid Fishes; Springer Science+Business Media: Dordrecht, The Netherland, 2015; p. 629. [Google Scholar] [CrossRef]
- Chourrout, D. Pressure-induced retention of second polar body and suppression of first cleavage in rainbow trout: Production of all-triploids, all-tetraploids, and heterozygous and homozygous diploid gynogenetics. Aquaculture 1984, 36, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Malison, J.A.; Terrence, B.K.; James, A.H.; Terence, P.B.; Clyde, H.A. Manipulation of ploidy in yellow perch (Perca flavescens) by heat shock, hydrostatic pressure shock, and spermatozoa inactivation. Aquaculture 1993, 110, 229–242. [Google Scholar] [CrossRef]
- Yamazakia, F.; Goodierb, J. Cytogenetic effects of hydrostatic pressure treatment to suppress the first cleavage of salmon embryos. Aquaculture 1993, 110, 51–59. [Google Scholar] [CrossRef]
- Glover, K.A.; Harvey, A.C.; Hansen, T.J.; Fjelldal, P.G.; Besnier, F.N.; Bos, J.B.; Ayllon, F.; Taggart, J.B.; Solberg, M.F. Chromosome aberrations in pressure induced triploid Atlantic salmon. BMC Gen. 2020, 21, 59. [Google Scholar] [CrossRef]
- Malison, J.A.; Held, J.A.; Weil, L.S.; Terrence, B.K.; Thorgaard, G.H. Manipulation of Ploidy in Walleyes by Heat Shock and Hydrostatic Pressure Shock. N. Am. J. Aquac. 2001, 63, 17–24. [Google Scholar] [CrossRef]
- Devlin, R.H.; Sakhrani, D.; Biagi, C.A.; Ki-Whan, E. Occurrence of incomplete paternal-chromosome retention in GH-transgenic coho salmon being assessed for reproductive containment by pressure-shock-induced triploidy. Aquaculture 2010, 304, 66–78. [Google Scholar] [CrossRef]
- Várkonyi, E.; Bercsényi, M.; Ozouf-Costaz, C.; Billard, R. Chromosomal and morphological abnormalities caused by oocyte ageing in Silurus glanis. J. Fish Biol. 1998, 52, 899–906. [Google Scholar] [CrossRef]
- Rougeot, C.; Minet, L.; Prignon, C.; Vanderplasschen, A.; Detry, B.; Pastoret, P.-P.; Mélard, C. Induce triploidy by heat shock in Eurasian perch, Perca fluviatilis. Aquat. Living Resour. 2003, 16, 90–94. [Google Scholar] [CrossRef]
- Piferrer, F.; Beaumont, A.; Falguière, J.C.; Flajšhans, M.; Haffray, P.; Colombo, L. Polyploidfish and shellfish: Production, biology and applications to aquaculture for performance improvement and genetic containment. Aquaculture 2009, 293, 125–156. [Google Scholar] [CrossRef] [Green Version]
- Fraser, W.K.T.; Fjelldal, P.G.; Hansen, T.; Mayer, I. Welfare Considerations of Triploid Fish. Rev. Fish. Sci. 2012, 20, 192–211. [Google Scholar] [CrossRef]
- Lee, S.; Katayama, N.; Yoshizaki, G. Generation of juvenile rainbow trout derived from cryopreserved whole ovaries by intraperitoneal transplantation of ovarian germ cells. Biochem. Biophys. Res. Commun. 2016, 478, 1478–1483. [Google Scholar] [CrossRef] [Green Version]
- Seki, S.; Kusano, K.; Lee, S.; Iwasaki, Y.; Yagisawa, M.; Ishida, M.; Hiratsuka, T.; Sasado, T.; Naruse, K.; Yoshizaki, G. Production of the medaka derived from vitrified whole testes by germ cell transplantation. Sci. Rep. 2017, 7, 43185. [Google Scholar] [CrossRef] [Green Version]
- Hamasaki, M.; Takeuchi, Y.; Yazawa, R.; Yoshikawa, S.; Kadomura, K.; Yamada, T.; Miyaki, K.; Kikuchi, K.; Yoshizaki, G. Production of tiger puffer Takifugu rubripes offspring from triploid grass puffer Takifugu niphobles parents. Mar. Biotechnol. 2017, 19, 579–591. [Google Scholar] [CrossRef]
- Franĕk, R.; Tichopád, T.; Fučíková, M.; Steinbach, C.; Pšenička, M. Production and use of triploid zebrafish for surrogate reproduction. Theriogenology 2019, 140, 33–43. [Google Scholar] [CrossRef]
- Marinovic, Z.; Lujic, J.; Kasa, E.; Csenki, Z.; Urbanyi, B.; Horvath, A. Cryopreservation of zebrafish spermatogonia by whole testes needle immersed ultra-rapid cooling. J. Vis. Exp. 2018, 133, 2–7. [Google Scholar] [CrossRef]
- Ljubobratovič, U.; Kwiatkowski, M.; Tóth, F.; Demény, F. Effects of hormonal treatment before water warming on synchronisation of spawning time, oocyte size, and egg quality in pikeperch (Sander lucioperca). Anim. Reprod. Sci. 2021, 226, 106712. [Google Scholar] [CrossRef]
- Kristan, J.; Blecha, M.; Policar, T. Alcalase treatment for elimination of stickiness in pikeperch (Sander lucioperca L.) eggs under controlled conditions. Aquac. Res. 2015, 47, 3998–4003. [Google Scholar] [CrossRef]
- Uroš Ljubobratović, Géza Péter, Rene Alvestad, Zoltán Horváth, András Rónyai: Alcalase enzyme treatment affects egg incubation and larval quality in pikeperch (Sander lucioperca). Aquac. Int. 2019, 27, 917–929. [CrossRef]
- Malison, J.A.; Procarione, L.S.; Held, J.A.; Kayes, T.B.; Amundson, C.H. The influence of triploidy and heat and hydrostatic pressure shocks on the growth and reproductive development of juvenile yellow perch (Perca flavescens). Aquaculture 1993, 116, 121–133. [Google Scholar] [CrossRef]
- Lemoine, H.L., Jr.; Smith, T.L. Polyploidy Induced in Brook Trout by Cold Shock. Trans. Am. Fish. Soc. 1980, 109, 626–631. [Google Scholar] [CrossRef]
- Smith, L.T.; Lemoine, H.L. (1979) Colchicine-Induced Polyploidy in Brook Trout. Progress. Fish-Cult. 1979, 41, 86–88. [Google Scholar] [CrossRef]
- Lebeda, I.; Ráb, P.; Majtánová, Z.; Flajšhans, M. Artifcial whole genome duplication in paleopolyploid sturgeons yields highest documented chromosome number in vertebrates. Sci. Rep. 2020, 10, 19705. [Google Scholar] [CrossRef]
- Allen, S.K., Jr.; Stanley, J.G. Polyploid Mosaics Induced by Cytochalasin B in Landlocked Atlantic Salmon Salmo salar. Trans. Am. Fish. Soc. 1979, 108, 462–466. [Google Scholar] [CrossRef]
- Refstie, T.; Vassvik, V.; Gjedrem, T. Induction of polyploidy in salmonids by cytochalasin B. Aquaculture 1977, 10, 65–74. [Google Scholar] [CrossRef]
- Fetherman, E.R.; Lepak, J.M.; Brown, B.L.; Harris, D.J. Optimizing Time of Initiation for Triploid Walleye Production Using Pressure Shock Treatment. North Am. J. Aquac. 2015, 77, 471–477. [Google Scholar] [CrossRef]
- Dadras, H.; Blecha, M.; Malinovskyi, O.; Flajšhans, M.; Lebeda, I.; Křišťan, J.; Policar, T. Triploidization in pikeperch (Sander lucioperca) induced by cold shock. Aquaculture 2021, 533, 736236. [Google Scholar] [CrossRef]
- Garcia-Abiado, M.A.R.; Lynch, W.E., Jr.; Dabrowski, K.; Hartman, T. Use of Thermal and Pressure Shocks to Induce Triploid Hybrid Saugeyes. North Am. J. Aquac. 2001, 63, 2. [Google Scholar] [CrossRef]
- Blecha, M.; Flajshans, M.; Lebeda, I.; Kristan, J.; Svacina, P.; Policar, T. Triploidisation of pikeperch (Sander lucioperca), first success. Aquaculture 2016, 462, 115–117. [Google Scholar] [CrossRef]
- Garcia-Abiado, M.A.; Penn, M.; Dabrowski, K.; Stafford, J. Evaluation of Two Commercially Available Pressure Chambers to Induce Triploidy in Saugeyes. N. Am. J. Aquac. 2007, 69, 197–201. [Google Scholar] [CrossRef]
- Swarup, H. Production of triploidy ingasterosteus aculeatus (L). J. Genet. 1959, 56, 129–142. [Google Scholar] [CrossRef]
- Svardson, G. Chromosome Studies on salmonidae. Ph.D. Thesis, Stockholm College, Stockholm, Sweden, 1945; p. 151. [Google Scholar]
- Iegorova, V.; Psenicka, M.; Lebeda, I.; Rodina, M.; Saito, T. Polyspermy produces viable haploid/diploid mosaics in sturgeon. Biol. Rep. 2018, 99, 695–706. [Google Scholar] [CrossRef]
- Iegorova, V.; Havelka, M.; Psenicka, M.; Saito, T. First evidence of viable progeny from three interspecific parents in sturgeon. Fish Phys. Biochem. 2018, 44, 1541–1550. [Google Scholar] [CrossRef]
- Briedis, A.; Elinson, R. Suppression of Male Pronuclear Movement in Frog Eggs by Hydrostatic Pressure and Deuterium Oxide Yields Androgenetic Haploids. J. Exp. Zool. 1982, 222, 45–57. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pressure (PSI) | Duration (min) | Type | Replicate | Identifier | Diploid Fish (pcs) | Triploid Fish (pcs) | Mosaic Fish (pcs) | Karyotyped Fish (pcs) | Diploid (%)/ Triploid (%) |
|---|---|---|---|---|---|---|---|---|---|
| - | - | Control | 1 | 710Ca | 11 | 0 | 0 | 11 | |
| 2 | 710Cb | 15 | 0 | 0 | 15 | 100.0 a/0.0 b | |||
| 3 | 710Cc | 16 | 0 | 0 | 16 | ||||
| 7000 | 10 | Treated | 1 | 710Ta | 1 | 9 | 0 | 10 | 12.3 ± 11.1 c/ 87.7 ± 11.1 d |
| 2 | 710Tb | 3 | 8 | 0 | 11 | ||||
| 3 | 710Tc | 0 | 11 | 0 | 11 | ||||
| - | - | Control | 1 | 720Ca | 15 | 0 | 0 | 15 | |
| 2 | 720Cb | 19 | 0 | 0 | 19 | 100.0 a/0.0 b | |||
| 3 | 720Cc | 16 | 0 | 0 | 16 | ||||
| 7000 | 20 | Treated | 1 | 720Ta | 2 | 24 | 0 | 26 | 11.3 ± 4.0 c/ 87.0 ± 3.7 d |
| 2 | 720Tb | 1 | 5 | 0 | 6 | ||||
| 3 | 720Tc | 2 | 20 | 1 | 23 | ||||
| - | - | Control | 1 | 810Ca | 27 | 0 | 0 | 27 | |
| 2 | 810Cb | 21 | 0 | 0 | 21 | 100.0 a/0.0 b | |||
| 3 | 810Cc | 13 | 0 | 0 | 13 | ||||
| 8000 | 10 | Treated | 1 | 810Ta | 0 | 22 | 3 | 25 | 0.0 e/ 96.0 ± 5.7 f |
| 2 | 810Tb | 0 | 14 | 0 | 14 | ||||
| 3 | 810Tc | 0 | 19 | 0 | 19 | ||||
| - | - | Control | 1 | 910Ca | 15 | 0 | 0 | 15 | |
| 2 | 910Cb | 26 | 0 | 0 | 26 | 100.0 a/0.0 b | |||
| 3 | 910Cc | 17 | 0 | 0 | 17 | ||||
| 9000 | 10 | Treated | 1 | 910Ta | 0 | 15 | 0 | 15 | 0.0 e/ 97.0 ± 3.0 df |
| 2 | 910Tb | 0 | 16 | 1 | 17 | ||||
| 3 | 910Tc | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Káldy, J.; Patakiné Várkonyi, E.; Fazekas, G.L.; Nagy, Z.; Sándor, Z.J.; Bogár, K.; Kovács, G.; Molnár, M.; Lázár, B.; Goda, K.; et al. Effects of Hydrostatic Pressure Treatment of Newly Fertilized Eggs on the Ploidy Level and Karyotype of Pikeperch Sander lucioperca (Linnaeus, 1758). Life 2021, 11, 1296. https://doi.org/10.3390/life11121296
Káldy J, Patakiné Várkonyi E, Fazekas GL, Nagy Z, Sándor ZJ, Bogár K, Kovács G, Molnár M, Lázár B, Goda K, et al. Effects of Hydrostatic Pressure Treatment of Newly Fertilized Eggs on the Ploidy Level and Karyotype of Pikeperch Sander lucioperca (Linnaeus, 1758). Life. 2021; 11(12):1296. https://doi.org/10.3390/life11121296
Chicago/Turabian StyleKáldy, Jenő, Eszter Patakiné Várkonyi, Georgina Lea Fazekas, Zoltán Nagy, Zsuzsanna J. Sándor, Katalin Bogár, Gyula Kovács, Mariann Molnár, Bence Lázár, Katalin Goda, and et al. 2021. "Effects of Hydrostatic Pressure Treatment of Newly Fertilized Eggs on the Ploidy Level and Karyotype of Pikeperch Sander lucioperca (Linnaeus, 1758)" Life 11, no. 12: 1296. https://doi.org/10.3390/life11121296
APA StyleKáldy, J., Patakiné Várkonyi, E., Fazekas, G. L., Nagy, Z., Sándor, Z. J., Bogár, K., Kovács, G., Molnár, M., Lázár, B., Goda, K., Gyöngy, Z., Ritter, Z., Nánási, P., Jr., Horváth, Á., & Ljubobratović, U. (2021). Effects of Hydrostatic Pressure Treatment of Newly Fertilized Eggs on the Ploidy Level and Karyotype of Pikeperch Sander lucioperca (Linnaeus, 1758). Life, 11(12), 1296. https://doi.org/10.3390/life11121296