
AC016745.3 Regulates the Transcription of AR Target Genes by Antagonizing NONO

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Androgen Treatment
2.2. RNA Interference and Transfection
2.3. qRT-PCR Analysis
2.4. RNA Pull down Assay
2.5. Mass-Spectrometric Sequencing Analysis
2.6. RNA Immunoprecipitation Assay
2.7. Chromatin Immunoprecipitation Assay
2.8. Co-Immunoprecipitation
2.9. Cell Migration Assay
2.10. Cell Proliferation Assay
2.11. Cell Cycle Assay
2.12. Annexin V-FITC Apoptosis Detection
2.13. Statistical Analysis
3. Results
3.1. AC016745.3 Is a Target of AR-Dependent Gene Repression and Is Downregulated in LNCaP Cells in Response to DHT Stimulation
3.2. AC016745.3 Acts as a Tumor Suppressor Gene
3.3. AC016745.3 Binds the AR Transcription Complex and Inhibits AR-Dependent Gene Transcription
3.4. Identification of AC016745.3 and AR Interacting Proteins within the AR Transcription Complex
3.5. NONO Is a Putative Proto-Oncogene That Promotes the Transcription of AR Target Genes
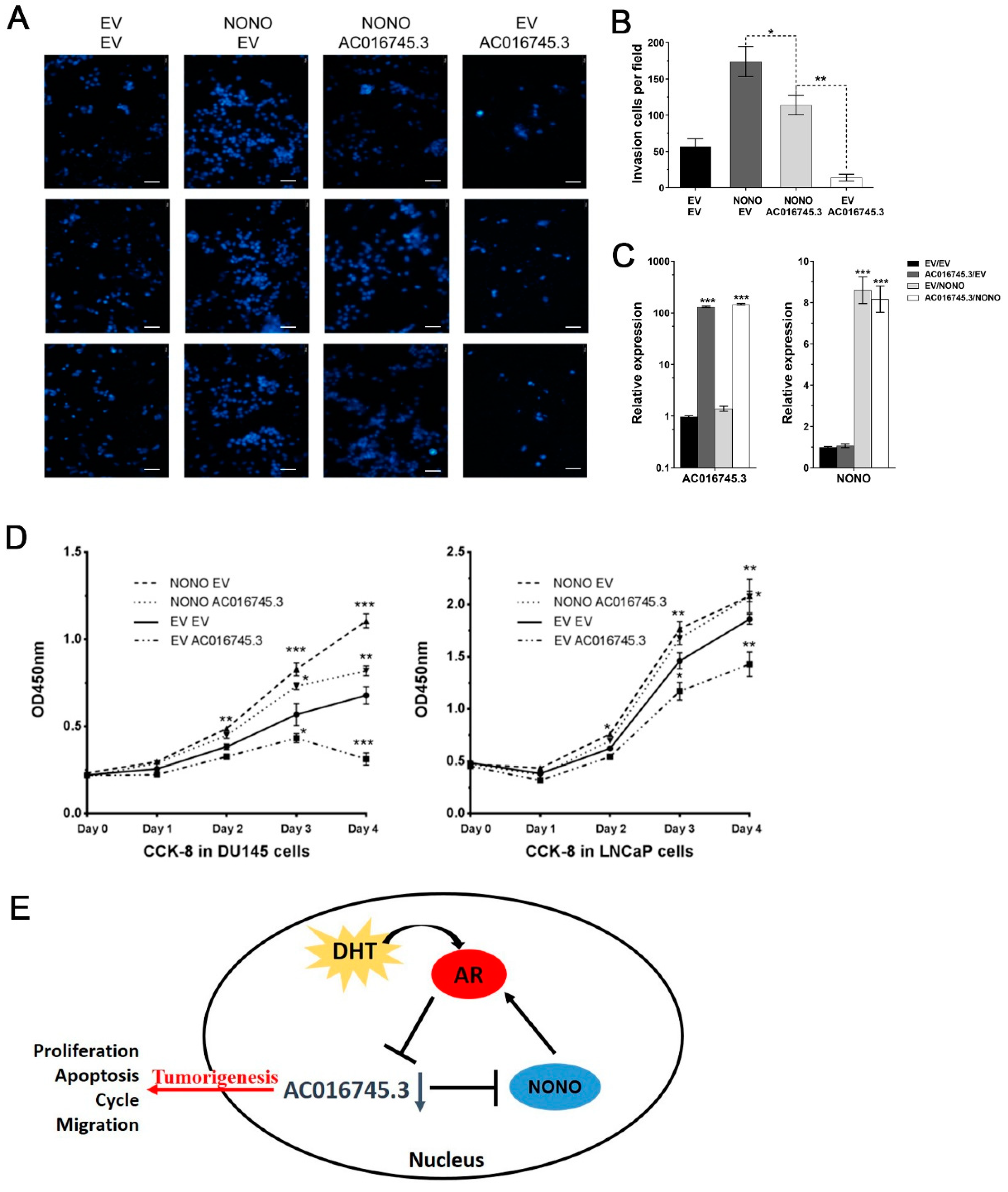
3.6. AC016745.3 Partially Restores NONO Function in PCa Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA-Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Uo, T.; Plymate, S.R.; Sprenger, C.C. Allosteric alterations in the androgen receptor and activity in prostate cancer. Endocr Relat Cancer. 2017, 24, R335–R348. [Google Scholar] [CrossRef] [Green Version]
- Leach, D.A.; Buchanan, G. Stromal Androgen Receptor in Prostate Cancer Development and Progression. Cancers 2017, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Ruan, H.; Xu, T.; Liu, L.; Liu, D.; Yang, H.; Zhang, X.; Chen, K. Recent advances on the progressive mechanism and therapy in castration-resistant prostate cancer. Onco Targets Ther. 2018, 11, 3167–3178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galletti, G.; Leach, B.I.; Lam, L.; Tagawa, S.T. Mechanisms of resistance to systemic therapy in metastatic castration-resistant prostate cancer. Cancer Treat. Rev. 2017, 57, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Rathkopf, D.E.; Smith, M.R.; Ryan, C.J.; Berry, W.R.; Shore, N.D.; Liu, G.; Higano, C.S.; Alumkal, J.J.; Hauke, R.; Tutrone, R.F.; et al. Androgen receptor mutations in patients with castration-resistant prostate cancer treated with apalutamide. Ann. Oncol. 2017, 28, 2264–2271. [Google Scholar] [CrossRef] [PubMed]
- Crona, D.J.; Whang, Y.E. Androgen Receptor-Dependent and -Independent Mechanisms Involved in Prostate Cancer Therapy Resistance. Cancers 2017, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Sahin, I.; Mega, A.E.; Carneiro, B.A. Androgen receptor-independent prostate cancer: An emerging clinical entity. Cancer Biol. Ther. 2018, 19, 347–348. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Zhai, W.; Sun, Y.; Guo, C.; Hu, G.; Wang, M.; Zheng, J.; Lin, W.; Huang, Q.; Li, G.; Zheng, J.; et al. LncRNA-SARCC suppresses renal cell carcinoma (RCC) progression via altering the androgen receptor(AR)/miRNA-143-3p signals. Cell Death Differ. 2017, 24, 1502–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yari, H.; Jin, L.; Teng, L.; Wang, Y.F.; Wu, Y.Y.; Liu, G.Z.; Gao, W.; Liang, J.; Xi, Y.; Feng, Y.C.; et al. LncRNA REG1CP promotes tumorigenesis through an enhancer complex to recruit FANCJ helicase for REG3A transcription. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.; Xu, C.; Li, Y.; Cai, X.; Ren, S.; Liu, H.; Wang, Y.; Wang, F.; Chen, R.; Qu, M.; et al. A feed-forward regulatory loop between androgen receptor and PlncRNA-1 promotes prostate cancer progression. Cancer Lett. 2016, 374, 62–74. [Google Scholar] [CrossRef]
- Pestell, R.G.; Yu, Z. Long and noncoding RNAs (lnc-RNAs) determine androgen receptor dependent gene expression in prostate cancer growth in vivo. Asian J. Androl. 2014, 16, 268–269. [Google Scholar] [CrossRef]
- Sattarifard, H.; Hashemi, M.; Hassanzarei, S.; Narouie, B.; Bahari, G. Association between genetic polymorphisms of long non-coding RNA PRNCR1 and prostate cancer risk in a sample of the Iranian population. Mol. Clin. Oncol. 2017, 7, 1152–1158. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.; Zhao, J.C.; Kim, J.; Fong, K.W.; Yang, Y.A.; Chakravarti, D.; Mo, Y.-Y.; Yu, J. LncRNA HOTAIR Enhances the Androgen-Receptor-Mediated Transcriptional Program and Drives Castration-Resistant Prostate Cancer. Cell Rep. 2015, 13, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J.; Zhang, Y.; Wan, X.; Zhang, C.; Huang, X.; Huang, W.; Pu, H.; Pei, C.; Wu, H.; et al. KDM1A triggers androgen-induced miRNA transcription via H3K4me2 demethylation and DNA oxidation. Prostate 2015, 75, 936–946. [Google Scholar] [CrossRef]
- Yang, L.; Lin, C.; Jin, C.; Yang, J.C.; Tanasa, B.; Li, W.; Merkurjev, D.; Ohgi, K.A.; Meng, D. lncRNA-dependent mechanisms of androgen-receptor-regulated gene activation programs. Nature 2013, 500, 598–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, L.F.; Beckedorff, F.C.; Ayupe, A.C.; Amaral, M.S.; Mesel, V.; Videira, A.; Reis, E.M.; Setubal, J.C.; Verjovski-Almeida, S. Chromatin Landscape Distinguishes the Genomic Loci of Hundreds of Androgen-Receptor-Associated LincRNAs From the Loci of Non-associated LincRNAs. Front Genet. 2018, 9, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, C.L.; Fei, T.; Chen, Y.W.; Li, T.T.; Gao, Y.F.; Wang, X.D.; Sun, T.; Sweeney, C.J.; Lee, G.-S.M.; Chen, S.; et al. Enhancer RNAs participate in androgen receptor-driven looping that selectively enhances gene activation. Proc. Natl. Acad. Sci. USA 2014, 111, 7319–7324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishitani, K.; Yoshida, T.; Kitagawa, H.; Ohta, H.; Nozawa, S.; Kato, S. p54nrb acts as a transcriptional coactivator for activation function 1 of the human androgen receptor. Biochem. Biophys. Res. Commun. 2003, 306, 660–665. [Google Scholar] [CrossRef]
- Zhang, P.; Lu, Y.; Kong, Z.; Zhang, Y.; Fu, F.; Su, X.; Huang, Y.; Wan, X.; Li, Y. Androgen-responsive lncRNA LINC00304 promotes cell cycle and proliferation via regulating CCNA1. Prostate 2019, 79, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Horie-Inoue, K.; Katayama, S.; Suzuki, T.; Tsutsumi, S.; Ikeda, K.; Urano, T.; Fujimura, T.; Takagi, K.; Takahashi, S.; et al. Androgen-responsive long noncoding RNA CTBP1-AS promotes prostate cancer. EMBO J. 2013, 32, 1665–1680. [Google Scholar] [CrossRef] [Green Version]
- Misawa, A.; Takayama, K.; Urano, T.; Inoue, S. Androgen-induced Long Noncoding RNA (lncRNA) SOCS2-AS1 Promotes Cell Growth and Inhibits Apoptosis in Prostate Cancer Cells. J. Biol. Chem. 2016, 291, 17861–17880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, T.T.; Huang, J.; Zhou, N.; Zhang, Z.; Koirala, P.; Zhou, X.; Wu, F.; Ding, X.; Mo, Y.-Y. Regulation of PCGEM1 by p54/nrb in prostate cancer. Sci. Rep. 2016, 6, 34529. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, R.; Osawa, T.; Sasaki, Y.; Yamamoto, S.; Anai, M.; Izumi, K.; Matsumura, Y.; Sakai, J.; Aburatani, H.; Mizokami, A.; et al. Overexpression of p54(nrb)/NONO induces differential EPHA6 splicing and contributes to castration-resistant prostate cancer growth. Oncotarget 2018, 9, 10510–10524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iino, K.; Mitobe, Y.; Ikeda, K.; Takayama, K.I.; Suzuki, T.; Kawabata, H.; Suzuki, Y.; Horie-Inoue, K.; Inoue, S. RNA-binding protein NONO promotes breast cancer proliferation by post-transcriptional regulation of SKP2 and E2F8. Cancer Sci. 2020, 111, 148–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Silva, H.C.; Lin, M.Z.; Phillips, L.; Martin, J.L.; Baxter, R.C. IGFBP-3 interacts with NONO and SFPQ in PARP-dependent DNA damage repair in triple-negative breast cancer. Cell Mol. Life Sci. 2019, 76, 2015–2030. [Google Scholar] [CrossRef]
- Lee, A.R.; Hung, W.; Xie, N.; Liu, L.; He, L.; Dong, X. Tyrosine Residues Regulate Multiple Nuclear Functions of P54nrb. J. Cell Physiol. 2017, 232, 852–861. [Google Scholar] [CrossRef]
- Knott, G.J.; Bond, C.S.; Fox, A.H. The DBHS proteins SFPQ, NONO and PSPC1: A multipurpose molecular scaffold. Nucleic Acids Res. 2016, 44, 3989–4004. [Google Scholar] [CrossRef]
- Feng, P.F.; Li, L.; Deng, T.G.; Liu, Y.; Ling, N.; Qiu, S.Y.; Zhang, L.; Peng, B.; Xiong, W.; Cao, L.; et al. NONO and tumorigenesis: More than splicing. J. Cell Mol. Med. 2020, 24, 4368–4376. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.; Wan, X.; Huang, W.; Zhang, L.; Luo, J.; Li, D.; Huang, Y.; Li, Y.; Xu, Y. AC016745.3 Regulates the Transcription of AR Target Genes by Antagonizing NONO. Life 2021, 11, 1208. https://doi.org/10.3390/life11111208
Lu Y, Wan X, Huang W, Zhang L, Luo J, Li D, Huang Y, Li Y, Xu Y. AC016745.3 Regulates the Transcription of AR Target Genes by Antagonizing NONO. Life. 2021; 11(11):1208. https://doi.org/10.3390/life11111208
Chicago/Turabian StyleLu, Yali, Xuechao Wan, Wenhua Huang, Lu Zhang, Jun Luo, Dujian Li, Yan Huang, Yao Li, and Yaoting Xu. 2021. "AC016745.3 Regulates the Transcription of AR Target Genes by Antagonizing NONO" Life 11, no. 11: 1208. https://doi.org/10.3390/life11111208
APA StyleLu, Y., Wan, X., Huang, W., Zhang, L., Luo, J., Li, D., Huang, Y., Li, Y., & Xu, Y. (2021). AC016745.3 Regulates the Transcription of AR Target Genes by Antagonizing NONO. Life, 11(11), 1208. https://doi.org/10.3390/life11111208