Human Umbilical Cord: Information Mine in Sex-Specific Medicine

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Strategy Search
3. Sex Differences in Fetal Growth and Placenta
4. Fetal Programming and Sex
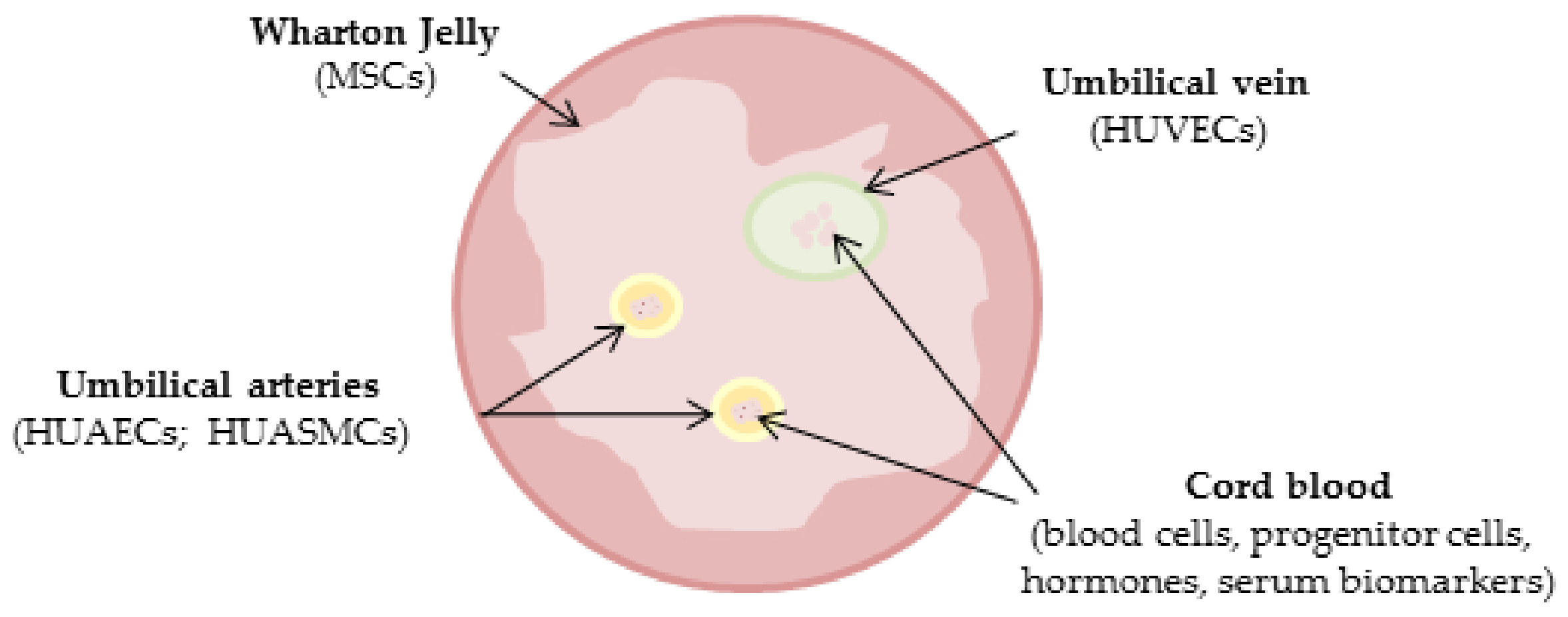
5. Umbilical Cord
6. Sex Differences in Cord Blood Cells, Plasma and Serum

{kind=link}
| Parameters | Source | M vs. F | Comments | Reference |
|---|---|---|---|---|
| Total and free testosterone | serum (venous) | M > F | Dehydroepiandrosterone sulfate from arterial serum > than that from vein only in F | [86] |
| Estradiol | serum (venous) | M > F | [86] | |
| Inhibin | serum (venous) | M > F | [86] | |
| Cortisol and corticosterone | serum (arterial and venous) | M < F | [107,108] | |
| Growth hormone | serum (venous) | M > F | [109] | |
| Leptin | serum (arterial and venous) | M < F | [92] | |
| Insulin and C-peptide | plasma | M < F | [110,111] | |
| 15-F(2t)-isoprostane | plasma | M > F | Premature twins | [93] |
| Gluthatione | umbilical cord vein | M > F | Segments of umbilical cord vein perfused with tert-butylhydroperoxide | [94] |
| Mononuclear cell mitochondrial DNA copies | cord blood | M < F | After prenatal exposure to carbon monoxide as air pollutant | [95] |
| DNA methylation | cord blood | M < F | Number of methylated CpGs sites | [116] |
| Red blood cells | cord blood | M > F | [117,118] | |
| Hematocrit | cord blood | M > F | [117,118] | |
| Hemoglobin | cord blood | M > F | [117,118] | |
| Mean corpuscular hemoglobin concentration | cord blood | M > F | [117,118] | |
| Mean corpuscular volume | cord blood | M < F | [117,118] | |
| Platelets | cord blood | M < F | [117,118] | |
| White blood cells | cord blood | M < F | Lymphocyte, monocyte, eosinophil, basophil > M, neutrophil, metamyelocyte, myelocyte, and promyelocyte ratios > F | [118] |
| CD34+ progenitor cells | cord blood | M > F | M have higher capacity to produce colonies | [119,120] |
| ILC2s | cord blood | M > F | [121] | |
| CD34 + KDR + progenitor cells | cord blood | M < F | [122] |
7. Sex Differences in -HUVECs and HUAECs
8. Sex Differences in HUASMCs
9. Sex Differences in WJ Cells
10. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMH | Anti-Müllerian hormone |
| AKT | protein kinase B |
| BIRC | baculoviral IAP repeat-containing protein |
| BPD | biparietal diameter |
| DNMT1 | DNA-methyltransferase |
| ERα | estrogen receptor alpha |
| ERβ | estrogen receptor beta |
| FGF9 | fibroblast growth factor 9 |
| GH | growth hormone |
| GPER | G protein-coupled estrogen receptor |
| HUAEC | human umbilical artery endothelial cells |
| HUASMC | human umbilical artery smooth muscle cells |
| HUVECs | human umbilical vein endothelial cells |
| IUGR | intrauterine growth restriction |
| LC3 | microtubule-associated protein 1A/1B-light chain 3 |
| MCP1 | monocyte chemoattractant protein 1 |
| MSCs | mesenchymal stem cells |
| mTOR | mammalian target of rapamycin |
| OCT4 | octamer-binding transcription factor 4 |
| PR | progesterone receptor |
| SOX | SRY-box transcription factor |
| SRY | sex-determining region Y |
| SUA | single umbilical artery |
| TGF-β | transforming growth factor-β |
| WJ | Wharton Jelly |
References
- Legato, M.J. Principles of Gender-Specific Medicine. Gender in the Genomic Era, 3rd ed.; Elsevier Academic Press: Cambridge, MA, USA, 2017; p. 792. [Google Scholar]
- Franconi, F.; Campesi, I. Sex impact on biomarkers, pharmacokinetics and pharmacodynamics. Curr. Med. Chem. 2017, 24, 2561–2575. [Google Scholar] [CrossRef]
- Franconi, F.; Campesi, I.; Colombo, D.; Antonini, P. Sex-gender variable: Methodological recommendations for increasing scientific value of clinical studies. Cells 2019, 8, 476. [Google Scholar] [CrossRef]
- Vige, A.; Gallou-Kabani, C.; Junien, C. Sexual dimorphism in non-Mendelian inheritance. Pediatr. Res. 2008, 63, 340–347. [Google Scholar] [CrossRef]
- Franconi, F.; Rosano, G.; Campesi, I. Need for gender-specific pre-analytical testing: The dark side of the moon in laboratory testing. Int. J. Cardiol. 2015, 179, 514–535. [Google Scholar] [CrossRef]
- Tzschoppe, A.; Rauh, M.; Goecke, T.W.; Yazdi, B.; Hart, N.; Siemer, J.; Schild, R.L.; Dotsch, J. Sex-specific differences in the concentration of tubular parameters in the amniotic fluid of second trimester fetuses. Prenat. Diagn. 2012, 32, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Challis, J.; Newnham, J.; Petraglia, F.; Yeganegi, M.; Bocking, A. Fetal sex and preterm birth. Placenta 2013, 34, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Franconi, F.; Brunelleschi, S.; Steardo, L.; Cuomo, V. Gender differences in drug responses. Pharmacol. Res. 2007, 55, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Minucci, D. Lifelong gender health programming in fetal life. Ital. J. Gend. Specif. Med. 2018, 4, 91–100. [Google Scholar]
- Glezerman, M. Intrauterine development of sex differences—fetal programming. In Principles of Gender-Specific Medicine. Gender in the Genomic Era; Legato, M.J., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 237–249. [Google Scholar]
- Shah, K.; McCormack, C.E.; Bradbury, N.A. Do you know the sex of your cells? Am. J. Physiol. Cell Physiol. 2013, 306, C3–C18. [Google Scholar] [CrossRef]
- Taylor, K.E.; Vallejo-Giraldo, C.; Schaible, N.S.; Zakeri, R.; Miller, V.M. Reporting of sex as a variable in cardiovascular studies using cultured cells. Biol. Sex Differ. 2011, 2, 11. [Google Scholar] [CrossRef]
- Beery, A.K.; Zucker, I. Sex bias in neuroscience and biomedical research. Neurosci. Biobehav. Rev. 2011, 35, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Maselli, A.; Matarrese, P.; Straface, E.; Canu, S.; Franconi, F.; Malorni, W. Cell sex: A new look at cell fate studies. FASEB J. 2009, 23, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Addis, R.; Campesi, I.; Fois, M.; Capobianco, G.; Dessole, S.; Fenu, G.; Montella, A.; Cattaneo, M.G.; Vicentini, L.M.; Franconi, F. Human umbilical endothelial cells (HUVECs) have a sex: Characterisation of the phenotype of male and female cells. Biol. Sex Differ. 2014, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Campesi, I.; Capobianco, G.; Dessole, S.; Occhioni, S.; Montella, A.; Franconi, F. Estrogenic compounds have divergent effects on human endothelial progenitor cell migration according to sex of the donor. J. Vasc. Res. 2015, 52, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Campesi, I.; Marino, M.; Montella, A.; Pais, S.; Franconi, F. Sex differences in estrogen receptor alpha and beta levels and activation status in LPS-stimulated human macrophages. J. Cell Physiol. 2016, 232, 340–345. [Google Scholar] [CrossRef]
- Campesi, I.; Sanna, M.; Zinellu, A.; Carru, C.; Rubattu, L.; Bulzomi, P.; Seghieri, G.; Tonolo, G.; Palermo, M.; Rosano, G.; et al. Oral contraceptives modify DNA methylation and monocyte-derived macrophage function. Biol. Sex Differ. 2012, 3, 4. [Google Scholar] [CrossRef]
- Pellegrini, M.; Bulzomi, P.; Lecis, M.; Leone, S.; Campesi, I.; Franconi, F.; Marino, M. Endocrine disruptors differently influence estrogen receptor beta and androgen receptor in male and female rat VSMC. J. Cell Physiol. 2014, 229, 1061–1068. [Google Scholar] [CrossRef]
- McCrohon, J.A.; Death, A.K.; Nakhla, S.; Jessup, W.; Handelsman, D.J.; Stanley, K.K.; Celermajer, D.S. Androgen receptor expression is greater in macrophages from male than from female donors. A sex difference with implications for atherogenesis. Circulation 2000, 101, 224–226. [Google Scholar] [CrossRef]
- Lopez-Ruiz, A.; Sartori-Valinotti, J.; Yanes, L.L.; Iliescu, R.; Reckelhoff, J.F. Sex differences in control of blood pressure: Role of oxidative stress in hypertension in females. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H466–H474. [Google Scholar] [CrossRef]
- Malorni, W.; Straface, E.; Matarrese, P.; Ascione, B.; Coinu, R.; Canu, S.; Galluzzo, P.; Marino, M.; Franconi, F. Redox state and gender differences in vascular smooth muscle cells. FEBS Lett. 2008, 582, 635–642. [Google Scholar] [CrossRef]
- Campesi, I.; Occhioni, S.; Capobianco, G.; Fois, M.; Montella, A.; Dessole, S.; Franconi, F. Sex-specific pharmacological modulation of autophagic process in human umbilical artery smooth muscle cells. Pharmacol. Res. 2016, 113, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.; Laranjeira, P.; Mendes, S.; Velada, I.; Leite, C.; Andrade, P.; Santos, F.; Henriques, A.; Graos, M.; Cardoso, C.M.; et al. Mesenchymal stem cells from umbilical cord matrix, adipose tissue and bone marrow exhibit different capability to suppress peripheral blood B, natural killer and T cells. Stem. Cell Res. Ther. 2013, 4, 125. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.; Reza, H.M. Short review on human umbilical cord lining epithelial cells and their potential clinical applications. Stem. Cell Res. Ther. 2017, 8, 222. [Google Scholar] [CrossRef]
- Parma, P.; Radi, O. Molecular mechanisms of sexual development. Sex Dev. 2012, 6, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Camerino, G.; Parma, P.; Radi, O.; Valentini, S. Sex determination and sex reversal. Curr. Opin. Genet. Dev. 2006, 16, 289–292. [Google Scholar] [CrossRef]
- Patel, S.; Bonora, G.; Sahakyan, A.; Kim, R.; Chronis, C.; Langerman, J.; Fitz-Gibbon, S.; Rubbi, L.; Skelton, R.J.P.; Ardehali, R.; et al. Human embryonic stem cells do not change their X inactivation status during differentiation. Cell Rep. 2017, 18, 54–67. [Google Scholar] [CrossRef]
- Patrat, C.; Ouimette, J.F.; Rougeulle, C. X chromosome inactivation in human development. Development 2020, 147. [Google Scholar] [CrossRef]
- Posynick, B.J.; Brown, C.J. Escape from X-Chromosome inactivation: An evolutionary perspective. Front. Cell Dev. Biol. 2019, 7, 241. [Google Scholar] [CrossRef]
- Berletch, J.B.; Yang, F.; Xu, J.; Carrel, L.; Disteche, C.M. Genes that escape from X inactivation. Hum. Genet. 2011, 130, 237–245. [Google Scholar] [CrossRef]
- Gribnau, J.; Barakat, S.T. X-chromosome inactivation and its implications for human disease. bioRxiv 2017, 076950. [Google Scholar] [CrossRef]
- Schmahl, J.; Eicher, E.M.; Washburn, L.L.; Capel, B. Sry induces cell proliferation in the mouse gonad. Development 2000, 127, 65–73. [Google Scholar] [PubMed]
- Vidal, V.P.; Chaboissier, M.C.; de Rooij, D.G.; Schedl, A. Sox9 induces testis development in XX transgenic mice. Nat. Genet. 2001, 28, 216–217. [Google Scholar] [CrossRef] [PubMed]
- Colvin, J.S.; Green, R.P.; Schmahl, J.; Capel, B.; Ornitz, D.M. Male-to-female sex reversal in mice lacking fibroblast growth factor 9. Cell 2001, 104, 875–889. [Google Scholar] [CrossRef]
- Lubchenco, L.O.; Hansman, C.; Dressler, M.; Boyd, E. Intrauterine Growth as Estimated from Liveborn Birth-Weight Data at 24 to 42 Weeks of Gestation. Pediatrics 1963, 32, 793–800. [Google Scholar]
- Mittwoch, U. Blastocysts prepare for the race to be male. Hum. Reprod. 1993, 8, 1550–1555. [Google Scholar] [CrossRef]
- Pedersen, J.F.; Mantoni, M. Difference in fetal size in the first trimester. Br. Med. J. (Clin. Res. Ed.) 1985, 291, 1278. [Google Scholar] [CrossRef][Green Version]
- Schwarzler, P.; Bland, J.M.; Holden, D.; Campbell, S.; Ville, Y. Sex-specific antenatal reference growth charts for uncomplicated singleton pregnancies at 15-40 weeks of gestation. Ultrasound Obstet. Gynecol. 2004, 23, 23–29. [Google Scholar] [CrossRef]
- Broere-Brown, Z.A.; Baan, E.; Schalekamp-Timmermans, S.; Verburg, B.O.; Jaddoe, V.W.; Steegers, E.A. Sex-specific differences in fetal and infant growth patterns: A prospective population-based cohort study. Biol. Sex Differ. 2016, 7, 65. [Google Scholar] [CrossRef]
- Moore, W.M.; Ward, B.S.; Jones, V.P.; Bamford, F.N. Sex difference in fetal head growth. Br. J. Obstet. Gynaecol. 1988, 95, 238–242. [Google Scholar] [CrossRef]
- Galjaard, S.; Ameye, L.; Lees, C.C.; Pexsters, A.; Bourne, T.; Timmerman, D.; Devlieger, R. Sex differences in fetal growth and immediate birth outcomes in a low-risk Caucasian population. Biol. Sex Differ. 2019, 10, 48. [Google Scholar] [CrossRef]
- Lee, E.; Ping, H. Sex differences in fetal activity and childhood hyperactivity. Res. J. Dev. Biol. 2014, 1, 1–5. [Google Scholar]
- Almli, C.R.; Ball, R.H.; Wheeler, M.E. Human fetal and neonatal movement patterns: Gender differences and fetal-to-neonatal continuity. Dev. Psychobiol. 2001, 38, 252–273. [Google Scholar] [CrossRef] [PubMed]
- DiPietro, J.A.; Hodgson, D.M.; Costigan, K.A.; Hilton, S.C.; Johnson, T.R. Fetal neurobehavioral development. Child. Dev. 1996, 67, 2553–2567. [Google Scholar] [CrossRef] [PubMed]
- Oreal, E.; Mazaud, S.; Picard, J.Y.; Magre, S.; Carre-Eusebe, D. Different patterns of anti-Mullerian hormone expression, as related to DMRT1, SF-1, WT1, GATA-4, Wnt-4, and Lhx9 expression, in the chick differentiating gonads. Dev. Dyn. 2002, 225, 221–232. [Google Scholar] [CrossRef]
- Novembri, R.; Funghi, L.; Voltolini, C.; Belmonte, G.; Vannuccini, S.; Torricelli, M.; Petraglia, F. Placenta expresses anti-Mullerian hormone and its receptor: Sex-related difference in fetal membranes. Placenta 2015, 36, 731–737. [Google Scholar] [CrossRef]
- Eriksson, J.G.; Kajantie, E.; Osmond, C.; Thornburg, K.; Barker, D.J. Boys live dangerously in the womb. Am. J. Hum. Biol. 2010, 22, 330–335. [Google Scholar] [CrossRef]
- Wallace, J.M.; Bhattacharya, S.; Horgan, G.W. Gestational age, gender and parity specific centile charts for placental weight for singleton deliveries in Aberdeen, UK. Placenta 2013, 34, 269–274. [Google Scholar] [CrossRef]
- Ogawa, M.; Matsuda, Y.; Nakai, A.; Hayashi, M.; Sato, S.; Matsubara, S. Standard curves of placental weight and fetal/placental weight ratio in Japanese population: Difference according to the delivery mode, fetal sex, or maternal parity. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 206, 225–231. [Google Scholar] [CrossRef]
- Bermejo-Alvarez, P.; Rizos, D.; Rath, D.; Lonergan, P.; Gutierrez-Adan, A. Epigenetic differences between male and female bovine blastocysts produced in vitro. Physiol. Genom. 2008, 32, 264–272. [Google Scholar] [CrossRef]
- Valdivia, R.P.; Kunieda, T.; Azuma, S.; Toyoda, Y. PCR sexing and developmental rate differences in preimplantation mouse embryos fertilized and cultured in vitro. Mol. Reprod. Dev. 1993, 35, 121–126. [Google Scholar] [CrossRef]
- Ray, P.F.; Conaghan, J.; Winston, R.M.; Handyside, A.H. Increased number of cells and metabolic activity in male human preimplantation embryos following in vitro fertilization. J. Reprod. Fertil. 1995, 104, 165–171. [Google Scholar] [CrossRef]
- Leon-Garcia, S.M.; Roeder, H.A.; Nelson, K.K.; Liao, X.; Pizzo, D.P.; Laurent, L.C.; Parast, M.M.; LaCoursiere, D.Y. Maternal obesity and sex-specific differences in placental pathology. Placenta 2016, 38, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Sheiner, E.; Levy, A.; Katz, M.; Hershkovitz, R.; Leron, E.; Mazor, M. Gender does matter in perinatal medicine. Fetal Diagn. Ther. 2004, 19, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Steier, J.A.; Myking, O.L.; Bergsjo, P.B. Correlation between fetal sex and human chorionic gonadotropin in peripheral maternal blood and amniotic fluid in second and third trimester normal pregnancies. Acta Obstet. Gynecol. Scand. 1999, 78, 367–371. [Google Scholar] [PubMed]
- Clifton, V.L.; Bisits, A.; Zarzycki, P.K. Characterization of human fetal cord blood steroid profiles in relation to fetal sex and mode of delivery using temperature-dependent inclusion chromatography and principal component analysis (PCA). J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 855, 249–254. [Google Scholar] [CrossRef]
- Goldenberg, R.L.; Andrews, W.W.; Faye-Petersen, O.M.; Goepfert, A.R.; Cliver, S.P.; Hauth, J.C. The Alabama Preterm Birth Study: Intrauterine infection and placental histologic findings in preterm births of males and females less than 32 weeks. Am. J. Obstet. Gynecol. 2006, 195, 1533–1537. [Google Scholar] [CrossRef]
- Scott, N.M.; Hodyl, N.A.; Murphy, V.E.; Osei-Kumah, A.; Wyper, H.; Hodgson, D.M.; Smith, R.; Clifton, V.L. Placental cytokine expression covaries with maternal asthma severity and fetal sex. J. Immunol. 2009, 182, 1411–1420. [Google Scholar] [CrossRef]
- Widnes, C.; Flo, K.; Wilsgaard, T.; Kiserud, T.; Acharya, G. Sex differences in umbilical artery Doppler indices: A longitudinal study. Biol. Sex Differ. 2018, 9, 16. [Google Scholar] [CrossRef]
- Dawes, N.W.; Dawes, G.S.; Moulden, M.; Redman, C.W. Fetal heart rate patterns in term labor vary with sex, gestational age, epidural analgesia, and fetal weight. Am. J. Obstet. Gynecol. 1999, 180, 181–187. [Google Scholar] [CrossRef]
- Bale, T.L. The placenta and neurodevelopment: Sex differences in prenatal vulnerability. Dialogues Clin. Neurosci. 2016, 18, 459–464. [Google Scholar]
- Sood, R.; Zehnder, J.L.; Druzin, M.L.; Brown, P.O. Gene expression patterns in human placenta. Proc. Natl. Acad. Sci. USA 2006, 103, 5478–5483. [Google Scholar] [CrossRef] [PubMed]
- Buckberry, S.; Bianco-Miotto, T.; Bent, S.J.; Dekker, G.A.; Roberts, C.T. Integrative transcriptome meta-analysis reveals widespread sex-biased gene expression at the human fetal-maternal interface. Mol. Hum. Reprod. 2014, 20, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Cvitic, S.; Longtine, M.S.; Hackl, H.; Wagner, K.; Nelson, M.D.; Desoye, G.; Hiden, U. The human placental sexome differs between trophoblast epithelium and villous vessel endothelium. PLoS ONE 2013, 8, e79233. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. Intrauterine programming of adult disease. Mol. Med. Today 1995, 1, 418–423. [Google Scholar] [CrossRef]
- Di Renzo, G.C.; Rosati, A.; Sarti, R.D.; Cruciani, L.; Cutuli, A.M. Does fetal sex affect pregnancy outcome? Gend. Med. 2007, 4, 19–30. [Google Scholar] [CrossRef]
- Verburg, P.E.; Tucker, G.; Scheil, W.; Erwich, J.J.; Dekker, G.A.; Roberts, C.T. Sexual Dimorphism in Adverse Pregnancy Outcomes—A Retrospective Australian Population Study 1981–2011. PLoS ONE 2016, 11, e0158807. [Google Scholar] [CrossRef]
- Martinez, D.; Pentinat, T.; Ribo, S.; Daviaud, C.; Bloks, V.W.; Cebria, J.; Villalmanzo, N.; Kalko, S.G.; Ramon-Krauel, M.; Diaz, R.; et al. In utero undernutrition in male mice programs liver lipid metabolism in the second-generation offspring involving altered Lxra DNA methylation. Cell Metab. 2014, 19, 941–951. [Google Scholar] [CrossRef]
- Moritz, K.M.; Cuffe, J.S.; Wilson, L.B.; Dickinson, H.; Wlodek, M.E.; Simmons, D.G.; Denton, K.M. Review: Sex specific programming: A critical role for the renal renin-angiotensin system. Placenta 2010, 31, S40–S46. [Google Scholar] [CrossRef]
- Grigore, D.; Ojeda, N.B.; Alexander, B.T. Sex differences in the fetal programming of hypertension. Gend. Med. 2008, 5 (Suppl. A), S121–S132. [Google Scholar] [CrossRef]
- Dasinger, J.H.; Alexander, B.T. Gender differences in developmental programming of cardiovascular diseases. Clin. Sci. (Lond.) 2016, 130, 337–348. [Google Scholar] [CrossRef]
- Clifton, V.L. Review: Sex and the human placenta: Mediating differential strategies of fetal growth and survival. Placenta 2010, 31, S33–S39. [Google Scholar] [CrossRef] [PubMed]
- Murphy, V.E.; Gibson, P.; Talbot, P.I.; Clifton, V.L. Severe asthma exacerbations during pregnancy. Obstet. Gynecol. 2005, 106, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.M.; Hulme, E.; Devendrakumar, V.; Turner, M.A.; Baker, P.N.; Sibley, C.P.; D’Souza, S.W. Effect of maternal asthma on birthweight and neonatal outcome in a British inner-city population. Paediatr. Perinat. Epidemiol. 2007, 21, 154–162. [Google Scholar] [CrossRef]
- Stark, M.J.; Clifton, V.L.; Wright, I.M. Neonates born to mothers with preeclampsia exhibit sex-specific alterations in microvascular function. Pediatr. Res. 2009, 65, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Stark, M.J.; Dierkx, L.; Clifton, V.L.; Wright, I.M. Alterations in the maternal peripheral microvascular response in pregnancies complicated by preeclampsia and the impact of fetal sex. J. Soc. Gynecol. Investig. 2006, 13, 573–578. [Google Scholar] [CrossRef]
- Sandman, C.A.; Glynn, L.M.; Davis, E.P. Is there a viability-vulnerability tradeoff? Sex differences in fetal programming. J. Psychosom. Res. 2013, 75, 327–335. [Google Scholar] [CrossRef]
- Wells, J.C. Natural selection and sex differences in morbidity and mortality in early life. J. Theor. Biol. 2000, 202, 65–76. [Google Scholar] [CrossRef]
- Cooperstock, M.; Campbell, J. Excess males in preterm birth: Interactions with gestational age, race, and multiple birth. Obstet. Gynecol. 1996, 88, 189–193. [Google Scholar] [CrossRef]
- Spurway, J.; Logan, P.; Pak, S. The development, structure and blood flow within the umbilical cord with particular reference to the venous system. Australas. J. Ultrasound Med. 2012, 15, 97–102. [Google Scholar] [CrossRef]
- Meyer, W.W.; Rumpelt, H.J.; Yao, A.C.; Lind, J. Structure and closure mechanism of the human umbilical artery. Eur. J. Pediatr. 1978, 128, 247–259. [Google Scholar] [CrossRef]
- Tavares Fortuna, J.F.; Lourdes Pratas, M. Coarctation of the umbilical cord: A cause of intrauterine fetal death. Int. J. Gynaecol. Obstet. 1978, 15, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Wharton, T.W. Adenographia; Freer, S., Ed.; Oxford University Press: Oxford, UK, 1996; p. 243. [Google Scholar]
- Takechi, K.; Kuwabara, Y.; Mizuno, M. Ultrastructural and immunohistochemical studies of Wharton’s jelly umbilical cord cells. Placenta 1993, 14, 235–245. [Google Scholar] [CrossRef]
- Kim, D.W.; Staples, M.; Shinozuka, K.; Pantcheva, P.; Kang, S.D.; Borlongan, C.V. Wharton’s jelly-derived mesenchymal stem cells: Phenotypic characterization and optimizing their therapeutic potential for clinical applications. Int. J. Mol. Sci. 2013, 14, 11692–11712. [Google Scholar] [CrossRef] [PubMed]
- Geipel, A.; Germer, U.; Welp, T.; Schwinger, E.; Gembruch, U. Prenatal diagnosis of single umbilical artery: Determination of the absent side, associated anomalies, Doppler findings and perinatal outcome. Ultrasound Obstet. Gynecol. 2000, 15, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Zhai, S.; Shi, N.; Li, M.; Cui, S.; Xu, Y.; Ran, L.; Ren, L.; Hong, T.; Liu, R. The risk factors and neonatal outcomes of isolated single umbilical artery in singleton pregnancy: A meta-analysis. Sci. Rep. 2017, 7, 7396. [Google Scholar] [CrossRef] [PubMed]
- Mu, S.C.; Lin, C.H.; Chen, Y.L.; Sung, T.C.; Bai, C.H.; Jow, G.M. The perinatal outcomes of asymptomatic isolated single umbilical artery in full-term neonates. Pediatr. Neonatol. 2008, 49, 230–233. [Google Scholar] [CrossRef]
- Elghazaly, A.E.; Jameel, A. Morphological features of the umbilical cord vessels at the delivery, in sudanese neonates. Int. J. Anat. Res. 2020, 8, 7200–7206. [Google Scholar] [CrossRef]
- Fujikura, T. Fused umbilical arteries near placental cord insertion. Am. J. Obstet. Gynecol. 2003, 188, 765–767. [Google Scholar] [CrossRef]
- Heifetz, S.A. Single umbilical artery. A statistical analysis of 237 autopsy cases and review of the literature. Perspect. Pediatr. Pathol. 1984, 8, 345–378. [Google Scholar]
- Lilja, M. Infants with single umbilical artery studied in a national registry. General epidemiological characteristics. Paediatr. Perinat. Epidemiol. 1991, 5, 27–36. [Google Scholar] [CrossRef]
- Chitra, T.; Sushanth, Y.S.; Raghavan, S. Umbilical coiling index as a marker of perinatal outcome: An analytical study. Obstet. Gynecol. Int. 2012, 2012, 213689. [Google Scholar] [CrossRef] [PubMed]
- Ochshorn, Y.; Ascher Landsberg, J.; Many, A.; Maslovitz, S.; Rimon, E.; Yogev, Y. Fetal gender and umbilical cord characteristics at birth. J. Matern. Fetal Neonatal Med. 2019, 1–4. [Google Scholar] [CrossRef]
- Barnwal, M.; Rathi, S.; Chhabra, S.; Nanda, S. Histomorphometry of umbilical cord and its vessels in pre- eclampsia as compared to normal pregnancies. Nepal J. Obstet. Gynaecol. 2013, 7, 28–32. [Google Scholar] [CrossRef]
- Rua Ede, A.; Porto, M.L.; Ramos, J.P.; Nogueira, B.V.; Meyrelles, S.S.; Vasquez, E.C.; Pereira, T.C. Effects of tobacco smoking during pregnancy on oxidative stress in the umbilical cord and mononuclear blood cells of neonates. J. Biomed. Sci. 2014, 21, 105. [Google Scholar] [PubMed]
- Milnerowicz-Nabzdyk, E.; Zimmer, M.; Tlolka, J.; Michniewicz, J.; Pomorski, M.; Wiatrowski, A. Umbilical cord morphology in pregnancies complicated by IUGR in cases of tobacco smoking and pregnancy-induced hypertension. Neuro Endocrinol. Lett. 2010, 31, 842–847. [Google Scholar]
- Broere-Brown, Z.A.; Adank, M.C.; Benschop, L.; Tielemans, M.; Muka, T.; Goncalves, R.; Bramer, W.M.; Schoufour, J.D.; Voortman, T.; Steegers, E.A.P.; et al. Fetal sex and maternal pregnancy outcomes: A systematic review and meta-analysis. Biol. Sex Differ. 2020, 11, 26. [Google Scholar] [CrossRef]
- Mittelstrass, K.; Ried, J.S.; Yu, Z.; Krumsiek, J.; Gieger, C.; Prehn, C.; Roemisch-Margl, W.; Polonikov, A.; Peters, A.; Theis, F.J.; et al. Discovery of sexual dimorphisms in metabolic and genetic biomarkers. PLoS Genet. 2011, 7, e1002215. [Google Scholar] [CrossRef]
- Caterino, M.; Ruoppolo, M.; Villani, G.R.D.; Marchese, E.; Costanzo, M.; Sotgiu, G.; Dore, S.; Franconi, F.; Campesi, I. Influence of sex on urinary organic acids: A cross-sectional study in children. Int. J. Mol. Sci. 2020, 21, 582. [Google Scholar] [CrossRef]
- Ruoppolo, M.; Campesi, I.; Scolamiero, E.; Pecce, R.; Caterino, M.; Cherchi, S.; Mercuro, G.; Tonolo, G.; Franconi, F. Serum metabolomic profiles suggest influence of sex and oral contraceptive use. Am. J. Transl. Res. 2014, 6, 614–624. [Google Scholar]
- Ruoppolo, M.; Scolamiero, E.; Caterino, M.; Mirisola, V.; Franconi, F.; Campesi, I. Female and male human babies have distinct blood metabolomic patterns. Mol. Biosyst. 2015, 11, 2483–2492. [Google Scholar] [CrossRef]
- Ishikawa, M.; Maekawa, K.; Saito, K.; Senoo, Y.; Urata, M.; Murayama, M.; Tajima, Y.; Kumagai, Y.; Saito, Y. Plasma and serum lipidomics of healthy white adults shows characteristic profiles by subjects’ gender and age. PLoS ONE 2014, 9, e91806. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.E.; Evans, A.M.; Chace, D.H.; Fornasini, G. Determination of the reference range of endogenous plasma carnitines in healthy adults. Ann. Clin. Biochem. 2008, 45, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.; France, J.T.; Keelan, J.A.; Song, L.; Knox, B.S. Sex differences in umbilical cord serum levels of inhibin, testosterone, oestradiol, dehydroepiandrosterone sulphate, and sex hormone-binding globulin in human term neonates. Biol. Neonate 1994, 65, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Wynne-Edwards, K.E.; Lee, K.; Zhou, R.; Edwards, H.E. Sex differences in substrates and clearance products of cortisol and corticosterone synthesis in full-term human umbilical circulation without labor: Substrate depletion matches synthesis in males, but not females. Psychoneuroendocrinology 2019, 109, 104381. [Google Scholar] [CrossRef] [PubMed]
- Giesbrecht, G.F.; Rash, J.A.; Edwards, H.E.; Wynne-Edwards, K.E. Full-term deliveries without antecedent labor reveal sex differences in umbilical cord glucocorticoid concentrations. Psychoneuroendocrinology 2016, 74, 121–125. [Google Scholar] [CrossRef]
- Fukaya, T.; Furuhashi, N.; Kono, H.; Tachibana, Y.; Shinkawa, O.; Takahashi, T.; Suzuki, M. Secretion and Sex differences in cord serum growth hormone levels of the human fetus (author’s transl). Nihon Sanka Fujinka Gakkai Zasshi 1982, 34, 491–496. [Google Scholar]
- Kuchay, M.S.; Kudyar, R.P.; Gupta, A.; Pandita, K.K.; Ganie, M.A. Gender differences in insulin and C-peptide concentrations at birth using cord blood collection. Arch. Endocrinol. Metab. 2016, 60, 264–266. [Google Scholar] [CrossRef]
- Shields, B.M.; Knight, B.; Hopper, H.; Hill, A.; Powell, R.J.; Hattersley, A.T.; Clark, P.M. Measurement of cord insulin and insulin-related peptides suggests that girls are more insulin resistant than boys at birth. Diabetes Care 2007, 30, 2661–2666. [Google Scholar] [CrossRef]
- Tome, M.A.; Lage, M.; Camina, J.P.; Garcia-Mayor, R.V.; Dieguez, C.; Casanueva, F.F. Sex-based differences in serum leptin concentrations from umbilical cord blood at delivery. Eur. J. Endocrinol. 1997, 137, 655–658. [Google Scholar] [CrossRef]
- Minghetti, L.; Greco, A.; Zanardo, V.; Suppiej, A. Early-life sex-dependent vulnerability to oxidative stress: The natural twining model. J. Matern. Fetal Neonatal. Med. 2013, 26, 259–262. [Google Scholar] [CrossRef]
- Lavoie, J.C.; Chessex, P. Gender-related response to a tert-butyl hydroperoxide-induced oxidation in human neonatal tissue. Free Radic. Biol. Med. 1994, 16, 307–313. [Google Scholar] [CrossRef]
- Kaali, S.; Jack, D.; Delimini, R.; Hu, L.; Burkart, K.; Opoku-Mensah, J.; Quinn, A.; Ae-Ngibise, K.A.; Wylie, B.; Boamah-Kaali, E.A.; et al. Prenatal Household Air Pollution Alters Cord Blood Mononuclear Cell Mitochondrial DNA Copy Number: Sex-Specific Associations. Int. J. Environ. Res. Public Health 2019, 16, 26. [Google Scholar] [CrossRef]
- Maschietto, M.; Bastos, L.C.; Tahira, A.C.; Bastos, E.P.; Euclydes, V.L.; Brentani, A.; Fink, G.; de Baumont, A.; Felipe-Silva, A.; Francisco, R.P.; et al. Sex differences in DNA methylation of the cord blood are related to sex-bias psychiatric diseases. Sci. Rep. 2017, 7, 44547. [Google Scholar] [CrossRef] [PubMed]
- Glasser, L.; Sutton, N.; Schmeling, M.; Machan, J.T. A comprehensive study of umbilical cord blood cell developmental changes and reference ranges by gestation, gender and mode of delivery. J. Perinatol. 2015, 35, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.H.; Yang, S.H.; Wang, T.F.; Lin, T.Y.; Yang, K.L.; Chen, S.H. Complete blood count reference values of cord blood in Taiwan and the influence of gender and delivery route on them. Pediatr. Neonatol. 2011, 52, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Che, Z.; Zhang, X.; Zhou, P.; Li, X.; Xu, X.; Shi, Q.; Li, D.; Ju, X. Influence of neonatal gender on cord blood CD34(+) cell amplification and gene expression. Exp. Ther. Med. 2019, 18, 105–118. [Google Scholar] [CrossRef]
- Aroviita, P.; Teramo, K.; Hiilesmaa, V.; Kekomaki, R. Cord blood hematopoietic progenitor cell concentration and infant sex. Transfusion 2005, 45, 613–621. [Google Scholar] [CrossRef]
- Forsberg, A.; Bengtsson, M.; Eringfalt, A.; Ernerudh, J.; Mjosberg, J.; Jenmalm, M.C. GATA binding protein 3(+) group 2 innate lymphoid cells are present in cord blood and in higher proportions in male than in female neonates. J. Allergy Clin. Immunol. 2014, 134, 228–230. [Google Scholar] [CrossRef]
- Fadini, G.P.; de Kreutzenberg, S.; Albiero, M.; Coracina, A.; Pagnin, E.; Baesso, I.; Cignarella, A.; Bolego, C.; Plebani, M.; Nardelli, G.B.; et al. Gender differences in endothelial progenitor cells and cardiovascular risk profile: The role of female estrogens. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 997–1004. [Google Scholar] [CrossRef]
- Onat, D.; Brillon, D.; Colombo, P.C.; Schmidt, A.M. Human vascular endothelial cells: A model system for studying vascular inflammation in diabetes and atherosclerosis. Curr. Diab. Rep. 2011, 11, 193–202. [Google Scholar] [CrossRef]
- Cines, D.B.; Pollak, E.S.; Buck, C.A.; Loscalzo, J.; Zimmerman, G.A.; McEver, R.P.; Pober, J.S.; Wick, T.M.; Konkle, B.A.; Schwartz, B.S.; et al. Endothelial cells in physiology and in the pathophysiology of vascular disorders. Blood 1998, 91, 3527–3561. [Google Scholar] [PubMed]
- Franconi, F.; Rosano, G.; Basili, S.; Montella, A.; Campesi, I. Human cells involved in atherosclerosis have a sex. Int. J. Cardiol. 2016, 228, 983–1001. [Google Scholar] [CrossRef] [PubMed]
- Koller, A. Perspectives: Microvascular endothelial dysfunction and gender. Eur. Heart J. Suppl. 2014, 16, A16–A19. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Hu, J.; Dong, Y.; Zhan, R.; Li, P.; Su, H.; Peng, Q.; Wu, T.; Lei, L.; Huang, X.; et al. Gender differences in the risk factors for endothelial dysfunction in Chinese hypertensive patients: Homocysteine is an independent risk factor in females. PLoS ONE 2015, 10, e0118686. [Google Scholar] [CrossRef]
- Cattaneo, M.G.; Vanetti, C.; Decimo, I.; Di Chio, M.; Martano, G.; Garrone, G.; Bifari, F.; Vicentini, L.M. Sex-specific eNOS activity and function in human endothelial cells. Sci. Rep. 2017, 7, 9612. [Google Scholar] [CrossRef]
- Oczypok, E.A.; Oury, T.D.; Chu, C.T. It’s a cell-eat-cell world: Autophagy and phagocytosis. Am. J. Pathol. 2013, 182, 612–622. [Google Scholar] [CrossRef]
- De Meyer, G.R.; Martinet, W. Autophagy in the cardiovascular system. Biochim. Biophys. Acta 2009, 1793, 1485–1495. [Google Scholar] [CrossRef]
- Cattaneo, M.G.; Banfi, C.; Brioschi, M.; Lattuada, D.; Vicentini, L.M. Sex-Dependent Differences in the Secretome of Human Endothelial Cells. Biol. Sex Differ. 2021, 12, 7. [Google Scholar] [CrossRef]
- Annibalini, G.; Agostini, D.; Calcabrini, C.; Martinelli, C.; Colombo, E.; Guescini, M.; Tibollo, P.; Stocchi, V.; Sestili, P. Effects of sex hormones on inflammatory response in male and female vascular endothelial cells. J. Endocrinol. Investig. 2014, 37, 861–869. [Google Scholar] [CrossRef]
- Toth, B.; Saadat, G.; Geller, A.; Scholz, C.; Schulze, S.; Friese, K.; Jeschke, U. Human umbilical vascular endothelial cells express estrogen receptor beta (ERbeta) and progesterone receptor A (PR-A), but not ERalpha and PR-B. Histochem. Cell Biol. 2008, 130, 399–405. [Google Scholar] [CrossRef]
- Batres, R.O.; Dupont, J. Gender differences in prostacyclin and prostaglandin E2 synthesis by human endothelial cells. Prostaglandins Leukot. Med. 1986, 22, 159–171. [Google Scholar] [CrossRef]
- Matarrese, P.; Colasanti, T.; Ascione, B.; Margutti, P.; Franconi, F.; Alessandri, C.; Conti, F.; Riccieri, V.; Rosano, G.; Ortona, E.; et al. Gender disparity in susceptibility to oxidative stress and apoptosis induced by autoantibodies specific to RLIP76 in vascular cells. Antioxid. Redox Signal. 2011, 15, 2825–2836. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, M.; Koschate, J.; Kaufmann, K.; Kreye, C.; Mertens, M.; Kuebler, W.M.; Baumann, G.; Gossing, G.; Marki, A.; Zakrzewicz, A.; et al. Does cellular sex matter? Dimorphic transcriptional differences between female and male endothelial cells. Atherosclerosis 2015, 240, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lingappan, K. Differential sex-specific effects of oxygen toxicity in human umbilical vein endothelial cells. Biochem. Biophys. Res. Commun. 2017, 486, 431–437. [Google Scholar] [CrossRef]
- Wijchers, P.J.; Festenstein, R.J. Epigenetic regulation of autosomal gene expression by sex chromosomes. Trends Genet. 2011, 27, 132–140. [Google Scholar] [CrossRef]
- Wilson, M.A. Searching for sex differences. Science 2020, 369, 1298–1299. [Google Scholar] [CrossRef]
- Lopes-Ramos, C.M.; Chen, C.-Y.; Kuijjer, M.L.; Paulson, J.N.; Sonawane, A.R.; Fagny, M.; Platig, J.; Glass, K.; Quackenbush, J.; DeMeo, D.L. Sex Differences in Gene Expression and Regulatory Networks across 29 Human Tissues. Cell Rep. 2020, 31, 107795. [Google Scholar] [CrossRef]
- Oettel, A.; Lorenz, M.; Stangl, V.; Costa, S.D.; Zenclussen, A.C.; Schumacher, A. Human umbilical vein endothelial cells foster conversion of CD4+CD25-Foxp3- T cells into CD4+Foxp3+ regulatory T cells via Transforming Growth Factor-beta. Sci. Rep. 2016, 6, 23278. [Google Scholar] [CrossRef]
- Lavoie, J.C.; Chessex, P. Gender and maturation affect glutathione status in human neonatal tissues. Free Radic. Biol. Med. 1997, 23, 648–657. [Google Scholar] [CrossRef]
- Hartman, R.J.G.; Kapteijn, D.M.C.; Haitjema, S.; Bekker, M.N.; Mokry, M.; Pasterkamp, G.; Civelek, M.; den Ruijter, H.M. Intrinsic transcriptomic sex differences in human endothelial cells at birth and in adults are associated with coronary artery disease targets. Sci. Rep. 2020, 10, 12367. [Google Scholar] [CrossRef]
- Montague, C.R.; Hunter, M.G.; Gavrilin, M.A.; Phillips, G.S.; Goldschmidt-Clermont, P.J.; Marsh, C.B. Activation of estrogen receptor-alpha reduces aortic smooth muscle differentiation. Circ. Res. 2006, 99, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Feng, Z.; Cui, S.; Hou, K.; Tang, L.; Zhou, J.; Cai, G.; Xie, Y.; Hong, Q.; Fu, B.; et al. Rapamycin upregulates autophagy by inhibiting the mTOR-ULK1 pathway, resulting in reduced podocyte injury. PLoS ONE 2013, 8, e63799. [Google Scholar] [CrossRef] [PubMed]
- Morey, A.K.; Pedram, A.; Razandi, M.; Prins, B.A.; Hu, R.M.; Biesiada, E.; Levin, E.R. Estrogen and progesterone inhibit vascular smooth muscle proliferation. Endocrinology 1997, 138, 3330–3339. [Google Scholar] [CrossRef] [PubMed]
- McElreavey, K.D.; Irvine, A.I.; Ennis, K.T.; McLean, W.H. Isolation, culture and characterisation of fibroblast-like cells derived from the Wharton’s jelly portion of human umbilical cord. Biochem. Soc. Trans. 1991, 19, 29S. [Google Scholar] [CrossRef] [PubMed]
- Corotchi, M.C.; Popa, M.A.; Simionescu, M. Testosterone stimulates proliferation and preserves stemness of human adult mesenchymal stem cells and endothelial progenitor cells. Rom. J. Morphol. Embryol. 2016, 57, 75–80. [Google Scholar]
- Mihai, M.C.; Popa, M.A.; Suica, V.I.; Antohe, F.; Jackson, E.K.; Simionescu, M.; Dubey, R.K. Mechanism of 17beta-estradiol stimulated integration of human mesenchymal stem cells in heart tissue. J. Mol. Cell Cardiol. 2019, 133, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Gao, J.; Jiang, Y.; Sun, B.; Lu, W.; Su, M.; Xu, Y.; Yang, X.; Zhang, Y. Differentiation of human umbilical cord Wharton’s jelly-derived mesenchymal stem cells into endometrial cells. Stem. Cell Res. Ther. 2017, 8, 246. [Google Scholar] [CrossRef]
- Tajiri, N.; Duncan, K.; Borlongan, M.C.; Pabon, M.; Acosta, S.; de la Pena, I.; Hernadez-Ontiveros, D.; Lozano, D.; Aguirre, D.; Reyes, S.; et al. Adult stem cell transplantation: Is gender a factor in stemness? Int. J. Mol. Sci. 2014, 15, 15225–15243. [Google Scholar] [CrossRef]
- Maioli, M.; Contini, G.; Santaniello, S.; Bandiera, P.; Pigliaru, G.; Sanna, R.; Rinaldi, S.; Delitala, A.P.; Montella, A.; Bagella, L.; et al. Amniotic fluid stem cells morph into a cardiovascular lineage: Analysis of a chemically induced cardiac and vascular commitment. Drug. Des. Devel. Ther. 2013, 7, 1063–1073. [Google Scholar]
- Balzano, F.; Bellu, E.; Basoli, V.; Dei Giudici, S.; Santaniello, S.; Cruciani, S.; Facchin, F.; Oggiano, A.; Capobianco, G.; Dessole, F.; et al. Lessons from human umbilical cord: Gender differences in stem cells from Wharton’s jelly. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 234, 143–148. [Google Scholar] [CrossRef]
- Balzano, F.; Campesi, I.; Cruciani, S.; Garroni, G.; Bellu, E.; Dei Giudici, S.; Angius, A.; Oggiano, A.; Rallo, V.; Capobianco, G.; et al. Epigenetics, stem cells, and autophagy: Exploring a path involving miRNA. Int. J. Mol. Sci. 2019, 20, 5091. [Google Scholar] [CrossRef] [PubMed]
- Gil-Kulik, P.; Swistowska, M.; Kondracka, A.; Chomik, P.; Krzyzanowski, A.; Kwasniewska, A.; Rahnama, M.; Kocki, J. Increased expression of BIRC2, BIRC3, and BIRC5 from the IAP family in mesenchymal stem cells of the umbilical cord Wharton’s jelly (WJSC) in younger women giving birth naturally. Oxid. Med. Cell. Longev. 2020, 2020, 9084730. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campesi, I.; Franconi, F.; Montella, A.; Dessole, S.; Capobianco, G. Human Umbilical Cord: Information Mine in Sex-Specific Medicine. Life 2021, 11, 52. https://doi.org/10.3390/life11010052
Campesi I, Franconi F, Montella A, Dessole S, Capobianco G. Human Umbilical Cord: Information Mine in Sex-Specific Medicine. Life. 2021; 11(1):52. https://doi.org/10.3390/life11010052
Chicago/Turabian StyleCampesi, Ilaria, Flavia Franconi, Andrea Montella, Salvatore Dessole, and Giampiero Capobianco. 2021. "Human Umbilical Cord: Information Mine in Sex-Specific Medicine" Life 11, no. 1: 52. https://doi.org/10.3390/life11010052
APA StyleCampesi, I., Franconi, F., Montella, A., Dessole, S., & Capobianco, G. (2021). Human Umbilical Cord: Information Mine in Sex-Specific Medicine. Life, 11(1), 52. https://doi.org/10.3390/life11010052