Studying GGDEF Domain in the Act: Minimize Conformational Frustration to Prevent Artefacts



 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
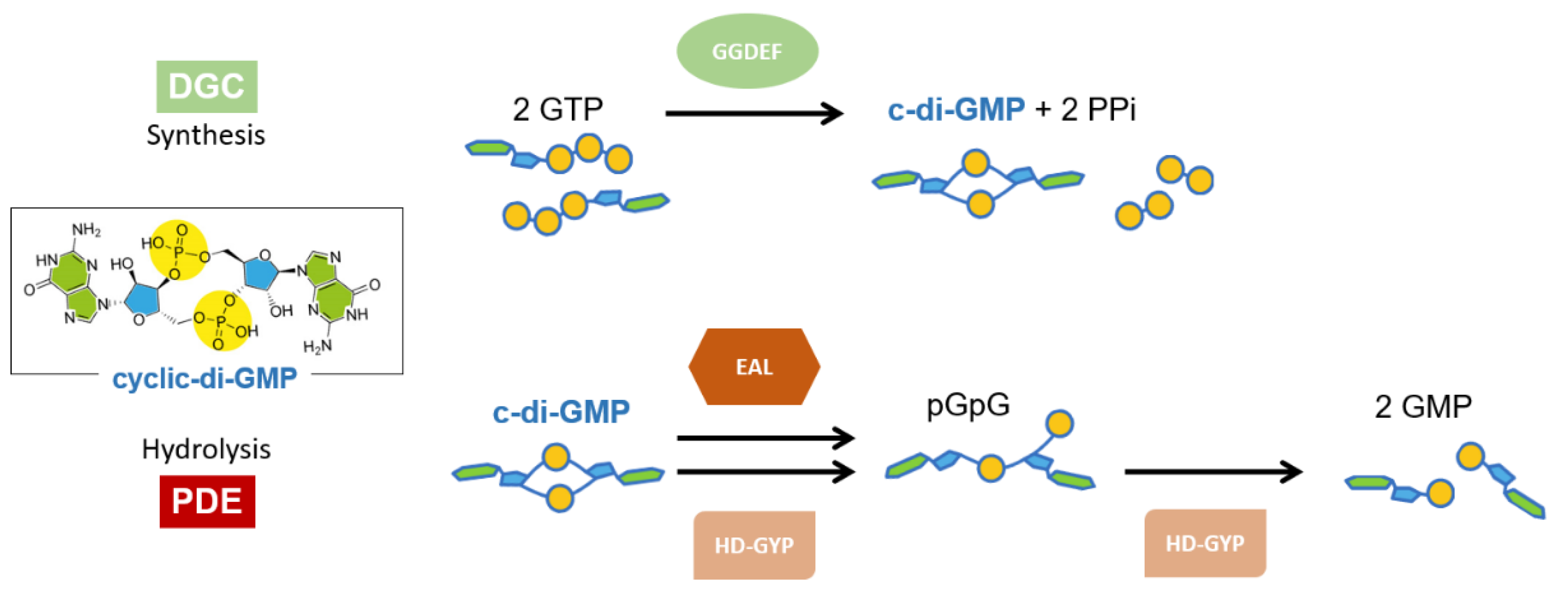
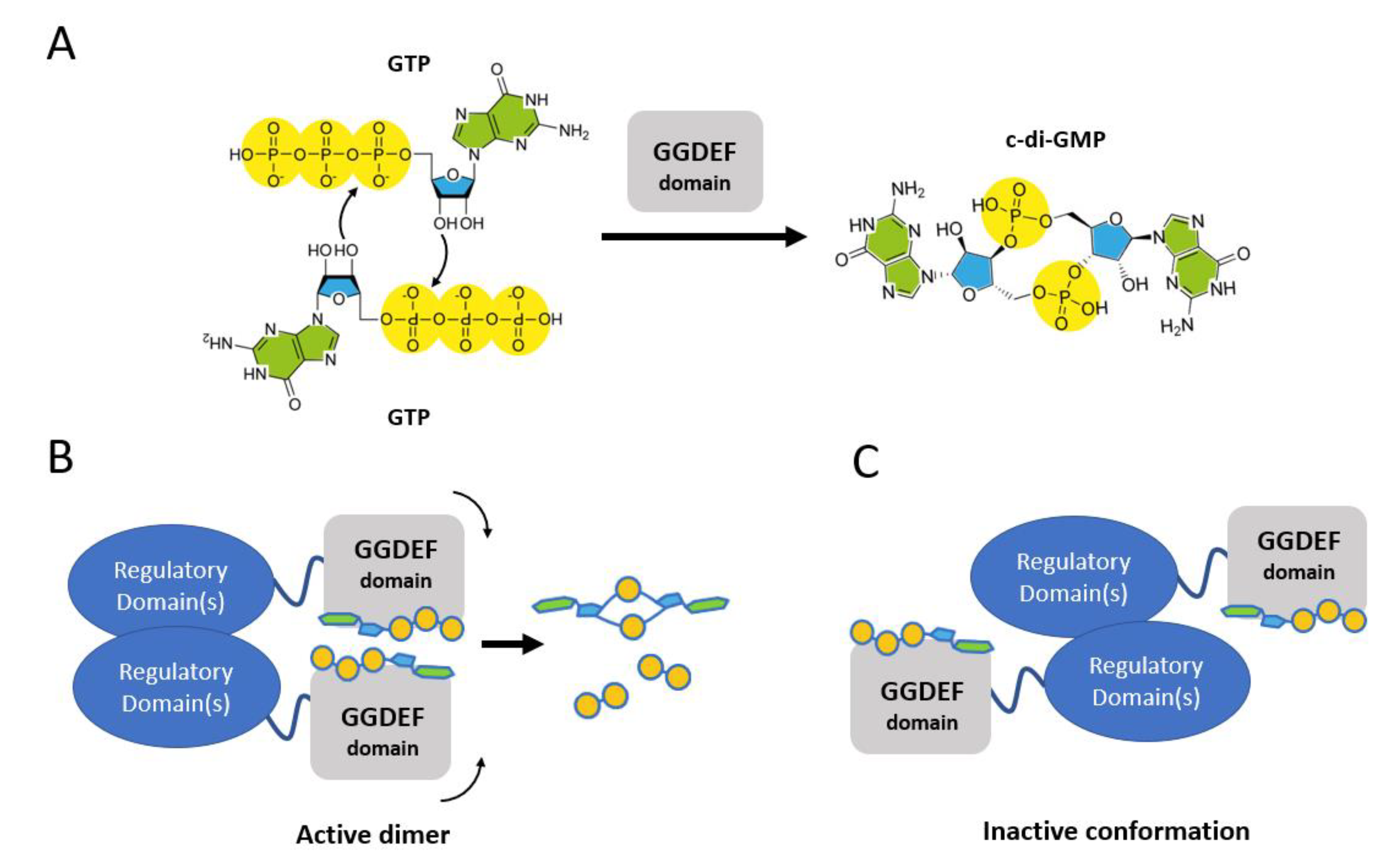
1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. Kinetic Assay GTPase Activity
2.3. Optimization of GMP Separation by RP-HPLC for Mass Spectrometry
2.4. Crystallography
3. Results
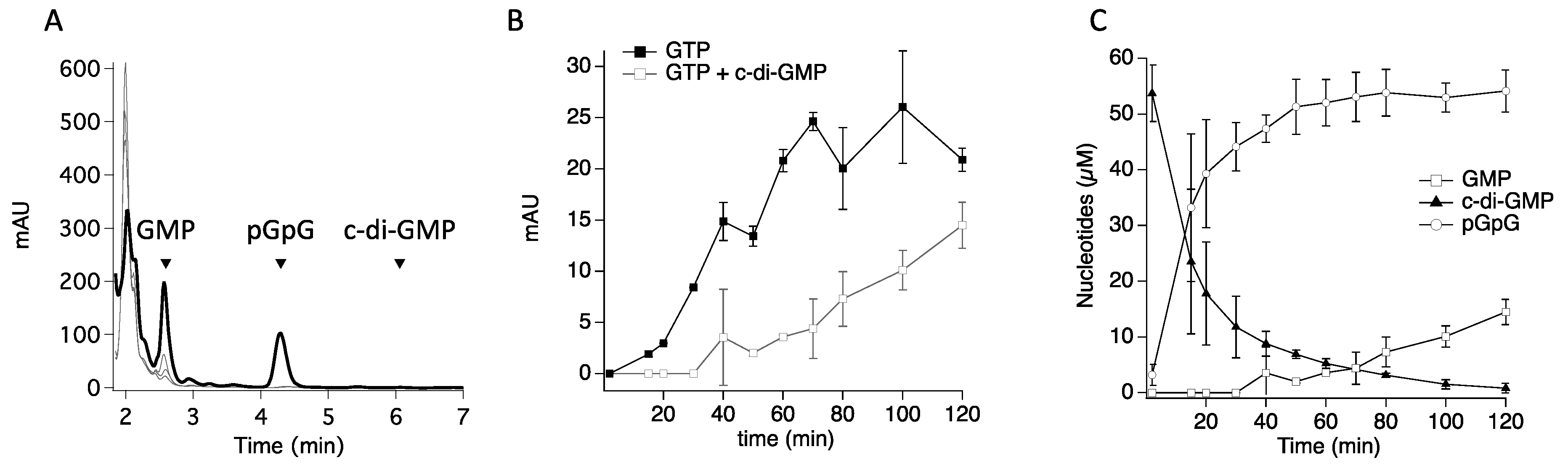
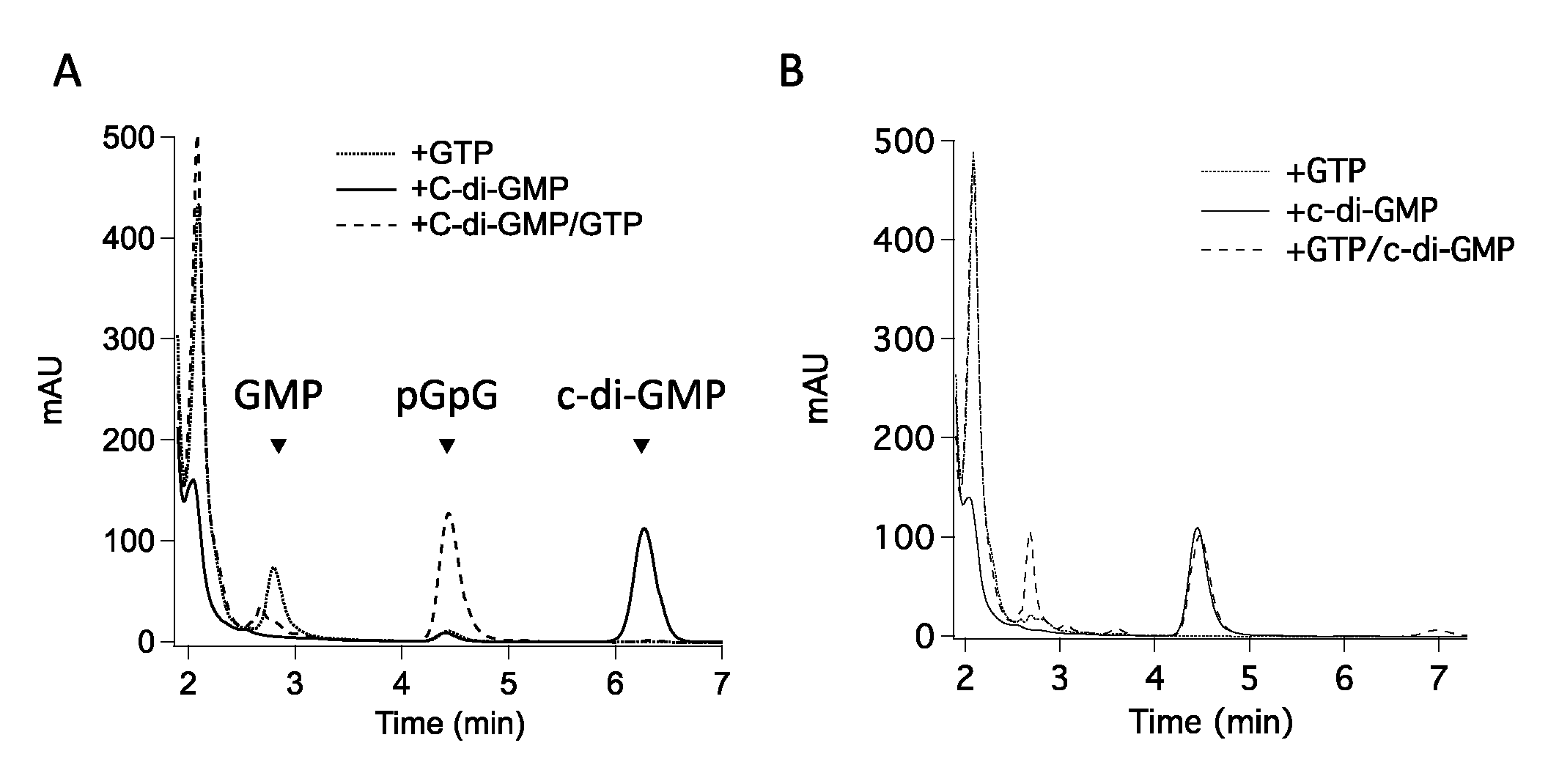
3.1. The GGDEF Domain of RmcA Is Able to Hydrolyze GTP
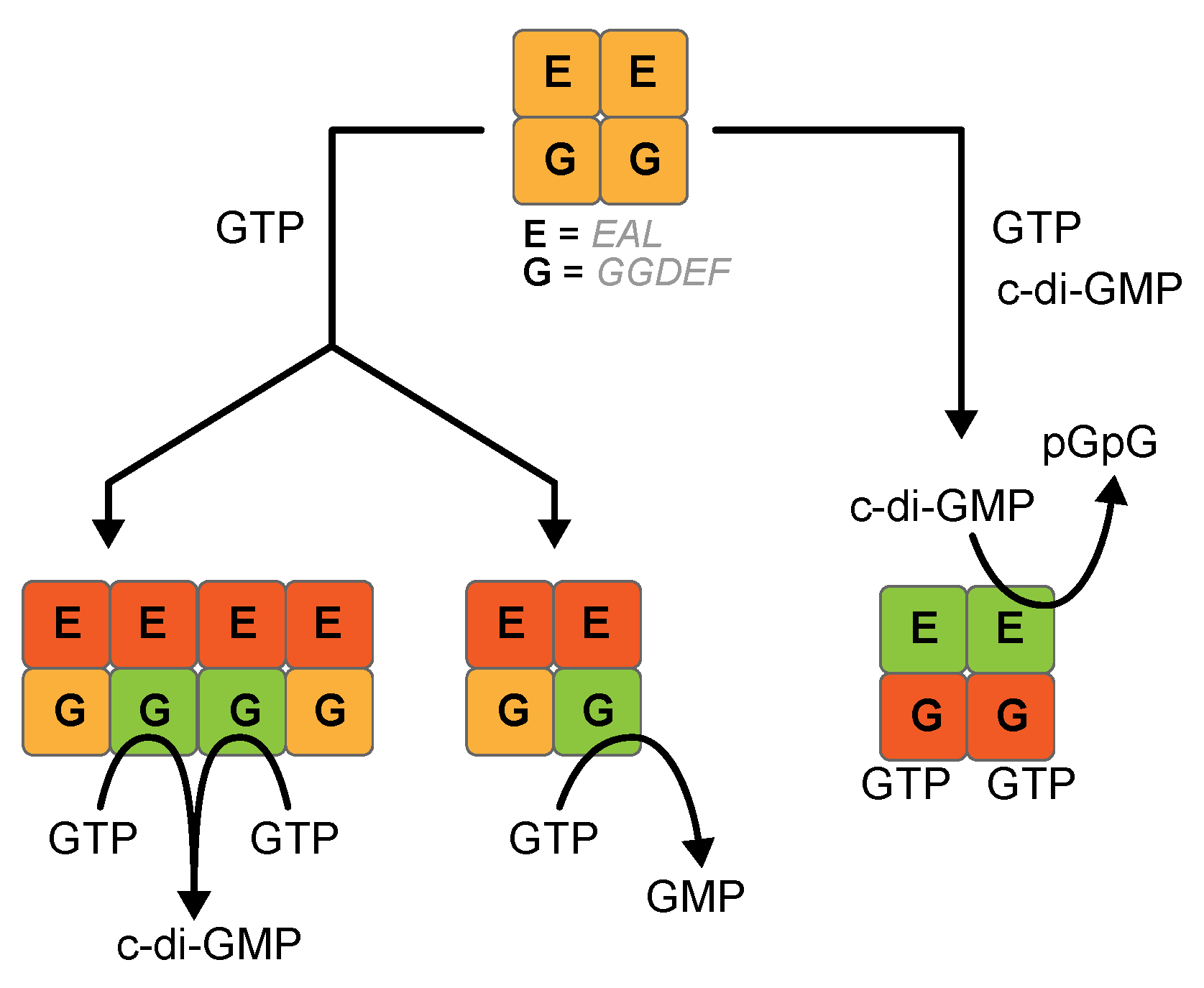
3.2. (α-β)-GTPase Activity on Other Hybrid Proteins
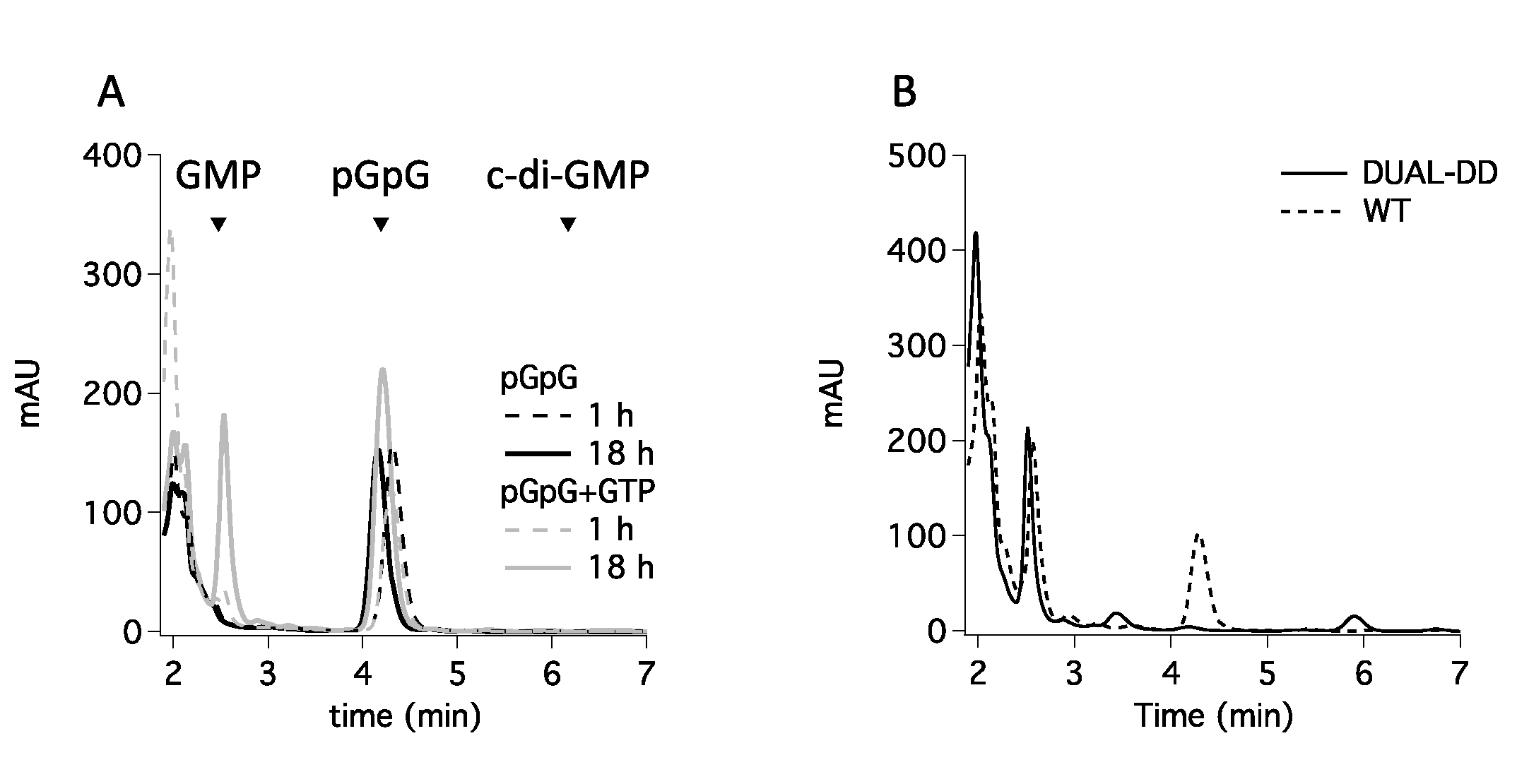
3.3. GMP Production Is Not Due to an Unexpected PDE-B Activity of the EAL Domain
3.4. GGDEF Incompetent Dimerization Promotes GMP Production
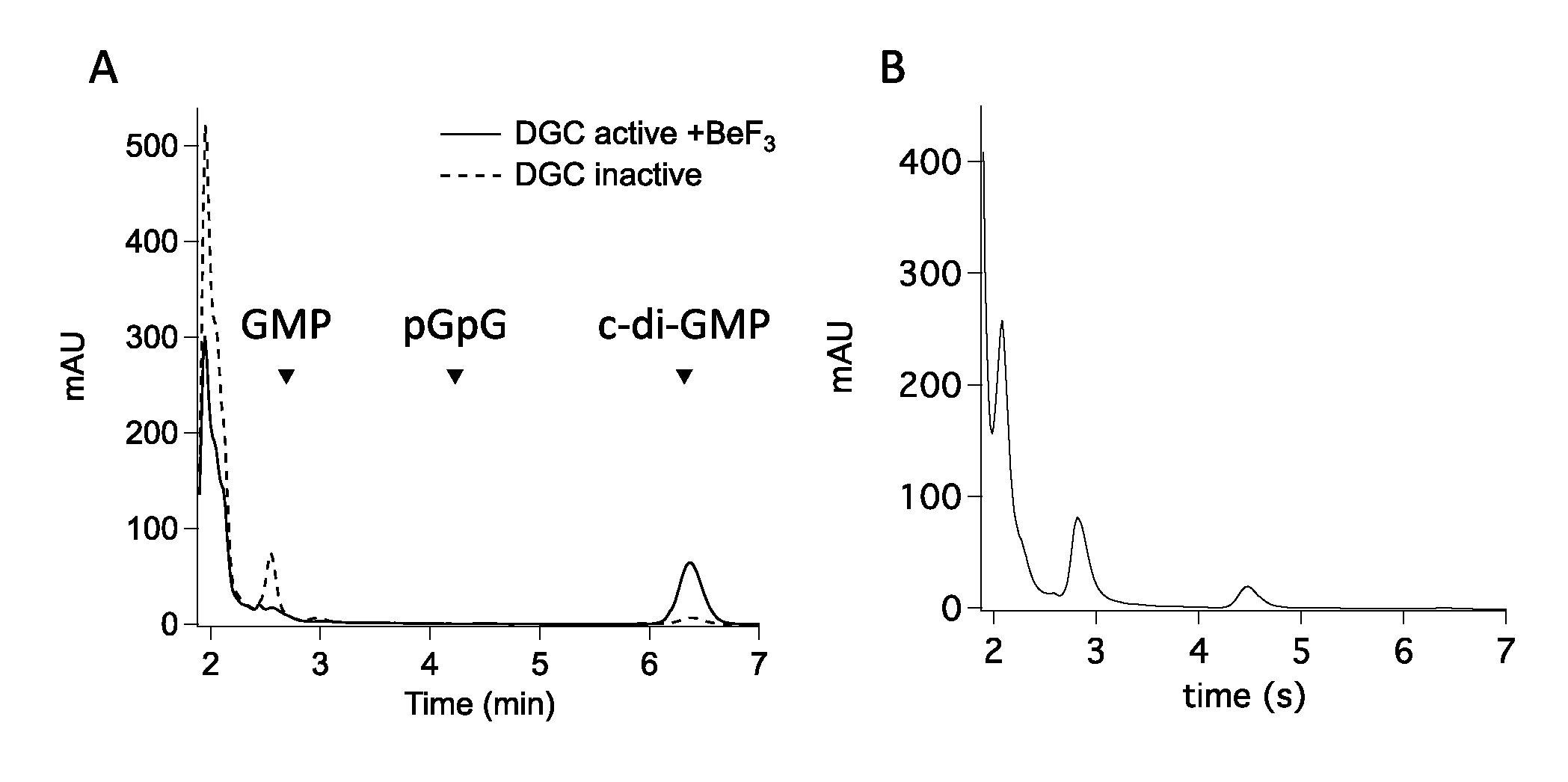
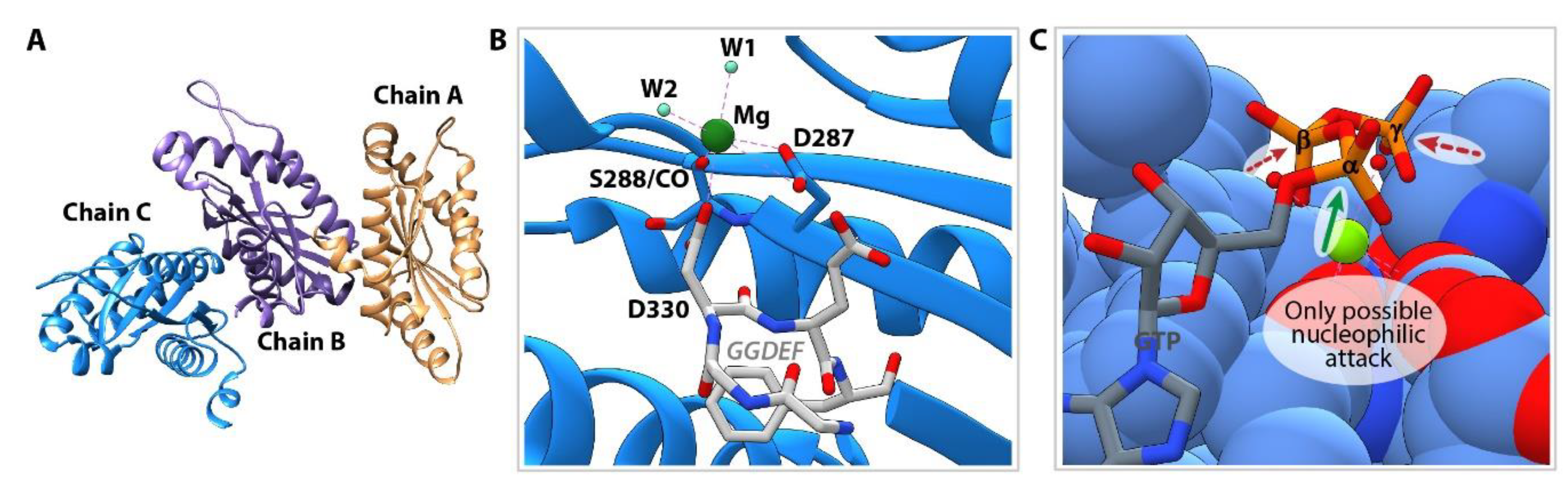
3.5. Structure of Inactive YfiNGGDEF
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Römling, U.; Liang, Z.-X.; Dow, J.M. Progress in Understanding the Molecular Basis Underlying Functional Diversification of Cyclic Dinucleotide Turnover Proteins. J. Bacteriol. 2016, 199, e00790-16. [Google Scholar] [CrossRef] [PubMed]
- Chou, S.-H.; Galperin, M.Y. Diversity of Cyclic Di-GMP-Binding Proteins and Mechanisms. J. Bacteriol. 2015, 198, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Schirmer, T.; Jenal, U. Structural and mechanistic determinants of c-di-GMP signalling. Nat. Rev. Genet. 2009, 7, 724–735. [Google Scholar] [CrossRef] [PubMed]
- Römling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The First 25 Years of a Universal Bacterial Second Messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [CrossRef]
- Mantoni, F.; Paiardini, A.; Brunotti, P.; D’Angelo, C.; Cervoni, L.; Paone, A.; Cappellacci, L.; Petrelli, R.; Ricciutelli, M.; Leoni, L.; et al. Insights into the GTP-dependent allosteric control of c-di-GMP hydrolysis from the crystal structure of PA0575 protein from Pseudomonas aeruginosa. FEBS J. 2018, 285, 3815–3834. [Google Scholar] [CrossRef] [PubMed]
- Bharati, B.K.; Mukherjee, R.; Chatterji, D. Substrate-induced domain movement in a bifunctional protein, DcpA, regulates cyclic di-GMP turnover: Functional implications of a highly conserved motif. J. Biol. Chem. 2018, 293, 14065–14079. [Google Scholar] [CrossRef]
- Phippen, C.W.; Mikolajek, H.; Schlaefli, H.G.; Keevil, C.W.; Webb, J.S.; Tews, I. Formation and dimerization of the phosphodiesterase active site of the Pseudomonas aeruginosa MorA, a bi-functional c-di-GMP regulator. FEBS Lett. 2014, 588, 4631–4636. [Google Scholar] [CrossRef]
- Liu, C.; Liew, C.W.; Wong, Y.H.; Tan, S.T.; Poh, W.H.; Manimekalai, M.S.S.; Rajan, S.; Xin, L.; Liang, Z.-X.; Grüber, G.; et al. Insights into Biofilm Dispersal Regulation from the Crystal Structure of the PAS-GGDEF-EAL Region of RbdA from Pseudomonas aeruginosa. J. Bacteriol. 2017, 200. [Google Scholar] [CrossRef]
- Christen, M.; Christen, B.; Folcher, M.; Schauerte, A.; Jenal, U. Identification and Characterization of a Cyclic di-GMP-specific Phosphodiesterase and Its Allosteric Control by GTP. J. Biol. Chem. 2005, 280, 30829–30837. [Google Scholar] [CrossRef]
- An, S.; Wu, J.; Zhang, L.-H. Modulation of Pseudomonas Aeruginosa Biofilm Dispersal by a Cyclic-Di-GMP Phosphodiesterase with a Putative Hypoxia-Sensing Domain. Appl. Environ. Microbiol. 2010, 76, 8160–8173. [Google Scholar] [CrossRef]
- Cho, K.H.; Tryon, R.G.; Kim, J.-H. Screening for Diguanylate Cyclase (DGC) Inhibitors Mitigating Bacterial Biofilm Formation. Front. Chem. 2020, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Sambanthamoorthy, K.; Sloup, R.E.; Parashar, V.; Smith, J.M.; Kim, E.E.; Semmelhack, M.F.; Neiditch, M.B.; Waters, C.M. Identification of Small Molecules That Antagonize Diguanylate Cyclase Enzymes to Inhibit Biofilm Formation. Antimicrob. Agents Chemother. 2012, 56, 5202–5211. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Watt, S.; Wang, J.; Nakayama, S.; Sayre, D.A.; Lam, Y.-F.; Lee, V.T.; Sintim, H.O. Potent suppression of c-di-GMP synthesis via I-site allosteric inhibition of diguanylate cyclases with 2′-F-c-di-GMP. Bioorg. Med. Chem. 2013, 21, 4396–4404. [Google Scholar] [CrossRef] [PubMed]
- Fernicola, S.; Paiardini, A.; Giardina, G.; Rampioni, G.; Leoni, L.; Cutruzzolà, F.; Rinaldo, S. In SilicoDiscovery andIn VitroValidation of Catechol-Containing Sulfonohydrazide Compounds as Potent Inhibitors of the Diguanylate Cyclase PleD. J. Bacteriol. 2015, 198, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, C.A.; Gaffney, B.L.; Jones, R.A.; Strobel, S.A. Identification of c-di-GMP Derivatives Resistant to an EAL Domain Phosphodiesterase. Biochemistry 2013, 52, 365–377. [Google Scholar] [CrossRef][Green Version]
- Fernicola, S.; Torquati, I.; Paiardini, A.; Giardina, G.; Rampioni, G.; Messina, M.; Leoni, L.; Del Bello, F.; Petrelli, R.; Rinaldo, S.; et al. Synthesis of Triazole-Linked Analogues of c-di-GMP and Their Interactions with Diguanylate Cyclase. J. Med. Chem. 2015, 58, 8269–8284. [Google Scholar] [CrossRef]
- Ching, S.M.; Tan, W.J.; Chua, K.L.; Lam, Y. Synthesis of cyclic di-nucleotidic acids as potential inhibitors targeting diguanylate cyclase. Bioorg. Med. Chem. 2010, 18, 6657–6665. [Google Scholar] [CrossRef]
- Giardina, G.; Paiardini, A.; Fernicola, S.; Franceschini, S.; Rinaldo, S.; Stelitano, V.; Cutruzzolà, F. Investigating the Allosteric Regulation of YfiN from Pseudomonas aeruginosa: Clues from the Structure of the Catalytic Domain. PLoS ONE 2013, 8, e81324. [Google Scholar] [CrossRef]
- Stelitano, V.; Brandt, A.; Fernicola, S.; Franceschini, S.; Giardina, G.; Pica, A.; Rinaldo, S.; Sica, F.; Cutruzzolà, F. Probing the activity of diguanylate cyclases and c-di-GMP phosphodiesterases in real-time by CD spectroscopy. Nucleic Acids Res. 2013, 41, e79. [Google Scholar] [CrossRef]
- Kabsch, W. Software XDS for Image Rotation, Recognition and Crystal Symmetry Assignment. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, D66, 125–132. [Google Scholar] [CrossRef]
- Evans, P.R.; Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Number 4 Collaborative Computational Project. The CCP4 suite: Programs for protein crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 1994, 50, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Vagin, A.A.; Teplyakov, A. Molecular replacement withMOLREP. Acta Crystallogr. Sect. D Biol. Crystallogr. 2009, 66, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Paiardini, A.; Mantoni, F.; Giardina, G.; Paone, A.; Janson, G.; Leoni, L.; Rampioni, G.; Cutruzzolà, F.; Rinaldo, S. A novel bacteriall-arginine sensor controlling c-di-GMP levels inPseudomonas aeruginosa. Proteins Struct. Funct. Bioinform. 2018, 86, 1088–1096. [Google Scholar] [CrossRef]
- Stelitano, V.; Giardina, G.; Paiardini, A.; Castiglione, N.; Cutruzzolà, F.; Rinaldo, S. C-di-GMP Hydrolysis by Pseudomonas aeruginosa HD-GYP Phosphodiesterases: Analysis of the Reaction Mechanism and Novel Roles for pGpG. PLoS ONE 2013, 8, e74920. [Google Scholar] [CrossRef]
- Paul, R.; Abel, S.; Wassmann, P.; Beck, A.; Heerklotz, H.; Jenal, U. Activation of the Diguanylate Cyclase PleD by Phosphorylation-mediated Dimerization. J. Biol. Chem. 2007, 282, 29170–29177. [Google Scholar] [CrossRef]
- Schirmer, T. C-di-GMP Synthesis: Structural Aspects of Evolution, Catalysis and Regulation. J. Mol. Biol. 2016, 428, 3683–3701. [Google Scholar] [CrossRef]
- Toulouse, G. Theory of Frustration Effects in Spin-Glasses: I. Comm. Phys. 1977, 2, 115. [Google Scholar]
- Gianni, S.; Camilloni, C.; Giri, R.; Toto, A.; Bonetti, D.; Morrone, A.; Sormanni, P.; Brunori, M.; Vendruscolo, M. Understanding the Frustration Arising from the Competition between Function, Misfolding, and Aggregation in a Globular Protein. Proc. Natl. Acad. Sci. USA 2014, 111, 14141–14146. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.C.; Teixeira, R.D.; Andrade, M.O.; Pinheiro, G.M.; Ramos, C.H.; Farah, C.S. Cooperative Substrate Binding by a Diguanylate Cyclase. J. Mol. Biol. 2015, 427, 415–432. [Google Scholar] [CrossRef] [PubMed]
- Anand, B.; Verma, S.K.; Prakash, B. Structural stabilization of GTP-binding domains in circularly permuted GTPases: Implications for RNA binding. Nucleic Acids Res. 2006, 34, 2196–2205. [Google Scholar] [CrossRef]
- Rocchio, S.; Santorelli, D.; Rinaldo, S.; Franceschini, M.; Malatesta, F.; Imperi, F.; Federici, L.; Travaglini-Allocatelli, C.; Di Matteo, A. Structural and functional investigation of the Small Ribosomal Subunit Biogenesis GTP ase A (RsgA) from Pseudomonas aeruginosa. FEBS J. 2019, 286, 4245–4260. [Google Scholar] [CrossRef] [PubMed]
- Anand, B.; Surana, P.; Bhogaraju, S.; Pahari, S.; Prakash, B. Circularly permuted GTPase YqeH binds 30S ribosomal subunit: Implications for its role in ribosome assembly. Biochem. Biophys. Res. Commun. 2009, 386, 602–606. [Google Scholar] [CrossRef][Green Version]
- Simm, R.; Morr, M.; Kader, A.; Nimtz, M.; Römling, U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol. Microbiol. 2004, 53, 1123–1134. [Google Scholar] [CrossRef]
- Weinhouse, H.; Sapir, S.; Amikam, D.; Shilo, Y.; Volman, G.; Ohana, P.; Benziman, M. c-di-GMP-binding protein, a new factor regulating cellulose synthesis inAcetobacter xylinum. FEBS Lett. 1997, 416, 207–211. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection | PA1120-E232start a |
|---|---|
| Beamline | Bessy ID14.1 |
| Space group | P212121 |
| Cell dimension; a b c (Å), α β γ (°) | 79.64 85.10 126.97 90.00 90.00 90.00 |
| Resolution range (Å) | 49.64–2.80 (2.95–2.80) |
| Half-set correlation CC(½) (%) b | 99.6 (68.2) |
| <I/σI> | 10.5 (1.6) |
| Completeness (%) | 100 (100) |
| N. unique reflection (total) | 21,951 (3175) |
| Multiplicity | 13.2 (13.5) |
| Wilson B-factor (Å2) | 56.1 |
| Refinement | |
| Resolution range (Å) | 49.0–2.80 |
| N° of unique reflections | 19,705 |
| Rfree test set | 2195 reflections (10.2%) |
| Rwork/Rfree (%) | 21.96/22.36 |
| Fo,Fc correlation | 0.94 |
| Total N. of atoms | 3809 |
| Average B-factor all atoms (Å2) | 64.3 |
| Bonds RMSD | |
| Length (Å) | 0.007 |
| Angle (°) | 1.520 |
| Ramachandran plot (n. residues, %) | |
| Favoured | 485 (94.4) |
| Allowed | 29 (5.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantoni, F.; Scribani Rossi, C.; Paiardini, A.; Di Matteo, A.; Cappellacci, L.; Petrelli, R.; Ricciutelli, M.; Paone, A.; Cutruzzolà, F.; Giardina, G.; et al. Studying GGDEF Domain in the Act: Minimize Conformational Frustration to Prevent Artefacts. Life 2021, 11, 31. https://doi.org/10.3390/life11010031
Mantoni F, Scribani Rossi C, Paiardini A, Di Matteo A, Cappellacci L, Petrelli R, Ricciutelli M, Paone A, Cutruzzolà F, Giardina G, et al. Studying GGDEF Domain in the Act: Minimize Conformational Frustration to Prevent Artefacts. Life. 2021; 11(1):31. https://doi.org/10.3390/life11010031
Chicago/Turabian StyleMantoni, Federico, Chiara Scribani Rossi, Alessandro Paiardini, Adele Di Matteo, Loredana Cappellacci, Riccardo Petrelli, Massimo Ricciutelli, Alessio Paone, Francesca Cutruzzolà, Giorgio Giardina, and et al. 2021. "Studying GGDEF Domain in the Act: Minimize Conformational Frustration to Prevent Artefacts" Life 11, no. 1: 31. https://doi.org/10.3390/life11010031
APA StyleMantoni, F., Scribani Rossi, C., Paiardini, A., Di Matteo, A., Cappellacci, L., Petrelli, R., Ricciutelli, M., Paone, A., Cutruzzolà, F., Giardina, G., & Rinaldo, S. (2021). Studying GGDEF Domain in the Act: Minimize Conformational Frustration to Prevent Artefacts. Life, 11(1), 31. https://doi.org/10.3390/life11010031