3.2.1. Cranial and Mandibular Elements
While cranial and mandibular material is rare among hesperornithiform fossils,
Parahesperornis alexi KUVP 2287 preserves a nearly complete (albeit crushed) skull. Comparative material is largely limited to two fairly complete skulls (KUVP 71012, YPM 1206) of
Hesperornis, which have been extensively described in the literature [
7,
8,
56,
57,
58,
69,
70,
71,
72,
73], and a number of specimens preserving fragmentary cranial elements. It should be noted that due to previous, but incorrect, assignment of KUVP 2287 to
Hesperornis gracilis, some previous works describe the skull of KUVP 2287 under that name [
58,
59]. In addition to some fragmentary material with other
Hesperornis specimens (FMNH 219, NMNH 4978, NMNH 6622, NMNH 13580, UNSM 10148, YPM 903, YPM 1207, YPM PU 18589), the only other skull material known from the Hesperornithiformes is the complete quadrate of
Potamornis skutchi [
49] (UCMP 73103), fragmentary portions of several unassociated skull elements assigned to
Pasquiaornis [
65], the caudal portion of the mandibular ramus of
Baptornis (FMNH 395), and three fragmentary braincases assigned to
Enaliornis [
72]. The braincase of
Enaliornis (SMC B54404) is exceptional in the lack of crushing of the specimen (see previous work [
72] for a complete description of the braincase of
Enaliornis). Very small fragments of the quadrate and frontal of
Baptornis (AMNH 5101) were previously reported [
19], however no such elements are included with the specimen today. Elzanowski [
49] noted that the accession record of AMNH 5101 never listed cranial elements, and so concluded the specimen was incorrectly attributed to AMNH 5101. Likewise, a fragment of the bill was reported with
Baptornis specimen KUVP 16112 [
19], however the specimen did not include material identifiable as such when consulted for this study. Outside of the hesperornithiforms, cranial material in Mesozoic ornithuromorphs is largely crushed, making detailed comparisons of morphology difficult. The skull of
Ichthyornis is known from a number of three-dimensional specimens [
61], and so is one of the only other non-neornithine ornithuromorphs for which comparisons of many aspects of the cranium can be made.
The skull of KUVP 2287 was originally preserved partially disarticulated. The majority of the cranium and upper jaw were articulated (
Figure 4), while the lower jaws, quadrates, pterygoids, palatines, and lacrimals were preserved as isolated elements. At some point, the cranium and upper jaws were disarticulated into two pieces, one including the braincase and frontals (
Figure 5 and
Figure 6) and the second piece consisting of the premaxillae, nasals, and a number of palatal elements (
Figure 7 and
Figure 8). This is important to note, as during this process some material from the specimen was lost (see
Figure 4). Possible identifications for these missing pieces are discussed below.
Braincase. The braincase of KUVP 2287 is highly crushed dorsoventrally and articulated to the frontals (
Figure 5 and
Figure 6). For comparisons, braincases of
Hesperornis (KUVP 71012) and
Enaliornis (SMC B54404) were examined (
Figure 9). In ventral view (
Figure 5B), the occipital region of KUVP 2287 is fairly clear. The occipital condyle is broken into halves and is slightly hook-shaped, with the rounded surface curving over the somewhat indented dorsal base of the condyle. Despite the breakage, the occipital condyle of
Parahesperornis appears to be much smaller proportionally than that of
Hesperornis (
Figure 9). The occipital condyle of
Enaliornis is not preserved.
Ventrally, rostral to the occipital condyle is a deep, round recess in the
lamina parasphenoidalis (rlp in
Figure 5). This recess appears to be narrower and more deeply excavated in KUVP 2287 than in
Hesperornis, however this may be enhanced by crushing. In
Enaliornis this recess is prominent and mediolaterally elongate, taking up a larger portion of the ventral skull than in either
Parahesperornis or
Hesperornis, perhaps indicating the degree of crushing in both KUVP 2287 and KUVP 71012.
In
Parahesperornis the recess of the
lamina parasphenoidalis is defined by the basioccipital and basisphenoid (BO and BS, respectively, in
Figure 5); the latter bones wrap around the recess and nearly meet at the base of the parasphenoid rostrum (PS in
Figure 5). The basioccipitals of
Parahesperornis are roughly kidney-shaped, with what appear to be prominent sutures on the caudal ends, the right of which is somewhat open (s in
Figure 5 and
Figure 6). In
Hesperornis the basioccipitals are more elongate with less curvature and lack any visible sutures. These elements are not preserved in
Enaliornis.The basisphenoids of
Parahesperornis are both somewhat crushed but appear to overlap the basioccipitals, with a flattened face that angles toward the parasphenoid rostrum, as in
Hesperornis. There appears to be a suture present between the basioccipital and basisphenoid of
Parahesperornis. A similar suture is present and more obvious in
Hesperornis. In
Enaliornis, the area rostral to the recess in the
lamina parasphenoidalis is very different, preserving a distinct pyramidal projection with two large fossae on the lateral and medial margins, tentatively identified as the
canalis orbitalis by Elzanowski and Galton [
72].
The parasphenoid rostrum of
Parahesperornis extends rostrally as a straight, paired bone with a smoothly rounded ventral surface (PS in
Figure 5). Lucas [
13] noted that the parasphenoid rostrum extended to the mesethemoid in KUVP 2287, however today it is broken considerably caudal to the mesethemoid. In
Hesperornis the ventral surface of parasphenoid rostrum is projected and forms a crest. The parsphenoid rostrum is not preserved in
Enaliornis.The basioccipitals are propped up on struts of bone, into which two foramina are visible piercing the base of the strut (
Figure 5 and
Figure 6). While the exact identity of these foramina is uncertain, it is likely that the smaller, more caudal foramen corresponds to the foramen for the exit of the hypoglossal nerve (XII) (fh in
Figure 5 and
Figure 6) while the larger, laterally facing one corresponds to the foramen for the vagus nerve (X) (fv in
Figure 6). This is unlike
Hesperornis, where three small foramina are seemingly associated with the hypoglossal nerve, while
Parahesperornis has only one. In
Enaliornis these foramina are present at the caudomedial and -lateral borders of the recess in the
lamina parsphenoidalis (
Figure 9).
Lateral to these foramina the exoccipital extends sideways, forming a prominent rounded paraoccipital process with a flattened ventral surface (pop in
Figure 5; sometimes referred to as the exoccipital process [
67]). This process is separated from the more rostrally-located zygomatic process (zp in
Figure 5) by a deep indentation in the lateral margin. This indent is much less dramatic in
Hesperornis, where the exoccipital process and prootic appear as a more continuous rectangular shape in ventral view. The indentation is lacking entirely in
Enaliornis, where the paraoccipital processes are much reduced and appear as very slight protuberances in dorsal view (
Figure 9).
The zygomatic process of
Parahesperornis extends laterally and slightly rostrally from the edge of the dorsal cranium and has a flattened face that has been crushed under ventrally on both sides. This face is better preserved in
Hesperornis KUVP 71012, where it is oval shaped with a slightly domed surface and appears to form a slight hook, however discrepancies between the left and right sides make it unclear how much of that shape is due to crushing (
Figure 9). The zygomatic processes of both
Parahesperornis and
Hesperornis are much reduced as compared to those of
Enaliornis, where they form broad triangular processes directed more rostrally than in hesperornithids (
Figure 9).
Dorsally, the area of the squamosals and exoccipitals medial to the zygomatic and paraoccipital process is developed as a broad, shallowly depressed face that is roughly rectangular. A few small foramina are preserved near the lateral indentation in the margin between the zygomatic and paraoccipital processes. In contrast to Parahesperornis, a single foramen is observed in Hesperornis; however, the region is obscured by crushing in KUVP 71012, the only skull available for study where this feature is observed. In Hesperornis this area also forms a broad depression; however, the shape approaches a square in Hesperornis as opposed to the rectangle in Parahesperornis. In Enaliornis this area is smaller and flattened as compared to the depressed surface in hesperornithids.
Ventrally, deep facets are developed medial to the zygomatic processes for the articulation with the otic processes of the quadrate (Qaf in
Figure 5). This facet is better preserved on the left side of the skull and is like that seen in
Hesperornis. In
Enaliornis the articulation for the quadrate is less defined, and forms a shallower depression divided into two similarly sized facets for articulation with the squamosal and otic heads of the quadrate, separated by the dorsal pneumatic foramen (
Figure 9). This foramen is not observed in the facet in KUVP 2287 or KVUP 71012.
In ventral view, the braincase of
Parahesperornis makes a sweeping curve at the margin of the temporal fossa that ends in a laterally projecting terminus where the braincase articulates with the frontals. This terminus does not appear to form a well-developed postorbital process (as in
Ichthyornis [
61]), however it is possible breakage has obscured this. The margin of the temporal fossa echoes that of the orbital margin of the frontals but is slightly shorter and indented to a lesser degree. It is unclear where the boundary between the parietals and the laterosphenoid occurs. As such our identifications of the boundaries of these bones (PR and LS in
Figure 5) should be considered tentative.
Hesperornis has a much more exaggerated lateral projection of the postorbital process than
Parahesperornis, while that of
Enaliornis is reduced (
Figure 9). The ventral walls of the braincase leading up to this margin are smooth but extremely crushed in
Parahesperornis. In
Hesperornis, they slope slightly ventrally to meet below the parasphenoid rostrum. The breakage pattern in KUVP 2287 might suggest that the walls of the braincase were not fused where they met at the midline, however this cannot be confirmed.
While crushing might obscure some details of the exact margins of the temporal fossa, it appears to be mostly open and only partially enclosed by the zygomatic process caudally and the postorbital process rostrally. The temporal fossa of
Hesperornis is similar, while that of
Enaliornis is much shorter caudorostrally. The structure of the temporal fossa is intermediate between most modern birds, where it is much reduced and only slightly enclosed by bone, and
Ichthyornis, where the temporal fossa is almost completely enclosed by bone, more similar to the primitive dinosaurian condition [
61]. The temporal fossa in some Neornithes, such as the extinct (Mio-Pliocene) flightless alcids Mancallinae, appear to be most similar to that of the hesperornithids [
74].
In dorsal view the caudal portion of the surface of the braincase of
Parahesperornis KUVP 2287 has split along the sagittal crest (sc in
Figure 5), obscuring the foramen magnum. While it is not possible to describe the sagittal crest in
Parahesperornis, in
Hesperornis it is a large, sharp crest tightly sutured along its length (
Figure 9). As the point of attachment for the deep fascia of the neck muscles
67, the robust sagittal crest indicates the significance of these muscles in stabilizing the neck while diving. In
Enaliornis the sagittal crest is only faintly developed caudally, between the squamosals, and appears to become less sutured rostrally, with the rostral extent partially open.
Rostral to the squamosal area on the dorsal side, the braincase of KUVP 2287 is heavily crushed, however a small piece in the center of the crushed area preserves a section of the parietals with the sagittal crest and a piece that appears to contain the frontoparietal suture (F-P in
Figure 5). It should be noted that this piece may have been positioned during preparation, as undated photographs show the specimen without this piece (
Figure 10). The frontoparietal suture is better preserved in
Hesperornis, where it runs in a broad transversal arc and forms a cross where it intersects the sagittal crest (
Figure 11). In
Hesperornis the frontoparietal contact appears to be fully sutured. The small portions of the preserved frontoparietal suture in
Parahesperornis KUVP 2287 agree with this morphology.
This observation contradicts Martin’s [
40] interpretation of the frontals and parietals as meeting in a moveable joint, giving
Parahesperornis a mesokinetic cranium, while the juncture in
Hesperornis was fused and immobile. Later work by Buhler et al. [
57] described the frontoparietal juncture of
Parahesperornis as consisting of a posterior ridge on the frontals that fit into an anterior groove on the parietals; however, due to the degree of crushing in KUVP 2287 this cannot be confirmed. Rather, the small portion of the frontoparietal suture preserved where it intersects the sagittal crest forms a crest and appears to be sutured.
In
Enaliornis, the three known braincases are broken along what has been interpreted to be the frontoparietal suture [
21]. If correct, then the frontoparietal suture was nearly linear in
Enaliornis, not arced as in hesperornithids. This consistent breakage pattern was also used to infer an open frontoparietal suture in
Enaliornis [
21]. One specimen assigned to
Pasquiaornis (RSM P2995.4) preserves a portion of the frontoparietal suture, which does not appear to be open [
65]. The suture forms a low, thin crest, that is fairly straight and angles caudally from the juncture with the sagittal crest. In all, despite apparent differences in the trajectories of the frontoparietal suture, with the possible exception of
Enaliornis, the frontals and parietals of hesperornithiforms appeared to have been sutured to each other.
The braincases of
Parahesperornis and
Hesperornis show much less pneumaticity than
Enaliornis. In
Enaliornis the braincase is highly pneumatic, with multiple foramina on the caudal end in the occipital region [
72]. While the crushing of the braincases of
Parahesperornis KUVP 2287 and
Hesperornis KUVP 71012 may have obscured some foramina in the rostral portions, the caudal braincase is fairly well preserved, and it is doubtful that either skull would have approached the level of pneumaticity seen in
Enaliornis. In KUVP 2287, internal views along the broken sagittal crest show some pneumatization of the interior of the bone (ip in
Figure 5). However, there is no evidence of internal pneumaticity in the broken edges of
Hesperornis KUVP 71012.
Frontals. The frontals of
Parahesperornis KUVP 2287 are preserved somewhat articulated with and flattened back upon the braincase (
Figure 5). For comparison, the frontals of
Hesperornis (KUVP 71012, YPM 1206) and
Pasquiaornis (RSM P2995.4 [
65]) were examined (
Figure 11). In dorsal view, the frontals of KUVP 2287 meet at the interfrontal suture, which forms a slight groove. To either side of this groove the frontals are flattened, with an arced, central crest that parallels the margin of the orbit, running along the length of the dorsal surface. Such morphology is very different from that of the frontals of
Hesperornis and
Pasquiaornis, where the interfrontal suture forms a longitudinal crest, continuous with the sagittal crest.
The central region of the sutured frontals is shortened in Parahesperornis as compared to Hesperornis. This gives the mediocaudal margin of the orbit, as formed by the lateral edge of the frontals, a more circular shape in Parahesperornis, whereas in Hesperornis the orbit is elongate and much more shallowly curved. It is difficult to compare the shape of the incomplete caudal orbit in Pasquiaornis; however, it appears to be intermediate between the morphologies of Hesperornis and Parahesperornis. In relative terms, Parahesperornis has a much broader skull than Hesperornis at the level of the orbits. The frontals of Hesperornis and Parahesperornis have similar widths, despite Parahesperornis being a much smaller animal overall. Pasquiaornis appears proportionally narrower as well, more like Hesperornis.
The frontals of
Parahesperornis appear to be tightly sutured along the majority of their length, however at the rostral-most end they become slightly separated from one another before the left and right frontals diverge and define a dramatic U-shaped rostral terminus (
Figure 5A). This U-shaped rostral end of the frontals is also seen in
Hesperornis and
Pasquiaornis, however the U-shape is narrower in the latter (
Figure 11). This contrasts with
Ichthyornis, where the frontals angle away from each other slightly, forming a Y-shape [
61]. In lateral view, the frontals of
Parahesperornis are fairly flat, as in
Pasquiaornis, however in
Hesperornis they are strongly domed caudally (
Figure 11C).
In
Parahesperornis, the dorsal margin of the frontals forming the upper orbit slope toward the orbit fairly steeply along their entire length, forming an angle dorsally where they flatten out medially before meeting at the interfrontal suture.
Pasquiaornis resembles
Parahesperornis in the angle of the flattened surface, however in
Hesperornis the angle is entirely lacking, and the sides of the frontals slope continuously from the juncture of the interfrontal and sagittal crests with the frontoparietal suture. Lucas [
13] described
Parahesperornis KUVP 2287 as having depressions for supraorbital glands (
fossa glandula nasales). In modern birds the lateral edges of the frontal dorsal to the orbit are often broad with a well-developed depression for the supraorbital gland. The morphologies seen in hesperornithiforms cast doubt about the interpretation of supraorbital glands, which if present, would have been rather small.
In ventral view, the orbital margin of the frontals forms a sharp lip or crest in
Parahesperornis (c in
Figure 5), as in
Pasquiaornis, while in
Hesperornis the frontals wrap over and nearly form a tunnel along the ventral orbital margin (
Figure 11). At its caudal-most end, this lip remains a consistent height in
Parahesperornis, while in
Hesperornis it forms an exaggerated ventrally directed projection, best seen in lateral view (
Figure 11C). At the caudal-most edge of the orbital margin the frontals narrow to a triangular projection with a facet for the postorbital process of the parietal (fpo in
Figure 5). This facet is much more strongly developed in
Hesperornis, where it is separated from the medial region of the frontals by a ridge that is only faintly visible in
Parahesperornis.
Nasals. The nasals of
Parahesperornis KUVP 2287 are poorly preserved. Both the left and right are preserved crushed against the premaxillae, with the right more easily observed (
Figure 7). For comparison, the nasals of
Hesperornis (KUVP 71012, YPM 1206), the only other hesperornithiform from which nasals have been reported, were examined (
Figure 12). The premaxillary and maxillary processes of the nasal of
Parahesperornis (Np and Nm, respectively, in
Figure 7) extend cranially in a similar orientation and run parallel to each other along their length, as in
Hesperornis. A small foramen is present on the main body of the nasal at the juncture of these processes in both
Parahesperornis and
Hesperornis (
Figure 12).
The dorsal surface of the premaxillary process of the nasal of
Parahesperornis appears to have a broad, faint groove caudally, near the juncture with the maxillary processes, that flattens rostrally (g1 in
Figure 7). In
Hesperornis, the dorsal surface of the premaxillary process is flattened along its length with a similar groove along the dorsal-most edge that does not extend as far caudally as in
Parahesperornis. The articulation between the maxillary processes of the nasal and premaxilla is clearly visible; the maxillary process of the premaxilla thins dramatically to a very thin sheet of bone that overlays the lateral face of the maxillary process of the nasal for a few centimeters, forming what Gingerich [
58] termed a subnarial bar. Details of how this subnarial bar articulated with the maxilla are not well preserved in KUVP 2287. However, the ventral surface of the maxillary process of the right premaxilla appears to be flattened with a very faint caudal groove (g2 in
Figure 7). Previously, Gingerich [
58] identified the subnarial bar as sliding into a prominent groove developed on the dorsal margin of the maxilla from observations of
Hesperornis YPM 1206. This is also seen in
Hesperornis KUVP 71012, but a maxilla is not preserved (or is not complete enough to be identified) with KUVP 2287.
Lacrimals. Portions of both lacrimals are preserved with
Parahesperornis KUVP 2287. The right lacrimal is nearly complete but lacks the cranial process (
Figure 13), while the left consists of only a small fragment of the central region. For comparison, the lacrimals of
Hesperornis KUVP 71012 were examined (
Figure 14). The lacrimal of
Parahesperornis is roughly T-shaped, with the caudal process extending dorsocaudally and the jugal process extending ventrally and slightly caudally.
The lacrimal of Parahesperornis differs from that of Hesperornis in several respects. Overall, the lacrimal of Parahesperornis is much more elongate than that of Hesperornis, with a relatively narrow body and proportionally longer jugal process. The end of the jugal process in Parahesperornis terminates in an angled, flattened face with an upturned lip around the margin for articulation with the jugal. However, the jugal process of Hesperornis terminates in a pointed foot. At the base of the caudal margin of the jugal process, Parahesperornis has a muscle scar that is absent in Hesperornis. Additionally, in caudal view, the face of the jugal process of Parahesperornis is highly irregular, with a deep ovoid depression along its surface. In Hesperornis the corresponding area is narrow and rounded, with only a slight depression centered at the distal end.
In lateral view, the side of the lacrimal of Parahesperornis has an ovoid depression that appears to shallow rostrally, such that it would not have extended beyond the broken edge of the cranial process. This region is more deeply excavated in Hesperornis, where it forms a groove that parallels the dorsal margin of the body of the lacrimal. Also in lateral view, the caudal process forms a dorsal curve in Parahesperornis but angles dorsally in a straight line in Hesperornis.
Witmer [
75] established that the lacrimals of both
Hesperornis and
Parahesperornis are pneumatic. In
Parahesperornis a large, funnel-like opening is present on the caudal margin of the depression in the lateral face of the lacrimal (best seen in craniomedial view,
Figure 13E). In
Hesperornis, the external evidence of this pneumaticity is limited to a small foramen located in the middle of the medial face of the lacrimal.
The overall shape of the lacrimal, and in particular the elongation of the jugal process, in
Parahesperornis is similar to that of other Mesozoic ornithuromorphs (e.g.,
Ichthyornis [
61] and
Dingavis [
76]), which share an overall T-shaped morphology with
Archeopteryx [
77], reflecting the ancestral state in birds. This highlights the more compact lacrimal of
Hesperornis, with a shortened jugal process, as an autapomorphy among Mesozoic birds.
Quadrate. Parahesperornis KUVP 2287 preserves the complete left quadrate (
Figure 15) and fragments of the otic process and dorsal articular region of the right quadrate. In addition to
Hesperornis (KUVP 71012), a well-preserved quadrate is also known for
Potamornis [
49] (UCMP 73013) (
Figure 16), and two poorly preserved quadrates have been reported for
Pasquiaornis (RSM P2988.25, RSM P2831.52) [
65]. As discussed above, a partial quadrate previously described [
19] as belonging to
Baptornis (AMNH 5101) was likely a mistaken attribution [
49]. Anatomical terminology for the discussion of the quadrate comes from Elzanowski [
49], who expanded upon the terminology of Baumel and Witmer [
67]. In general, the quadrates of hesperornithiforms are similar, with some minor differences in proportion and details of morphology, and much like those of modern birds and
Ichthyornis.
The otic process of the quadrate is poorly divided in Parahesperornis, as in other hesperornithids and modern paleognathes, but unlike the better defined otic and squamosal head in Ichthyornis and many modern birds. The otic head is slightly smaller than the squamosal head, while in Hesperornis and Potamornis the otic head is much smaller than the squamosal. While the quadrate of Enaliornis is not known, the division of the quadrate’s articular facet on the squamosal into visible medial and lateral cotylae indicate the otic and squamosal heads of the quadrate of Enaliornis may have been better divided.
In caudal view, the intercapitular incision wraps down onto the caudal face of the otic process and around the ventral margin of the otic head forming a caudomedial depression. This depression is much better developed in Parahesperornis than in Potamornis, while in Hesperornis the depression is absent. In medial view the otic process hooks backward over the neck and projects rostrally, as in Hesperornis and unlike Potamornis, where it is not hooked at all. In both Parahesperornis and Hesperornis, a small pocket is present beneath the lip of the head on the rostral surface of the neck. The neck is more robust in Parahesperornis and Potamornis, where it narrows very little before the otic process, while in Hesperornis the neck is more restricted.
The dorsal margin of the quadrate, between the otic process and the orbital process, is broad and u-shaped in
Parahesperornis, while in
Hesperornis it is slightly deeper (resulting from the longer neck of the otic process in
Hesperornis) and more v-shaped. The orbital process of
Parahesperornis KUVP 2287 lacks the anterior end, but the preserved portion is very thin mediolaterally and tapers anteriorly, as in
Hesperornis. Elzanowksi reported the orbital process of
Potamornis as only missing the anterior-most end. If this is true, then the orbital process was dramatically reduced in
Potamornis as compared to hesperornithids. The thin, flattened orbital process plays a role in the mobility of the quadrate, as that is where the m. protractor pterygoidei et quadrati, which pulls the quadrate dorsorostrally, inserts, and the m. pseudotemporalis profundus, which adducts the mandible [
78], originates [
57].
In medial view, the body of the quadrate of Parahesperornis just above the medial condyle is deeply excavated in a triangular pocket, as in Pasquiaornis. Potamornis has a shallow depression in this area, while the corresponding region in Hesperornis is only faintly depressed. In lateral view the body of the quadrate of Parahesperornis is fairly featureless and flat, while in Hesperornis and Potamornis the central region is slightly depressed. Also in Hesperornis and Potamornis, the neck is slightly twisted, such that the lateral crest crosses the lateral face just below the otic process, while in Parahesperornis the lateral crest is restricted to the lateral margin.
The caudal condyle on the mandibular process of Parahesperornis is small and thin, located to the medial side of the distocaudal surface. In Hesperornis this condyle is more robust, present as a thick, triangular projection that occupies most of the caudal surface of the pterygoid condyle. Potamornis has a small caudal condyle, similar in size to that of Parahesperornis, but developed as a hook projecting caudodorsally, unlike in other hesperornithiforms.
The pterygoid condyle is very well-developed and like that of modern birds. In Parahesperornis and Potamornis the pterygoid condyle is broad and triangular, while in Hesperornis it is oval with a slightly concave surface. The medial condyle of Parahesperornis, Potamornis, and Pasquiaornis is elongate and narrower at the caudal end than at the rostral end, with a slightly convex dorsal margin. That of Hesperornis is more uniform in width with much deeper curvature along the dorsal margin.
The lateral condyle of
Parahesperornis is narrower rostrocaudally than the pterygoid condyle, with an oval quadratojugal cotyla facing laterally and slightly rostrally. The margins of the cotyla are slightly pinched in toward the rostral end, creating a rostral projection that may correspond to the quadratojugal buttress identified in
Potamornis by Elzanowski [
49]. The quadratojugal cotyla is nearly circular in
Hesperornis, where it faces directly laterally. That of
Potamornis is oval in shape, however the long axis of the cotyla is oriented dorsoventrally, while in
Parahesperornis the long axis is oriented rostrodorsally to caudoventrally. The pneumaticity of the quadrate in hesperornithids was discussed by Witmer [
75] and Elzanowski [
49]. A single visible pneumatopore is located on the middle portion of the caudal face of the quadrate of
Parahesperornis. In
Hesperornis this pneumatopore is displaced toward the distal end. In
Potamornis no foramen is visible in this area.
Palatines. The palatines of
Parahesperornis and
Hesperornis have been somewhat contradictorily described by previous authors. What Marsh [
7] initially identified as a vomer of
Hesperornis (YPM 1206), Gingerich [
58], working with both KUVP 2287 and YPM 1206, later described as a palatine, and what Marsh [
7] referred to as a palatine Gingrich [
58] described as a vomer. Gingerich [
58,
69] described the palatine as articulating with the pterygoid and tapering anteriorly to where it is fused with the vomer. There is no evidence of this relationship preserved today with
Parahesperornis KUVP 2287. Working with KUVP 2287 before it was disarticulated, Gingerich [
69] described both palatines as “virtually complete” and identified the left palatine as preserved alongside the frontals and a portion of the right palatine alongside the premaxilla (see
Figure 4).
In his palatal reconstruction, Gingerich [
69] placed the palatines as articulating with the pterygoids (
Figure 4). While work by Witmer and Martin [
70] supported Gingerich’s interpretation, later work by Elzanowski [
71] rejected Gingerich’s [
69] reassignment of the bones of YPM 1206 as the palatine and vomer, reverting to Marsh’s [
7] original identifications. While Elzanowski did not specifically identify these elements in KUVP 2287, the bones identified as the palatines by Gingerich [
58,
69] in his reconstruction would be equivalent to the hemipterygoids of Elzanowski [
56]. The present study supports Elzanowski’s interpretation of the hemipterygoids.
Today, a portion of the left palatine is preserved pressed against the ventral side of the frontals of KUVP 2287 (PT in
Figure 5). The identification of the palatine of
Parahesperornis KUVP 2287 as supported by this study is based on the distinctive hook present at the cranial end of this bone, which is difficult to interpret as belonging to any other element. The remainder of this palatine is difficult to identify. Historic photos demonstrate that the small rod preserved alongside this hook is not associated with the palatine and is instead most likely the hemipterygoid (see discussion above). It seems likely that some portion of the palatines is preserved in the jumble of thin bones crushed into the ventral surface of the premaxillae (
Figure 7), however precisely identifying these bones is not possible.
The preserved portion of the left palatine of
Parahesperornis consists of a small, caudally directed hook, somewhat crushed into the shaft, which is elongate and very thin mediolaterally. The hook is much more compact than that of
Hesperornis, which is elongate and reaches back further over the shaft (
Figure 17). The midsection of the shaft of the left palatine of
Parahesperornis, immediately rostral to the hook, appears to have a weakly-developed groove, which is more clearly seen in
Hesperornis.
Pterygoids.
Parahesperornis KUVP 2287 preserves both pterygoids; the right is articulated with the right hemipterygoid (
Figure 18), while the left is disarticulated (
Figure 19). For comparison the pterygoid of
Hesperornis (KUVP 71012, YPM 1206) was examined (
Figure 20). The body of the pterygoid of
Parahesperornis is triangular and very thin, with nearly flat sides. In
Hesperornis, the sides are not as flat, with a depression near the articulation for the quadrate on the medial face and a broad groove developed dorsoventrally across the lateral face. The ventral margin of the lateral face of the pterygoid in
Parahesperornis forms a broad, flattened c-shape, while in
Hesperornis it is a deep, nearly enclosed u-shape.
The ventral surface of the pterygoid of Parahesperornis is semicircular and shallowly depressed with a thin crest along the lateral margin, while in Hesperornis it is almost rectangular with a thickened lateral margin bordered by a deep recess. The quadrate articulation of the pterygoid of Parahesperornis is a tiny facet at the caudal end of the triangular body that is more shallowly excavated than in Hesperornis. In Hesperornis a small pointed process is present on the medioventral margin of the pedicel for the parasphenoid articulation (best seen in dorsal view), however this process is absent in Parahesperornis, where the margin is smoothly rounded. In caudal view, the palatine articulation is more concave in Parahesperornis and flattened in Hesperornis with a well-developed lip around the caudodorsal margin.
Hemipterygoid. Parahesperornis KUVP 2287 preserves both hemipterygoids; the left is preserved alongside the braincase and frontals and overlapping the left palatine (
Figure 5), while the right is preserved separately and articulated with the right pterygoid (
Figure 18). For comparison, the hemipterygoid of
Hesperornis (KUVP 71012) was examined (
Figure 21). While the left hemipterygoid identified here was described by Gingerich [
69] as the left palatine, the preserved caudal end agrees much more with that of the right hemipterygoid, as both have a rounded ventral margin and an angled caudal margin.
The hemipterygoid is a very long, thin bone that widens caudally at the articulation with the pterygoid. This caudal expansion is more dramatic in Parahesperornis than in Hesperornis. The hemipterygoid appears not to have been fused or strongly sutured to the pterygoids, as indicated by the disarticulation of the elements. At the caudal end, the lateral and medial surfaces are relatively flat in Parahesperornis, while in Hesperornis the lateral surface is rounded and the medial is broadly indented.
Premaxilla. The premaxillae of
Parahesperornis KUVP 2287 were preserved in articulation with the bulk of the skull (
Figure 4) and are slightly crushed dorsoventrally and to the left side (
Figure 7). The caudal ends appear to narrow before the broken ends, indicating that perhaps not much length has been lost due to breakage. For comparison the premaxilla of
Hesperornis (KUVP 71012, YPM 1206) was examined (
Figure 22). Despite being of a similar width at the rostral end of the nares as
Hesperornis (and accounting for the slight crushing to the element), the length from the tip of the premaxilla to the opening of the nares is much shorter in
Parahesperornis (width at the nares is approximately 35% the pre-nares length) than in
Hesperornis (width at the nares is approximately 20% the pre-nares length), implying a more elongate skull in the latter.
The premaxillae are edentulous with a straight tomial margin, ending in a slight terminal hook, similar to that in
Hesperornis and other Mesozoic ornithuromorphs (
Dingavis [
76],
Changzuiornis [
79]) but much shallower than in
Ichthyornis. The premaxillae of hesperornithids were quite long and narrow, making up close to half the length of the rostrum, as compared to the shorter premaxillae in some other ornithuromorphs (i.e.,
Ichthyornis, Yanornis). This is likely a foraging adaptation for the marine lifestyle of hesperornithiforms. Across Mesozoic birds, lengthening of the rostrum is commonly accomplished by lengthening of the maxilla, contrasted by the elongation of the premaxilla common in Neornithes [
76]. While the maxilla does not appear to be preserved in
Parahesperornis KUVP 2287,
Hesperornis KUVP 71012 preserves a fairly complete maxilla that indicates
Hesperornis achieved rostral elongation in the same way that modern birds do, through elongation of the premaxillae, and unlike the maxillary elongation documented in other Mesozoic birds [
76].
In dorsal view, the suture between the left and right premaxillae of
Parahesperornis is obvious caudally along the entire length of the frontal processes; it becomes fainter rostrally, disappearing about an inch before the rostral-most extent of the premaxillae. In
Hesperornis, this suture is not visible for as much of its length. Both birds have a series of pinhole neurovascular foramina dotting the rostral-most end of the premaxilla, possibly indicating the presence of a keratinized ramphotheca. The maxillary process of the premaxilla is articulated to the maxillary process of the nasal in
Parahesperornis KUVP 2287 (
Figure 7). In ventral view the premaxillae of
Parahesperornis lack impressions for the fitting of the dentary teeth, which are visible in
Hesperornis and some other toothed birds (e.g.,
Ichthyornis). Both
Parahesperornis and
Hesperornis have a small pair of teardrop-shaped pits in the very tip of the ventral premaxillae (f in
Figure 7).
Mesethmoid. A bone which may represent the mesethmoid in
Parahesperornis KUVP 2287 is preserved crushed against the ventral surface of the frontals, adjacent to the palatine (ME in
Figure 5). If this identification is correct, then the bone is lying on its side and has been crushed into the frontals. This bone is broad along the dorsal margin and narrow at the ventral edge. This is similar to the more complete mesethmoid of
Hesperornis as preserved with KUVP 71012 (
Figure 23).
The mesethemoid has been reconstructed as fitting into the gap between the separate rostral-most ends of the frontals [
57], terminating at the anterior end of the frontals, as has recently been described for
Ichthyornis [
61]. Unfortunately, the degree of crushing in KUVP 2287 does not provide additional information about this arrangement, however the current placement of the mesethemoid and the degree of separation of the frontals does not contradict this interpretation.
Angulars. Both angulars of
Parahesperornis KUVP 2287 are preserved and have been glued into rough articulation with the articulars and surangulars (
Figure 24). For comparison the angular of
Hesperornis (KUVP 71012, YPM 1206) was examined (
Figure 25). While angulars have been assigned to
Pasquiaornis [
65], these bones are highly fragmentary and provide little comparative information.
The angular of Parahesperornis is straight along its ventral margin and bows slightly in dorsal or ventral view, as in Hesperornis. The articular surfaces are somewhat obscured caudally by the surangulars. In medial view, the articular surface forms a faint, broad ridge bounded laterally by the thin, flat lateral side of the bone. At the caudal-most end, the articular surface transitions to a flat, dorsally facing surface that is glued onto the surangular in KUVP 2287. In Hesperornis the surangulars are disarticulated, making the entire articular surface visible. The caudal-most end of this articular surface is broad and concave, narrowing rostrally and developing a lateral ridge as in Parahesperornis. However, at the rostral-most end the articular surface of Hesperornis forms a slightly expanded, concave surface unlike the flattened surface of Parahesperornis. The morphology of the dorsal surface implies that the angular Parahesperornis articulated with the surangular through a broader, sheeted surface along its length, while that of Hesperornis was somewhat more restricted to the concave surface.
It should be noted that due to the breakage of the ventral margin of the surangular, the angulars of KUVP 2287 have been glued on imperfectly. The present reconstruction of the left, and more complete, mandible has a slight gap between the surangular and rostral angular. This gap has also been figured in reconstructions of the lower jaw of
Hesperornis that were based off this specimen [
58,
59]. However, the surangulars and angulars of
Hesperornis KUVP 71012 are complete enough to allow a good articulation, and the angular fits smoothly and completely over the surangular, with no gap.
Surangulars. The surangulars of
Parahesperornis KUVP 2287 are preserved in articulation with the articulars and glued to the angulars but crushed and poorly preserved (
Figure 24). For comparison the surangulars of
Baptornis (FMNH 395) and
Hesperornis (KUVP 71012, YPM 1206) (
Figure 25) were examined. The surangular of
Parahesperornis is a very thin bone that articulates with the dorsal surface of the articular just cranial to the pseudotemporale tubercle, where a suture is visible (
Figure 24). In dorsal view, both surangulars of
Parahesperornis are convex, bowing out laterally to a much greater degree than in
Hesperornis.
The medial face of the surangular of Parahesperornis has a broad, shallow depression. A mandibular foramen is not present. The mandibular foramen is also absent in Hesperornis, contra Marsh7. A slight groove is present running diagonally from the ventral margin to the dorsal margin, opening into this depressed area. In Hesperornis this groove is not present, and the depression on the medial face is much more deeply excavated than in Parahesperornis. The lateral face of the right surangular of Parahesperornis KUVP 2287 preserves three foramina, one located near the dorsal margin by the articular and two nearly centered on the lateral surface. While the foramen near the dorsal margin is not present in Hesperornis, the central ones are visible in a similar location. The surangular of Baptornis is minimally preserved, with FMNH 395 only preserving the caudal-most end, which is fully fused to the angular, with a groove marking this fusion in medial view, as in hesperornithids.
Articulars. Both articulars of
Parahesperornis KUVP 2287 are preserved in articulation with the surangulars (
Figure 24). For comparison the articulars of
Baptornis (FMNH 395),
Hesperornis (KUVP 71012;
Figure 25), and
Pasquiaornis (RSM P2989.21, RSM P2989.19) [
65] were examined. The retroarticular process of
Parahesperornis is somewhat oval and dorsally concave, with raised edges that almost form a lip, while that of
Hesperornis appears to be proportionally broader and flatter. The retroarticular process of
Parahesperornis is extremely elongate, slightly more so than in
Hesperornis, however the elongation in both taxa is extreme compared to modern birds. The retroarticular process in
Baptornis is more like that of modern birds, with the caudal end the same width as the remainder of the element and extending only slightly caudally beyond the cotylae. The articular is not fully fused to the surangular, with a suture visible in both
Parahesperornis and
Hesperornis. In
Baptornis these elements appear fully fused, with no suture present, at least at the preserved caudal end.
Rostral to the retroarticular process is a small but very deep depression separating the retroarticular process from the medial articular cotyla of the quadrate articulation. This deep fossa is not present in Hesperornis or Baptornis but is in Pasquiaornis. The medial cotyla is developed as a deep groove oriented almost transversely and angled such that the lateral edge is ventral to the medial edge. In Hesperornis the medial cotyla is oriented even more transversely than in Parahesperornis. The degree of excavation of the medial cotyla is highest in Parahesperornis, reduced but still prominent in Hesperornis and Pasquiaornis, and faint in Baptornis. This cotyla is located primarily on the dorsal surface of the mandible in Parahesperornis, as is also the case in Baptornis, but slightly more offset to the medial side of the mandible in Hesperornis and located almost entirely on the medial side of the mandible in Pasquiaornis.
Medial and slightly rostral to the medial cotyla in Parahesperornis, the caudal cotyla is developed as a small oval facet that is bounded by prominent ridges on all but the rostral margin, which is indistinct. The caudal cotyla in Hesperornis is directly medial to the medial cotyla and not offset rostrally. It is only faintly developed and lacks the prominent margins in Parahesperornis. The caudal cotyla is not discernible in Baptornis FMNH 395. The lateral cotyla of Parahesperornis is very faint and not clearly divided from the caudal, constituting a very faint, flattened face rostral to the caudal condyle. This is also true for Hesperornis.
Splenials. Portions of both splenials are preserved as isolated elements with
Parahesperornis KUVP 2287 (
Figure 26). For comparison the splenials of
Hesperornis (KUVP 71012) and
Pasquiaornis (RSM P2985.9) [
65] were examined (
Figure 27). The splenial of
Parahesperornis has a slightly rounded lateral surface and a medial surface divided lengthwise by Meckel’s groove, which forms a slight shelf for the dentary. Above this shelf the splenial is very thin and appears to form a sheet over the ventral bladed portion of the dentary. This sheeted overlay of the splenial on the dentary is unusual in birds, but is also seen in
Ichthyornis [
61].
The caudal end of the splenial angles ventrally to form an articular surface for the angular. The splenial of Parahesperornis is very similar to that of Hesperornis, including in size, despite the overall larger size of the mandible of Hesperornis. In Parahesperornis the caudal end of the articular surface for the dentary angles more sharply ventrally at the caudal end than in Hesperornis, while in Pasquiaornis65 the transition is smooth and broad, more a curve than an angle.
Dentaries. The dentaries of
Parahesperornis KUVP 2287 are poorly preserved, being incomplete and crushed (
Figure 28). The right dentary preserves more of the length than the left, including the symphysis. For comparison, the dentaries of
Hesperornis (KUVP 71012, YPM 1206) (
Figure 29) and
Pasquiaornis (RSM P 2831.60, RSM P 2985.10, RSM P 2988.11) [
65] were examined, as well as a partial dentary assigned to
Asiahesperornis (IZASK 4/KM 97, but see discussion below) [
80]. All hesperornithiform dentaries show partial individual sockets for the teeth, opening into a dental groove running along the dorsal surface of the dentary [
81] (a dental implantation similar to the ‘aulacodonty’ of non-avian reptiles [
82]. While these partial sockets have been described as “faint” and insufficient to reduce the width of the dental groove [
7], specimens that have broken open show the sockets to be clear divisions within the dentary, but not true sockets as in
Ichthyornis (e.g.,
Hesperornis YPM 1206 and
Pasquiaornis RSM P 2985.10 and RSMP P 2988.11). The dentary fragment assigned to
Asiahesperornis (IZASK 4/KM 97) [
80] shows clear alveoli for the teeth. As this is not the case among other hesperornithiforms, and the
Asiahesperornis material consists entirely of unassociated elements, it is unlikely this specimen belongs to a hesperornithiform bird.
In Parahesperornis, the dental grooves appear similar to those of Hesperornis and expand at the caudal-most end, however the degree of crushing makes identification of more specific features difficult. The dentaries of Parahesperornis appear to preserve a longitudinal groove along the lateral face, as in Hesperornis and Pasquiaornis, however the degree of crushing makes this identification tentative. A pair of medial grooves have been reported in Pasquiaornis, and the right dentary of Parahesperornis appears to have a short groove present near the ventral margin. Hesperornis appears to have a single groove running down both the lateral and medial faces of the dentary.
The symphysis preserved on the right dentary consists of two small bulbous projections stacked vertically. This configuration is also present in
Pasquiaornis (RSM P 2831.6). Some specimens of
Hesperornis (i.e., KUVP 71012) do not show this, having instead a smooth symphysis that tapers to a point (
Figure 29), while others do appear to have the rounded projections (YPM 1206).
Predentaries. Martin first reported the existence of a predentary bone in ornithuromorphs when he described those of hesperornithids [
83]. Within the Hesperornithiformes to date, this bone has only been reported for a single specimen of
Hesperornis (KUVP 71012) and
Parahesperornis (KUVP 2287) [
73] (
Figure 30). The predentary is known in other Mesozoic ornithuromorphs, including
Ichthyornis,
Yanornis, Yixianornis, Hongshanornis, and
Jianchangornis [
84,
85].
The predentary of
Parahesperornis possesses a pair of small facets on the caudal face, the right of which is deformed and flattened. In
Hesperornis KUVP 71012, a single facet appears to be present; however, it is possible that weathering of the element could have obscured the division of the facets. The predentary of
Parahesperornis is more elongate than that of
Hesperornis. While the predentary of
Hesperornis is certainly more weathered than that of
Parahesperornis, this is not enough to account fully for the size discrepancy. Recent work has hypothesized a unique mandibular kinesis in ornithuromorphs that possess a predentary, with a synovial joint involving ventral flexion of the dentaries and translation and compression of the predentary on the dentaries [
85].
Teeth. Parahesperornis KUVP 2287 preserves several teeth, however none are in place in the dentaries, the only identifiable tooth-bearing bone. At least four teeth are visible in the underside of the premaxilla. These teeth could have come from the maxilla or dentaries. Two are easily visible and include the root and crown (
Figure 7 and
Figure 8). Martin [
40] described the teeth of
Parahesperornis as less pointed and recurved than those of
Hesperornis. That observation is likely not accurate, as along the jaw the teeth of
Hesperornis change shape slightly, becoming more recurved rostrally and less recurved caudally [
81]. This appears to be true for
Parahesperornis as well, as the two teeth preserved near each other in the ventral premaxillae are of different curvatures, however their placement within the jaw is unknown so this supposition cannot be confirmed (
Figure 8). A comparison of both teeth preserved in the ventral surface of the premaxilla with the isolated tooth of
Hesperornis KUVP 71012 preserved near the palatine (
Figure 17) shows them to be largely similar, but smaller in size. Similar teeth have also been reported for
Pasquiaornis [
65], however none have been found associated with identifiable skeletal material.
Unidentified Palatal Elements. The ventral surfaces of the caudal premaxillae are filled with numerous small, thin bones jumbled together (
Figure 7 and
Figure 8). While the interpretations of the nasals and premaxillae presented here seem likely, other identifications are less so. At first glance it appears that there are two bones that form a V-shape and would perhaps be the left and right of the same element. However, close examination reveals that the left “bone” is actually three separate bones stacked together. Furthermore, none of these bones have the smoothly rounded ventral margin displayed by the right bone; rather, all three have thin, sharp edges. Of these three bones on the left, the lateral-most bone is the longest, extending as a thin blade past the nares.
Unfortunately, none of these bones can be positively identified at this time, however it is possible they may be portions of the jugals, quadratojugals, maxillae, or vomers, none of which have been identified thus far. It should be noted that there appears to be a tooth preserved in the center of this area in historical photographs (
Figure 4), however the tooth is no longer present today.
3.2.2. Vertebrae
Parahesperornis KUVP 2287 preserves 22 presacral vertebrae, including the axis, and five caudal vertebrae, while KUVP 24090 preserves at least 17 presacral vertebrae, four caudal vertebrae, and the pygostyle. The posterior cervical and thoracic vertebrae of KUVP 24090 are predominately articulated and encased in chalk while those of KUVP 2287 are fully prepared, with some isolated and some articulated sections. The vertebrae of
Parahesperornis discussed here have been identified through a comparison to Marsh’s [
7] figures of
Hesperornis, which were based primarily on YPM 1207, YPM 1476, YPM 1477, and YPM PU 18589, as well as articulations that can be recreated between disarticulated elements.
The process of identification is complicated by the general similarity of neighboring vertebrae in some sections of the vertebral column. For example, vertebrae 18 to 21 are very similar in hesperornithiforms, making their precise identification in
Parahesperornis difficult. However, the morphology of the more cranial vertebrae of hesperornithiforms is distinctive, making certain vertebrae easily identifiable. The following specimens were used for comparison of the presacral vertebrae:
Baptornis (AMNH 5101, KUVP 2290, KUVP 16112);
Canadaga (NMC 41050);
Chupkaornis (MCM.A773) [
12];
Enaliornis (SMC B55277, SMC B55279, SMC B55280);
Fumicollis (UNSM 20030);
Hesperornis (KUVP 2280, KUVP 2289, KUVP 71012, UNSM 4-19-5-36, YPM 1200, YPM 1207, YPM 1476, YPM PU 18589); and
Pasquiaornis (RSM P2831, RSM P2988) [
65].
The presacral vertebrae of hesperornithids can be divided into five general morphotypes to facilitate description [
20]. While each vertebra is unique in terms of small details of morphology, there are several trends that can be used to identify the approximate location in the vertebral column. The presacral vertebrae of
Parahesperornis are described below, in terms of the main morphotypes occurring along the sequence. Marsh [
7] designated the first seventeen vertebrae of
Hesperornis as the cervicals, with the last three (15 to 17) having free ribs. The atlas, or 1st cervical vertebra, is unknown for both
Hesperornis and
Parahesperornis.
Axis. The axis, or second cervical vertebra, is preserved in
Parahesperornis KUVP 2287. It is somewhat crushed laterally and wedge-shaped in side view (
Figure 31). The cranial articular surface forms a shallow, ovate dish, below which is a small, partially preserved ventral process. This process is not seen on the axis of
Hesperornis (YPM 1207, KUVP 71012). The dens is crushed back against the body of the axis, with no discernable features. In
Hesperornis (YPM 1207) the dens extends quite far cranially from the front of the axis, ending in a point in lateral view. Much of the sides of the centrum of KUVP 2287 are obscured by crushing. Near the craniocaudal midpoint of the centrum, the complete spinal process expands upward from the dorsal surface, sloping caudally. The caudal-most portion of the spinal process is rounded and blunt, extending to a height roughly one-third of the total dorsoventral height of the vertebra. This is similar in
Hesperornis. However, the spinal process extends caudally past the end of the vertebral body.
The left postzygapophysis is completely preserved and consists of a domed dorsal surface over a shallow, ventrally directed facet that extends laterally. In
Hesperornis the postzygapophyses are capped by high crests roughly 75% the height of the spinal process. In the axis of
Parahesperornis, the caudal articular surface is taller dorsoventrally than it is wide, with a slight depression along the dorsal margin and a deeper depression on the ventral margin. The center of the caudal articular surface is marked by a small, circular depression that has been suggested to represent the trace of the notochord [
19]. A small projection extends ventrally from the caudal body of the axis in
Parahesperornis, while in
Hesperornis, this expansion forms a prominent crest nearly as large as the spinal process.
Cervical Vertebrae 3 to5. The third throughfifth cervicals of
Parahesperornis KUVP 2287 are preserved in articulation (
Figure 32). Their placement was identified by the ability of the third vertebra to articulate with the axis, as well as the similar shapes of the centra in dorsal view to those of
Hesperornis. Most of the fourth and the dorsocaudal portion of the fifth cervical vertebrae are preserved as isolated elements in
Parahesperornis KUVP 24090 (
Figure 33). The placement of these vertebrae was identified primarily by the shape of the postzygapophyses, which are broad and fan shaped, with only a slight indentation between the zygapophyses.
In hesperornithids, cervical vertebrae three through five are characterized by a triangular shape in dorsal view with a narrow cranial end and broad caudal end. These vertebrae in
Parahesperornis are very narrow mediolaterally with a grooved ventral surface that expands into a fan at the caudal end. The ventral surface is very narrow in the third cervical and broadens along the series. While the anterior cervical vertebrae of
Baptornis are not well known, AMNH 5101 preserves two fragmentary vertebrae that are likely the third and fourth or fourth and fifth cervicals. The ventral surface of these vertebrae is much narrower than in hesperornithids, compressed into a thin crest at the caudal end of the third or fourth vertebra. A cervical proposed as being in the interval of cervical three to six was reported with the holotype of
Brodavis varneri (SDSM 68430) [
31], however the specimen is encrusted with gypsum and preserves little detail. Anterior cervical vertebrae have not been reported for any other hesperornithiform taxa.
In lateral view the centrum of the third cervical has a profile that is dorsoventrally tallest at the caudal end and narrows dramatically toward the cranial end, similar to, but less dramatic than, that of the third cervical of Hesperornis. In hesperornithids, this profile becomes progressively less exaggerated in the fourth and fifth cervicals, such that the fifth appears more rectangular in lateral profile. The lateral sides of the centra are nearly flat, with a slight craniocaudal groove along the ventral margin in Parahesperornis and Hesperornis. The costal processes of these anterior cervicals in Parahesperornis are only known from a short fragment preserved with the fifth cervical of KUVP 2287. Dorsal to where these processes would be in Parahesperornis is a small, oval depression.
The cranial end of the third vertebra of Parahesperornis KUVP 2287 is crushed and that of the others are obscured by articulation. The prezygapophyses are small and closely set, with oval articular surfaces facing dorsomedially, as in Hesperornis. Progressively along the series the prezygapophyses become more widely spaced. The spinal processes are well-preserved and offset caudally on the vertebral body. The spinal processes of these vertebrae are not preserved in any Hesperornis specimen available for study; however, from the broken surfaces they appear similarly located. In lateral view, the spinal processes of Parahesperornis become progressively craniocaudally longer and dorsoventrally shorter in the series, such that the spinal process of the fifth vertebra is almost twice the length of the third but only 75% the height.
The postzygapophyses are robust and expand laterally, making the dorsal outline of the vertebrae triangular. Along the series the articular facets become larger. The dorsal surfaces form peaks that become progressively lower and more rounded along the series. Breakage of the corresponding vertebra of Hesperornis makes these features difficult to compare; however, in the fifth cervical of YPM 1207 the dorsal surface of the left postzygapophysis is very highly peaked and does not appear rounded like that of Parahesperornis. Unlike the articular surfaces of the postzygapophyses of the preceding vertebrae, which are directed ventrally, those of the fifth cervical of Parahesperornis are directed slightly ventrolaterally, similar to the orientation of the fourth and fifth cervicals in Hesperornis.
Cervical Vertebrae 6 to 9. Within this series,
Parahesperornis KUVP 2287 preserves the articulated sixth and seventh vertebrae, the isolated eighth, and the ninth articulated to the tenth (
Figure 32), while KUVP 24090 preserves the isolated sixth or seventh vertebrae (
Figure 33). The sixth to ninth cervical vertebrae of hesperornithids can be characterized by the division of the postzygapophyses into two distinct articular surfaces in dorsal view, which become progressively separated along the series. In dorsal view these vertebrae resemble an elongate X-shape instead of the triangular outline of the more anterior vertebrae. Other trends that began in the previous vertebrae are continued, such as the broadening of the ventral surface of the centra.
In cranial view, the articular surface of the sixth cervical narrows dramatically dorsoventrally in the center and wraps around and under the ventral margin of the centrum, expanding into the costal process on the sides (were they preserved). Further along the series, the center expands and is not as narrow. This is also seen in Hesperornis. The prezygapophyses are more circular than in the preceding vertebrae, with the articular facets steeply angled toward the midline of the centrum.
In lateral view, vertebrae six and seven are nearly rectangular in outline, with the caudal end only slightly taller than the cranial, and slightly more elongate than the preceding vertebrae. Progressing along the series shows this changing as the ventral margin angles downward, resulting in a more wedge-shaped outline in side view. The ventral margin flattens from the slight arch seen in the preceding vertebrae, and unlike the pronounced arch seen in the corresponding vertebrae of Hesperornis. Neither the costal nor spinal processes are preserved on these vertebrae in either specimen of Parahesperornis. From the broken area on the centrum it appears the spinal processes are longer and more centrally located on the centrum than in the previous vertebrae, as is also seen in Hesperornis.
The ventral surfaces of these vertebrae are shaped like an elongate hourglass, with a slight indent toward the caudal end. As in the more anterior vertebrae, the ventral surface is grooved along its length. This is also observed in
Hesperornis and perhaps in
Baptornis, however the cervical vertebrae are poorly preserved for that animal. In
Baptornis the ventral surface remains very narrow, much more so than in the corresponding vertebrae of hesperornithids. A vertebra belonging to this range has been assigned to
Pasquiaornis (RSM P2626.15) [
65] which is like
Baptornis in having a very narrow constriction of the ventral surface.
Unlike the previous vertebrae, in caudal view, the articular facets of the postzygapophyses face ventrally and slightly laterally, a trend that will continue in the subsequent vertebrae. This is similar to the case in Hesperornis but possibly different from that of Baptornis, where the postzygapophyses are spread further apart, angling outward to a degree not seen until around the fifteenth cervical in Hesperornis and Parahesperornis. In Parahesperornis, the postzygapophyses of these vertebrae have rounded margins in dorsal view and are separated by a slight indentation, as in Hesperornis, while in Pasquiaornis the postzygapophyses are more broadly spread, but without the projection seen in Baptornis. The eighth cervical of Parahesperornis is the first in the series to have high projections on the lateral margins of the prezygapophyses, as opposed to the rounded surfaces in the preceding vertebrae. The corresponding region is not preserved in Hesperornis.
Cervical Vertebrae 10 to 13. Within this series,
Parahesperornis KUVP 2287 preserves the tenth cervical articulated to the ninth (
Figure 32), the isolated eleventh, and the articulated twelfth, 13th, and 14th (
Figure 34), while KUVP 24090 preserves the eleventh through 13th articulated in a slab (
Figure 35,
Figure 36,
Figure 37 and
Figure 38). The tenth to 13th cervical vertebrae of
Parahesperornis are characterized by shortening of the centrum, as in
Hesperornis, and the development of the postzygapophyses into widely spaced structures extending out from the centrum, a trend that is continued in the subsequent vertebrae. In
Hesperornis these vertebrae are more compact than in
Parahesperornis. A cervical vertebra proposed to be the twelfth, 13th, or 14th has been assigned to
Enaliornis (YORYMG 507) [
21]. Very little detail is preserved on that specimen; however, it is very compact and not elongated like those of other hesperornithiforms.
Cranially, the prezygapophyses are widely spaced and extend laterally past the edges of the cranial articular surface, which is rectangular. The articular facets of the prezygapophyses are angled to face dorsomedially, as in the preceding vertebrae. This is not seen in Hesperornis, where the articular facets face dorsally and are not angled. The cranial articular surfaces are narrower than in preceding vertebrae, as in Hesperornis. In lateral view, beginning with the eleventh cervical, the ventral margin arches dorsally, more like the state in the preceding vertebrae of Hesperornis. The sides of the centrum are nearly flat in the eleventh cervical but become broadly depressed in the subsequent vertebrae.
The ventral surfaces of these vertebrae are similar to those of the sixthto ninth cervicals, but do not narrow at the midlength. An isolated vertebra that corresponds most closely to the eleventh of hesperornithids has been reported for
Pasquiaornis (RSM P2831.8) [
65]. This vertebra is very similar in overall morphology, but more elongate and with a narrower ventral surface, including the cranial and caudal articular faces, as is also the case in the corresponding vertebrae of
Baptornis (AMNH 5101).
The postzygapophyses are widely spaced and elevated above the height of the centrum, extending laterally from the caudal end of the centrum, as in Hesperornis. The postzygapophyses of Baptornis are exaggerated in this regard, extending dramatically from the end of the centrum on long necks. In dorsal view, the margins of the postzygapophyses are not smoothly rounded, as in the preceding vertebrae, but rather have more angular margins, a pattern which begins earlier, in the ninth cervical of Hesperornis. The postzygapophyses have peaked dorsal surfaces, as in Hesperornis. The peaks are more prominent and the postzygapophyses more elongate and widely spaced in Baptornis. The caudal articular surfaces jut outwards from the centrum, forming a shelf onto which the neural foramen opens. The articular surface is very narrow and curved, wrapping around the caudal end of the vertebrae and onto the sides to a greater degree than in the previous vertebrae, a trend continued in the subsequent vertebrae. This is also seen in Hesperornis and Baptornis.
Cervical Vertebrae 14 to 15. Within this series,
Parahesperornis KUVP 2287 preserves the 14th vertebrae articulated to the twelfth and 13th and the isolated 15th (
Figure 34), while the 14th and 15th vertebrae of KUVP 24090 are preserved articulated but unprepared (
Figure 35 and
Figure 36). The 14th and 15th cervical vertebrae of
Parahesperornis are characterized by continued shortening of the centra. These are the first vertebrae to have transverse processes, as is also the case in
Hesperornis. It is unclear whether the 14th cervical of
Baptornis had transverse processes. Elaborate ventral processes are found in these vertebrae originating from the cranial end of the centrum. In
Parahesperornis the 16th cervical has more in common morphologically with the subsequent vertebrae than with the preceding. This is very different from
Hesperornis, where the 16th cervical shares the blocky, robust appearance of the 14th and 15th vertebrae, as well as possessing a short, robust ventral process. A 14th or 15th cervical vertebra has been assigned to
Asiahesperornis (IZASK 2/KM 97) [
80], which is morphologically similar to the cervical vertebrae of
Hesperornis despite being closer in size to those of
Parahesperornis. The cranial articular surfaces of these vertebrae are much more robust than in either previous or subsequent vertebrae in the series, with thickened lateral margins that extend almost as wide as the prezygapophyses. In Parahesperornis and Hesperornis the ventral margins of the cranial articular surfaces form a broad arch, while in Pasquiaornis this arch is much narrower and deeper. The prezygapophyses are widely spaced with the articular surfaces facing dorsomedially, as in previous vertebrae. This is also seen in Hesperornis, whereas the orientation of the prezygapophyses in the preceding vertebrae was less angled. The neural spines are not preserved, but the scar indicates they were very short and caudally positioned on the centrum, as in Hesperornis. There is a deep, teardrop-shaped foramen present just caudal to the neural spine. This foramen is not seen in the corresponding cervicals of Hesperornis YPM 1206 but is present on the 15th cervical of Canadaga.
In lateral view, the 14th and 15th cervicals are the first vertebrae in the series with well-developed lateral concavities, as in Hesperornis. The lateral concavities of Parahesperornis are generally shallower than those of Hesperornis, while Baptornis has lateral concavities more deeply excavated than either hesperornithid. It should be noted that the excavation of the lateral concavity is uneven between the left and right sides of individual vertebrae in hesperornithiforms, as well as modern birds, making this feature of limited use in identification or diagnosis. A rudimentary facet that may be an articulation for a free rib is present on the cranioventral corner of the centrum in the 15th vertebrae, while the first well-developed rib articulation is present on the 16th cervical in Parahesperornis.
Beginning with the 14th vertebrae, the ventral surfaces are characterized by elaborate, often unique processes that are rarely preserved. While Parahesperornis KUVP 2287 does not preserve many of these processes, their general position can be determined by the pattern of breakage on the ventral surface. The corresponding vertebrae of Baptornis show similar evidence of breakage that may also indicate the presence of these processes. The ventral process of the 14th and 15th vertebrae differ from those of subsequent vertebrae by being very robust in their origin on the cranial end of the ventral surface and not as long, while the ventral processes of the thoracic vertebrae are very compressed mediolaterally and originate from the center of the ventral surface.
The 14th cervical of Parahesperornis has a broad, flaring surface extending from the ventral margins of the cranial articular surface that is broken along both margins. This paired breakage is suggestive of the pair of elongate, ventrally projecting flanges that protrude from the cranial end of the ventral surface of the centrum in Hesperornis. This morphology is unique to the 14th cervical of Hesperornis, making it readily identifiable. This is not the case in Parahesperornis, where the 15th cervical shows a similar pattern of breakage, suggesting a second vertebrae with paired ventral projections. In Hesperornis, however, the 15th cervical has a broad, thick ventral process descending from the central portion of the centrum and ending in a forked structure. While Canadaga NMC 41050 does not preserve the ventral processes completely, the remnant present in the 15th cervical resembles that of Hesperornis.
The ventral surfaces of the 14th and 15th vertebrae of Parahesperornis are the first in the series to have the exaggerated hourglass shape typical of the thoracic vertebrae of hesperornithiforms, with a narrow waist near the midline of the centrum and a flared, fan-shaped caudal margin. The waist is less pronounced in most hesperornithids than in Baptornis. The exception is Canadaga, a hesperornithid with extreme expansion of the cranial and caudal articular surfaces and a dramatic central waist in ventral view.
The postzygapophyses of these vertebrae have highly elongate necks that extend far past the centrum caudally, as is seen in earlier cervical vertebrae in Baptornis. The articular facets are ventrolaterally directed, with rounded caudal margins that do not extend past the margins of the neck, unlike in Hesperornis where the caudal ends are highly circular and expanded. In caudal view, the articular surface is narrow, with a waisted appearance and a flared ventral margin. The dorsal margin of the articular surface is narrower than that of Hesperornis.
Cervical vertebra 16 to 17 and thoracic vertebrae 18 to 23. Marsh [
7] began the series of thoracic vertebrae with the 18th vertebra, as it was the first vertebra to articulate with the sternum through a sternal rib. Despite this, both
Hesperornis (the 17th vertebra) and
Parahesperornis (the 16th and 17th) have preceding vertebrae that bear a closer overall resemblance to the 18th vertebrae (
Figure 39), and so these vertebrae are discussed together.
The appearance of the 16th vertebra of Parahesperornis represents a transition along the vertebral column. The anterior cervical vertebrae of hesperornithids, particularly eleven through 15, each show unique features that make identifying them relatively easy as compared to the thoracic vertebrae, which are more similar. These features include things like the elaborate ventral processes and the shapes of the postzygapophyses in both dorsal and lateral views, as described above. The 16th vertebrae of Parahesperornis is interesting in that it clearly shows the beginnings of a number of features common among the subsequent vertebra, but not as developed. This includes features such as comparatively shorter and caudally-directed postzygapophyses, the sides of the centrum less waisted in ventral view than in the previous vertebrae, and the loss of the robust lateral expansions of the ventral half of the cranial articular surface. In Hesperornis, this sort of transitional appearance is instead found in the 17th vertebra.
As described above, the 16th to 23rd vertebrae of
Parahesperornis are remarkably similar in form (
Figure 38,
Figure 39 and
Figure 40), a departure from the previous vertebrae which were more variable (
Figure 32,
Figure 33 and
Figure 34). This similarity makes definitive placement within the series difficult unless articulated. While KUVP 24090 preserves the articulated cervicothoracic series, the chalk they are preserved in obscures many details of morphology necessary for specific vertebral identification (
Figure 35 and
Figure 37). For the disarticulated KUVP 2287, identifications were made through the fit of articulations as well as small morphological differences, in particular, the placement of the 18th through 20th vertebrae is tentative.
Two fairly well-preserved thoracic vertebrae have been assigned to
Pasquiaornis (RSMP P2957.15, and an additional un-numbered vertebra [
65]). They have been described as “slightly amphicoelus” [
65], which one specimen appears to be (RSM P2957.15) [
65]. Another specimen (incorrectly labeled as RSM P2957.15 [
65]), however, is heterocoelus. They appear to be highly pneumatic as well, with numerous foramina on the centra around the transverse processes and toward the caudal margin. Breakage along the spinal and transverse processes shows extensive pneumatization within the bone, more so than is seen in corresponding areas of breakage in hesperornithids. Thoracic vertebrae are also known for
Brodavis varneri (SDSM 6843) [
31], however they are heavily encrusted in gypsum and very little can be said about the morphology other than that they seem to agree with other hesperornithiforms.
Chupkaornis preserves a vertebra identified as the 16th [
12], which is poorly preserved but agrees in general appearance with that of
Parahesperornis.
The 16th to 23rd vertebrae of
Parahesperornis are characterized by shortened and oval-shaped postzygapophyses; tall and centrally-located spinal processes; deep lateral concavities on the centra; progressively dorsally-displaced costal fovea; cranial and caudal articular surfaces expanded, but not as much as in the previous posterior cervical vertebrae; centrally-located transverse processes with deep depressions on the cranial and caudal faces; narrow and centrally-placed ventral processes that decrease in size progressively in the series; and are fully heterocoelous. Overall, this is very similar to the thoracic vertebrae of
Hesperornis and
Baptornis. As noted earlier, the degree of excavation of the lateral concavities is highly variable within species and even individuals. Some specimens assigned to
Baptornis have very faint lateral concavities on the centra of the thoracic vertebrae (AMNH 5101, SDSM 5893), while in others the concavities are developed to a similar degree as seen in hesperornithids (KUVP 2280, KUVP 2290). A single well-preserved anterior thoracic vertebra assigned to
Pasquiaornis (RSM P2988.12) [
65] has deeply excavated lateral concavities, as does
Canadaga. The preserved 15th–18th vertebrae of
Canadaga (NMC 41050, NUVF 284) show extreme excavation of the lateral concavities, almost to the extent of looking like large fossae.
The spinal processes of these vertebrae originate at the caudal end of the centrum and become progressively longer along the series. The spinal process of the 16th vertebrae occupies roughly half the length of the centrum, while that of the 23rd is almost twice as long. This is also the case in
Hesperornis, however in
Baptornis and
Pasquiaornis the spinal processes occupy most of the dorsal surface of the centrum in the preserved thoracic vertebrae. Little can be said about the height of the spinal processes; however, that of the 23rd is nearly complete and extends very straight dorsally to a height roughly equivalent to that of the centrum (
Figure 40). In
Hesperornis the spinal process of the 23rd vertebrae is even taller, exceeding the height of the centrum.
The transverse processes are not well preserved in any of these vertebrae of Parahesperornis. They are located slightly toward the cranial end of the centrum and become narrower along the series, as in Hesperornis. The origination of the transverse process on the centrum bears deep triangular fossae on both the cranial and caudal sides. In Hesperornis, deep grooves are present in the same position but extending out along the transverse process. In Baptornis, these fossae are limited to the sides of the centrum, and do not extend onto the transverse processes. The preserved portions of the transverse processes are oriented directly laterally in Parahesperornis. This is similar in Hesperornis for the more anterior thoracics, but those of the 22nd and 23rd are oriented slightly cranially.
In Parahesperornis, the 16th is the first vertebra in the series with a well-developed, oval fossa for an articulation to a free rib. This fossa is located on the cranioventral margin of the lateral side of the centrum and is slightly angled to face cranially. This fossa is well-developed in the corresponding vertebra of both Hesperornis and Baptornis, however in both of those taxa it is oriented directly laterally. The costal foveae of the subsequent vertebrae in Parahesperornis are directed laterally, as in other hesperornithiforms. The placement of the costal foveae on the centra changes along the series, migrating dorsally such that in the 22nd vertebra it is nearly at the height of the transverse process. The shape also changes, being oval in most of the vertebrae but circular in the 19th to 21st. These trends are also seen in Hesperornis. In Baptornis the position of the costal foveae changes similarly, however they are more consistently circular in that taxon. The 23rd vertebra has only a faint oval face and not a well-developed costal fovea, as in Hesperornis and Baptornis.
While most of the ventral processes are not preserved in KUVP 2287, the scar along the ventral surface indicates they were very narrow and centrally located, unlike the previous ventral processes that were offset cranially and robust. Like in
Hesperornis, the ventral processes took up most of the ventral surface of the centra in the more cranial thoracic vertebrae, but they become craniocaudally shorter along the series. In
Parahesperornis the reduction of the ventral processes is greater than in the corresponding vertebrae of
Hesperornis. This shortening of the ventral process is present only in the 22nd vertebra of
Baptornis, while the 21st appears to have had the same width of process as the preceding thoracic vertebrae. Ventral processes appear to only be present on the more cranial thoracic vertebrae of
Pasquiaornis [
65]. However, definitive identification of the isolated specimens is not possible.
Only the complete ventral process of the 20th vertebra is known for Parahesperornis. It extends ventrally to a length approximately equal to the height of the centrum and has a forked end that has been somewhat crushed. Hesperornis UNSM 4-19-5-36 has an identical ventral process on the corresponding vertebra. It should be noted that the articulated vertebral column of KUVP 24090 may well preserve these processes, however without preparation these features cannot be studied. The ventral surfaces of these vertebrae continue the hourglass outline typical of hesperornithiforms and seen in the 14th and 15th vertebrae. This shape becomes more symmetrical along the series, with the fan-shaped expansion of the caudal end being almost as wide as that of the cranial end.
The 23rd vertebra is unique among the thoracics in not having a ventral process, as is also the condition in other hesperornithiforms. This vertebra has also been reported for
Enaliornis (SMC B55277) [
21]. While the identification as the 23rd is not definitive for an isolated specimen, it does lack a ventral process. It is also clearly heterocoelus [
16,
21], contra Tanaka et al. [
12]
The postzygapophyses of these vertebrae are in general closely spaced, short, and oriented directly caudally. Those of the 16th are somewhat intermediate between the elongate postzygapophyses of the previous vertebrae and the reduced form seen in the latter. The caudal articular surfaces are robust and indented on all margins, being narrower on the dorsal margin than the ventral. This discrepancy in widths increases along the series. In cranial view the thoracics of Parahesperornis and very similar to those of Hesperornis.
Synsacrum. The synsacrum of
Parahesperornis KUVP 2287 is preserved in articulation within the pelvis, and thus most anatomical features of this compound bone are obscured (
Figure 41). It appears to contain eleven sutured or fused centra, however breakage makes this number difficult to confirm. The sacrum of KUVP 24090 is preserved primarily within the pelvis and encased in matrix. On one side of the slab, the first two sutured centra are exposed (
Figure 37 and
Figure 38). These are identified as belonging to the sacral vertebrae, and not to the caudal-most thoracic vertebrae, by the completely smooth ventral surfaces, lacking even the faint ventral crest visible on the 23rd thoracic vertebra of
Parahesperornis. Marsh [
7] described the synsacrum of
Hesperornis YPM 1207 as having fourteen centra in the ankylosed section, more than in
Parahesperornis, however the degree of fusion in YPM 1207 makes this assertion difficult to confirm. Lucas [
13] and Martin and Tate [
19] both reported the number of centra in the synsacrum of
Baptornis (AMNH 5101) as ten; however, due to crushing this is also difficult to confirm. In other hesperornithiforms preserving the synsacrum, such as
Enaliornis and
Brodavis varneri, this compound bone is not complete.
The vertebrae incorporated into the synsacrum of Parahesperornis increase in the degree of joining along the series, with the first and second sutured and their margins clearly delineated, while further along the synsacrum the vertebrae are fully fused, with margins obscured. This is also seen in Hesperornis and Pasquiaornis. However, in Baptornis, the cranial centra are fully fused, with no discernable boundaries between them, while the caudal centra appear to be sutured and not fully fused. The first centrum in the synsacrum of Parahesperornis KUVP 2287 is crushed dorsally, obscuring the neural spine. The cranial articular surface is broad, shallowly excavated, and rectangular in outline. In dorsal view the centrum is similar to that of the 23rd vertebra. However, the caudal articular surface is much narrower than the cranial. This appears to also be the case in the second centrum. However, the states in the other centra are difficult to determine due to the degree of fusion. In Baptornis, the cranial articular end of the first centrum in the synsacrum is only slightly wider than the caudal end in dorsal view (as determined from the indentation in the sides of the centrum in dorsal view, as the exact caudal margin is fused and not visible).
A very short, somewhat stubby transverse process is preserved on the right side of the first centrum in KUVP 2287. A reduced transverse process is also present on the first centrum of the synsacrum in Hesperornis. Dorsally, the width of the synsacrum narrows dramatically from around the level of the acetabulum caudally, as in Hesperornis, while in Baptornis the caudal synsacrum does not become mediolaterally restricted until well past the level of the acetabulum. Overall, the portions of the synsacrum of Parahesperornis KUVP 2287 that are visible suggest a great similarity with this compound bone in Hesperornis.
Caudal vertebrae. Parahesperornis KUVP 2287 preserves four isolated free caudal vertebrae (
Figure 42), while KUVP 24090 preserves one isolated and three articulated free caudal vertebrae (
Figure 43 and
Figure 44). For comparison, the caudal vertebrae of
Hesperornis YPM 1200 were examined. The caudal vertebrae of
Parahesperornis possess extremely wide transverse processes that curve slightly cranially in dorsal view. While the width is similar between
Parahesperornis and
Hesperornis, the transverse processes of
Hesperornis do not curve, but instead extend straight but at an angle cranially from the centrum. The caudal-most vertebrae have thinner and progressively shorter transverse processes, despite having only slightly shorter centrum lengths.
None of the spinal processes are preserved in KUVP 24090, however some of the free caudal vertebrae of KUVP 2287 preserve very tall spinal processes. The most complete caudal series of Hesperornis consists of ten free caudal vertebrae preserved in YPM 1200; however, it cannot be determined if the caudal series of this specimen is complete, as the individual elements are disarticulated. Given the overall similarities of Parahesperornis caudal vertebrae with those of Hesperornis, it is reasonable to posit that Parahesperornis may have had at least ten caudal vertebrae, the same as in Hesperornis.
Pygostyle. The complete pygostyle is preserved in
Parahesperornis KUVP 24090 in dorsal view in a slab containing part of the pelvis and hindlimb (
Figure 43 and
Figure 44) and is unknown for KUVP 2287. A partial pygostyle preserving two full centra and a partial third is known from
Hesperornis (YPM 1200) (as described by Marsh [
7] and contra Martin and Tate [
19]), and another preserving three centra is known from
Fumicollis (UNSM 20030). Martin and Tate [
19] described the pygostyle of
Baptornis on the basis of UNSM 20030, which has since been reassigned as the holotype of
Fumicollis [
20], and FNMH 395, which does not include a pygostyle today. It is therefore not possible to confirm Martin and Tate’s assertion [
19] of five fused centra in the pygostgyle of
Baptornis.
The pygostyle of Parahesperornis preserves three fused centra. The first centrum has moderate transverse processes, while those on the second are rudimentary and those on the third are absent (though possibly broken). A dorsal ridge is present along the center of the fused centra. The transverse processes of the first centrum of the pygostyle of Hesperornis appear to be proportionally of the same length as those of Parahesperornis, however much more robust, while those of the second centrum are only partially preserved. Similarities among these elements indicate that the pygostyle of more derived hesperornithiforms was small and formed by just a few vertebrae.
3.2.6. Hindlimb
Femur. The left and right femora of
Parahesperornis are preserved with both KUVP 2287 (
Figure 54) and KUVP 24090 (
Figure 43 and
Figure 55). A large number of hesperornithiform femora are known; it is one of the best represented elements for the group. The following specimens were used for comparison to
Parahesperornis:
Baptornis (KUVP 2290, FMNH 395);
Brodavis varneri (SDSM 68430);
Enaliornis (BMNH A163, BMNH A479, BMNH A5801, SMC B55289, SMC B55300, SMC B55287);
Fumicollis (UNSM 20030);
Hesperornis (YPM 1200, YPM 1207, YPM 1476, YPM 1477); and
Pasquiaornis (RSM P2997.4) (
Figure 56).
The femur of Parahesperornis displays a number of typical hesperornithiform traits, such as an expanded trochanter and fibular condyle. The proximal and distal ends are evenly expanded mediolaterally, with a narrowed shaft. Slightly proximal to the midpoint of the shaft is waisted, intermediate to the dramatic waist seen in Hesperornis and the straighter shaft of other hesperornithiforms.
In medial and lateral views, the femur is bowed, an appearance exaggerated by the high arch of the cranial surface. This condition is better developed in KUVP 2287 than in KUVP 24090. When compared to other hesperornithiforms, KUVP 24090 is most similar in this regard to Hesperornis, however the cranial arch is more dramatic in KUVP 2287. Other hesperornithiform femora vary widely in this feature, with Pasquiaornis and Enaliornis having strongly bowed shafts, Fumicollis only bowed at the caudal end, and a nearly straight shaft on Baptornis.
In proximal view, the articular surface of the femur of Parahesperornis is shaped like a figure eight, with the nearly circular head and the ovate trochanter separated by a narrow, pinched region. The depression for the capitol ligament on the head of the femur is directed proximally, such that it is visible in proximal view. The trochanter narrows to a point on the craniolateral margin of the proximal surface, with a flattened face on the lateral half. In proximal view the craniocaudal width of the head is greater than that of the trochanter.
In cranial or caudal view, the head is directed proximally and extends past the top of the trochanter, more so in KUVP 2287 than KUVP 24090. This condition differs from that of other hesperornithiforms where the head is either even with the trochanter in proximal extent (Hesperornis, Baptornis) or does not extend as far proximally (Fumicollis, Pasquiaornis, possibly Enaliornis). The head and trochanter are separated along the proximal margin by a deep indentation, as in other hesperornithiforms. Distal to this indentation on the caudal face of the proximal end, a pronounced ridge is present running mediolaterally across the cranial face, just below the articular surface, extending from the distal margin of the head to the trochanter. In KUVP 2287 this ridge is straight, while in KUVP 24090 it arches proximally, with the peak just below the indentation between the head and trochanter. This ridge is also seen in Hesperornis, where it is straight.
In medial view, the head of the femur forms a rounded ball with a slightly flattened distal margin set atop a much narrower neck, as in Hesperornis and Fumicollis. Baptornis also has a distinct neck below the femoral head, but the distal margin of the head is smoothly rounded. In both Pasquiaornis and Enaliornis, the neck appears to be less distinct, however the known femora of Enaliornis are also highly eroded, which could contribute to the appearance of lacking a neck.
In caudal view, the trochanter forms a broad ridge that wraps around the lateral margin of the femur and ends abruptly about one-fourth the way down the shaft. A small projection is present where the trochanter meets the shaft. Along both the cranial and caudal faces of the femur the margins of the trochanter are clearly delineated. This is also seen in Hesperornis. However, in other hesperornithiforms, the trochanter merges more smoothly into the shaft and is less laterally projected. The caudal face of the proximal femur of Parahesperornis forms a deep depression medial to the margin of the trochanter, whereas this area is flattened in Hesperornis. In lateral view the trochanter of Parahesperornis is highly rugose and irregular, as in Hesperornis. Due to the bowing of the shaft, the trochanter is angled slightly caudally at the proximal end.
The femoral shaft of
Parahesperornis is marked by several rugose muscle scars, as in
Hesperornis, to a somewhat lesser degree in
Fumicollis, and to a much lesser degree in other hesperornithiforms. One of the most prominent muscle scars is a large tuberosity for the m. gastrocnemius lateralis, which extends the foot during the propulsive stroke [
86], located toward the distal end of the lateral side of the shaft. Also prominent is a narrow ridge for the m. puboischiofemorales medialis and the area of attachment of the aponeuroses of the m. popliteus which runs diagonally across the proximal end of the caudal shaft to the distal end of the medial shaft. The m. puboischiofemorales medialis retracts the femur while the m. popliteus stabilizes the knee and inwardly rotates the tarsometatarsus [
86]. Galton and Martin [
21] describe the attachment for the m. femorotibialis externus as being level with that for the m. gastrocnemius lateralis in
Parahesperornis; however, the scar for the m. femorotibialis externus is actually distal to that of the m. gastrocnemius lateralis, as has been described for
Hesperornis [
86]. Additionally, the scar for the m. iliofemoralis internus is slightly proximal to that of the m. iliotrochantericus cranialis in
Parahesperornis, while in
Hesperornis they are nearly even [
86], also contra Galton and Martin [
21].
The shaft of the femur narrows into a slight waist mediolaterally just above the midline of the shaft, with the lateral side narrowing more than the medial, which is nearly straight. In medial and lateral views, the shaft is thickest the point of maximum inflection of the cranial surface, at and proximal to midshaft, as in Hesperornis. In Pasquiaornis, Enaliornis, and Baptornis, the femoral shaft is only slightly thicker in the midshaft region. The distal lateral shaft of the femur of Parahesperornis narrows proximal to the lateral condyle, but to a lesser degree than in Hesperornis and Fumicollis, where the distal shaft of the femur narrows into a neck just above the lateral condyle.
In cranial and caudal views, the femur of Parahesperornis widens from the distal shaft primarily on the lateral side through the dramatic expansion of the lateral condyle common in hesperornithids. The lateral condyle is made up of the caudally directed fibular trochlea and a depression for the articulation of the fibular head, with the tibiofibular crest on the medial margin and a second crest on the lateral margin. The fibular trochlea is oriented to face caudolaterally, as is the case in other hesperornithiforms.
Overall, the lateral condyle is much larger than the medial, as in most hesperornithiforms, but to a greater degree in Parahesperornis and Hesperornis. In most hesperornithiforms the lateral condyle extends much further distally than the medial condyle, more so than in Parahesperornis. However, in Hesperornis, the medial and lateral condyles are of similar distal extent, making Parahesperornis intermediate to Hesperornis and other hesperornithiforms in this regard.
In lateral view, the lateral condyle is semicircular, with a depressed center and a sharp angle at the proximal edge of the caudal margin. The caudal surface of the lateral condyle is a smooth, rounded bulge, as in other hesperornithiforms. The distal surface of the lateral condyle is flattened and bulges slightly, as in Hesperornis and Fumicollis; in other hesperornithiforms the distal surface of the lateral condyle is smoothly rounded. In distal view the lateral orientation of the condyle is most obvious, being intermediate between Hesperornis and Fumicollis, which are angled more laterally, and Baptornis and Brodavis, which are slightly less angled.
The medial and lateral condyles of Parahesperornis are separated by a deep intercondylar sulcus, as in other hesperornithiforms. In cranial view the distal surface of the shaft above the intercondylar sulcus forms a depressed patellar sulcus that is bounded distally by a ridge, giving it a pocketed appearance as in Hesperornis and to a lesser degree in Fumicollis; this distinct morphology is absent in other hesperornithiforms. In caudal view, the distal shaft between the condyles forms a deep popliteal fossa that extends downward into a deep excavation behind the ridge of bone separating the medial and lateral condyles.
The medial condyle is ovate and somewhat kidney-shaped in caudal view and set at an angle to the long axis of the shaft, as in Hesperornis and Fumicollis. In medial view the medial condyle resembles the lateral but is distinctly smaller. The caudal margin of the medial condyle forms a lip that projects medially, and an elongate tuberosity for the medial collateral ligament is present above the face of the condyle. This condyle is more elongate in medial view in Hesperornis, with a rounder tuberosity. The distal shaft narrows into a slight neck just proximal to the medial condyle, as in Hesperornis. In distal view the long axis of the medial condyle is oriented perpendicular to the mediolateral axis of the distal end, as in other hesperornithiforms. The medially expanded lip on the cranial margin gives the medial margin of the condyle a c-shaped contour in distal view; this condition is similar to that of Hesperornis and more pronounced than the condition in other hesperornithiforms.
Patella. The patella of
Parahesperornis is preserved with both KUVP 2287 and KUVP 24090 (
Figure 57), the left of which is encased in chalk (
Figure 43 and
Figure 44). Comparisons were made with the following hesperornithiform specimens:
Baptornis (KUVP 2290);
Fumicollis (UNSM 20030); and
Hesperornis (YPM 1200, YPM 1207) (
Figure 58). Patellae are unknown for other hesperornithiform taxa.
In general, the patella of Parahesperornis is very similar to that of Hesperornis. Hesperornithid patellae differ dramatically from those of other hesperornithiforms in being extremely elongate, with a shape like an isosceles pyramid with elongate sides and a very short base, as opposed to the nearly equilateral pyramid seen in Baptornis and Fumicollis, where the sides and base are similar lengths. The patella of Parahesperornis is proportionally much less elongated than that of Hesperornis. The distal width is approximately 50% of the length in Parahesperornis, while in Hesperornis it is approximately 35%. In cranial view the medial margin is curved while the lateral margin is straight and the distal margin angles down to the medial margin, as in Hesperornis. The laterodistal corner of the cranial face is penetrated by a small foramen for the ambiens tendon, which emerges as a larger foramen on the lateral side of the patella. This foramen is a thinner slit in Baptornis and a broader teardrop shape in Fumicollis.
In lateral view the patella of Parahesperornis is extremely thin, like that of Hesperornis. The patellae of both Baptornis and Fumicollis are much wider in this view, comparable to the width in cranial view. The proximal portion of the patella is very thin craniocaudally, leading to a high degree of deformation in most specimens of both Parahesperornis and Hesperornis. The distal face of the patella is set at a high angle, such that it is visible in caudal and medial views, as in Hesperornis. The completely preserved patella of KUVP 2287 is also curved caudally along the proximal half, as is the patella of Baptornis, while those of KUVP 24090 and Hesperornis are straight. This difference does not seem to be the result of taphonomy.
Fibula. The left fibula of
Parahesperornis is preserved with KUVP 2287 (
Figure 59) and both are preserved with KUVP 24090 (
Figure 43 and
Figure 44). The total length of KUVP 2287 is preserved, however the distal extent is broken as a separate piece that can be cleanly fitted back to the remainder of the shaft. The left fibula of KUVP 24090 is completely preserved and exposed in caudal view, in articulation with the tibiotarsus, encased in chalk. To the best of the authors’ knowledge, these are the best preserved hesperornithiform fibulae, as all other specimens examined or published preserve a lesser portion of the shaft or are broken and reconstructed. Comparisons were made with the following specimens:
Baptornis (KUVP 2290);
Brodavis varneri (SDSM 68430);
Fumicollis (UNSM 20030);
Hesperornis (AMNH 2181, YPM 1499, YPM 1200, YPM PU 17193) (
Figure 60).
The fibula of Parahesperornis is crutch-shaped, with the proximal surface expanded on the cranial side. The shaft is roughly oval in cross section and tapers along its length to a thin sliver at the distal-most end, very similar to the fibula of other hesperornithiforms. The proximal surface forms a deep depression, which resembles the seat of a saddle in medial and lateral view. This is intermediate between Hesperornis and other hesperornithiforms, which is less deeply excavated, and Baptornis, which is more deeply excavated. The medial face of the fibula of Parahesperornis has a triangular depression, with a small foramen centered on the proximal margin. A similar depression is only faintly developed in Hesperornis and Fumicollis, while in Baptornis no depression is evident. In lateral view the face of the proximal end bulges outward on the cranial side, as in other hesperornithiforms. A large muscle scar is visible on the cranioproximal end of the fibula, as in Hesperornis, but which is not visible in other hesperornithiforms.
The shaft of the fibula of Parahesperornis is irregularly shaped. The tubercle for the m. iliofibularis is present almost one-fourth of the way down the shaft on the caudal margin (best seen in medial view). Distal to this tubercle, just below the midpoint of the shaft, the bone narrows dramatically, with its sides forming a sharp angle in medial and lateral view, as in Hesperornis and Fumicollis. A crest is present along the medial face of the shaft where the fibula abuts the tibiotarsal shaft. This crest is also present in Hesperornis but does not extend as far distally. This crest may serve as the attachment of the ligaments that connect the fibula to the tibiotarsus.
Tibiotarsus. Both tibiotarsi of
Parahesperornis are preserved with KUVP 2287 (
Figure 61) and KUVP 24090. In KUVP 24090the right is incomplete (
Figure 62) while the left is complete and encased in chalk in caudal view (
Figure 43 and
Figure 44). For comparison, the following specimens were used:
Baptornis (AMNH 5101, KUVP 2290, FMNH 395);
Brodavis varneri (SDSM 68430);
Enaliornis (SMC B55315, SMC B55314);
Fumicollis (UNSM 20030);
Hesperornis (AMNH 2181, YPM 1200, YPM 1476); and
Pasquiaornis (RSM P2957.21, RSM P2957.22) (
Figure 63). The tibiotarsus of
Parahesperornis has several typical hesperornithiform features, in particular an expanded proximal process for the cnemial crests, a gap in the fibular crest, and a medially deflected medial condyle.
The proximal portion of the tibiotarsus of Parahesperornis is very similar to that of Hesperornis. In proximal view, the medial cotyla is large and kidney shaped, separated from the cnemial expansion on the cranial margin by a groove. In medial view, the articular surface of the medial cotyla slopes caudally, with the margin of the cotyla thickened and rugose. The lateral cotyla is circular, much smaller than the medial, and displaced caudally, such that it has a similar caudal extent as that of the much larger medial cotyla. While the cotylae are similarly unevenly sized in Hesperornis, they are not levelled caudally, and the lateral cotyla is elongate and oval shaped. In Fumicollis the size discrepancy between the cotylae is not as exaggerated as in hesperornithids, a reflection of the smaller discrepancy between femoral condyles, as is also seen in other hesperornithiforms. Cranial to the lateral cotyla is a deep indentation between this cotyla and the cnemial expansion, for articulation with the proximal end of the fibula.
In cranial view, the cnemial expansion is triangular, with the medial corner higher proximally than the lateral. While the cnemial expansion of KUVP 24090 appears to be narrower than that of KUVP 2287, this is due to the crushing of the proximal end of KUVP 24090. The lateral cnemial crest is sharp and narrow, running along the lateral margin of the cnemial expansion. The lateral cnemial crest extends distally approximately one-fifth the length of the shaft, while the medial cnemial crest extends to about midshaft. On the cnemial expansion, the medial cnemial crest is broader than the lateral. The cnemial expansion of Hesperornis is more oval in cranial view, while that of Fumicollis and Brodavis varneri is also triangular. Across hesperornithiforms, the cnemial crests are equally well-developed, varying only in their shape. While the cnemial crests of Enaliornis and Brodavis varneri are rounded proximally, that is likely due to the poor preservation of the specimens, which are all heavily eroded.
The proximal shaft of the tibiotarsus of Parahesperornis narrows very slightly below the medial cotyla, which forms a slight lip at its caudal-most extent. In caudal view, the articular surface is angled, with the medial side highest, and expanded mediolaterally past the margins of the shaft, as in Hesperornis. In Fumicollis the articular surface is also angled, but not as greatly expanded mediolaterally, while in Baptornis and Brodavis the articular surface is only slightly angled and expanded. A slight depression is present on the proximal face of the caudal shaft just below the overhanging articular surfaces. This depression is deeper in Hesperornis and faint in Fumicollis and Baptornis.
The tibiotarsal shaft of Parahesperornis is straight in caudal view and oval in cross-section. In both cranial and caudal views, the shaft of the tibiotarsus is narrowest proximally and very gradually expands distally, while in medial and lateral view the shaft is a constant width until just above the distal condyles, where it then narrows. The shaft is slightly twisted, such that when the distal end is in cranial view the proximal end is in craniomedial view. This is also seen in Hesperornis and Fumicollis. The shaft is slightly bowed along its length, such that in medial or lateral view the shaft is cranially convex just below midshaft. This bowing is more pronounced in KUVP 24090 than in KUVP 2287. In KUVP 24090 the shaft appears to be sinusoidal in cranial and caudal view, while that of KUVP 2287 is straight. The poor preservation of many of the tibiotarsi assigned to Hesperornis make observation of the straightness of the shaft difficult, as some specimens appear to be perfectly straight (e.g., YPM 1200), while others more closely resemble KUVP 24090 with a prominent bow and sinusoidal shape (e.g., YPM 1476). The tibiotarsus of Fumicollis is slightly bowed and sinusoidal.
In Parahesperornis, the fibular crest extends from the neck of the lateral cotyla distally to just below midshaft, as in Hesperornis. In Fumicollis and Baptornis the fibular crest ends at midshaft. At this point there is a gap in the fibular crest before the proximal end of the less prominent distal fibular crest that originates just above the lateral condyle, as is also the case for Hesperornis and Fumicollis. Baptornis lacks this distal part of the fibular crest. In cranial view, the fibular crest has minimal lateral projection, being about the same as that of the medial cnemial crest, however in caudal view the fibular crest is much wider, with a faint groove running along the medial margin separating it from the shaft of the tibiotarsus. This groove is more prominent in KUVP 24090 than KUVP 2287, where the fibular crest also appears to be less laterally projected than in KUVP 2287.
The distal end of the tibiotarsus expands slightly on the lateral side but flares widely on the medial side, such that the center of the medial condyle is in line with the medial margin of the shaft. This is like the condition in Fumicollis and Baptornis, and more exaggerated than in Hesperornis. In other hesperornithiforms the distal end is centered on the shaft and evenly expanded mediolaterally. In cranial view, the distal shaft has a deep central extensor groove that ends in a slight pocket behind the intercondylar bridge. The cranial face of the lateral shaft is broad and flat, extending over the extensor groove. A slight depression is present on the shaft just proximal to the lateral condyle, as in other hesperornithiforms. A large tuberosity is present on the medial side of the cranial shaft, extending into the extensor groove. The tuberosity is also seen in Hesperornis but not in other hesperornithiforms. No true supratendinal bridge is present in hesperornithiforms, however a ridge is present between the condyles that spans the extensor groove distally.
The medial and lateral condyles of
Parahesperornis are of a similar size in cranial view, with a deep intercondylar groove between them and the medial condyle extending slightly further distally than the lateral, as in
Hesperornis. In other hesperornithiforms the medial condyle extends past the lateral to a greater degree. In medial view, the medial condyle is much smaller than the lateral, such that the lateral condyle can be seen on either side of the medial. The caudal margin of the medial condyle forms a lip around the medial face of the condyle, which has a prominent tubercle for the attachment of the medial collateral intertarsal ligament [
86] on its face. The entire condyle appears to be angled caudally in medial view, and overall appears very much like the medial condyle of
Hesperornis. In
Fumicollis and
Baptornis, the medial condyle is in line with the shaft. However, it is not as reduced as in hesperornithids and expanded cranially.
In caudal view, both condyles form narrow epicondylar crests that wrap up onto the cranial surface of the shaft. In lateral view, the lateral condyle is somewhat rounded, being nearly circular along the caudal margin but flattened along the cranial margin. The cranial margin of the lateral condyle projects past the shaft. In contrast, the lateral condyle of Hesperornis is nearly circular in lateral view and centered on the distal shaft. In Fumicollis and Baptornis, the lateral condyle is further projected cranially than in Parahesperornis. The center of the lateral face of the condyle has a round tuberosity with a deep depression on its craniodistal border.
In distal view the condyles are aligned mediolaterally, with the lateral condyle much wider craniocaudally than the medial. This is also the case in Hesperornis, however in Fumicollis the condyles are more similarly sized. The craniocaudal axes of the condyles angle toward each other cranially. This feature may be highly subject to taphonomic distortion, as in some specimens of Hesperornis the orientation of the craniocaudal axes of the distal condyles varies dramatically between left and right elements of an individual (e.g., YPM 1476).
Tarsometatarsus. The left and right tarsometatarsi of
Parahesperornis are preserved with KUVP 2287 (
Figure 64). KUVP 24090 does not preserve either tarsometatarsus. An additional tarsometatarsus, FHSM VP-17312, has previously been assigned to the genus
Parahesperornis [
26]. The following specimens were used for comparisons:
Baptornis (AMNH 5101, FMNH 395, YPM 1465);
Brodavis baileyi (UNSM 50665);
Brodavis varneri (SDSM 68430);
Fumicollis (UNSM 20030);
Hesperornis regalis (YPM 1200, YPM 1207, YPM 1476);
Hesperornis gracilis (YPM 1473, YPM 1478); and
Pasquiaornis (RSM P2997.18, RSM P2997.81) (
Figure 65). The tarsometatarsus of
Parahesperornis exhibits the typical hesperornithiform features of a prominent intercotylar eminence, dorsal surface of the fourth metatarsal forming a sharp ridge, expanded trochlea on the fourth metatarsal, reduced and proximoplantarly displaced second metatarsal trochlea, and shingled or stacked arrangement of the metatarsals.
In proximal view, the tarsometatarsus of Parahesperornis is somewhat rectangular in outline, with a longer mediolateral width than craniocaudal depth, but with the medial and lateral margins angled such that the outline is rhombic overall. The proximal articular surfaces of both Hesperornis and Fumicollis are more rectangular, with a greater discrepancy between the craniocaudal and mediolateral dimensions, while that of Baptornis is more like Parahesperornis. Both cotylae are oval and craniocaudally expanded, with the medial slightly longer than the lateral, as in most other hesperornithiforms except for Baptornis, where the lateral cotyla is longer than the medial. The plantar margins of both cotylae are smoothly rounded and lack the flange that is prominent in Hesperornis and less developed in Fumicollis and Baptornis.
The intercotylar eminence is broad and rounded, as in all other hesperornithiforms except Hesperornis, where it is narrow and peaked. The intercotylar eminence is centrally located along the dorsal margin of the articular surface, as in other hesperornithiforms. In dorsal view, the intercotylar eminence projects proximally and is slightly angled laterally with a small depression on the medial margin, as in most other hesperornithiforms except Baptornis, where it is not angled. On the plantar slope of the intercotylar eminence, there is a slight depression that is found in all hesperornithiforms. In medial and lateral views, the intercotylar eminence of the isolated Parahesperornis tarsometatarsus FHSM VP-17312 is narrower and more pointed than in KUVP 2287.
In plantar view, the proximal articular surface of the tarsometatarsus of Parahesperornis is angled, with the medial side highest. This is typical in hesperornithiforms, to a similar degree in Fumicollis and Baptornis and to a greater degree in Hesperornis. In medial view, the proximal end of metatarsal II of Parahesperornis has a flattened surface that angles dorsoplantarly, with a rounded plantar margin and a sharper dorsal margin. Below this the shaft of metatarsal II narrows at a slight neck, less pronounced than that seen in Hesperornis and more like that of Fumicollis.
The plantar margin of the articular surface forms the proximal extent of a rudimentary hypotarsus. Most hesperornithiforms have a triangular, roughened patch of bone that covers the proximal plantar surface of the tarsometatarsus and wraps up onto the dorsal edge of the articular surface. In Parahesperornis this surface is delineated by grooves on the proximal articular surface, as well as on the medial and lateral margins of the plantar surface, as in Hesperornis. In other hesperornithiforms these edges are not well marked. The hypotarsus is more elongate in Parahesperornis than in Hesperornis. This area is only faintly developed in Fumicollis and Baptornis. In Brodavis varneri the hypotarsal area is not preserved, however two very deep grooves or slits are preserved that coincide with the margins of metatarsal III.
On the dorsal surface of the shaft, the proximal portions of metatarsals II and IV are developed as ridges, with metatarsal III deeply depressed between them to form the extensor groove, as in other hesperornithiforms. Within the proximal-most part of this depression are two oval foramina, the medial of which is slightly proximal to the lateral. These foramina do not penetrate to the plantar surface of the tarsometatarsus, as in modern birds, and they are found in all hesperornithiforms. In Parahesperornis, Hesperornis, Fumicollis, and Brodavis varneri, the proximal foramina are fairly large and either oval or teardrop shaped, while in Baptornis they are much smaller and circular.
Distal to the proximal foramina is an elongate depression or small groove with a tuberosity along the distal margin. This is similar to the condition in Fumicollis, while in Hesperornis the depression is reduced or absent and in Baptornis the tuberosity is reduced. This tuberosity is interpreted as the attachment of the m. tibialis cranialis. In lateral view the margin of metatarsal IV is evenly rounded with a depression on the proximolateral face, as in Fumicollis. In Hesperornis the corresponding area is flattened and tilted dorsally.
The shaft of the tarsometatarsus of Parahesperornis is straight and narrows gradually along its length in dorsal view. Also in this view, the metatarsals are separated by grooves along their length, with the dorsal surface of metatarsal IV forming a high, rounded ridge, as in Hesperornis. Distally, these grooves become much more deeply excavated, while in Hesperornis they are deeply excavated along the length of the shaft. The grooves extend the full length of the shaft but are less prominent in Fumicollis, while in Baptornis and Brodavis they only extend to the midshaft.
The shaft of the tarsometatarsus of Parahesperornis is twisted, such that when the proximal end is in dorsal view the distal end is in dorsomedial view. While all hesperornithiforms share this trait to some degree, the degree of rotation in Parahesperornis is most like that of Fumicollis, as Hesperornis is more twisted and Baptornis and Brodavis are less twisted. In plantar view the shaft of Parahesperornis forms a continuous flattened surface with a slight central depression bordered on the medial and lateral edges by thin ridges, as in Hesperornis and Fumicollis, while the depression is very faint in Baptornis. In medial view the shaft of metatarsal II is slightly sinusoidal in shape, curving proximally and distally, as in Fumicollis. Hesperornis has a similar, but less dramatic, shape, while that of Brodavis is more dramatically curved. In Baptornis the shaft of metatarsal II is bowed in medial view.
Distally, the tarsometatarsus of Parahesperornis is like those of Hesperornis and Fumicollis. Between the necks of trochlea III and IV, the deep groove between the metatarsals terminates in a distal foramen that penetrates through to the plantar surface. In plantar view, the distal foramen is oval and set within a larger tear-drop shaped hollow between the metatarsals, as in Hesperornis and perhaps Fumicollis, although in UNSM 20030 the area is obscured partially by sediment. While the distal foramen is seen in all hesperornithiforms, the spacing of the metatarsal trochlea distal to the foramen varies. In Parahesperornis, like in Hesperornis and Fumicollis, the trochleae are pressed tightly together, while in other hesperornithiforms the trochleae are slightly further apart, including the isolated tarsometatarsus assigned to Parahesperornis (FHSM VP-17312).
The metatarsal trochleae of hesperornithiforms are offset dorsoplantarly to varying degrees, giving the metatarsals a shingled or stacked appearance in medial view, with trochlea II displaced plantarly such that it is almost completely behind metatarsal III. The most extreme form of this morphology is seen in Hesperornis. In Parahesperornis and other hesperornithiforms trochlea II is less displaced plantarly and is more set on the medial side of metatarsal III. In Parahesperornis trochlea IV is larger than II or III, being slightly wider dorsoplantarly (best seen in medial view) and almost twice as wide mediolaterally (best seen in distal view). Trochlea IV also extends slightly further distally than III. This difference is seen to varying degrees in other hesperornithiforms. In Hesperornis the size discrepancy is greater with trochlea IV extending much further distally, while in Pasquiaornis it is less than in Parahesperornis. Fumicollis is the most like Parahesperornis in terms of the size discrepancy and distal extent of trochlea III and IV.
Trochlea II appears somewhat triangular in dorsal view, with the medial trochlear ridge displaced proximally, as in other hesperornithiforms. In medial view, the plantar margin of the medial face of the trochlea extends into a small flange, like that seen in
Hesperornis. In
Baptornis this flange is somewhat better developed and slightly medially directed, while in
Fumicollis the margin is evenly rounded without a flange. The medial face of trochlea III and the lateral face of trochlea IV are marked with a circular depression for the attachment of the collateral ligaments [
86]. In distal view, the size discrepancy in the medial and lateral trochlear ridges of trochlea IV is easily seen, with the medial ridge about 70% the length of the lateral. This is very similar to the condition in
Hesperornis and much more exaggerated than in other hesperornithiforms. The plantar-most margin of the lateral trochlear ridge flips laterally, to a lesser degree than in
Hesperornis, giving the trochlea a slight c-shape on the lateral margin. Trochlea III is triangular in shape in distal view with similarly sized medial and lateral trochlear ridges, as in other hesperornithiforms.
Phalanges. Nearly all the phalanges of
Parahesperornis are preserved in KUVP 2287: complete digits III and IV, including the ungual phalanges, from both the left and right feet; the first phalanx and ungual phalanges are preserved from both the left and right side of digit II, while the second phalanx is only preserved from the left side and the third is only preserved from the right, and from digit I the terminal phalanges are preserved from the left and right side while only the right ungual phalanx is preserved (
Figure 66). These phalanges are very similar in form to those of
Hesperornis. For comparison the following specimens were used:
Baptornis (FMNH 395);
Fumicollis (UNSM 20030); and
Hesperornis (FHSM VP- 2069, YPM 1200, YPM 1476) (
Figure 67).
The phalanges of digit IV of Parahesperornis are overall very similar to those of Hesperornis and reflect the unique morphology of the enlarged metatarsal IV trochlea and the enlarged medial ridge of that trochlea in hesperornithids. Unlike Fumicollis and Baptornis, the phalanges of digit IV of hesperornithids are robust and cylindrical, with a triangular cross-section distally and a circular cross section proximally. The proximal articular surface has an enlarged, oval medial cotyla and a much smaller, circular lateral cotyla. At the plantar end of the medial cotyla the articular surface curves proximally into a high peak. This peak is the proximal-most extent of a dramatic flange that extends plantarly from the medial side of the phalanx. This flange is also present in Hesperornis, where it is proportionally larger. The plantar surface of digit IV has a flattened dorsal surface with a depression along its length. The medial and lateral sides of digit IV phalanges are smoothly rounded. Distally, the body of the phalanx narrows, with a slight neck just before the distal trochlea. The medial and lateral trochlea are unevenly sized, with the lateral a small peg projected much further distally than the broader, crescent-shaped medial condyle, as in Hesperornis.
This morphology is extremely different from that of Baptornis. The phalanges of the fourth toe of Baptornis are much more elongate with a circular cross section. Proximally, the articular surface is not divided into separate cotylae for the trochlear ridges of metatarsal IV, and the distal end is only weakly divided into trochleae. The plantar face is slightly flattened, however the groove along its surface is lacking.
The first toe of digit IV is longer than the second and third, which are similarly sized, while the fourth toe is longest and less robust than the preceding phalanges. The ungual phalanx, or claw, is small and triangular, with a hooked appearance in dorsal or plantar view not seen in the other unguals. In Hesperornis the phalanges of the fourth toe have a similar morphology, however the fourth phalanx is much thinner in Parahesperornis, being almost flattened dorsoplantarly. It is possible this is due to preservational factors. The ungual phalanx for the fourth toe is most likely not preserved in any of the Hesperornis specimens accessible for this study, as none display the prominent hook shape seen in Parahesperornis.
The size discrepancy between the articular surfaces of metatarsal trochlea IV and the phalanges of digit IV has implications for the movement of the toes. As digit IV is bent, at each joint the proximal bone will swivel around the smaller medial condyle on the longer, crescent-shaped lateral condyle. This, combined with the stacked orientation of the metatarsals within the tarsometatarsus, functions to streamline the foot and increase the range of motion of the longest toe. This has led to the conclusion that hesperornithiforms may have had lobed feet, as in modern grebes [
19]. This conclusion is reasonable, as grebes share the size discrepancy in the articular surfaces of the toes and generate lift during the swimming stroke by separating and rotating their toes, thus using each lobe as a separate hydrofoil [
87]. However, it should be noted that coots also have lobed toes but do not have unevenly sized condyles, and only hesperornithids show this size discrepancy in the condyles of the fourth toe.
The phalanges of digit III of Parahesperornis are very different from those of digit IV, reflecting the much narrower third metatarsal trochlea. In Parahesperornis these phalanges are very narrow mediolaterally and triangular in medial or lateral view, with a greatly expanded proximal end and a long neck, as in Hesperornis. The phalanges of digit III of Fumicollis are similarly shaped but more elongate. In Baptornis the proximal end is not as greatly expanded, and these phalanges are not as compressed mediolaterally. In Parahesperornis, a deep, triangular depression is present on the proximal end of the plantar surface, as in other hesperornithiforms. The proximal articular surface is oval shaped in proximal view, with the plantar end somewhat flattened and the dorsal end coming to a peak, with the surface deeply depressed, as in Hesperornis and Fumicollis. In Baptornis, the proximal end is more circular.
The distal ends of the digit III phalanges in Parahesperornis are divided into two similarly sized trochlea, the medial of which extends very slightly further distally than the lateral, as in Hesperornis. In Baptornis the trochleae are less clearly defined and have an equal distal extent. In distal view, the dorsoplantar axes of the trochleae intersect dorsally. The phalanges of digit III of Parahesperornis get progressively shorter away from the foot, as in other hesperornithiforms. The ungual phalanx, or claw, is small and triangular, flattened dorsoplantarly, and with a bulbous proximal end, as in Hesperornis.
The phalanges of digit II of Parahesperornis are highly elongate and fairly consistent along their length, widening slightly at the distal end in medial or lateral view and narrowing to a very slight neck just above the trochlea. A very faint groove runs along the dorsal surface. These phalanges are somewhat flattened dorsoplantarly, particularly the second phalanx. The proximal articular surface of the first phalanx is triangular and faintly divided into two cotylae for articulation with the uneven second trochlea of the tarsometatarsus. The distal end is divided into two subequal trochlea. The proximal surface of the second phalanx is narrower and oval in outline, with only faint margins of the cotylae. The distal trochleae of the second phalanx are only weakly developed. The ungual phalanx is much like that of the third toe, such that they seem interchangeable. The morphology of these phalanges is consistent with that of Hesperornis and Fumicollis.
The first phalanx of digit I of Parahesperornis is small and rectangular in dorsal or plantar view. It is highly flattened dorsoplantarly and slightly arched in medial or lateral view. Both the proximal and distal ends are weakly developed, with little development of the articular surfaces. The morphology of the first phalanx is very similar to that of Fumicollis. The ungual phalanx of the first toe is triangular and flattened dorsoplantarly and is slightly more elongate than the other unguals.







































































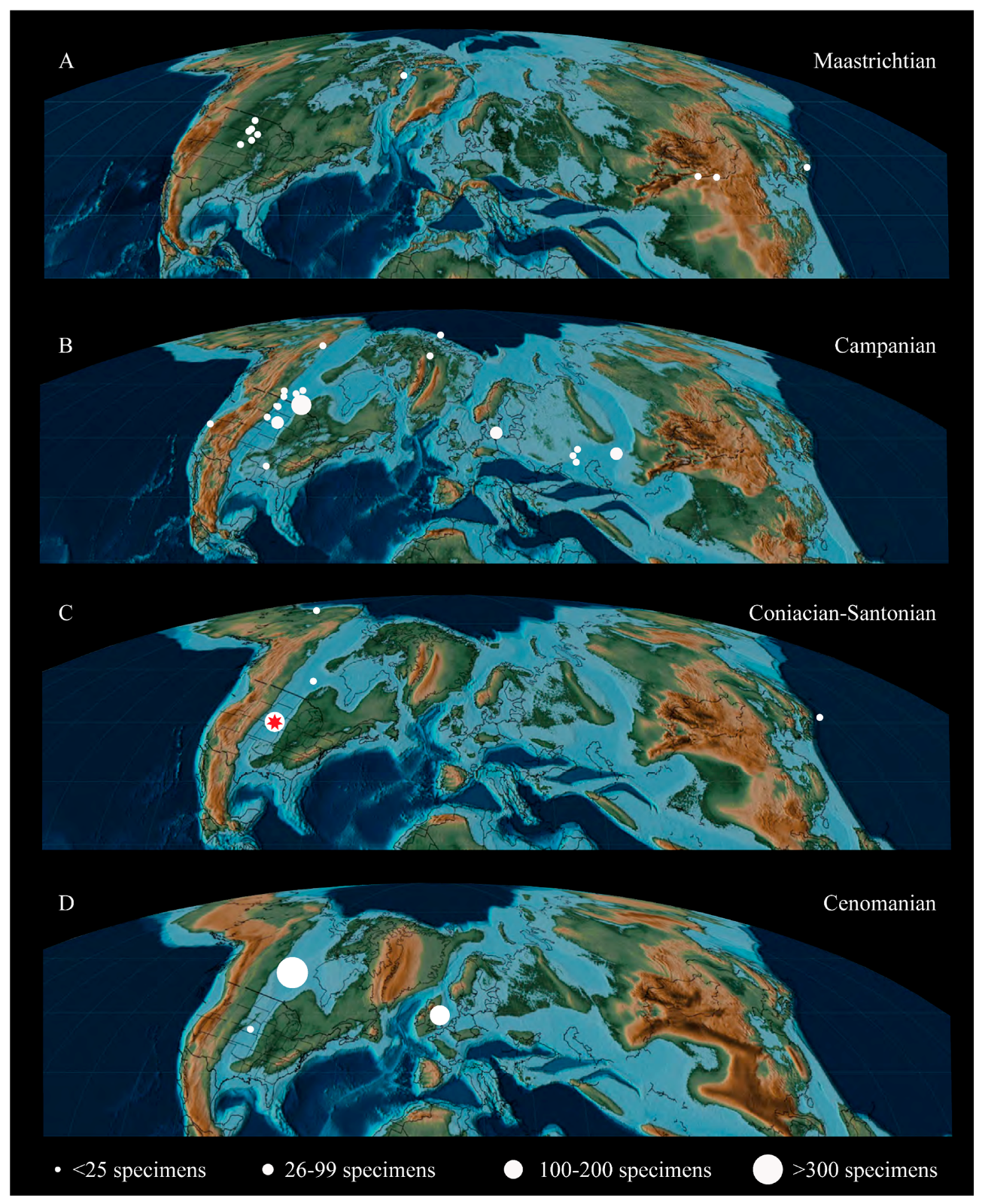{kind=link}
{kind=link}
{kind=link}
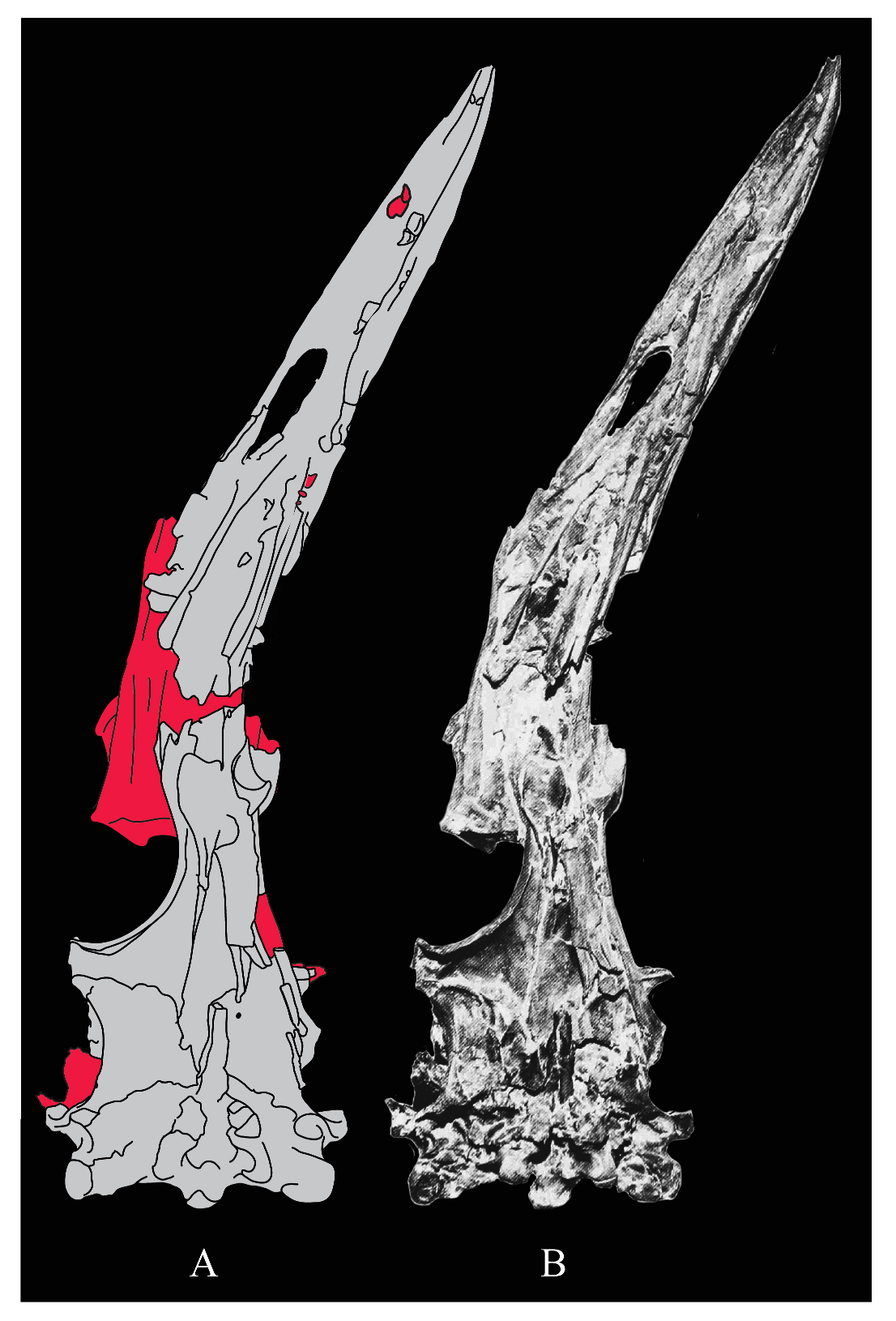{kind=link}
{kind=link}
{kind=link}
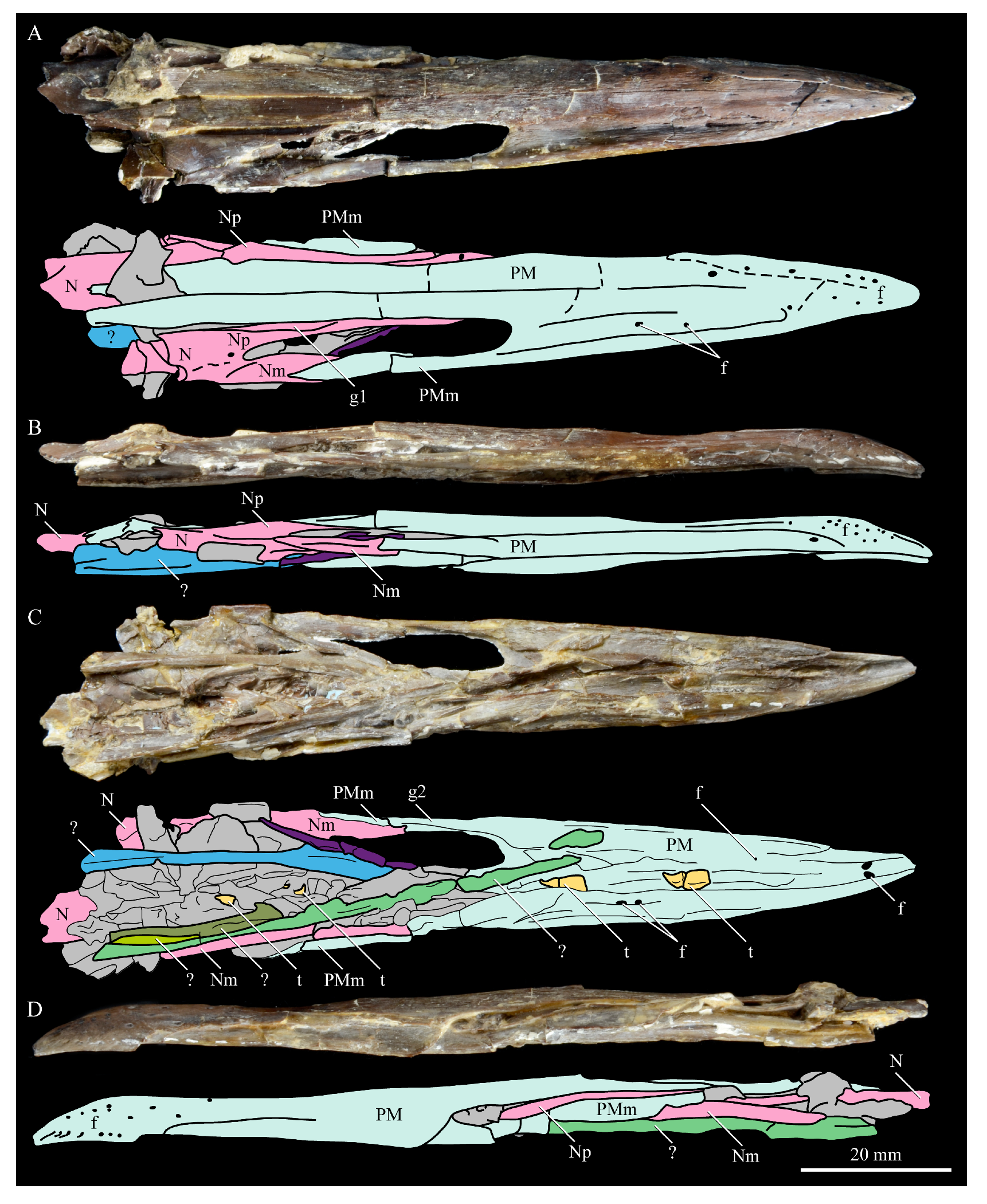{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
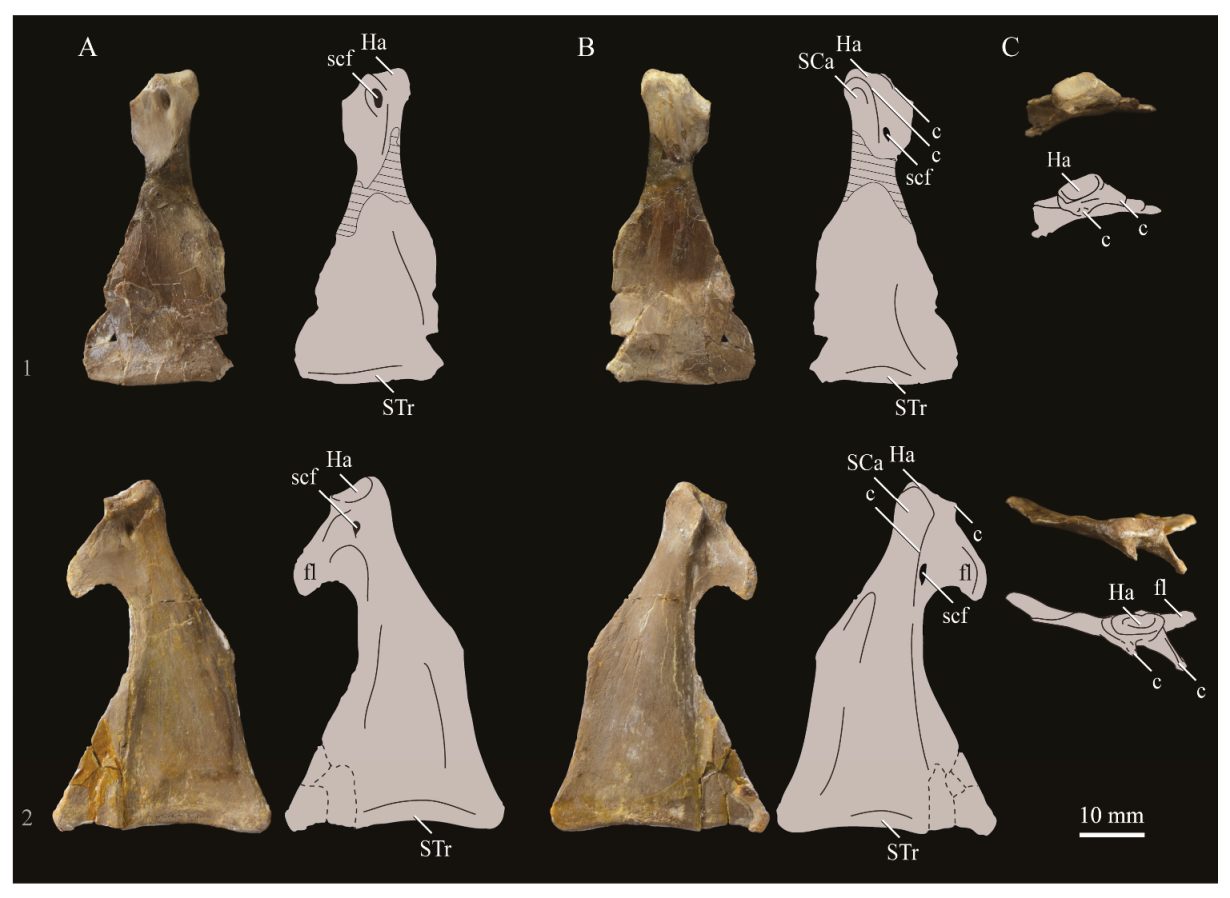{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
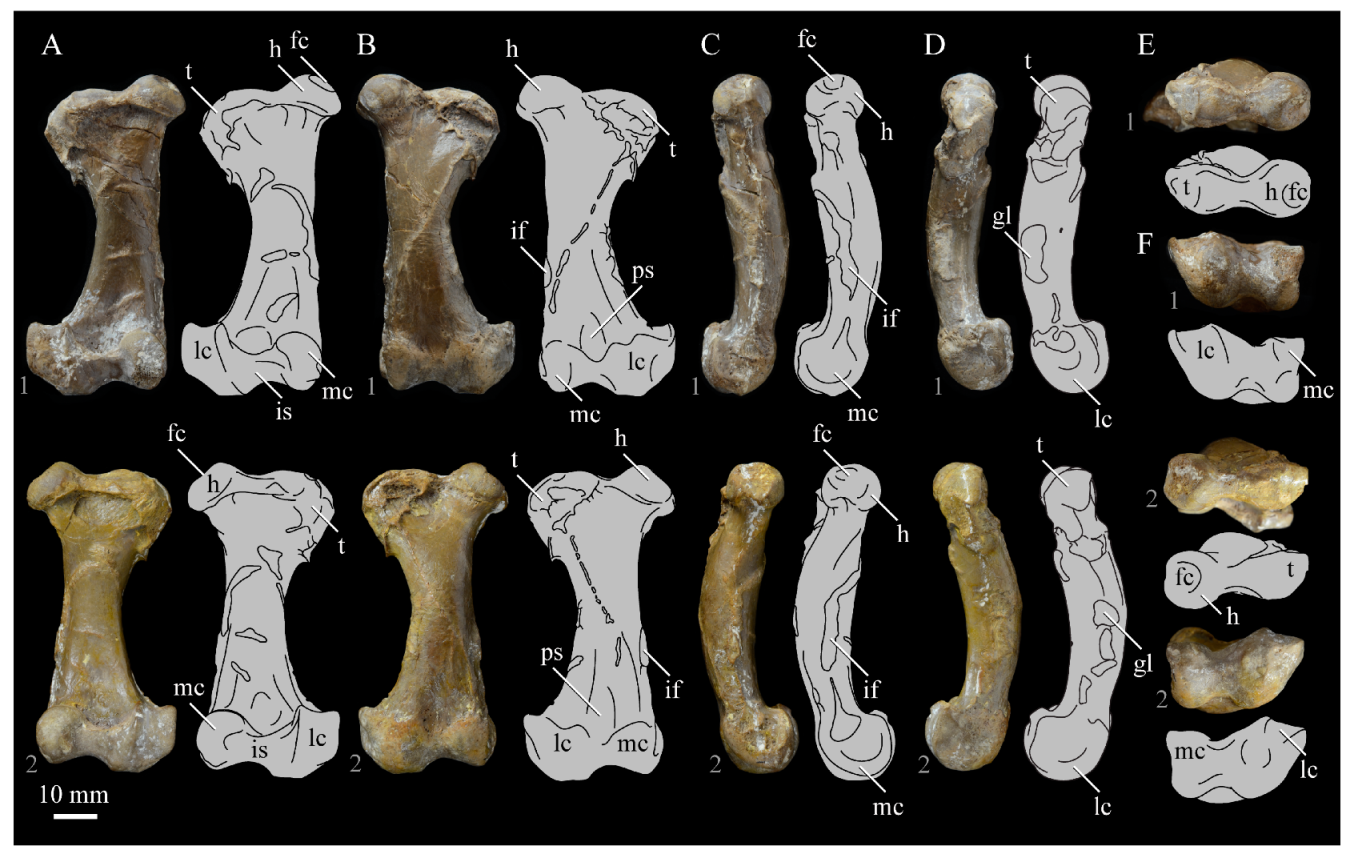{kind=link}
{kind=link}
{kind=link}
{kind=link}
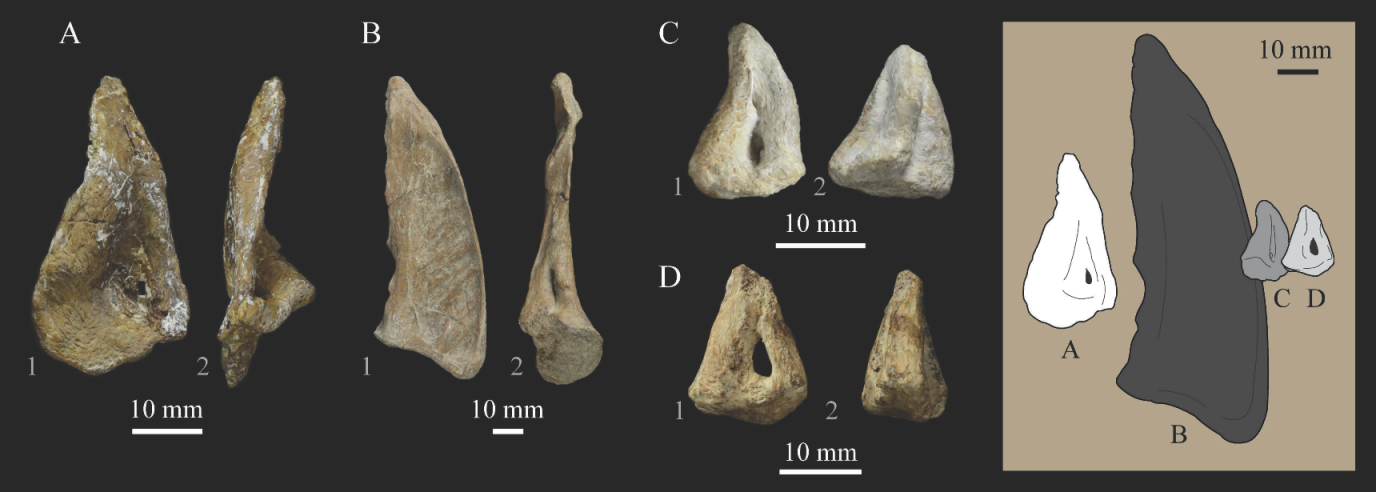{kind=link}
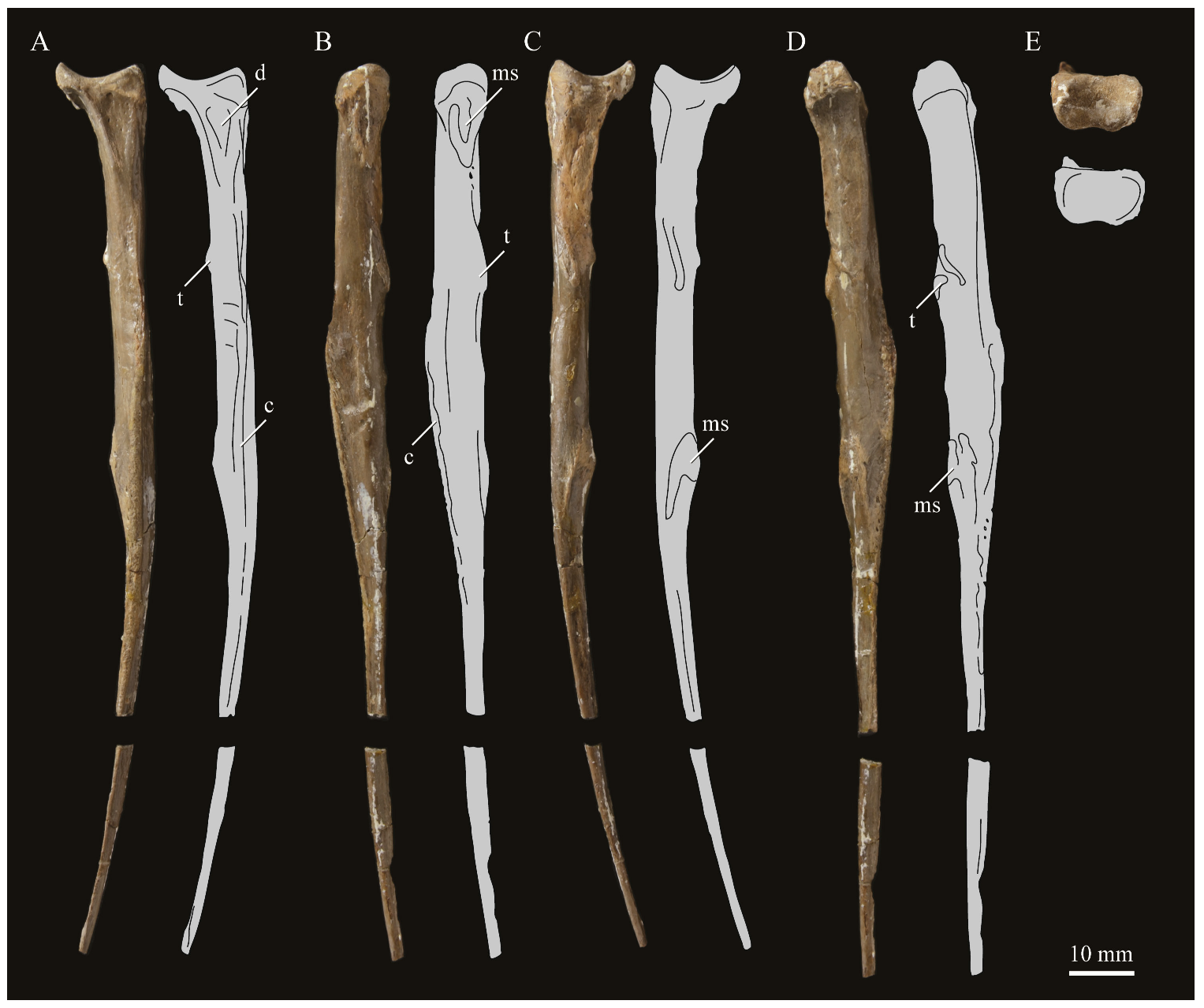{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
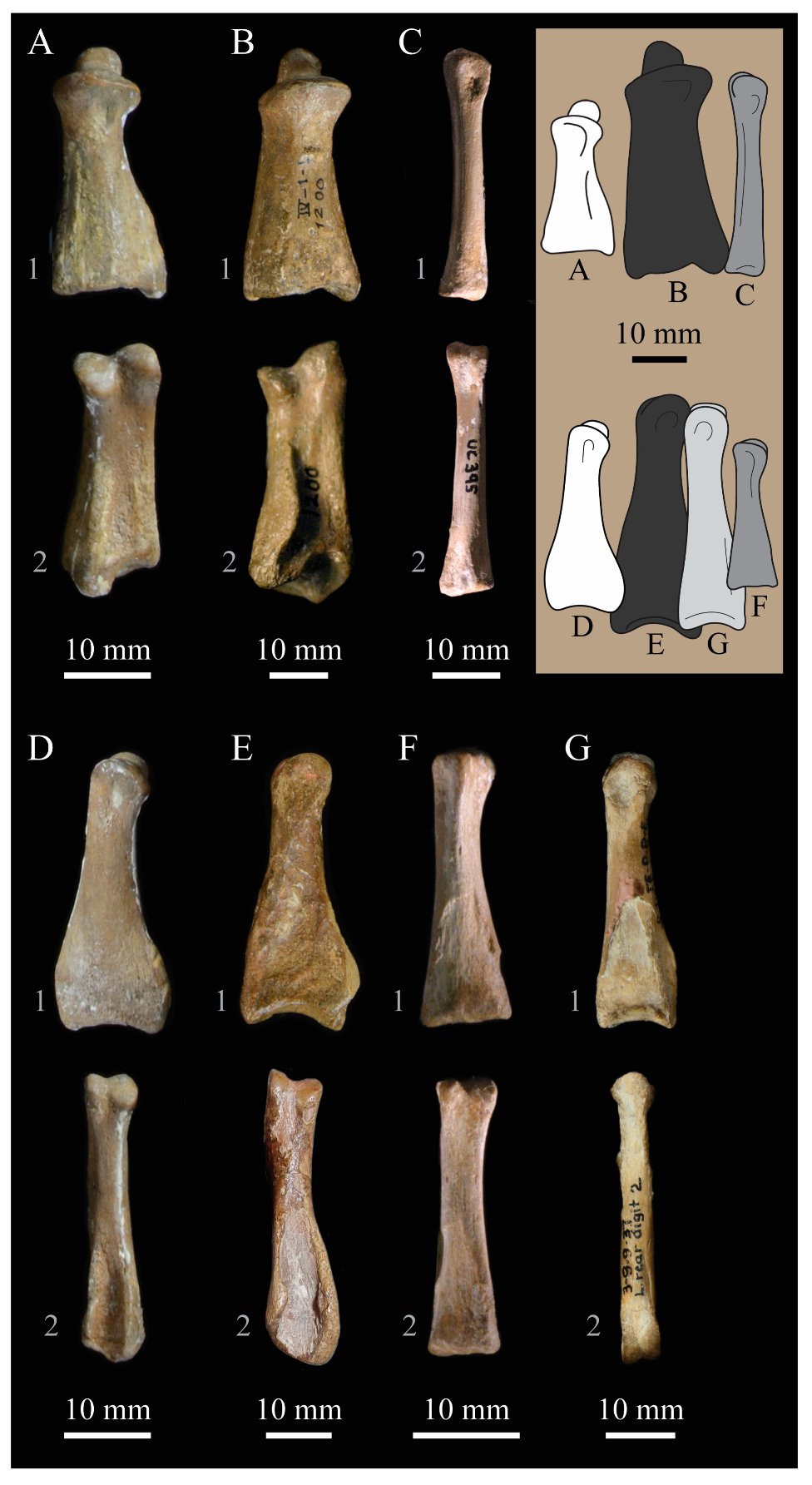{kind=link}
{kind=link}
{kind=link}
{kind=link}