

Dynamic Analysis of a Delayed Differential Equation for Ips subelongatus Motschulsky-Larix spp.


Abstract
1. Introduction
2. Mathematical Modeling
- (a)
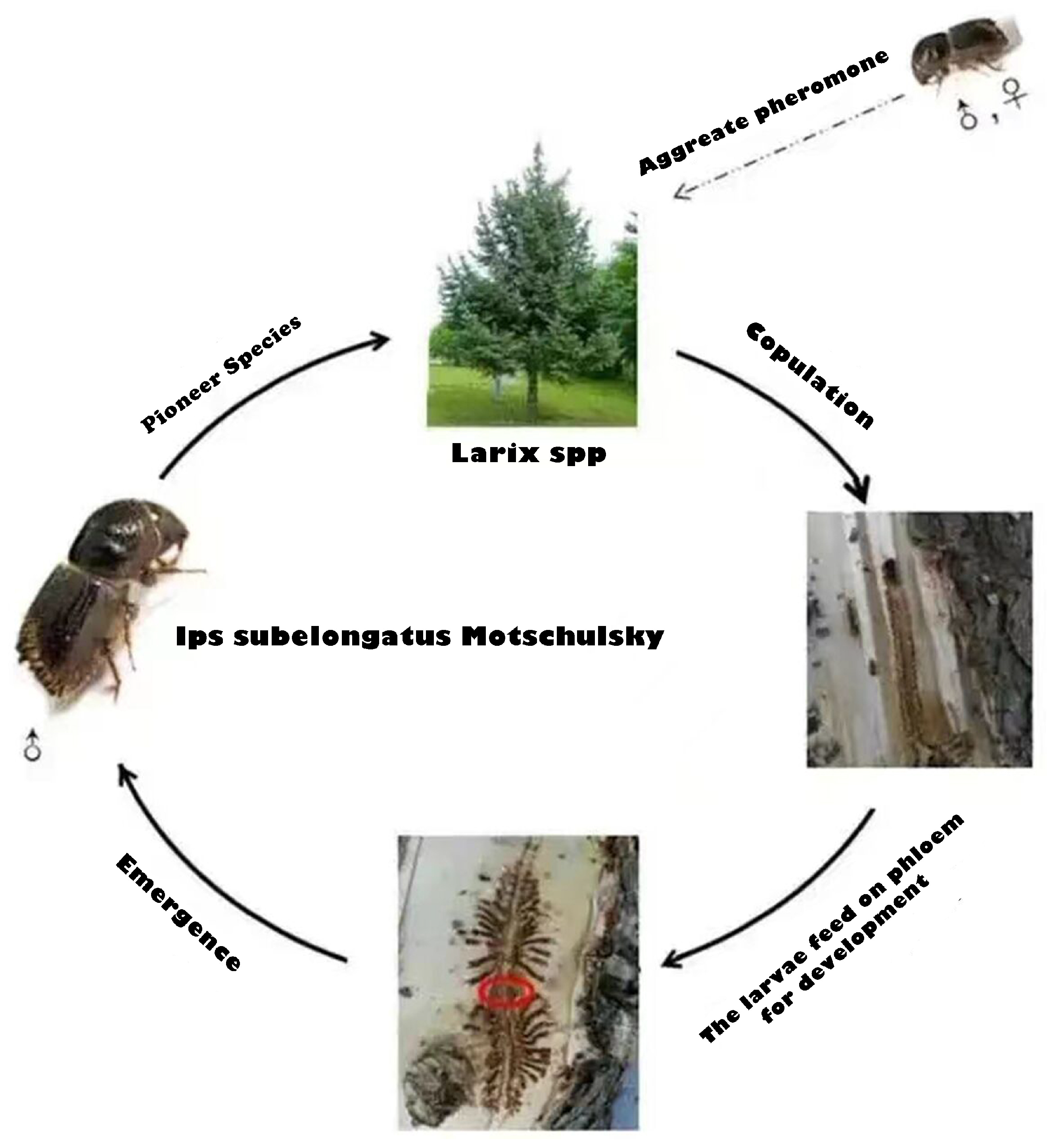
- Based on the biological characteristics of Ips subelongatus Motschulsky, we denote as the population size of Ips subelongatus Motschulsky. The reproductive capacity of pests is subject to resource constraints, resulting in a gradual decrease in their growth rate as they approach the environmental capacity N, we assume that the growth rate of pests follows the logistic model; that is, . Moreover, the mortality rate pests experience not only depends on their own mortality rate , but also depends on predation by natural predators M. Based on this analysis, we establish the first equation of system (1) to describe the rate of change in the pest population.
- (b)
- We assume that susceptible Larix spp. becomes infected Larix spp. after being infected by pests. Infected Larix spp. can be cured and transformed back into susceptible Larix spp. . The process of curing requires a certain amount of time, taking into account practical considerations; thus, we introduce as the curing duration. The rate at which cured infected trees change is denoted as .
- (c)
- In the conventional infectious disease model, the infection rate is constant, but the infection ability of Ips subelongatus Motschulsky is stronger when the canopy density of the trees is lower, indicating that pests have a stronger ability to infect as more trees die. Therefore, we establish an infection rate function , where k represents the infection rate coefficient and represents the rate of change in infected trees that have died. This function reflects that the infection rate is directly proportional to . At this point, the infectiousness of the pest is denoted . Thus, the transmission rate from susceptible trees to infected trees can be denoted . Since the forest is not isolated, new Larix spp. trees will be planted at a certain rate R. Based on the above analysis, we can establish a relationship between susceptible and infected trees.
3. Stability Analysis of Equilibrium and Existence of Hopf Bifurcation
3.1. The Existence of Equilibrium
3.2. The Stability of Equilibrium and Existence of Hopf Bifurcation
4. Normal Form of Hopf Bifurcation
- (i)
- If , the periodic solution reduced on the center manifold is unstable, when , the Hopf bifurcating periodic solution is forward (backward).
- (ii)
- If , the periodic solution reduced on the center manifold is stable, when , the Hopf bifurcating periodic solution is forward (backward).
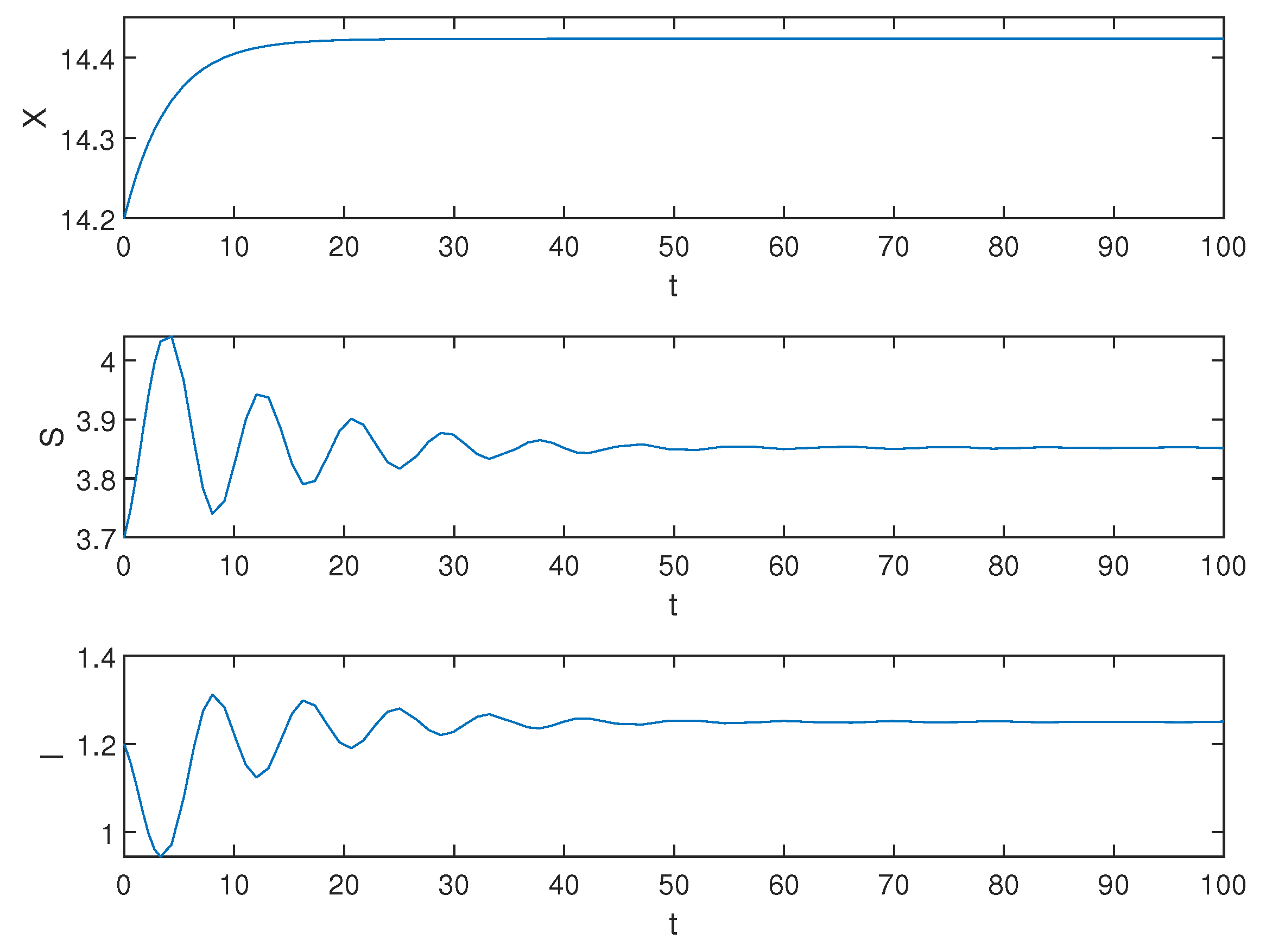
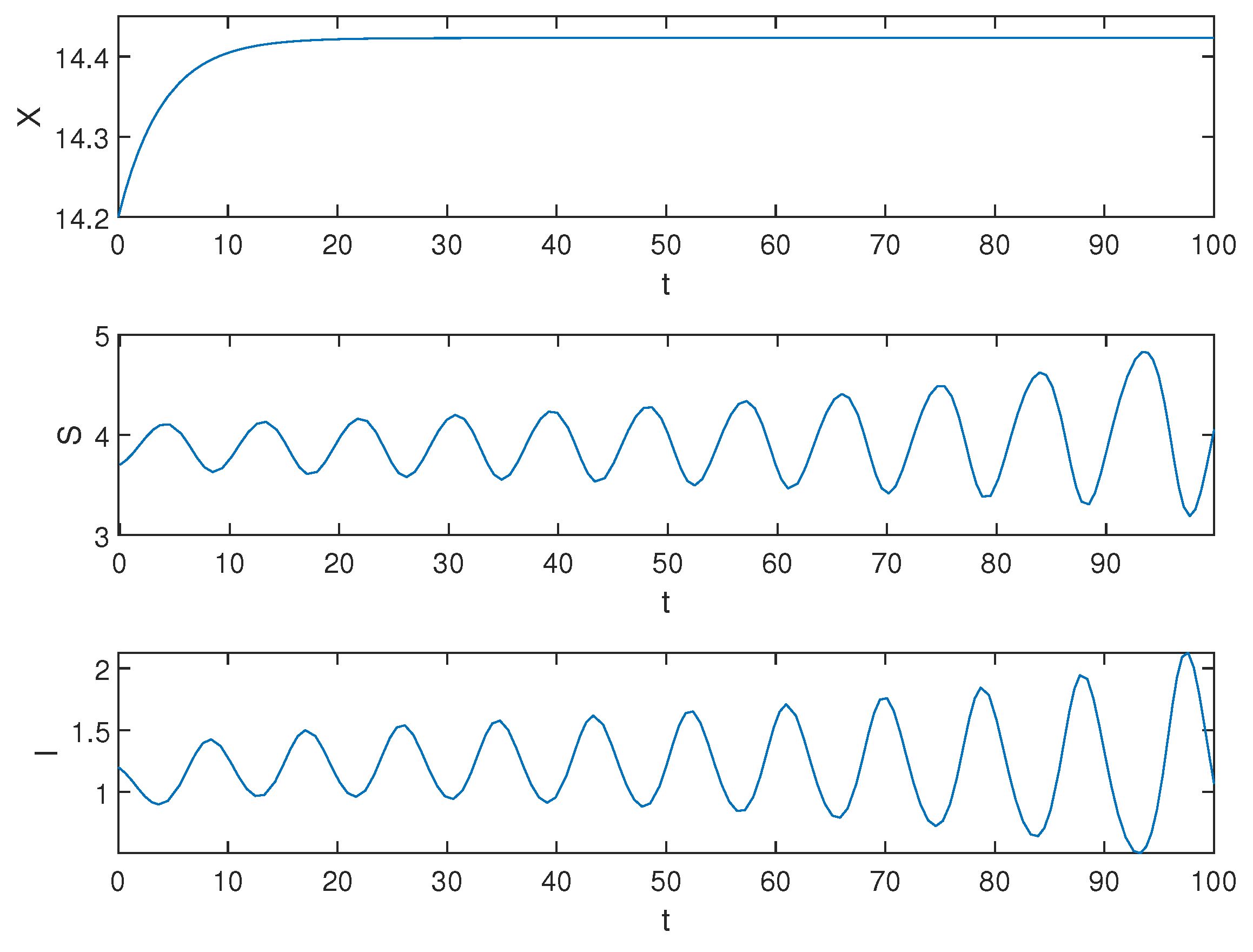
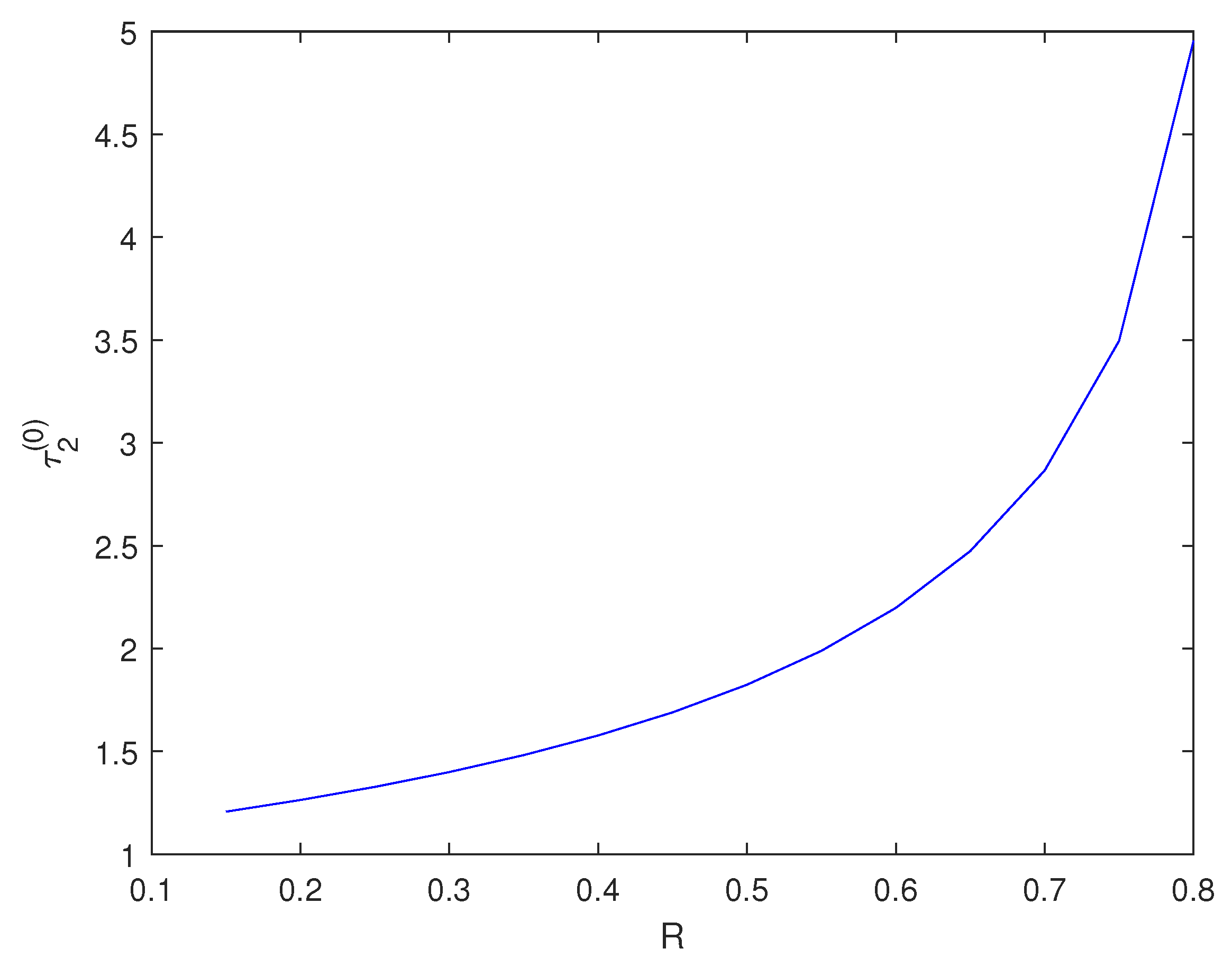
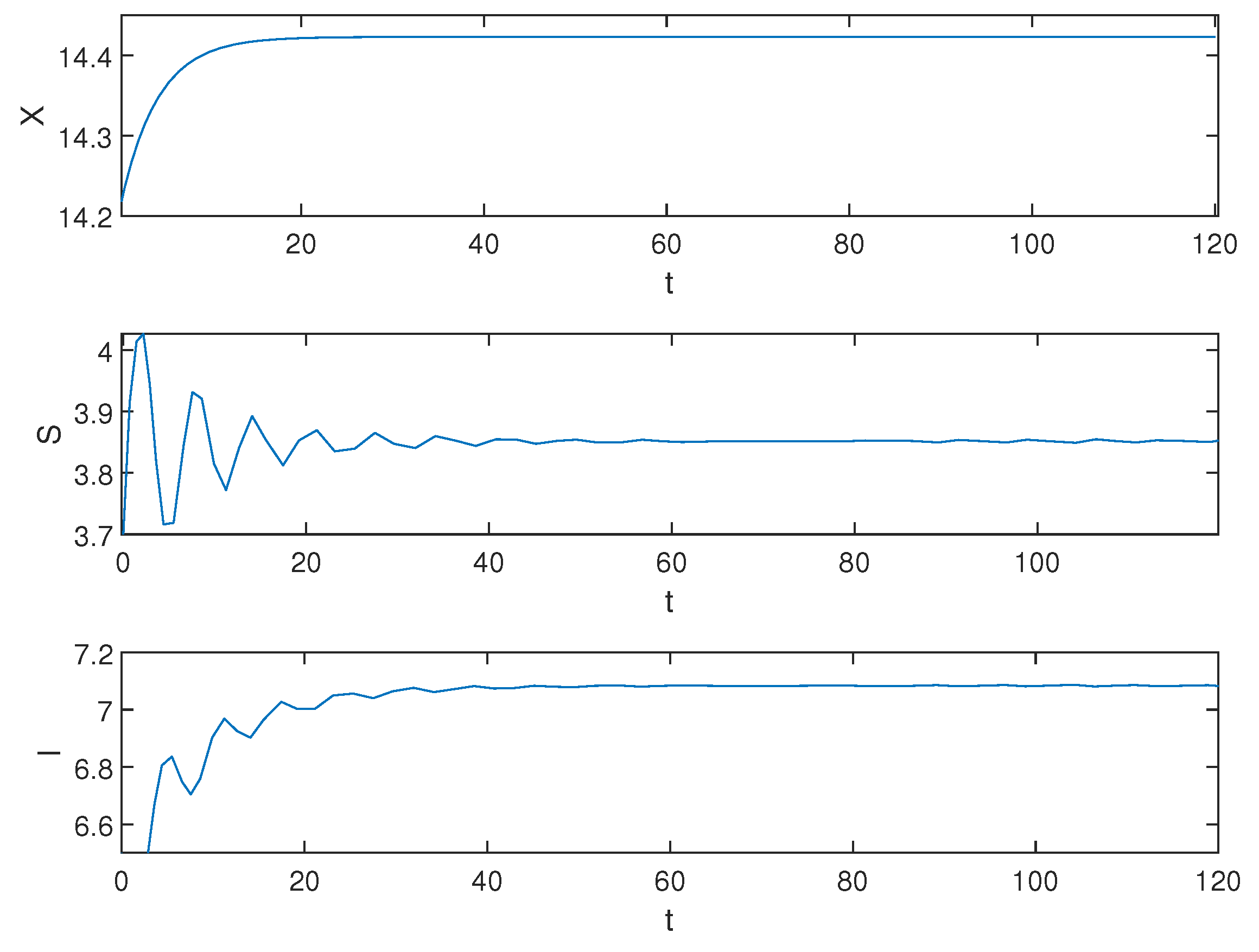
5. Numerical Simulations
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Augusto, L.; Boča, A. Tree functional traits, forest biomass, and tree species diversity interact with site properties to drive forest soil carbon. Nat. Commun. 2022, 13, 1097. [Google Scholar] [CrossRef] [PubMed]
- Quirion, B.R.; Domke, G.M.; Walters, B.F.; Lovett, G.M.; Fargione, J.E.; Greenwood, L. Insect and disease disturbances correlate with reduced carbon sequestration in forests of the contiguous United States. Front. For. Glob. Chang. 2021, 4, 716582. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Boguś, M.I. Fungi of entomopathogenic potential in Chytridiomycota and Blastocladiomycota, and in fungal allies of the Oomycota and Microsporidia. IMA Fungus 2021, 12, 29. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Ramos, A.P.; Lnácio, M.L. Insights into the role of fungi in pine wilt disease. J. Fungi 2021, 7, 780. [Google Scholar] [CrossRef] [PubMed]
- Dou, G.; Yan, D. Research progress on biocontrol of pine wilt disease by microorganisms. Forests 2022, 13, 1047. [Google Scholar] [CrossRef]
- Luo, Y.; Huang, H.; Roques, A. Early monitoring of forest wood-boring pests with remote sensing. Annu. Rev. Entomol. 2023, 68, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Borralho, N.; Cabral, P.; Caetano, M. Recent advances in forest insect pests and diseases monitoring using UAV-based data: A systematic review. Forests 2022, 13, 911. [Google Scholar] [CrossRef]
- Bozzini, A.; Francini, S.; Chirici, G.; Battisti, A.; Faccoli, M. Spruce Bark Beetle Outbreak Prediction through Automatic Classification of Sentinel-2 Imagery. Forests 2023, 14, 1116. [Google Scholar] [CrossRef]
- Jentsch, P.C.; Bauch, C.T.; Anand, M. Fire mitigates bark beetle outbreaks in serotinous forests. Theor. Ecol. 2021, 14, 611–621. [Google Scholar] [CrossRef]
- Xu, W.; Chen, S.; Chen, L. Modeling of the prevention and control of forest pest. J. Biol. Syst. 2007, 15, 539–550. [Google Scholar] [CrossRef]
- Kishi, S.; Sun, J.; Kawaguchi, A.; Ochi, S.; Yoshida, M.; Yamanaka, T. Characteristic features of statistical models and machine learning methods derived from pest and disease monitoring datasets. R. Soc. Open Sci. 2023, 10, 230079. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Jara, J.P.; Vogt-Geisse, K.; Correa, M.C.G.; Vilches-Ponce, K.; Pérez, L.M.; Chowell, G. Modeling the impact of agricultural mitigation measures on the spread of Sharka disease in sweet cherry orchards. Plants 2023, 12, 3442. [Google Scholar] [CrossRef] [PubMed]
- Buch, K.R.; Fefferman, N.H. Mathematical Model of Basal Sprout Production in Vector-Borne Tree Disease. Forests 2023, 14, 349. [Google Scholar] [CrossRef]
- Song, Y.; Peng, Y.; Zhang, T. The spatially inhomogeneous Hopf bifurcation induced by memory delay in a memory-based diffusion system. J. Differ. Equ. 2021, 300, 597–624. [Google Scholar] [CrossRef]
- Jiang, W.; An, Q.; Shi, J. Formulation of the normal form of Turing–Hopf bifurcation in partial functional differential equations. J. Differ. Equ. 2020, 268, 705–715. [Google Scholar] [CrossRef]
- Sawada, Y.; Takeuchi, Y.; Dong, Y. Stability analysis of a single-species logistic model with time delay and constant inflow. Appl. Math. Lett. 2023, 138, 108514. [Google Scholar] [CrossRef]
- Chen, S.; Ma, W. Integrable nonlocal PT-symmetric generalized so(3,ℝ)-mKdV equations. Commun. Theor. Phys. 2023, 75, 125003. [Google Scholar] [CrossRef]
- Ma, W. AKNS Type Reduced Integrable Hierarchies with Hamiltonian Formulations. Rom. J. Phys. 2023, 68, 116. [Google Scholar] [CrossRef]
- Guo, Q.; Li, H.; Zheng, W.; Pan, J.; Lu, J.; Li, J. Analysis of genetic diversity and prediction of Larix species distribution in the Qinghai–Tibet Plateau, China. J. For. Res. 2023, 34, 705–715. [Google Scholar] [CrossRef]
- Mishima, K.; Hirakawa, H.; Iki, T.; Fukuda, Y.; Hirao, T.; Tamure, A.; Makoto, T. Comprehensive collection of genes and comparative analysis of full-length transcriptome sequences from Japanese larch (Larix kaempferi) and Kuril larch (Larix gmelinii var. japonica). BMC Plant Biol. 2022, 22, 470. [Google Scholar] [CrossRef]
- Skrzecz, I.; Perlińska, A. Current problems and tasks of forest protection in Poland. Folia For. Polonica 2018, 60, 161–172. [Google Scholar] [CrossRef]
- Chen, D.; Li, Y.; Zhang, Q.; Zhang, S.; Wang, H.; Zhen, Z.; Zhao, L.-L.; Kong, X.-B. Population divergence of aggregation pheromone responses in Ips subelongatus in northeastern China. Insect Sci. 2016, 23, 728–738. [Google Scholar] [CrossRef]
- Lee, C.Y.; Nam, Y.; Park, C.; Bae, Y.J.; Choi, W.I. Forecasting spring flight of Ips subelongatus (Coleoptera: Curculionidae: Scolytinae) in Japanese larch Larix kaempferi (Pinales: Pinaceae) forests in the Republic of Korea. Environ. Entomol. 2019, 48, 1481–1488. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, H.; Lu, Q.; Decock, C.; Li, Y.; Zhang, X. Taxonomy and pathogenicity of Leptographium species associated with Ips subelongatus infestations of Larix spp. in northern China, including two new species. Mycol. Prog. 2017, 16, 1–13. [Google Scholar] [CrossRef]
- Du, H.; Fang, J.; Shi, X.; Yu, C.; Deng, M.; Zhang, S.; Liu, F.; Zhang, Z.; Han, F.; Kong, X. Insights into the divergence of Chinese Ips bark beetles during evolutionary adaptation. Biology 2022, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Zhang, S.; Liu, F.; Xu, F.; Zhang, F.; Guo, X.; Zhang, Z.; Kong, X.B. SEM analysis of sensilla on the mouthparts and antennae of Asian larch bark beetle Ips subelongatus. Micron 2021, 140, 102976. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Fang, J.; Shi, X.; Zhang, S.; Liu, F.; Yu, C.; Zhang, Z.; Kong, X. Comparative analysis of eight mitogenomes of bark beetles and their phylogenetic implications. Insects 2021, 12, 949. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Liu, M.; Zhang, S.; Liu, F.; Zhang, Z.; Zhang, Q.; Kong, X. Chemical signal interactions of the bark beetle with fungal symbionts, and host/non-host trees. J. Exp. Bot. 2020, 71, 6084–6091. [Google Scholar] [CrossRef] [PubMed]
- Majee, S.; Adak, S.; Jana, S.; Mandal, M.; Kar, T.K. Complex dynamics of a fractional-order SIR system in the context of COVID-19. J. Appl. Math. Comput. 2022, 21, 4051–4074. [Google Scholar] [CrossRef]
- Li, B.; Eskandari, Z.; Avazzadeh, Z. Dynamical Behaviors of an SIR Epidemic Model with Discrete Time. Fractal Fract. 2022, 6, 659. [Google Scholar] [CrossRef]
- Dong, L.; Lin, X.; Bettinger, P.; Liu, Z. The contributions of stand characteristics on carbon sequestration potential are triple that of climate variables for Larix spp. plantations in northeast China. Sci. Total Environ. 2024, 911, 168726. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, C.; Wang, H. Diffusive Spatial Movement with Memory. J. Diff. Equ. 2020, 32, 979–1002. [Google Scholar] [CrossRef]
- Yu, J. Existence and stability of a unique and exact two periodic orbits for an interactive wild and sterile mosquito model. J. Diff. Equ. 2020, 269, 10395–10415. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Descriptions | Unit |
|---|---|---|
| X | The population size of the Ips subelongatus Motschulsky | PCS |
| S | The population size of susceptible Larix spp. | trees |
| I | The population size of infected Larix spp. | trees |
| r | The proliferation rate of Ips subelongatus Motschulsky | - |
| N | Environmental capacity of Ips subelongatus Motschulsky | PCS |
| The natural mortality rate of Ips subelongatus Motschulsky | - | |
| Mortality rate of infected trees | - | |
| The cure rate of infected trees | - | |
| The time required for infected trees to recover their health | Month | |
| k | Infection rate coefficient | - |
| M | The mortality rate caused by pests being preyed upon by natural enemies | - |
| R | The input rate of artificially planted trees | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Ding, Y. Dynamic Analysis of a Delayed Differential Equation for Ips subelongatus Motschulsky-Larix spp. Axioms 2024, 13, 232. https://doi.org/10.3390/axioms13040232
Li Z, Ding Y. Dynamic Analysis of a Delayed Differential Equation for Ips subelongatus Motschulsky-Larix spp. Axioms. 2024; 13(4):232. https://doi.org/10.3390/axioms13040232
Chicago/Turabian StyleLi, Zhenwei, and Yuting Ding. 2024. "Dynamic Analysis of a Delayed Differential Equation for Ips subelongatus Motschulsky-Larix spp." Axioms 13, no. 4: 232. https://doi.org/10.3390/axioms13040232
APA StyleLi, Z., & Ding, Y. (2024). Dynamic Analysis of a Delayed Differential Equation for Ips subelongatus Motschulsky-Larix spp. Axioms, 13(4), 232. https://doi.org/10.3390/axioms13040232