
Microbe-Mediated Mn Oxidation—A Proposed Model of Mineral Formation

 , ,
, , 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
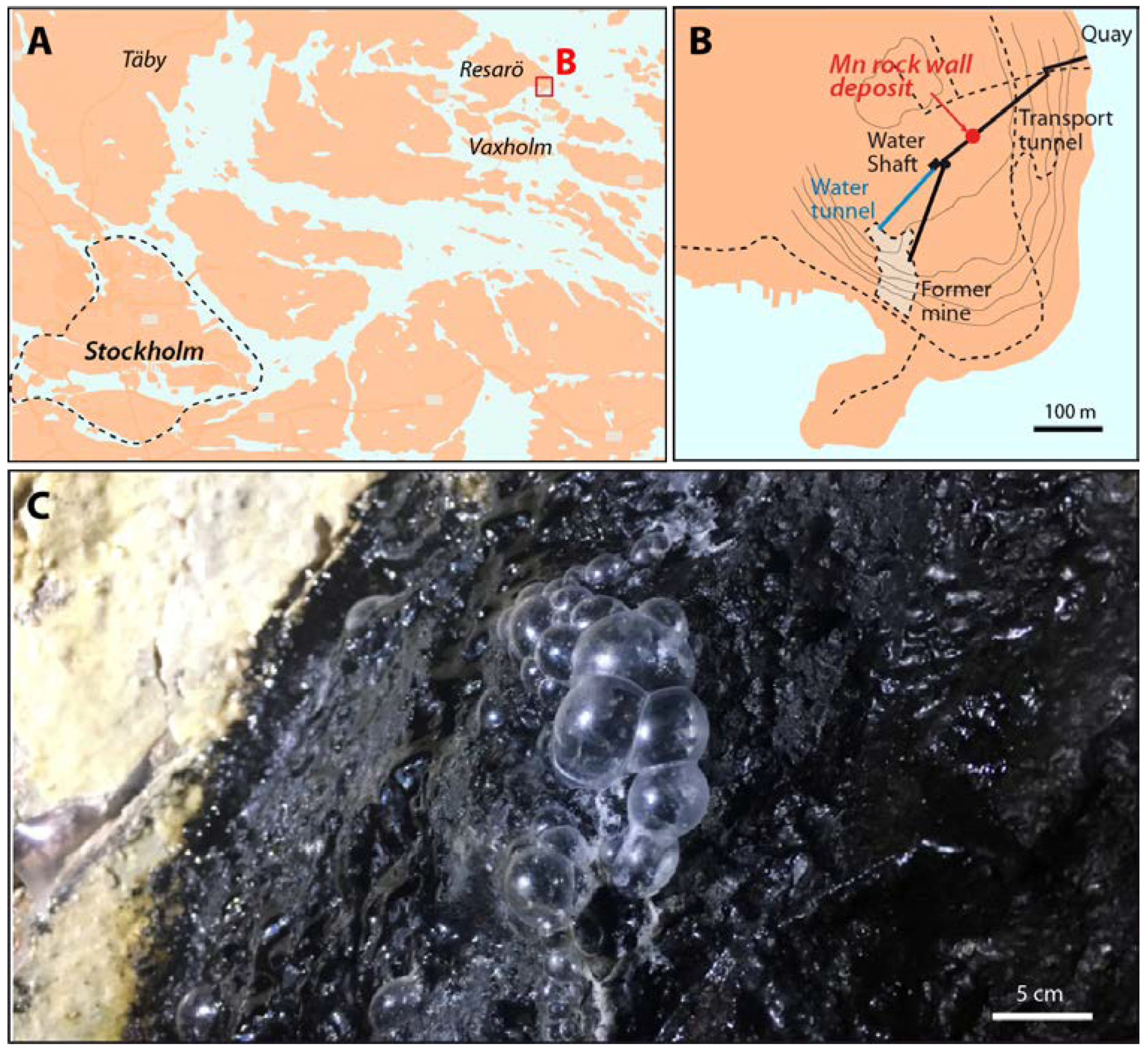
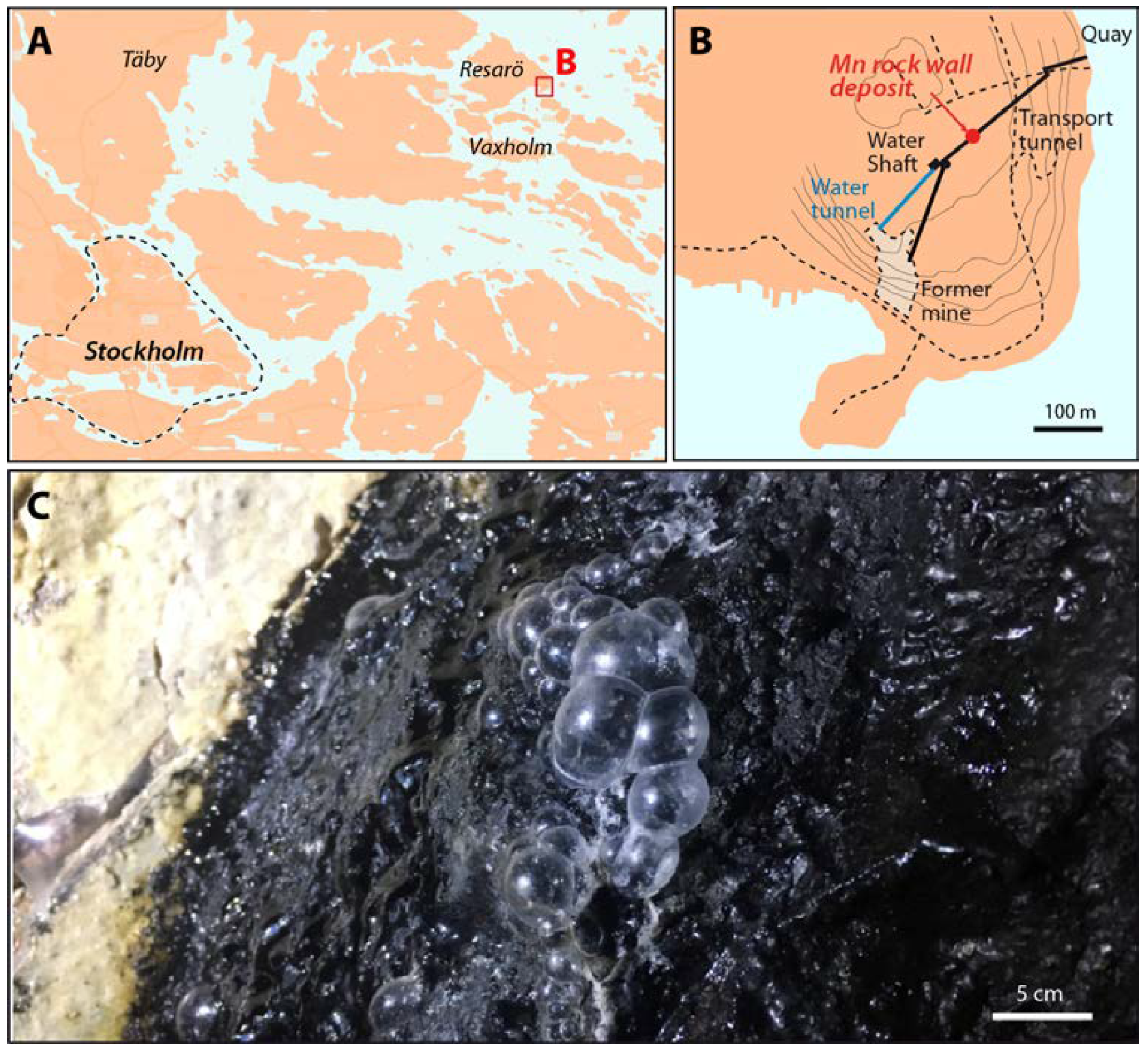
2. Study Site
3. Materials and Methods
3.1. Isolation and Identification of Mn-Oxidizing Species
3.2. Characterization of Mn Minerals in Cultures and the Field Site Samples
3.2.1. Environmental Scanning Electron Microscopy
3.2.2. Transmission Electron Microscopy
3.2.3. X-ray Absorption Spectroscopy
4. Results and Discussion
4.1. Isolated Mn-Oxidizing Microorganisms
4.2. Products of Mn(II) Oxidation in Cultures
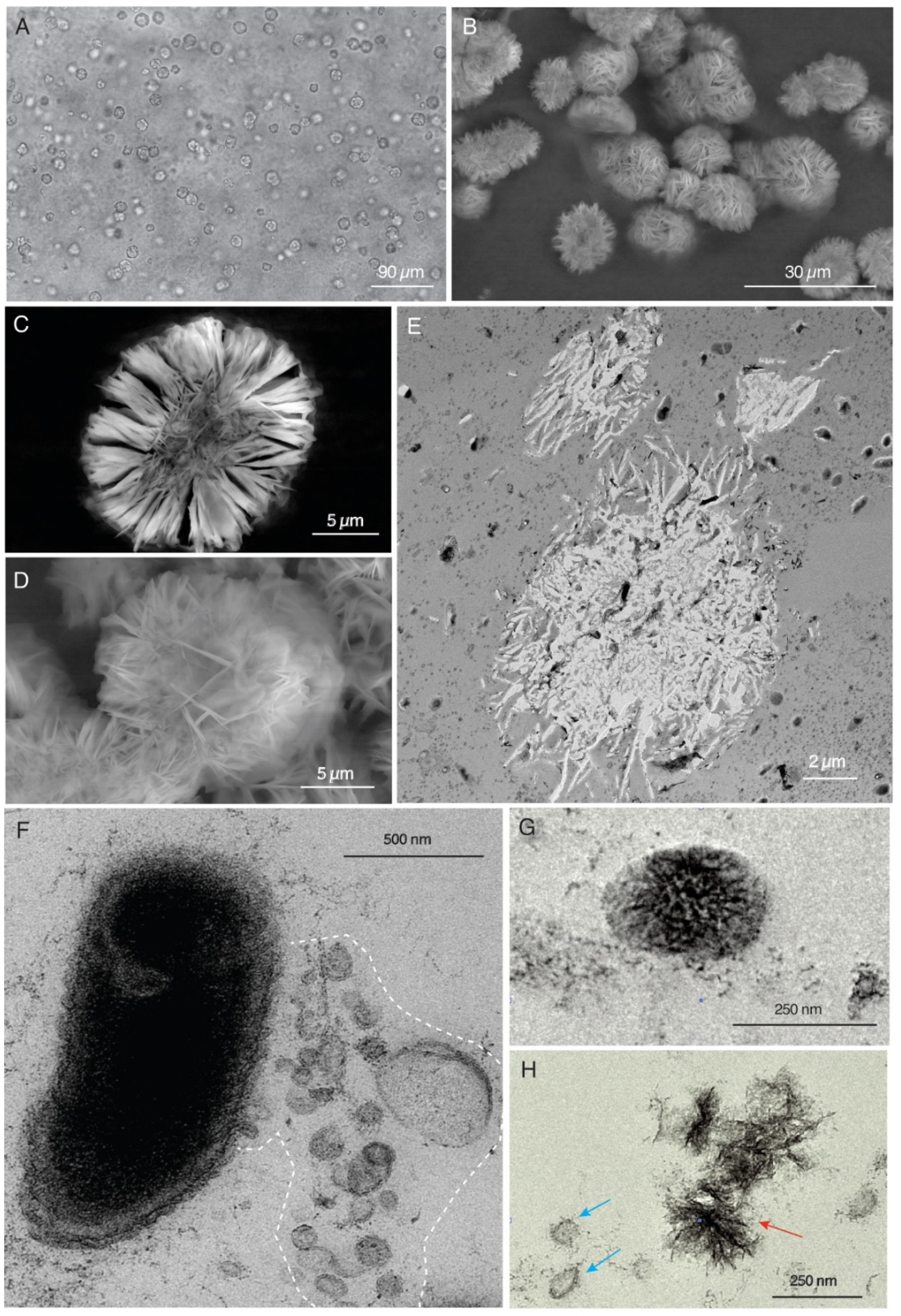
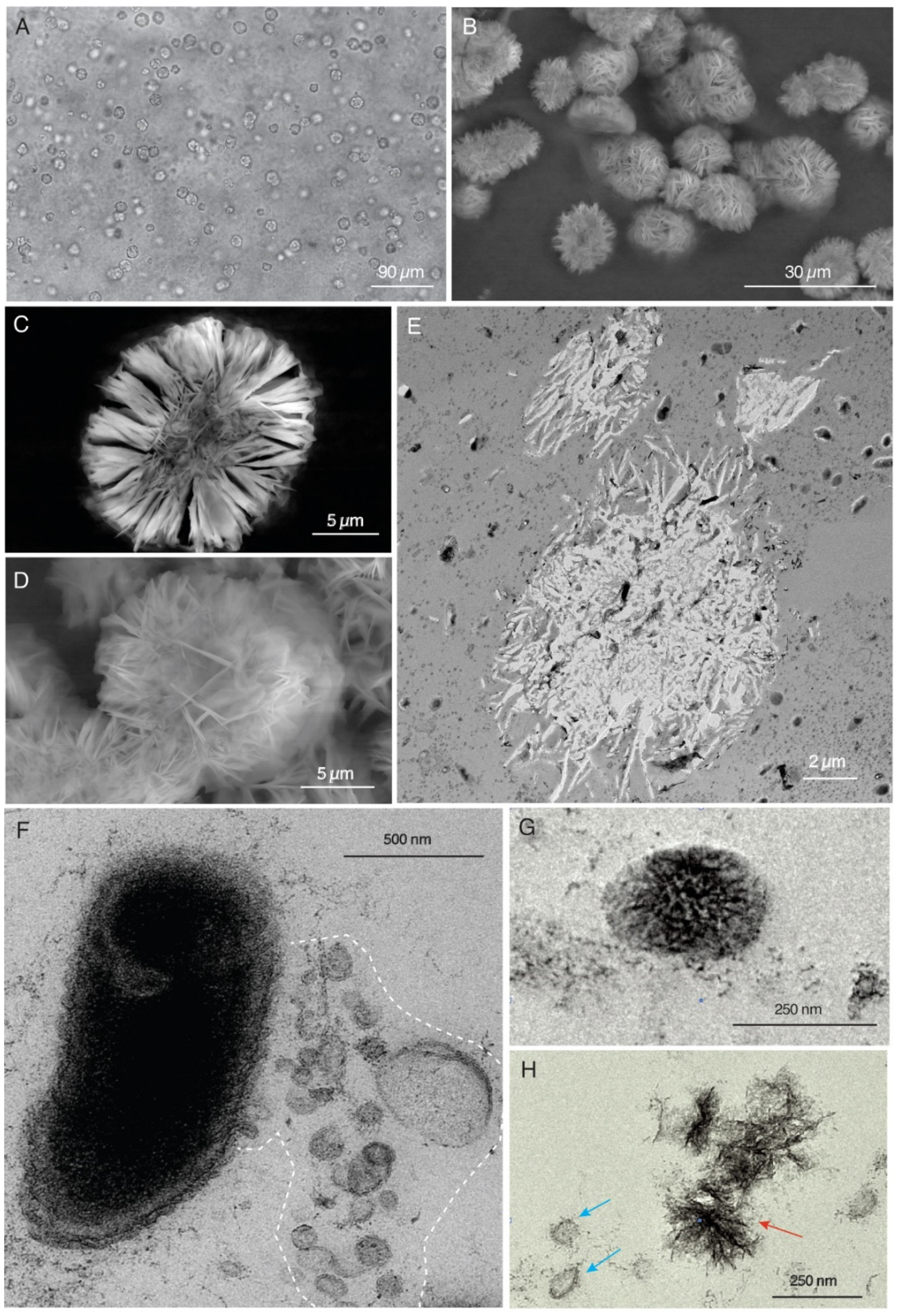
4.2.1. Hydrogenophaga sp.
4.2.2. Pedobacter sp.
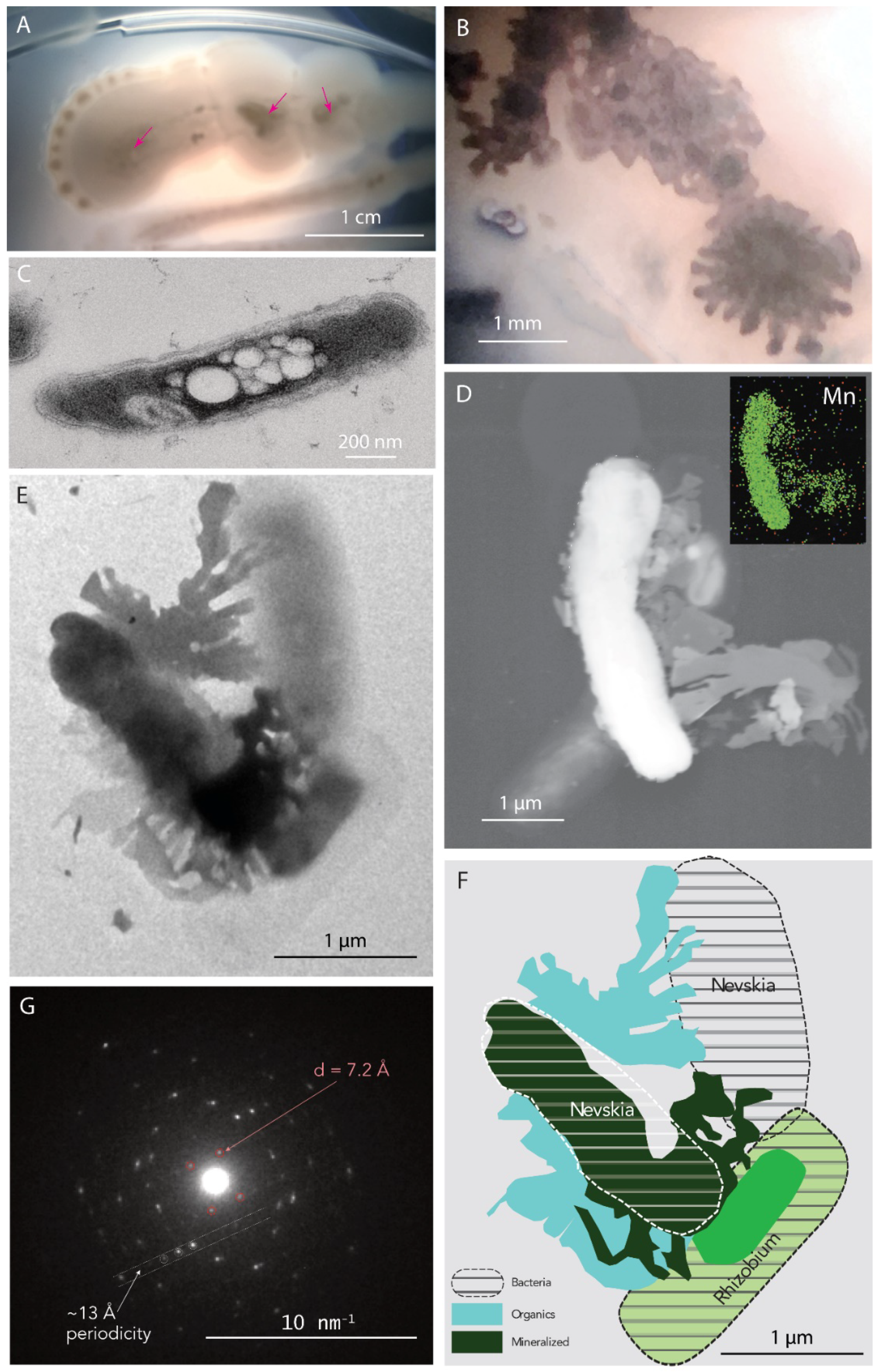
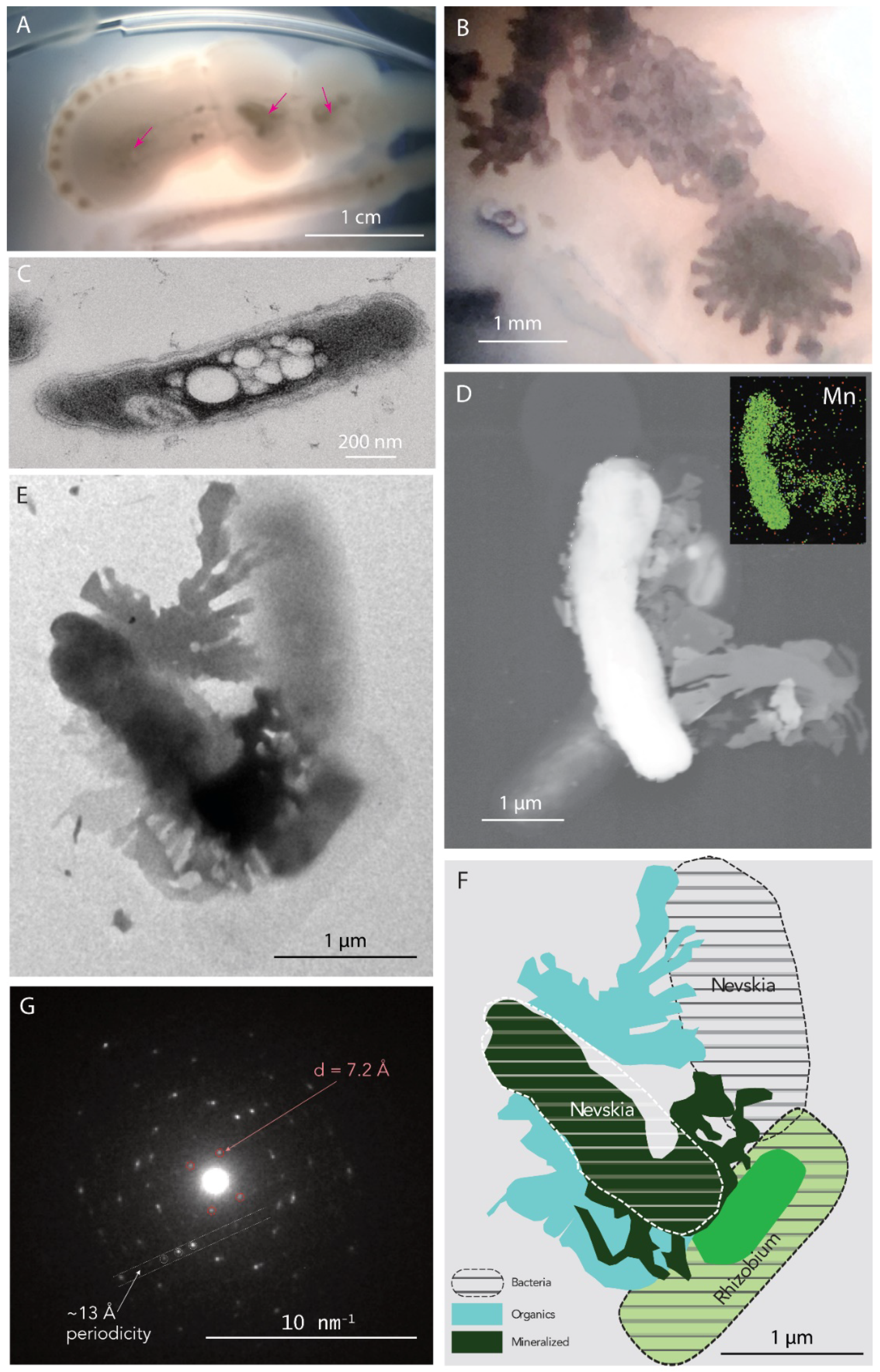
4.2.3. Nevskia sp.—Rhizobium sp.
4.2.4. Cladosporium sp.—Hydrogenophaga sp.—Rhizobium sp.—Nevskia sp.
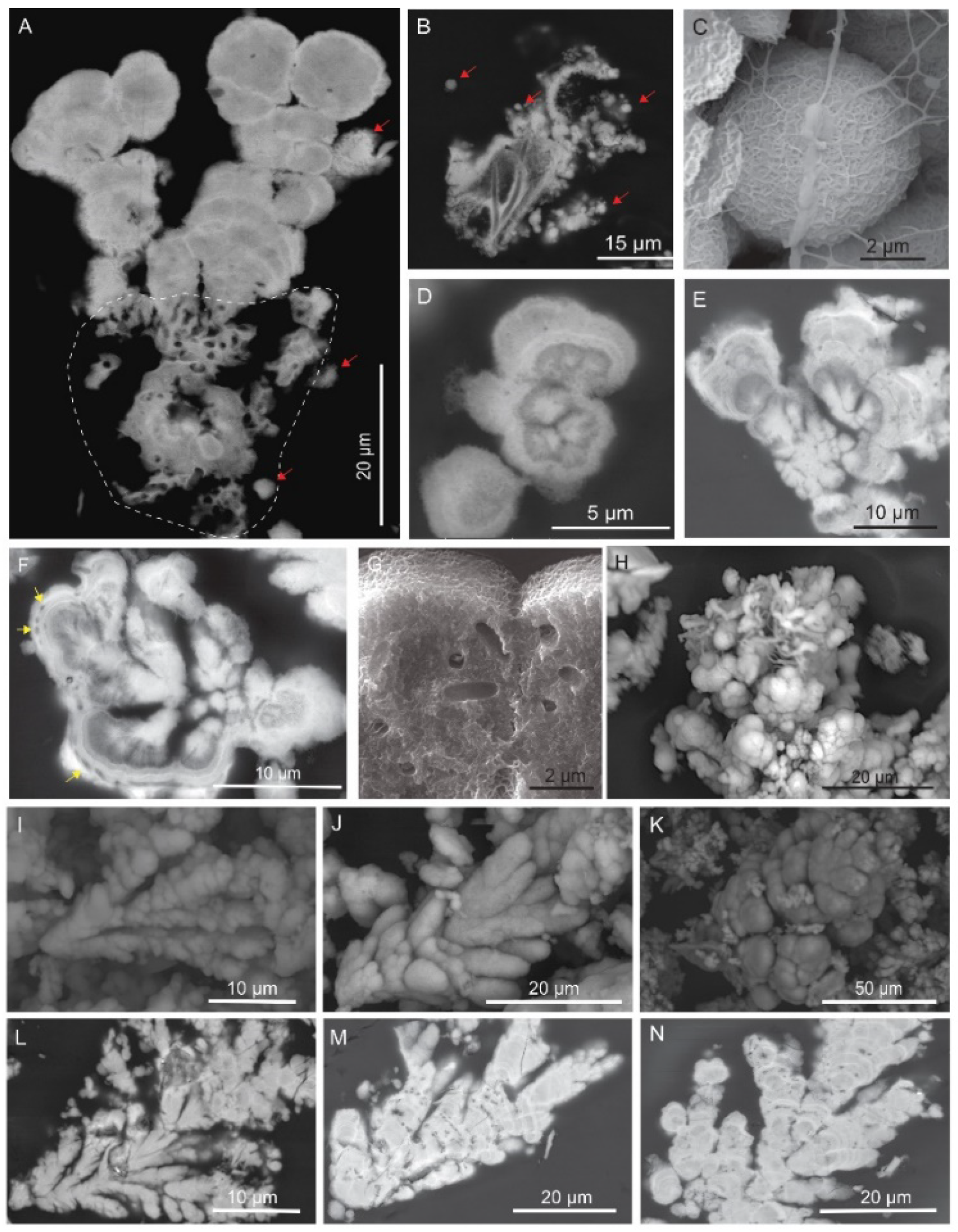
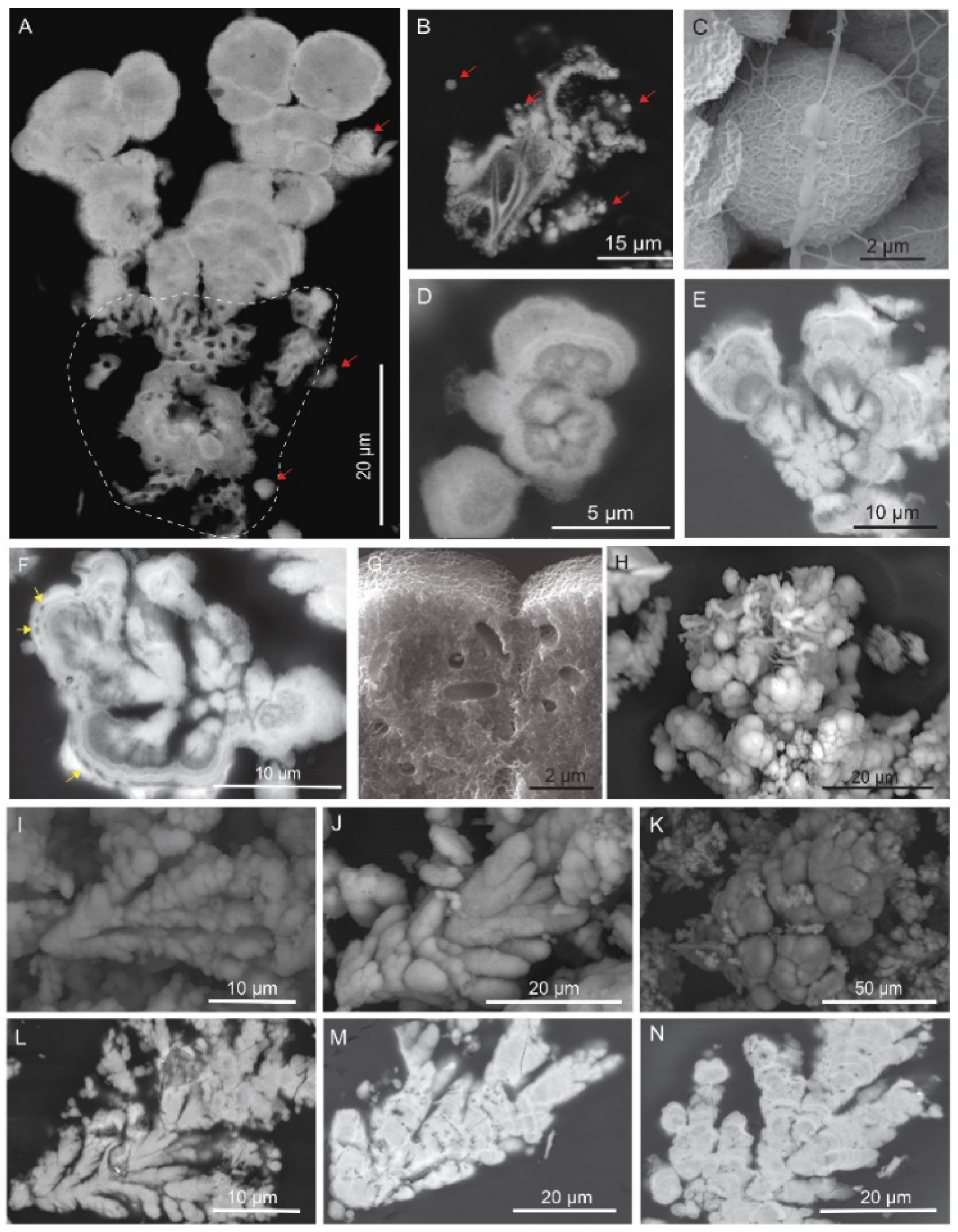
4.3. Products of Mn(II) Oxidation at the Field Site
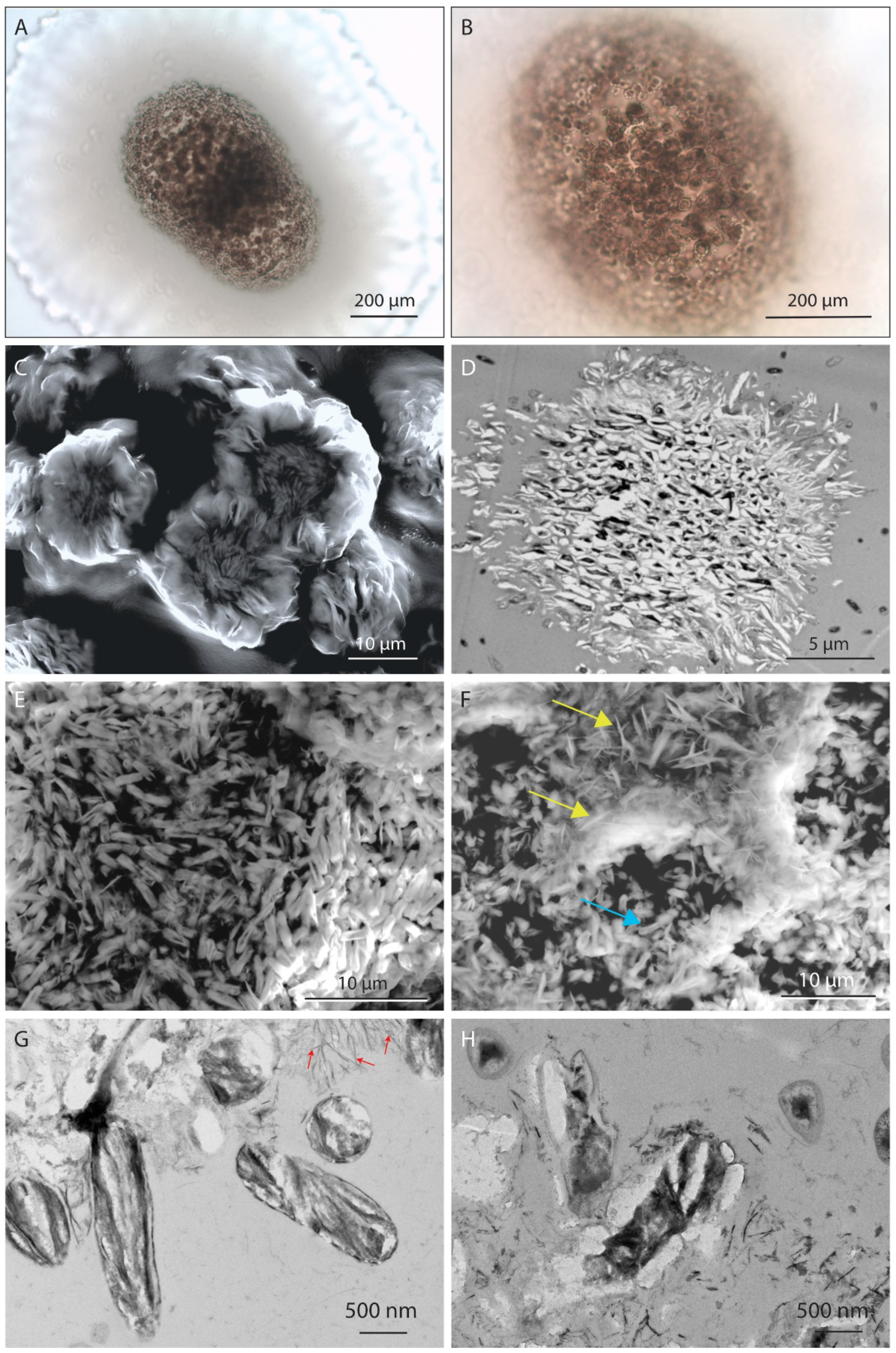
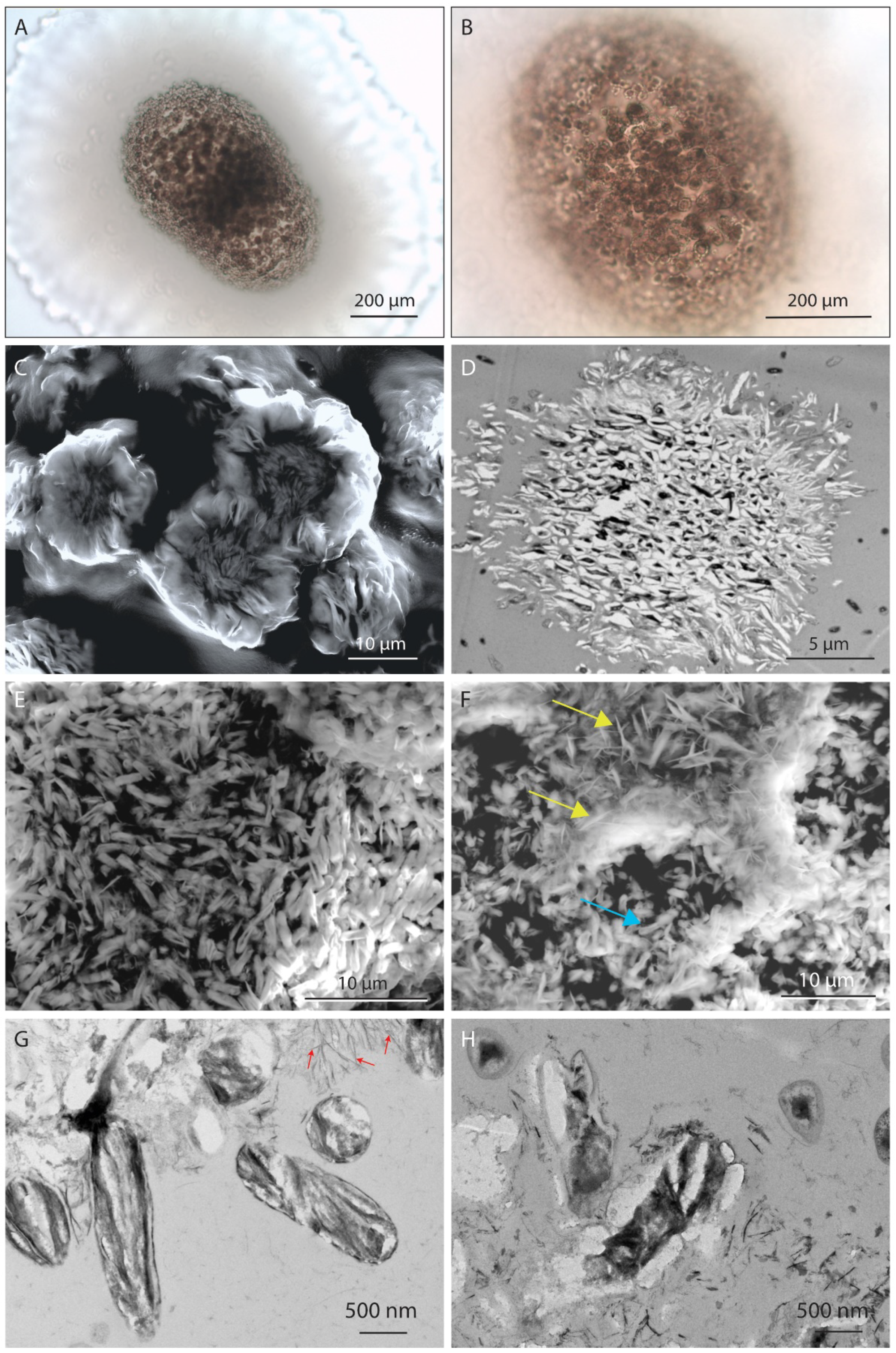
4.3.1. Microscale
4.3.2. Nanoscale
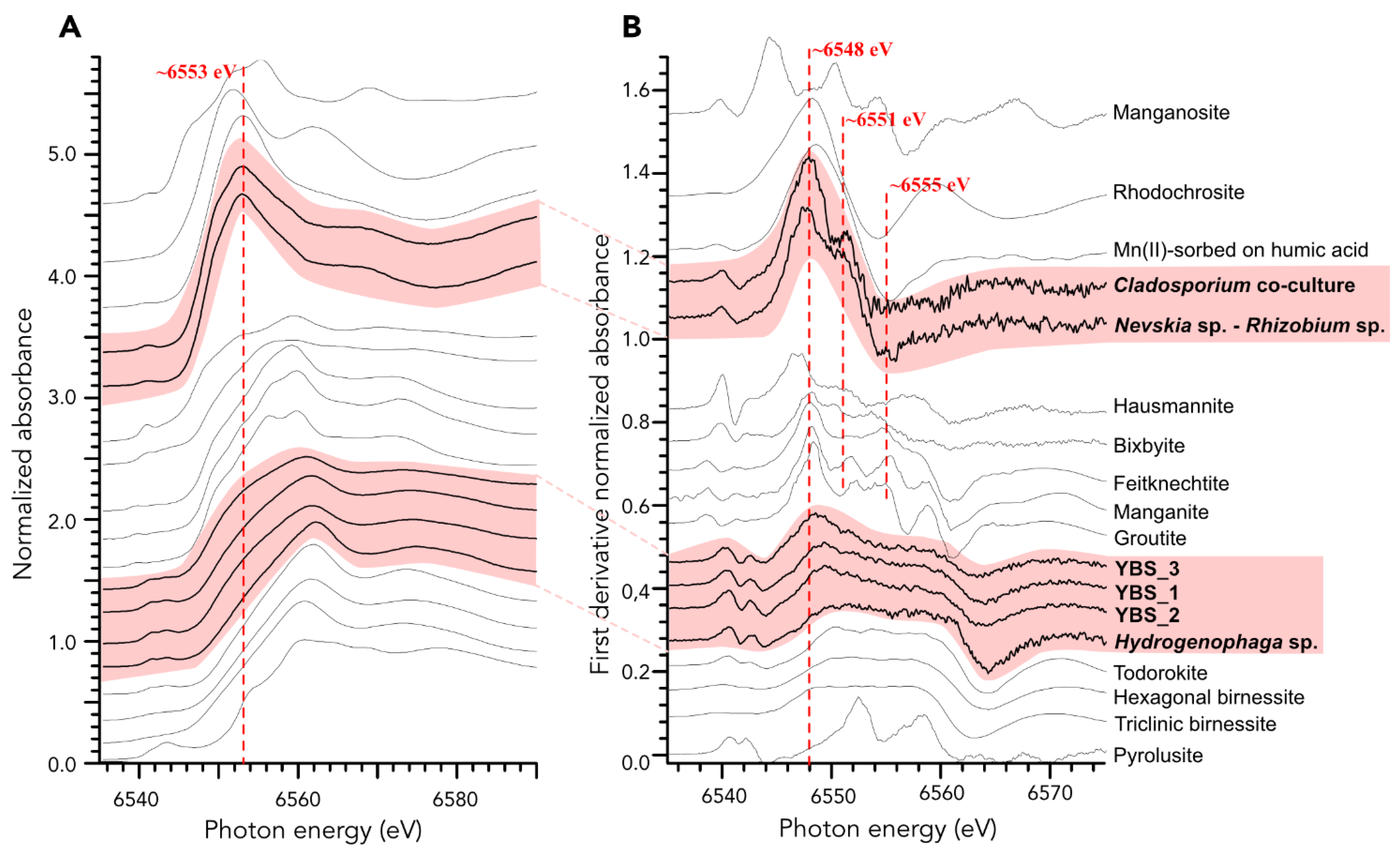
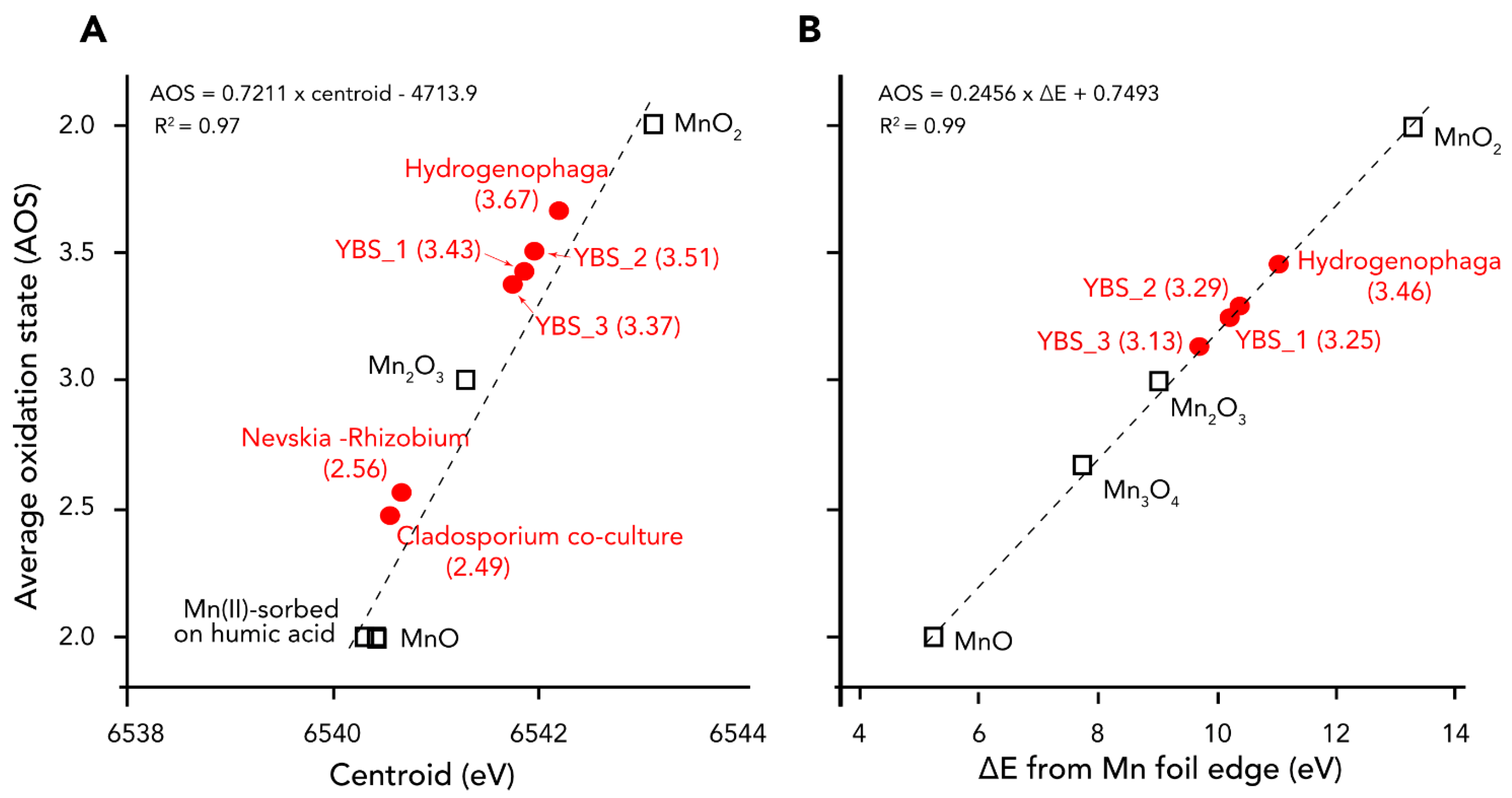
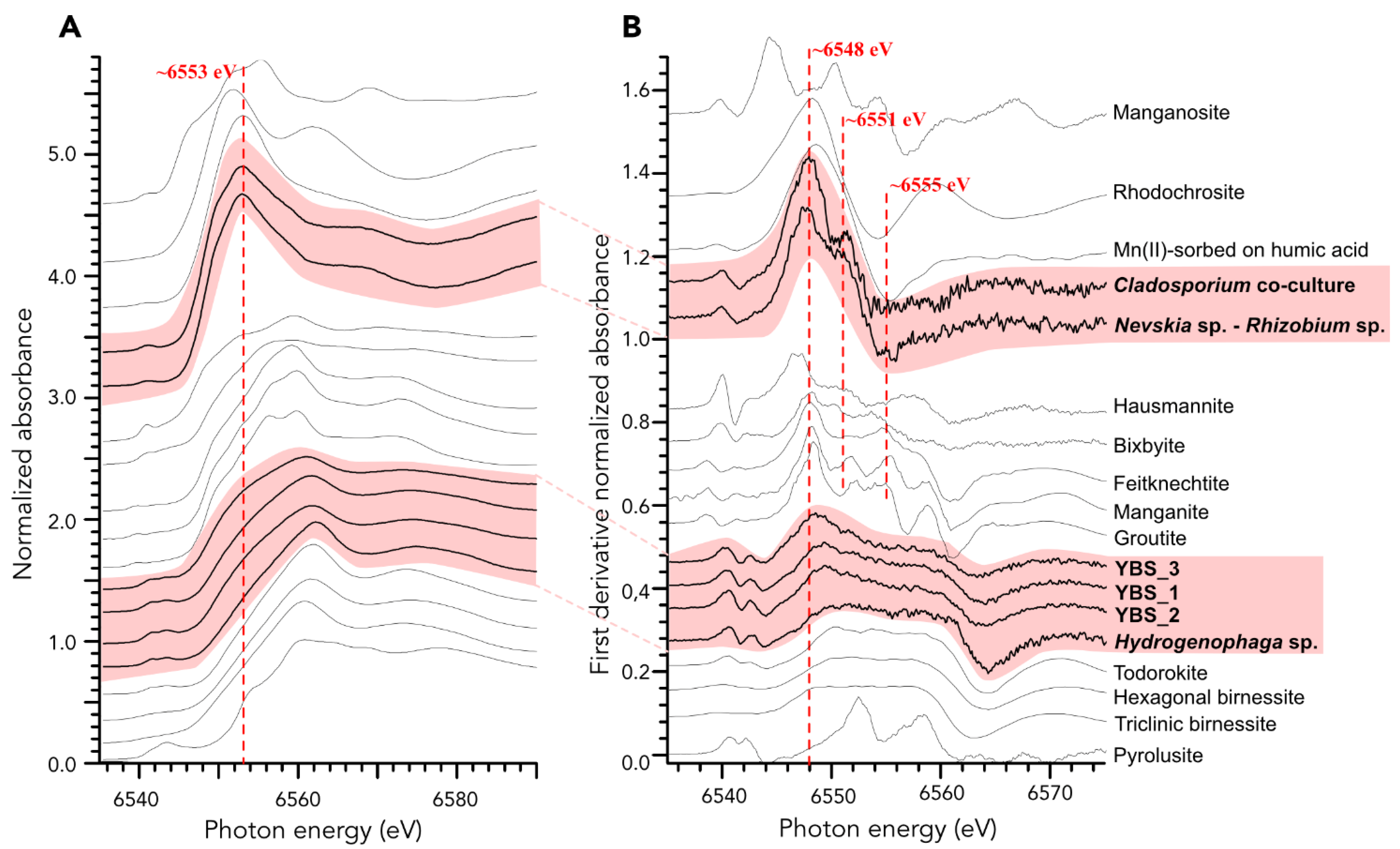
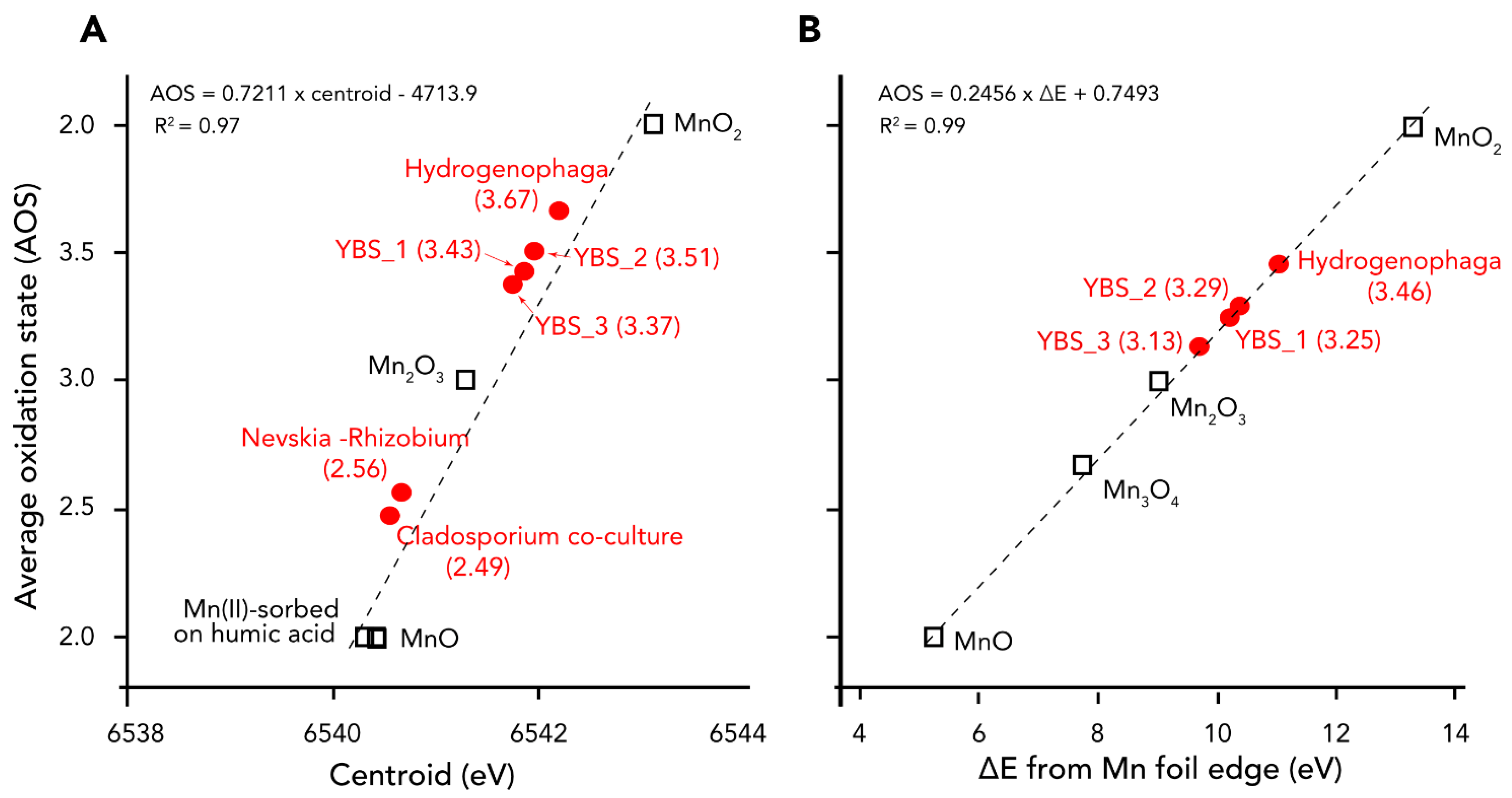
4.4. Manganese Phases and Oxidation States in the Samples Based on XANES Analyses
4.5. Promotion of Mn Oxidation by Biofilm Formation
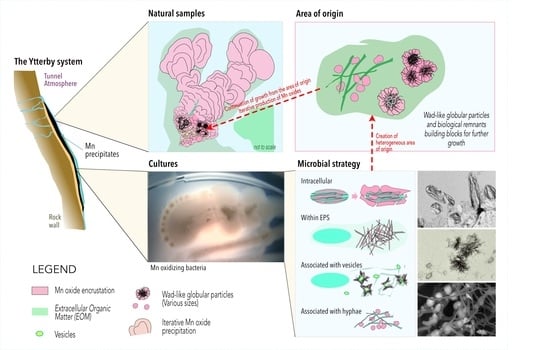
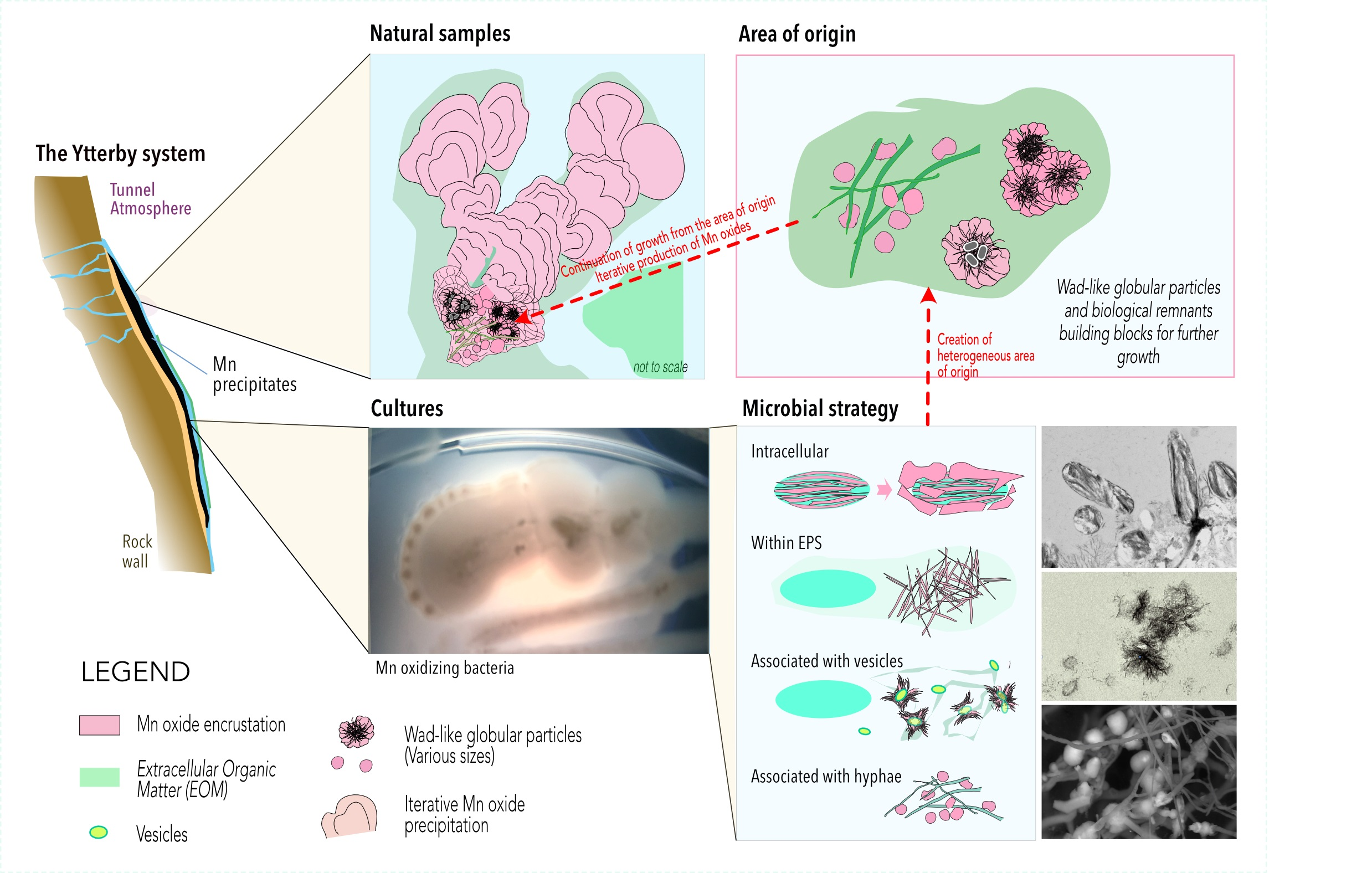
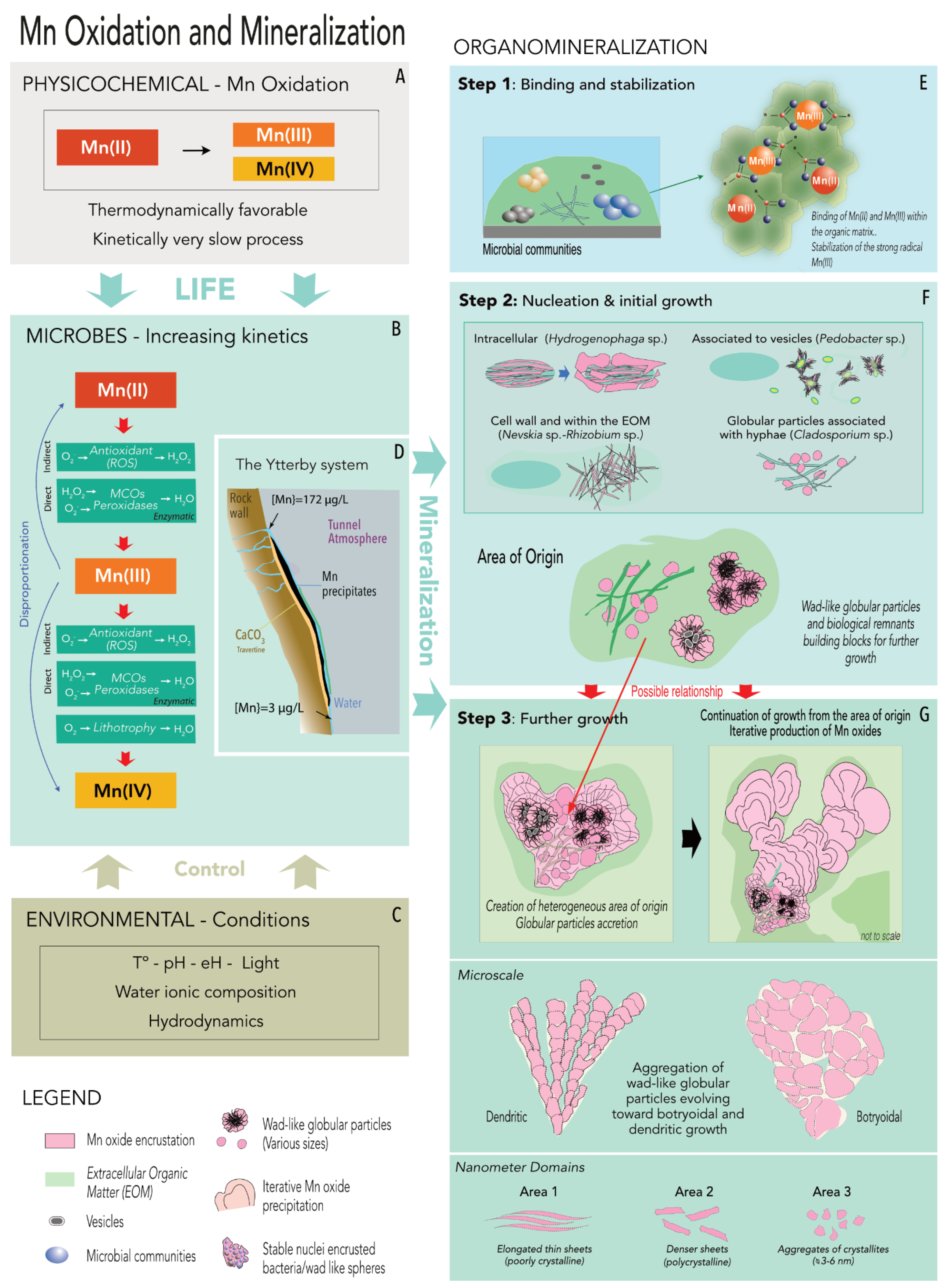
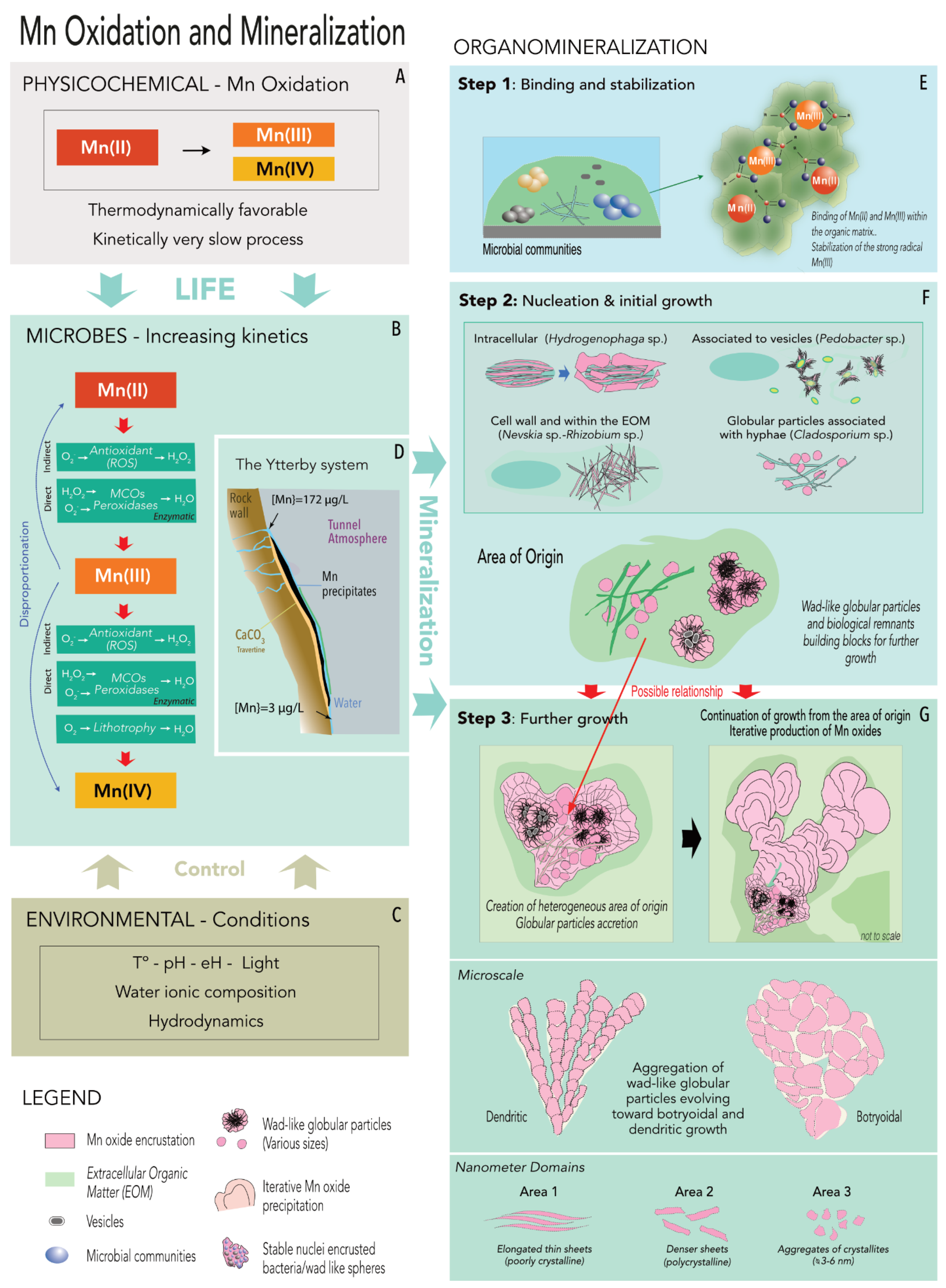
4.6. Sequestration of Mn Oxides in the Ytterby Mine Tunnel—A Proposed Model of Formation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tebo, B.M.; Bargar, J.R.; Clement, B.G.; Dick, G.J.; Murray, K.J.; Parker, D.; Verity, R.; Webb, S.M. Biogenic manganese oxides: Properties and mechanisms of formation. Annu. Rev. Earth Planet. Sci. 2004, 32, 287–328. [Google Scholar] [CrossRef] [Green Version]
- Luther, G.W., III. The role of one- and two-electron transfer reactions in forming thermodynamically unstable intermediates as barriers in multi-electron redox reactions. Aquat. Geochem. 2010, 16, 395–420. [Google Scholar] [CrossRef]
- Morgan, J.J. Kinetics of reaction of O2 and Mn(II) species in aqueous solutions. Geochim. Cosmochim. Acta 2005, 69, 35–48. [Google Scholar] [CrossRef]
- Hansel, C.M.; Learman, D.R. Geomicrobiology of manganese. In Ehrlich’s Geomicrobiology, 6th ed.; Ehrlich, H.L., Newman, D.K., Kappler, A., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 401–452. [Google Scholar]
- Zhou, D.; Kim, D.-G.; Ko, S.-O. Heavy metal adsorption with biogenic manganese oxides generated by Pseudomonas putida strain MnB1. J. Ind. Eng. Chem. 2015, 24, 132–139. [Google Scholar] [CrossRef]
- Bargar, J.R.; Tebo, B.M.; Bergmann, U.; Webb, S.M.; Glatzel, P.; Chiu, V.Q.; Villalobos, M. Biotic and abiotic products of Mn(II) oxidation by spores of the marine Bacillus sp. strain SG-1. Am. Mineral. 2005, 90, 143–154. [Google Scholar] [CrossRef]
- Webb, S.M.; Tebo, B.M.; Bargar, J.R. Structural influences of sodium and calcium ions on the biogenic manganese oxides produced by the marine Bacillus sp., strain SG-1. Geomicrobiol. J. 2005, 22, 181–193. [Google Scholar] [CrossRef]
- Sjöberg, S.; Stairs, C.W.; Allard, B.; Homa, F.; Martin, T.; Sjöberg, V.; Ettema, T.J.; Dupraz, C. Microbiomes in a manganese oxide producing ecosystem in the Ytterby mine, Sweden: Impact on metal mobility. FEMS Microbiol. Ecol. 2020, 96, 11. [Google Scholar] [CrossRef]
- Sun, W.; Kitchaev, D.A.; Kramer, D.; Ceder, G. Non-equilibrium crystallization pathways of manganese oxides in aqueous solution. Nat. Commun. 2019, 10, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enghag, P. Jordens Grundämnen Och Deras Upptäckt, Sällsynt-Ädelt-Aktivt; Industrilitteratur AB: Stockholm, Sweden, 1999. [Google Scholar]
- Nordenskjöld, I. Ytterby Fältspatbrott. Skandinavisk Tidskrift för Lervaru- och Stenindustri. Organ för Tegel, Cement, Beton, Kalk, Sten Och Byggnadsmaterialier; N:o 6 &7; Stockholm, Sweden, 1904. (In Swedish) [Google Scholar]
- Sjöberg, S. The Ytterby Mine—A Historical Review and an Evaluation of Its Suggested Spatial Coupling to Multiple Sclerosis (MS). Bachelor’s Thesis, Stockholm University, Stockholm, Sweden, 2012. [Google Scholar]
- Sjöberg, S. Characterization of an REE-Enriched Black Substance in Fractured Bedrock in the Ytterby Mine. Master’s Thesis, Stockholm University, Stockholm, Sweden, 2014. [Google Scholar]
- Cailleau, G.; Verrecchia, E.P.; Braissant, O.; Emmanuel, L. The biogenic origin of needle fibre calcite. Sedimentology 2009, 56, 1858–1875. [Google Scholar] [CrossRef]
- Sjöberg, S.; Allard, B.; Rattray, J.E.; Callac, N.; Grawunder, A.; Ivarsson, M.; Sjöberg, V.; Karlsson, S.; Skelton, A.; Dupraz, C. Rare earth element enriched birnessite in water-bearing fractures, the Ytterby mine, Sweden. Appl. Geochem. 2017, 78, 158–171. [Google Scholar] [CrossRef]
- Swedish Fortifications Agency (Fortifikationsverket). Utredningsprogram Ytterby, Vaxholms Kommun; 4:610, 4:564, 4:611. 4:9, Rev, A; Swedish Fortifications Agency: Eskilstuna, Sweden, 2012. [Google Scholar]
- Krumbein, W.E.; Altmann, H.J. A new method for the detection and enumeration of manganese oxidizing and reducing microorganisms. Helgoländer Wiss. Meeresunters. 1973, 25, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Tebo, B.M.; Clement, B.G.; Dick, G.D. Biotransformations of manganese. In Manual of Environmental Microbiology, 3rd ed.; Hurst, C.H., Ed.; ASM Press: Washington, DC, USA, 2007; pp. 1223–1238. [Google Scholar]
- Spang, A.; Saw, J.H.; Jørgensen, S.L.; Zaremba-Niedzwiedzka, K.; Martijn, J.; Lind, A.E.; van Eijk, R.; Schleper, C.; Guy, L.; Ettema, T.J.G. Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature 2015, 521, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Bokulich, N.; Robeson, M.; Dillon, M.; Ziemski, M.; Kaehler, B.; O’Rourke, D. Bokulich-Lab/RESCRIPt: 2021.8.0.dev0 (Zenodo, 2021). 2021. Available online: https://zenodo.org/record/4811136#.YWlYdRpByUk (accessed on 20 August 2021).
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- NCBI Sequence Read Archive (SRA). Available online: https://www.ncbi.nlm.nih.gov/sra (accessed on 20 August 2021).
- Dupraz, C.; Fowler, A.; Tobias, C.; Visscher, P.T. Stromatolic knobs in Storr’s lake (San Salvador, Bahamas): A model system for formation and alternation of laminae. Geobiology 2013, 11, 527–548. [Google Scholar] [CrossRef] [PubMed]
- Hawes, P.; Netherton, C.L.; Mueller, M.; Wileman, T.; Monaghan, P. Rapid freeze- substitution preserves membranes in high pressure frozen tissue culture cells. J. Microsc. 2007, 226, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klementiev, K.; Norén, K.; Carlson, S.; Clauss Sigridsson, K.; Persson, I. The BALDER Beamline at the MAX IV Laboratory. J. Phys. Conf. Ser. 2016, 712, 01223. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.X.; Virtasalo, J.J.; Österholm, P.; Burton, E.D.; Peltola, P.; Ojala, A.E.K.; Hogmalm, J.K.; Åström, M.E. Manganese accumulation and solid-phase speciation in a 3.5 m thick mud sequence from the estuary of an acidic and Mn-rich creek, northern Baltic Sea. Chem. Geol. 2016, 437, 56–66. [Google Scholar] [CrossRef]
- Ravel, B.; Newville, M. ATHENA, ARTEMIS, HEPHAESTUS: Data analysis for X-ray absorption spectroscopy using IFEFFIT. J. Synchrotron Radiat. 2005, 12, 537–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.H.; Zhu, M.; Ginder-Vogel, M.; Ni, C.; Parikh, S.J.; Sparks, D.L. Formation of nano-crystalline todorokite from biogenic Mn oxides. Geochim. Cosmochim. Acta 2010, 74, 3232–3245. [Google Scholar] [CrossRef]
- Yu, C.; Drake, H.; Mathurin, F.A.; Åström, M.E. Cerium sequestration and accumulation in fractured crystalline bedrock: The role of Mn-Fe (hydr-)oxides and clay minerals. Geochim. Et Cosmochim. Acta 2017, 199, 370–389. [Google Scholar] [CrossRef]
- Sjöberg, S.; Stairs, C.; Allard, B.; Hallberg, R.; Homa, F.; Martin, T.; Ettema, T.J.G.; Dupraz, C. Bubble biofilm: Bacterial colonization of air-air interface. Biofilm 2020, 2, 100030. [Google Scholar] [CrossRef] [PubMed]
- Marcus, D.N.; Pinto, A.; Anantharaman, K.; Ruberg, S.A.; Kramer, E.L.; Raskin, L.; Dick, G.J. Diverse manganese(II)-oxidizing bacteria are prevalent in drinking water systems. Environ. Microbiol. Rep. 2017, 9, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Dangeti, S.; McBeth, J.M.; Roshani, B.; Vyskocil, J.M.; Rindall, B.; Chang, W. Microbial communities and biogenic Mn-oxides in an on-site biofiltration system for cold Fe-(II)- and Mn(II)-rich groundwater treatment. Sci. Total Environ. 2020, 710, 136386. [Google Scholar] [CrossRef] [PubMed]
- Bohu, T.; Akob, D.M.; Abratis, M.; Lazar, C.S.; Küsel, K. Biological low-pH Mn(II) oxidation in a manganese deposit influenced by metal-rich groundwater. Appl. Environ. Microbiol. 2016, 82, 3009–3021. [Google Scholar] [CrossRef] [Green Version]
- Mota, E.A.; Felestrino, E.B.; Leão, V.A.; Guerra-Sá, R. Manganese (II) removal from aqueous solution by Cladosporium Halotolerans and Hypocrea jecorina. Biotechn. Rep. 2020, 25, e00431. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Sun, F. Intracellular sequestration of manganese and phosphorous in a metal-resistant fungus Cladosporium cladosporioides from deep-sea sediment. Extremophiles. 2007, 11, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Drits, V.A.; Silvester, E.; Gorshkov, A.I.; Manceau, A. Structure of synthetic monoclinic Na-rich birnessite and hexagonal birnessite: I. Results from X-ray diffraction and selected-area electron diffraction. Am. Mineral. 1997, 82, 946–961. [Google Scholar] [CrossRef]
- Villalobos, M.; Toner, B.; Bargar, J.; Sposito, G. Characterization of the manganese oxide produced by Pseudomonas putida strain MnB1. Geochim. Cosmochim. Acta 2003, 67, 2649–2662. [Google Scholar] [CrossRef]
- Villalobos, M.; Lanson, B.; Manceau, A.; Toner, B.; Sposito, G. Structural model for the biogenic Mn oxide produced by Pseudomonas putida. Am. Mineral. 2006, 91, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Atkins, A.L.; Shaw, S.; Peacock, C.L. Nucleation and growth of todorokite from birnessite: Implications for trace-metal cycling in marine sediments. Geochim. Cosmochim. Acta 2014, 144, 109–125. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Taillefert, M.; Sun, J.; Wang, Q.; Borkiewicz, O.J.; Liu, P.; Yang, L.; Chen, S.; Chen, H.; Tang, Y. Redox cycling driven transformation of layered manganese oxides to tunnel structures. J. Am. Chem. Soc. 2020, 142, 2506–2513. [Google Scholar] [CrossRef] [PubMed]
- Gabr, R.M.; Hassan, S.H.A.; Shoreit, A.A.M. Biosorption of lead and nickel by living and non-living cells of Pseudomonas aeruginosa ASU 6a. Int. Biodeterior. Biodegrad. 2008, 62, 195–203. [Google Scholar] [CrossRef]
- Larsen, E.I.; Sly, L.I.; McEwan, A.G. Manganese(II) adsorption and oxidation by whole cells and a membrane fraction of Pedomicrobium sp. ACM 3067. Arch. Microbiol. 1999, 171, 257–264. [Google Scholar] [CrossRef]
- Ghiorse, W.C.; Hirsch, P. An ultrastructural study of iron and manganese deposition associated with extracellular polymers of Pedomicrobium-like budding bacteria. Arch. Microbiol. 1979, 123, 213–226. [Google Scholar] [CrossRef]
- De Vrind, J.; de Groot, A.; Brouwers, G.J.; Tommassen, J.; de Vrind–de Jong, E. Identification of a novel Gsp-related pathway required for secretion of the manganese-oxidizing factor of Pseudomonas putida strain GB-1. Mol. Microbiol. 2003, 47, 993–1006. [Google Scholar] [CrossRef]
- Emerson, D.; Ghiorse, W.C. Ultrastructure and chemical composition of the sheath of Lepthothrix discophora SP-6. J. Bacteriol. 1993, 175, 7808–7818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, M.; Sugita, T.; Shimizu, M.; Ohode, Y.; Iwamoto, K.; de Vrind de Jong, E.W.; de Vrind, J.P.M.; Corstjens, P.L.A.M. Partial purification and characteriazation of manganese oxidizing factors of Pseudomonas fluorescens GB-1. Appl. Environ. Microbiol. 1997, 63, 4793–4799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, C.A.; Tebo, B.M. Marine Bacillus spores as catalysts for oxidative precipitation and sorption of metals. J. Molec. Microbiol. Biotechnol. 1999, 1, 71–78. [Google Scholar]
- Francis, C.A.; Casciotti, K.L.; Tebo, B.M. Localization of Mn(II)-oxidizing activity and the putative multicopper oxidase, MnxG, to the exosporium of the marine Bacillus sp. strain SG-1. Arch. Microbiol. 2002, 178, 450–456. [Google Scholar] [CrossRef]
- Nealson, K.H.; Tebo, B. Structural features of manganese precipitating bacteria. Orig. Life Evol. Biosph. 1980, 10, 117–126. [Google Scholar] [CrossRef]
- Daye, M.; Klepac-Ceraj, V.; Pajusalu, M.; Rowland, S.; Farrell-Sherman, A.; Beukes, N.; Tamura, N.; Fournier, G.; Bosak, T. Light-driven anaerobic microbial oxidation of manganese. Nature 2019, 576, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.L.; Salerno, J.C. Energy coupling in Mn2+ oxidation by a marine bacterium. Arch. Microbiol. 1990, 154, 12–17. [Google Scholar]
- Babenzien, H.D.; Cypionka, H.N. Nevskia. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons Inc., in Association with Bergey’s Manual Trust: Hoboken, NJ, USA, 2015; pp. 1–6. [Google Scholar]
- Heldal, M.; Tumyr, O. Morphology and content of dry matter and some elements in cells and stalks of Nevskia from an eutrophic lake. Can. J. Microbiol. 1986, 32, 89–92. [Google Scholar] [CrossRef]
- Kim, S.J. New characterization of takanelite. Am. Mineral. 1991, 76, 1426–1430. [Google Scholar]
- Post, J.E.; Heaney, P.J.; Ertl, A. Rietveld refinement of the ranciéite structure using synchrotron powder diffraction data. Powder Diffr. 2007, 23, 10–14. [Google Scholar] [CrossRef]
- Cygan, R.T.; Post, J.E.; Heaney, P.J.; Kubicki, J.D. Molecular models of birnessite and related hydrated layered minerals. Am. Mineral. 2012, 97, 1505–1514. [Google Scholar] [CrossRef]
- Grangeon, S.; Warmont, F.; Tournassat, C.; Lanson, B.; Lanson, M.; Elkaïm, E.; Claret, F. Nucleation and growth of feitknechtite from nanocrystalline vernadite precursor. Eur. J. Mineral. 2017, 29, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Wegorzewski, A.V.; Kuhn, T.; Dohrmann, R.; Wirth, R.; Grangeon, S. Mineralogical characterization of individual growth structures of Mn-nodules with different Ni + Cu content from the central Pacific Ocean. Am. Mineral. 2015, 100, 2497–2508. [Google Scholar] [CrossRef]
- Chukrov, F.V. New mineral phases of oceanic manganese micronodules. Int. Geol. Rev. 1980, 22, 154–160. [Google Scholar] [CrossRef]
- Chukrov, F.V.; Drits, V.A.; Gorshkov, A.I.; Sakharov, B.A.; Dikov, Y.P. Structural models of vernadite. Int. Geol. Rev. 1987, 29, 1337–1347. [Google Scholar] [CrossRef]
- Learman, D.R.; Wankel, S.D.; Webb, S.M.; Martinez, N.; Madden, A.S.; Hansel, C.M. Coupled biotic-abiotic Mn (II) oxidation pathways mediates the formation and structural evolution of biogenic Mn oxides. Geochim. Cosmochim. Acta 2011, 75, 6048–6063. [Google Scholar] [CrossRef]
- Webb, S.M.; Tebo, B.M.; Bargar, J.R. Structural characterization of biogenic Mn oxides produced in seawater by the marine Bacillus sp. strain SG-1. Am. Mineral. 2005, 90, 1342–1357. [Google Scholar] [CrossRef]
- Nealson, K.H.; Ford, J. Surface enhancement of bacterial manganese oxidation: Implications for aquatic environments. Geomicrobiol. J. 1980, 2, 21–37. [Google Scholar] [CrossRef]
- Couasnon, T. Determination and Quantification of Reactive Oxygen Species (ROS) in Bacterial Biofilms. Rôle des Espèces Réactives de L’oxygène et Substance Exopolymériques des Bactéries Dans le Processus de Biominéralisation du Manganese. Ph.D. Thesis, Institute de Physique du Globe de Paris, Université Sorbonne Paris Cité, Paris, France, 2019. Chapter 3. pp. 51–88. [Google Scholar]
- Sjöberg, S. Microbially Mediated Manganese Oxides Enriched in Yttrium and Rare Earth Elements in the Ytterby Mine. Ph.D. Thesis, Stockholm University, Stockholm, Sweden, 2019. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sjöberg, S.; Yu, C.; Stairs, C.W.; Allard, B.; Hallberg, R.; Henriksson, S.; Åström, M.; Dupraz, C. Microbe-Mediated Mn Oxidation—A Proposed Model of Mineral Formation. Minerals 2021, 11, 1146. https://doi.org/10.3390/min11101146
Sjöberg S, Yu C, Stairs CW, Allard B, Hallberg R, Henriksson S, Åström M, Dupraz C. Microbe-Mediated Mn Oxidation—A Proposed Model of Mineral Formation. Minerals. 2021; 11(10):1146. https://doi.org/10.3390/min11101146
Chicago/Turabian StyleSjöberg, Susanne, Changxun Yu, Courtney W. Stairs, Bert Allard, Rolf Hallberg, Sara Henriksson, Mats Åström, and Christophe Dupraz. 2021. "Microbe-Mediated Mn Oxidation—A Proposed Model of Mineral Formation" Minerals 11, no. 10: 1146. https://doi.org/10.3390/min11101146
APA StyleSjöberg, S., Yu, C., Stairs, C. W., Allard, B., Hallberg, R., Henriksson, S., Åström, M., & Dupraz, C. (2021). Microbe-Mediated Mn Oxidation—A Proposed Model of Mineral Formation. Minerals, 11(10), 1146. https://doi.org/10.3390/min11101146