
Developmental Origins of Limb Developmental Instability in Human Fetuses: Many Abnormalities Make the Difference

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fetuses and Asymmetry Measurements
2.2. Fluctuating Asymmetry Measurement, Measurement Error and Directional Asymmetry
2.3. The Number of Malformed Organ Systems, the Germ Layer of Origin and Associations, Patterning Defects with Fluctuating Asymmetry
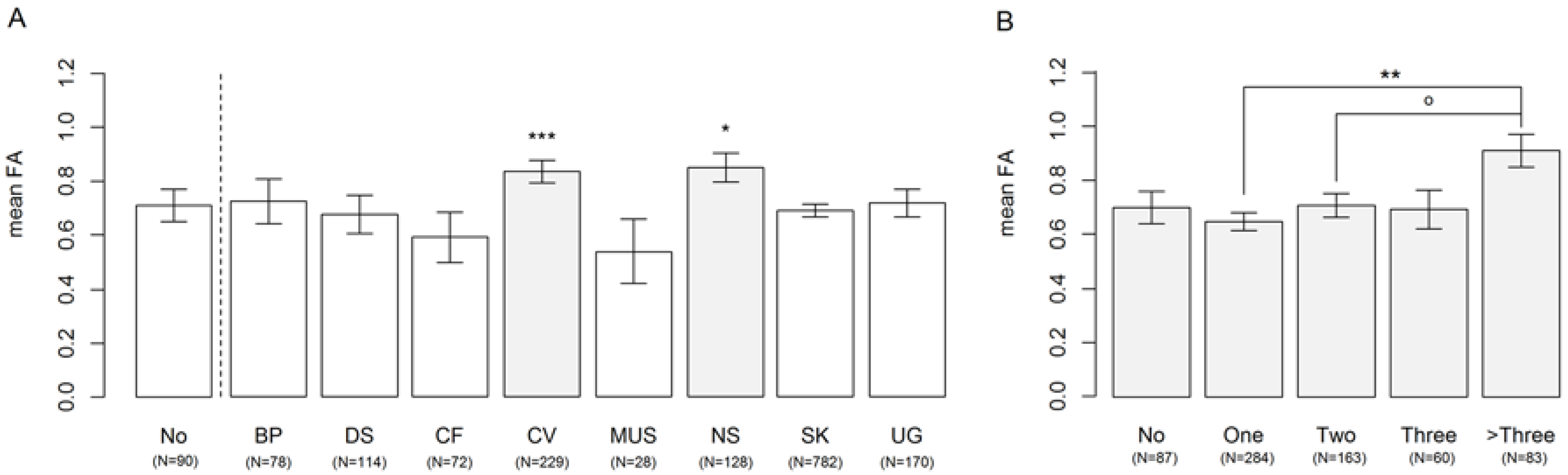
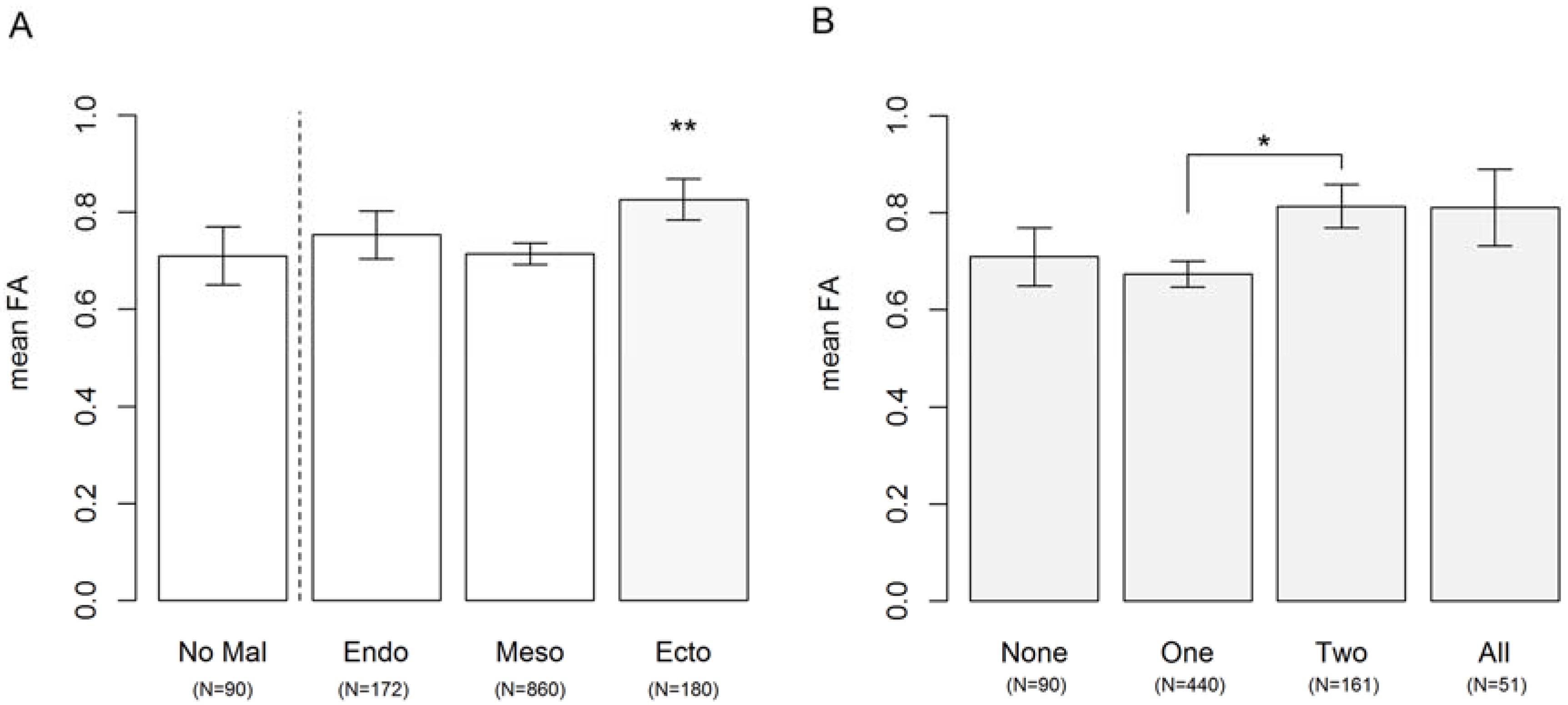
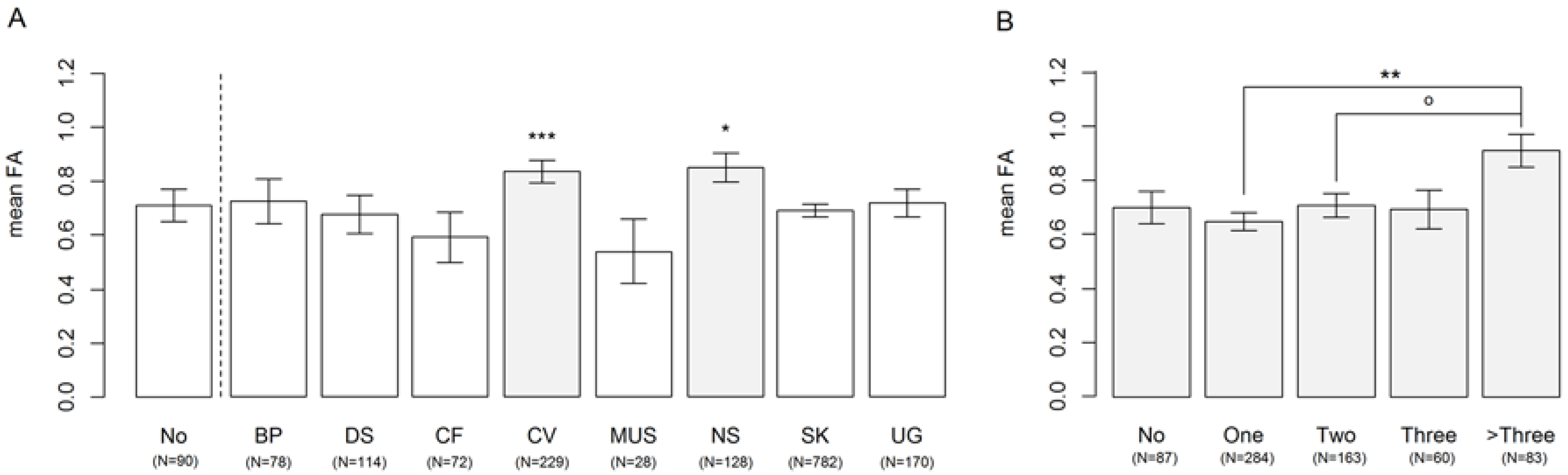
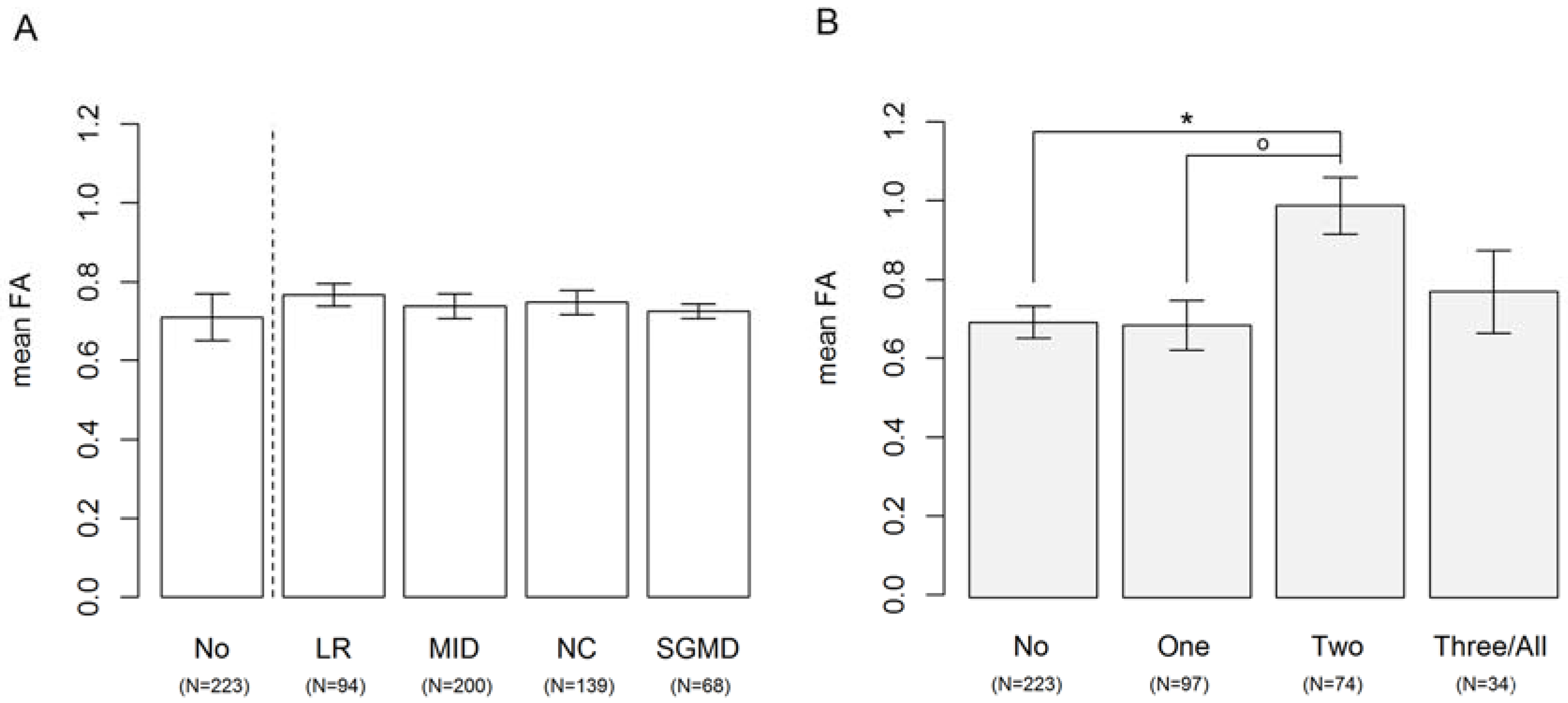
3. Results
3.1. Affected Organ Systems
3.2. Congenital Abnormalities Arising from Different Germ Layers during Development
3.3. Disturbances in Development of Left-Right Patterning, Midline Development, Neural Crest Development and Segementation
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Ethics Statement
References
- Klingenberg, C.P. A developmental perspective on developmental instability: Theory, models, and mechanisms. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 14–34. [Google Scholar]
- Van Dongen, S. Fluctuating asymmetry and developmental instability in evolutionary biology: Past, present and future. J. Evol. Biol. 2006, 19, 1727–1743. [Google Scholar] [CrossRef] [PubMed]
- Van Valen, L. A study of fluctuating asymmetry. Evolution 1962, 16, 125–142. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry: Measurement, analysis, patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Møller, A.P. Developmental stability and fitness: A review. Am. Nat. 1997, 149, 916–932. [Google Scholar] [CrossRef] [PubMed]
- Møller, A.P. Developmental stability is related to fitness. Am. Nat. 1999, 153, 556–560. [Google Scholar] [CrossRef]
- Møller, A.P. A review of developmental instability, parasitism and disease: Infection, genetics and evolution. Infect. Genet. Evol. 2006, 6, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, R.; Møller, A.P. Developmental stability, disease and medicine. Biol. Rev. 1997, 72, 497–548. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, S.; Gangestad, S.W. Human fluctuating asymmetry in relation to health and quality: A meta-analysis. Evol. Hum. Behav. 2011, 32, 380–398. [Google Scholar] [CrossRef]
- Bots, J.; ten Broek, C.M.A.; Belien, J.A.M.; Bugiani, M.; Galis, F.; Van Dongen, S. Higher limb asymmetry in deceased human fetuses and infants with aneuploidy. Sci. Rep. 2014, 4, 3703. [Google Scholar] [CrossRef] [PubMed]
- Ten Broek, C.M.A.; Bakker, A.J.; Varela-Lasheras, I.; Bugiani, M.; Van Dongen, S.; Galis, F. Evo-devo of the human vertebral column: On homeotic transformations, pathologies and prenatal selection. Evol. Boil. 2012, 39, 456–471. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, S. The statistical analysis of fluctuating asymmetry: Reml estimation of a mixed regression model. J. Evol. Biol. 1999, 12, 94–102. [Google Scholar] [CrossRef]
- Ten Broek, C.M.A.; Bots, J.; Varela-Lasheras, I.; Bugiani, M.; Galis, F.; Van Dongen, S. Amniotic fluid deficiency and congenital abnormalities both influence fluctuating asymmetry in developing limbs of human deceased fetuses. PLoS ONE 2013, 8, e81824. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, S.; Wijnaendts, L.C.D.; ten Broek, C.M.A.; Galis, F. Fluctuating asymmetry does not consistently reflect severe developmental disorders in human fetuses. Evolution 2009, 63, 1832–1844. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen, S.; ten Broek, C.M.A; Bots, J.; Galis, F. Changes of fluctuating asymmetry in human fetuses with age. Symmetry 2017, 9, 44. [Google Scholar] [CrossRef]
- Williams, V.S.; Jones, L.V.; Tukey, J.W. Controlling error in multiple comparisons, with examples from state-to-state differences in educational achievement. J. Educ. Behav. Stat. 1999, 24, 42–69. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Lme4: Linear mixed-Effects Models Using Eigen and s4, R package version 1.0-5. 2013.
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- ten Broek, C.M.A.; Bots, J.; Bugiani, M.; Galis, F.; Van Dongen, S. No relationship between vertebral column shifts and limb fluctuating asymmetry in human foetuses. PeerJ. submitted for publication.
- Strachan, T.; Read, A. Human Molecular Genetics; Taylor and Francis Inc.: Boca Raton, FL, USA, 2003; Volume 3, p. 675. [Google Scholar]
- Özener, B. Fluctuating and directional asymmetry in young human males: Effect of heavy working condition and socioeconomic status. Am. J. Phys. Anthropol. 2010, 143, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Pound, N.; Lawson, D.W.; Toma, A.M.; Richmond, S.; Zhurov, A.I.; Penton-Voak, I.S. Facial fluctuating asymmetry is not associated with childhood ill-health in a large british cohort study. Proc. R. Soc. Lond. B Biol. Sci. 2014, 281, 20141639. [Google Scholar]

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broek, C.M.A.t.; Bots, J.; Bugiani, M.; Galis, F.; Dongen, S.v. Developmental Origins of Limb Developmental Instability in Human Fetuses: Many Abnormalities Make the Difference. Symmetry 2017, 9, 51. https://doi.org/10.3390/sym9040051
Broek CMAt, Bots J, Bugiani M, Galis F, Dongen Sv. Developmental Origins of Limb Developmental Instability in Human Fetuses: Many Abnormalities Make the Difference. Symmetry. 2017; 9(4):51. https://doi.org/10.3390/sym9040051
Chicago/Turabian StyleBroek, Clara M. A. ten, Jessica Bots, Marianna Bugiani, Frietson Galis, and Stefan van Dongen. 2017. "Developmental Origins of Limb Developmental Instability in Human Fetuses: Many Abnormalities Make the Difference" Symmetry 9, no. 4: 51. https://doi.org/10.3390/sym9040051
APA StyleBroek, C. M. A. t., Bots, J., Bugiani, M., Galis, F., & Dongen, S. v. (2017). Developmental Origins of Limb Developmental Instability in Human Fetuses: Many Abnormalities Make the Difference. Symmetry, 9(4), 51. https://doi.org/10.3390/sym9040051