1. Introduction
To vision scientists,
symmetry typically means bilateral (mirror) symmetry, and less often it denotes multifold symmetry. Papers devoted to the study of mirror symmetry frequently note the apparently innate appreciation for symmetry and its prominent place in human artifacts of many sorts. In fact we can report that many presentations of psychophysical work on symmetry perception begin with slides of the Notre Dame Cathedral in Paris and front-on views of tigers and mandrills. The structural symmetry of many biological forms is frequently hypothesized to underlie our putative sensitivity to visual symmetry and our fascination with it. Sensitivity to symmetry, it is argued, would have advantages to predator and prey. In fact, studies have shown that both female swallows [
1] and finches [
2] prefer symmetric males, and honey bees [
3] can discriminate symmetry from non-symmetry. Such studies have suggested to some that symmetry in the visual world has played a causal role in the evolution of our visual systems and hence our sensitivity to symmetry, or
vice versa [
4].
Although animate biological forms are structurally symmetrical they rarely project perfect bilateral symmetry to the retinas. In part this is because animate objects do not often position their limbs to produce perfect, object-centered symmetry, and in part because even perfectly symmetrical objects rarely present themselves perpendicular to the line of sight. Therefore, perfectly symmetrical objects typically project to the retinas as
skewed symmetries [
5].
Notwithstanding the rarity of mirror symmetrical images on the retinas, early work on symmetry perception in the vision science literature typically took the position that mirror symmetry is a salient (and perhaps) special image feature of our visual world (see Wagemans [
6], 1995 for a useful review). In fact, human subjects are able to detect symmetry in noise with high efficiency [
7] and we are more sensitive to symmetry than other patterns with similar information content [
8]. So, in these ways symmetry may be special to the visual system but there are other ways in which it is not. Vision scientists view feature contrasts as important when they
pop out of cluttered backgrounds, in the way that a single red letter on a page of black text would pop-out (
i.e., draw attention [
9]). Because red is unique in the display it is readily distinguished from the surrounding black text. Therefore, it is somewhat surprising that a symmetrical target does not pop out of a display of non-symmetrical items that are otherwise identical to the target; the time required to detect a symmetrical item embedded in a background of non-symmetrical items increases rather steeply with the number of non-symmetrical items [
10,
11]. Therefore, the symmetry/non-symmetry contrast does not seem important (special) to the visual system. Similarly, symmetry does not seem to serve an alerting function because it is extremely difficult to detect in peripheral vision when embedded in non-symmetrical noise [
12]. In this paper we extend our explorations of sensitivity to symmetry across the visual field.
A great deal of study has focused on our sensitivity to
isolated patches of symmetry presented to different positions in the visual field. It is clear to anyone possessing a normally functioning visual system that sensitivity to spatial structure decreases as stimuli are moved from fixation into the visual periphery. (Distance from fixation is called “eccentricity” and measured in degrees of visual angle.) It has long been known that in many cases stimulus magnification is sufficient to compensate for eccentricity-dependent sensitivity loss and that the required magnification is often a linear function of eccentricity [
13]. Studies of eccentricity-dependent loss of sensitivity to isolated symmetrical targets have shown that stimulus magnification is sufficient to compensate for this loss [
14,
15,
16]. The rate at which stimuli need to be magnified (
M) with eccentricity is characterized by
k such that
M = 1 +
kE, where
E is eccentricity in degrees visual angle. This research shows that perception of symmetry at fixation is not qualitatively different from perception of symmetry in the periphery in the sense that eccentricity-dependent stimulus magnification, using
, with
k ≈ 1.25, seems sufficient to equate sensitivity to symmetry across the visual field. (However, there is very interesting work on symmetry and peripheral vision that deviates from this simple model [
17]). This gradient (
k ≈ 1.25) has been found in many other studies for a variety of stimuli [
18,
19,
20,
21] and should not be viewed as exceptional in any way.
In addition to simple eccentricity-dependent resolution loss (requiring stimulus magnification to equate performance across the visual field), neighboring (henceforth
flanking) items impair discrimination of targets that are well above the resolution limits of the visual system. Such interference effects are known in the vision science literature as
crowding. Crowding by flanking stimuli increases with eccentricity [
22,
23] and is arguably non-existent at fixation [
24]. Furthermore, crowding regions (the regions within which flankers impair target detection or discrimination) are elliptical and oriented towards the fovea [
23]. This means that when a target is presented on the horizontal meridian, flankers placed to the right and left of the target (
i.e., on a line connected to the point of fixation) will have a more disruptive effect than when placed above and below the target (
i.e., perpendicular to the line connecting the target and the point of fixation).
We have noted that symmetry is difficult to distinguish from background clutter in peripheral vision [
11,
12] but these effects have not been studied in the context of crowding. Therefore, in this paper we ask how flankers affect the perception of symmetry across the visual field. Using a symmetrical target in a number of target/flanker configurations we investigate whether symmetry does indeed exhibit the common characteristics of crowding such as: (i) the decrease or absence of effect at fixation; (ii) increasing magnitude of spatial interference in the peripheral visual field; (iii) the presence of an anisotropy with respect to target/flanker configuration; and (iv) whether linear magnification factors are sufficient to characterize performance across the visual field.
Our approach to these questions relies on a method developed by Latham and Whitaker [
22]. Configurations are created with targets (symmetrical patches) flanked by non-symmetrical “flankers” at a range of separations that are proportional to target size. The size of an entire configuration is varied across trials until the target size eliciting a fixed level of discrimination accuracy is obtained. At this point we have, simultaneously, the target size at threshold and the target/flanker separation at threshold.
Recently we have shown (using other types of stimuli) that target size at threshold increases at a modest rate in the absence of flankers [
25]. The presence of flankers increases the rate at which target size-thresholds increase with eccentricity, and the smaller the separation the greater the rate of size-threshold increase with eccentricity [
22,
23,
25]. For the smallest possible target/flanker separations, the rate at which size thresholds increase with eccentricity is far greater than the rate of increase in the unflanked condition. Recent work has shown that when targets are distinguished from flankers by a feature such as color, orientation or spatial frequency then the flankers have little effect on sensitivity to the target [
26,
27]. Therefore, if symmetry were special to the visual system in this way we would expect to see size thresholds increase at about the same rate in the presence of flankers as in their absence. However, as noted, previous results have shown that mirror symmetry does not pop out of non-symmetrical distractors in a visual search task [
10,
11] and that it is difficult to detect when surrounded by noise in the periphery. These results suggest that symmetry/non-symmetry contrasts are not special to the early visual system and so we expect to see size thresholds increase at a much faster rate in the presence of flankers than in their absence [
25].
3. Results
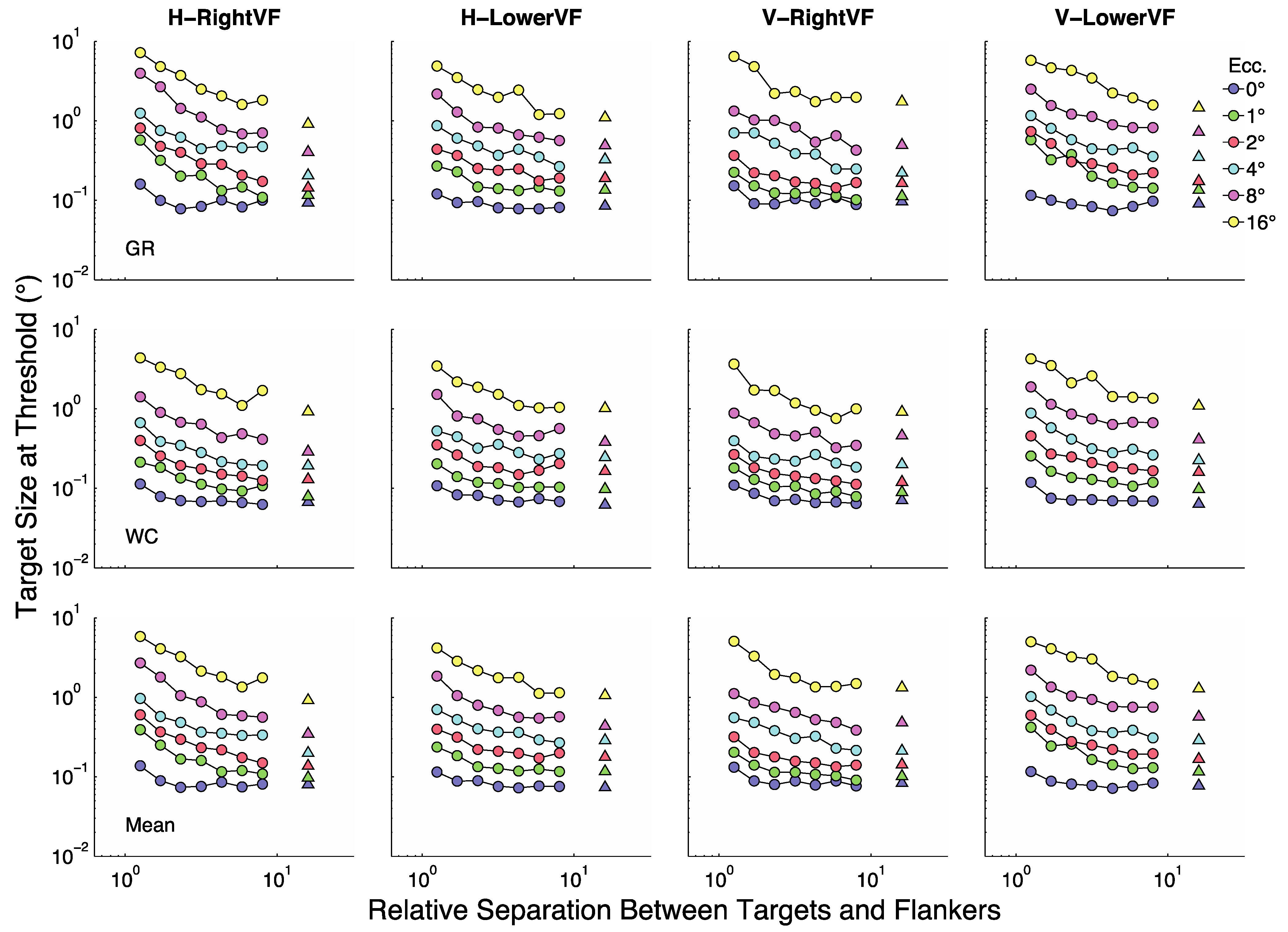
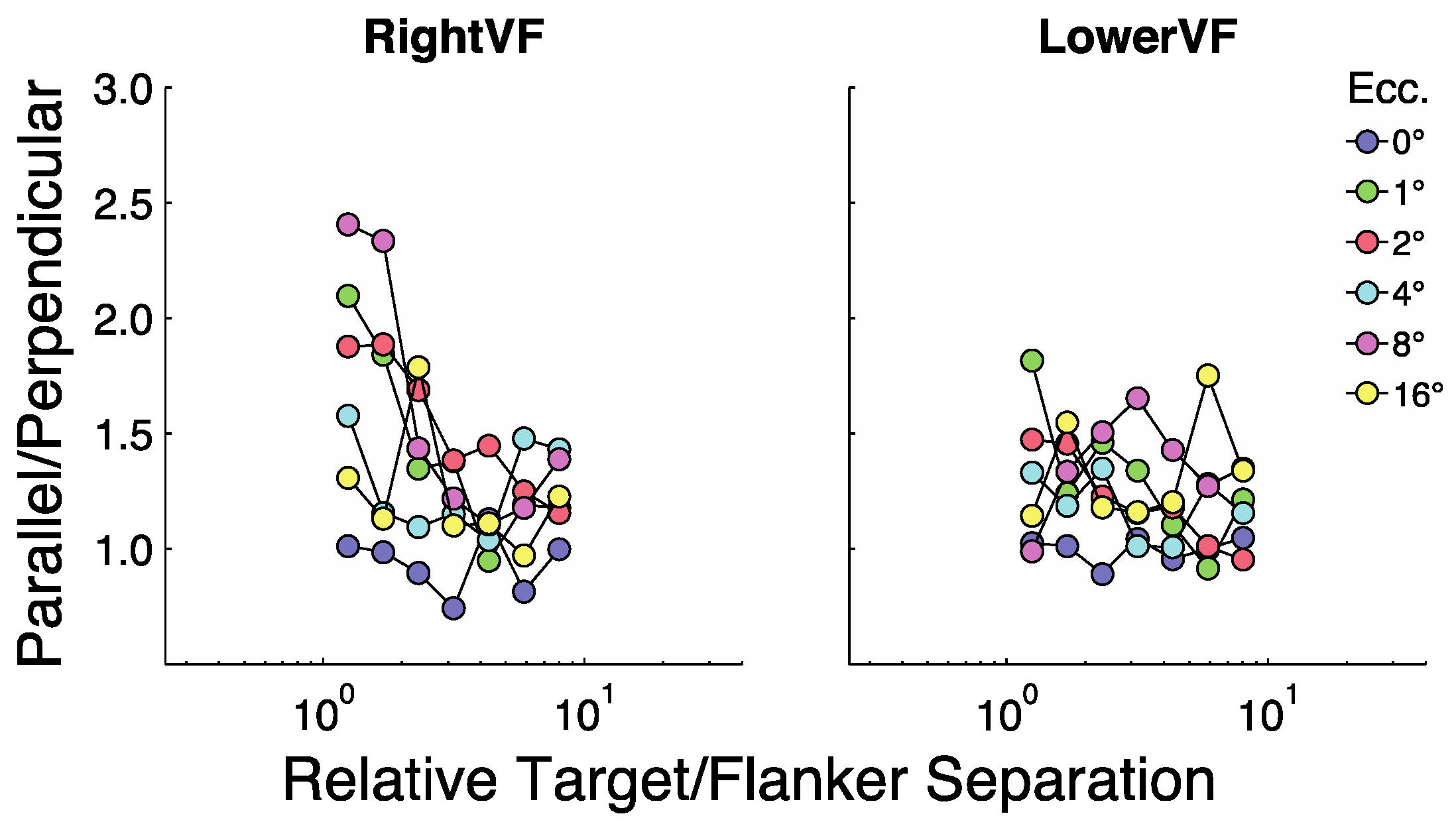
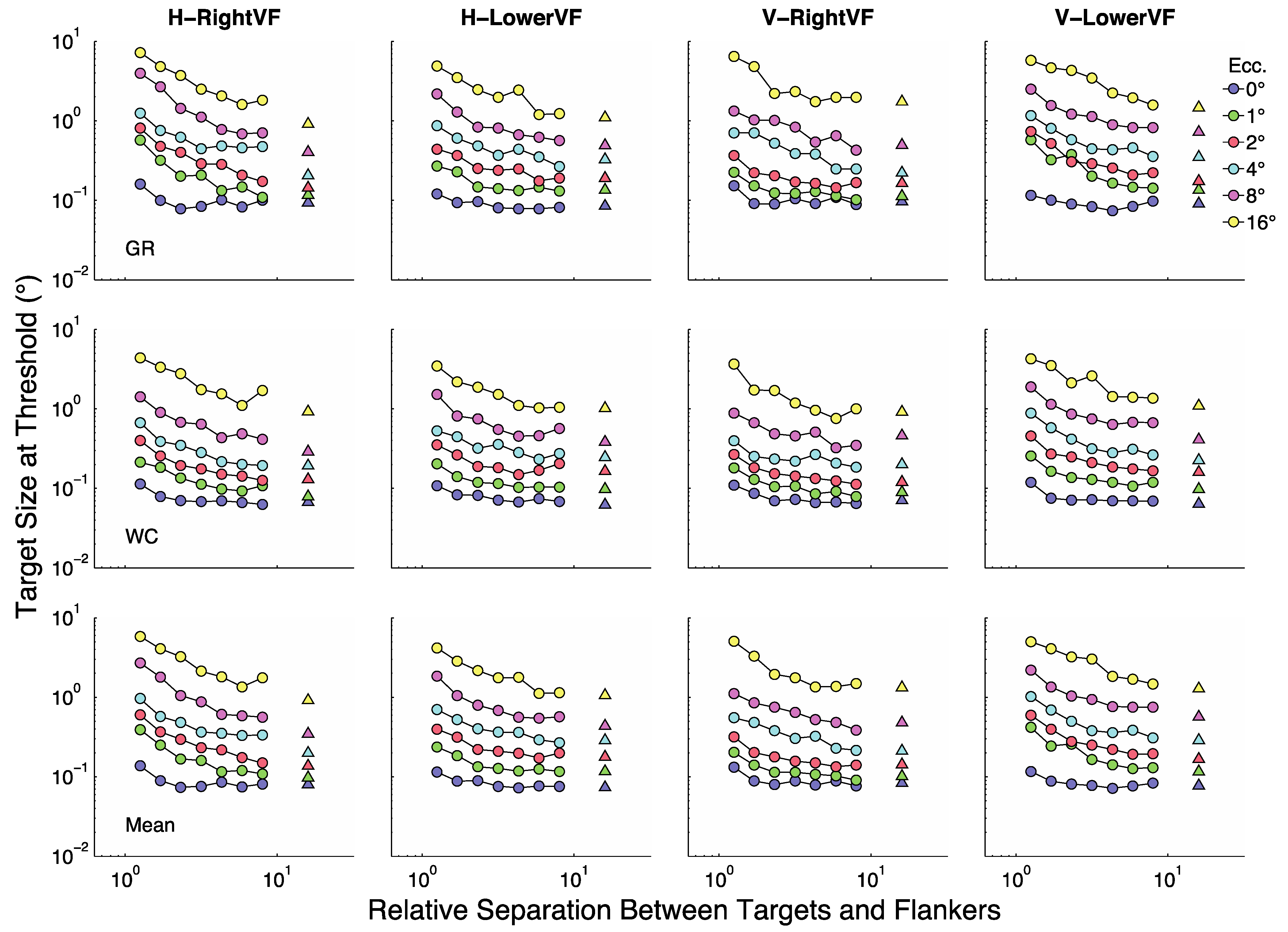
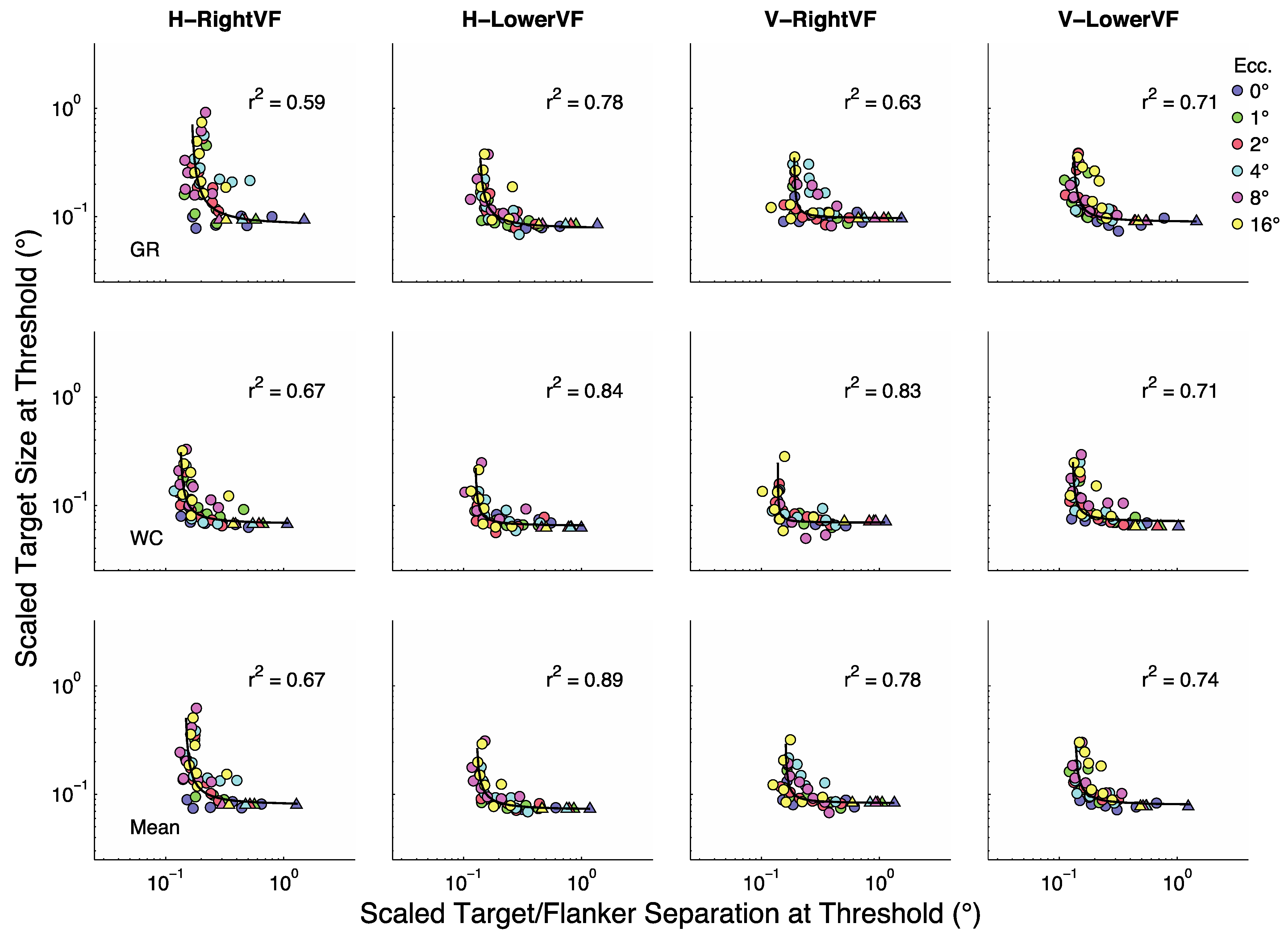
Figure 2 summarizes the results for the two participants (rows 1 and 2) as well as their averaged results (row 3). In each panel the y-axis represents target size (in degrees visual angle) at threshold and the X-axis represents target/flanker separation in multiples of target size (relative separations). Each line (color) represents a given eccentricity as shown in the legend. The four panels in each row show results for the horizontal (H) and vertical (V) flankers when presented in the RightVF or LowerVF.
Figure 3 replots the same data with the x-axis as actual separation (in degrees visual angle). Actual separation is obtained by multiplying target size at threshold by the relative separation. The colored circles represent conditions in which the relative separations were 1.25 to 8× target size. Triangles represent the unflanked condition (plotted at 16 times target size in
Figure 2 and
Figure 3).
Figure 3 shows that at fixation (blue symbols, 0°) size thresholds are largely independent of target-flanker separation for most separations, with modest threshold elevation (increases over the unflanked-condition) at the two smallest separations. At all other eccentricities threshold elevation is considerably greater at the smallest sizes and decreases as separation increases until asymptote is reached at the largest separations. Roughly speaking there are two major parts to most non-foveal curves in
Figure 3. At small separations the curves tend to asymptote parallel to the y-axis and at large separations they tend to asymptote parallel to the X-axis. Sections of a curve parallel to the y-axis indicate points at which crowding is independent of target size. That is, the centre-to-centre spacing between targets and flankers does not change with changes in target size. Sections of a curve parallel to the X-axis indicate points at which the flankers have no influence on size thresholds. That is, size thresholds are unchanged as the centre-to-centre spacing between targets and flankers varies.
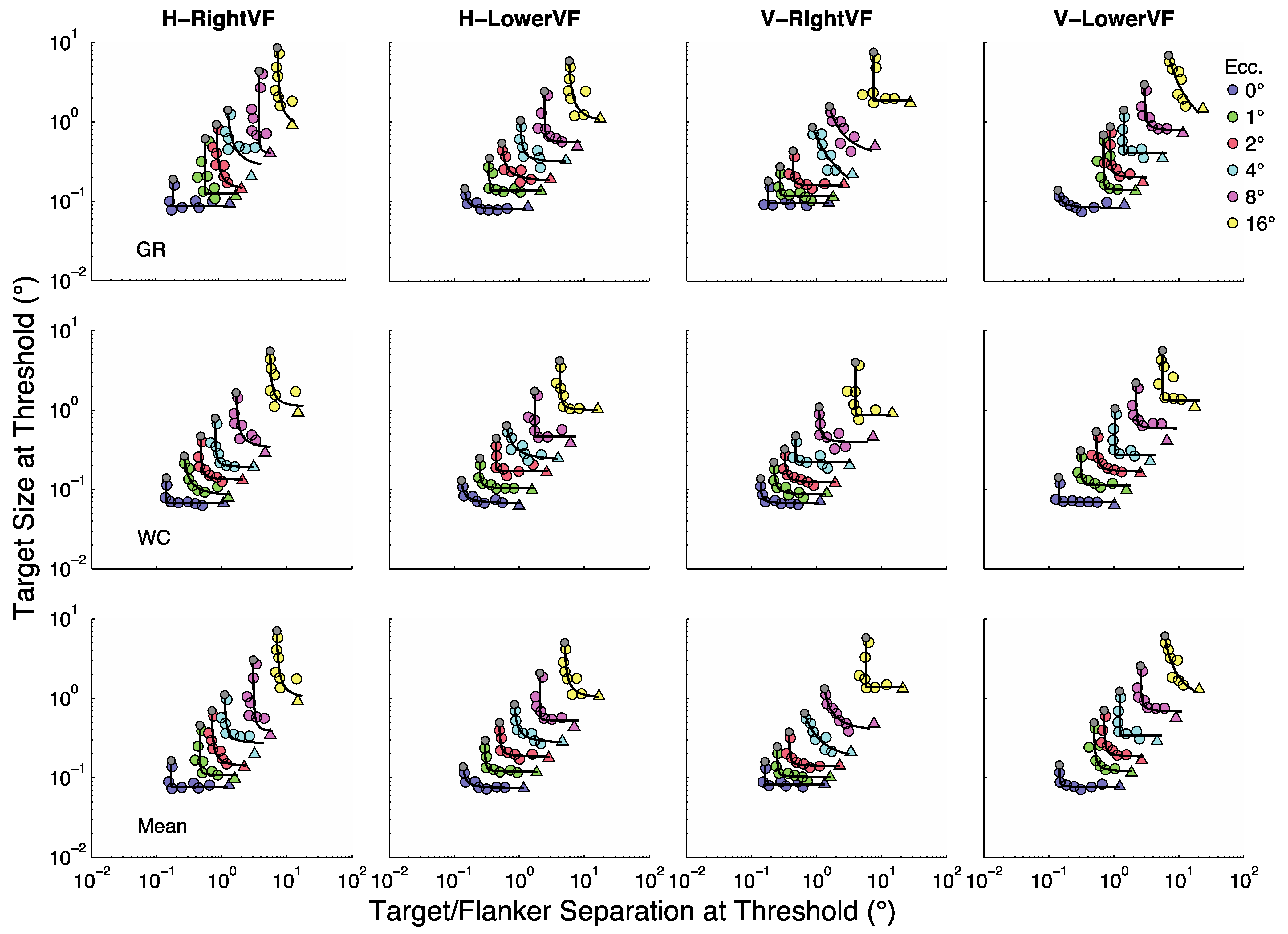
The general form of the functions in
Figure 3 is well captured by a rectangular parabola [
31], as defined in Equation 1,
In this case
represents the limiting size-at-threshold obtained when target/flanker separation increases towards infinity (
i.e., the unflanked condition). In other words,
represents the stimulus size at which the curve becomes parallel to the separation axis. And,
represents the limiting separation-at-threshold obtained when target/flanker separation decreases towards 1 (
i.e., the smallest centre-to-centre spacing for which targets and flankers do not overlap). In other words,
represents the target/flanker separation at which the curve becomes parallel to the size axis. The best fitting rectangular parabola for each curve in
Figure 3 is shown as a continuous black line.
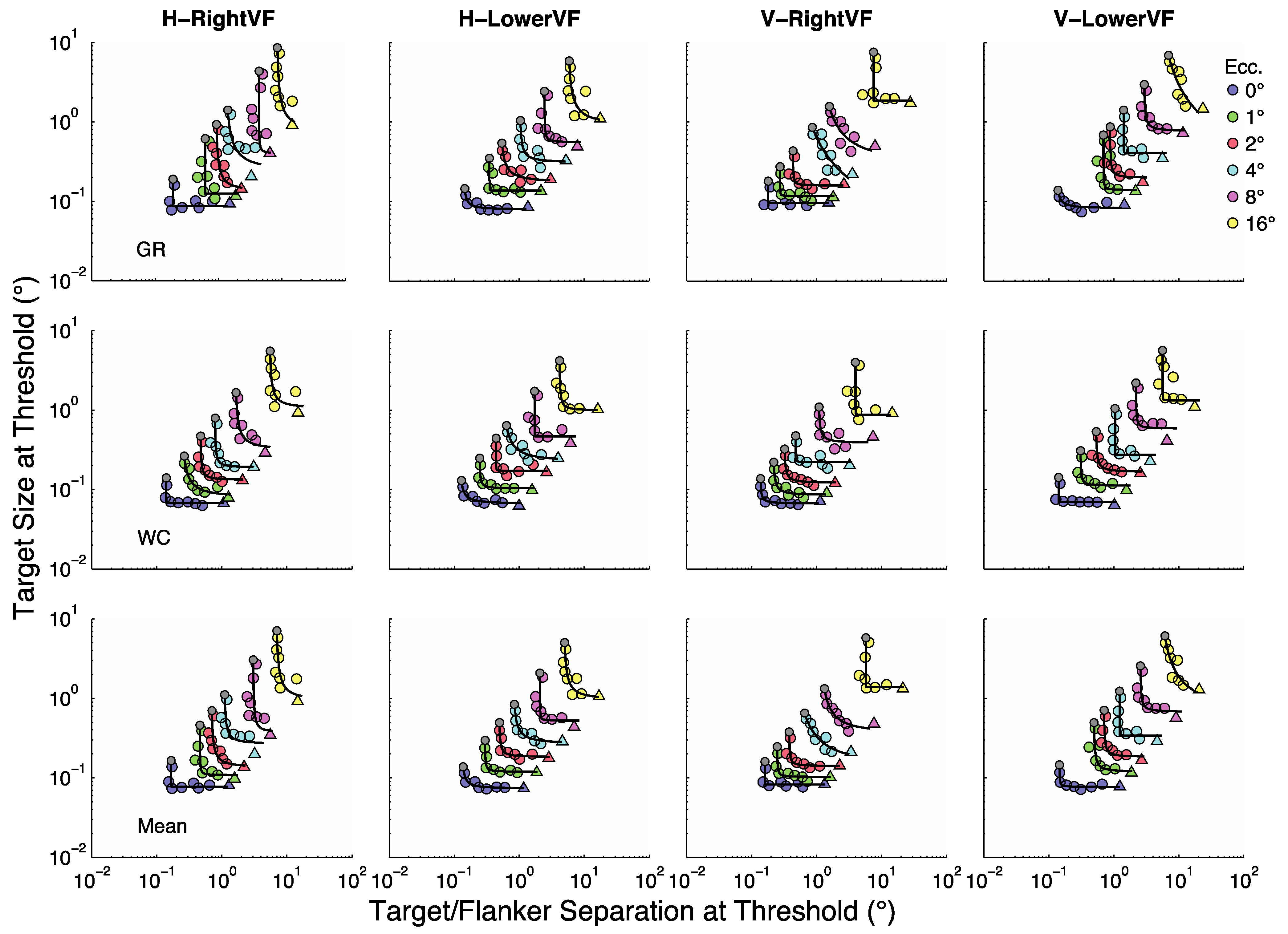
Our objective is to characterize how stimulus size and target/flanker separation combine to determine thresholds across the visual field. A first step in this direction is to consider the limiting cases;
i.e., the unflanked thresholds and the smallest possible target/flanker separations. The blue circles in
Figure 4 show size thresholds at 1 to 16° relative to size thresholds at fixation;
i.e.,
. The green circles in
Figure 4 show the points on the rectangular parabolas at 1 to 16° for which size at threshold (
μsize) = separation at threshold (
μsep);
i.e.,
μsize /
μsep = 1. In other words, the green circles in
Figure 4 represent the sizes (or separations) corresponding to the grey dots in
Figure 3 at eccentricities of 1 to 16° divided by the sizes (or separations) corresponding to the grey dots at fixation (0°); i.e.,
μE/
μ0. These limiting cases (
and
μE/
μ0) show how the rectangular parabolas in
Figure 3 shift up and to the right with eccentricity, respectively, within each panel.
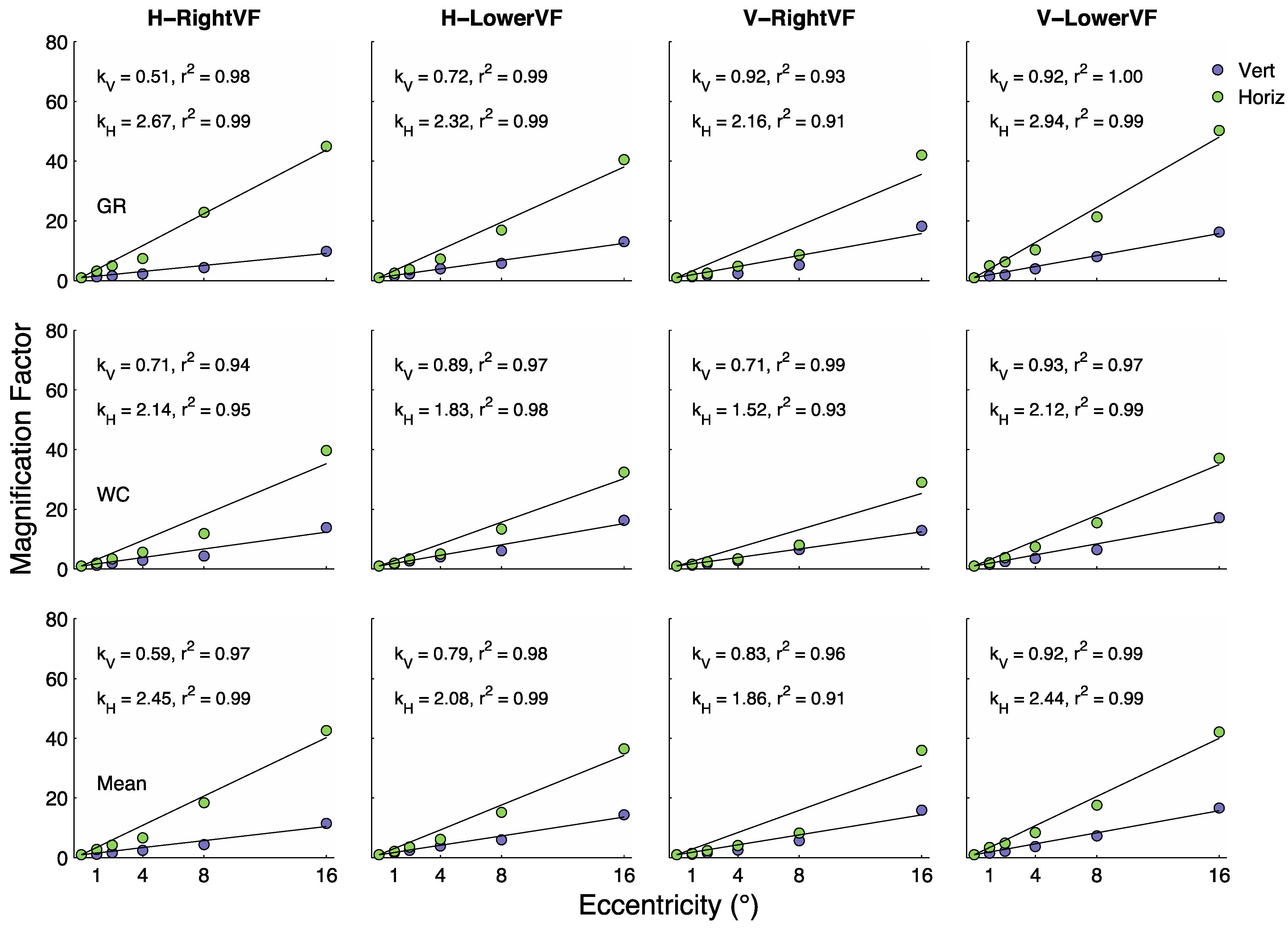
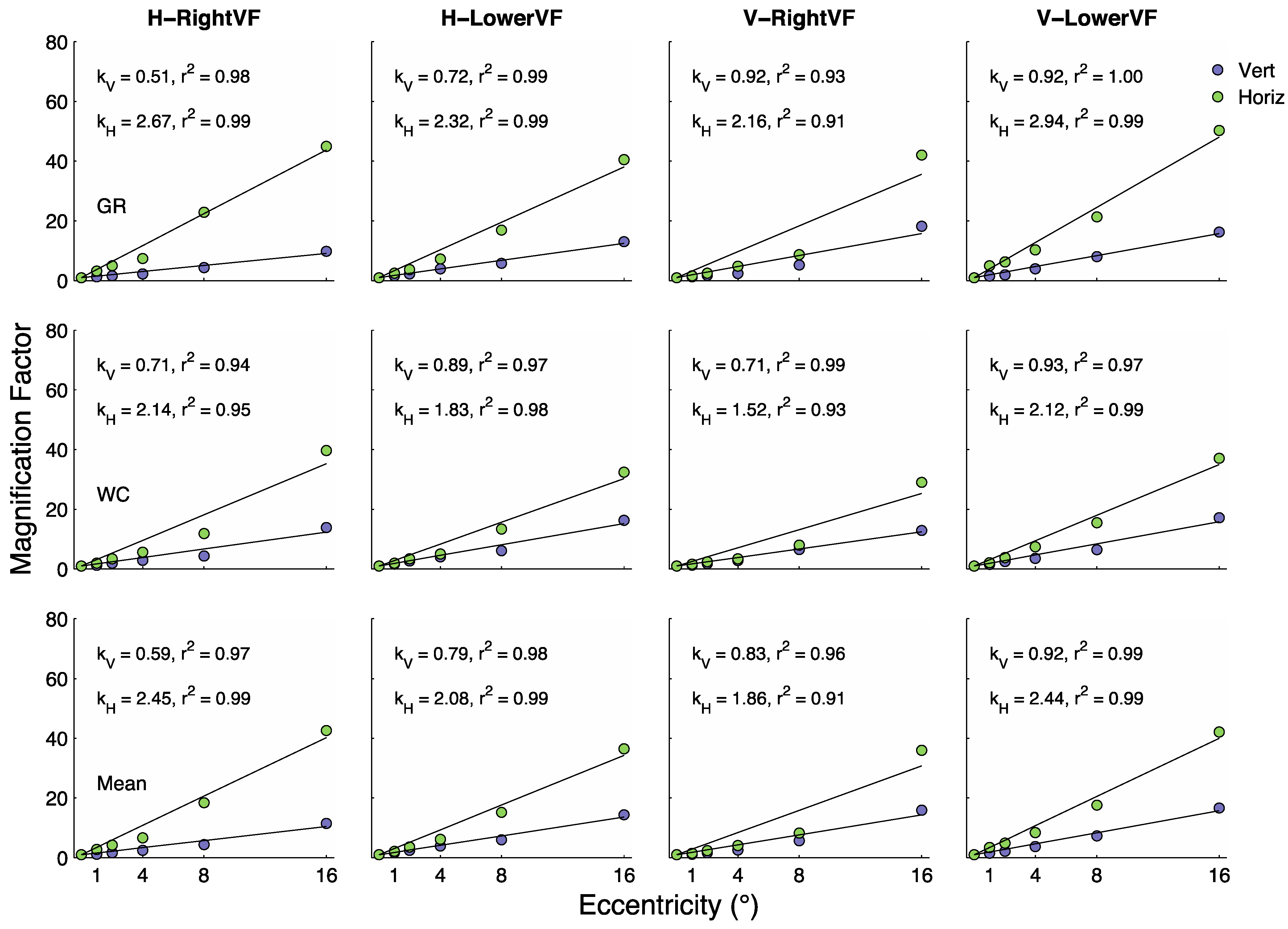
In each panel of
Figure 4 the blue and green data points have been fit with a function of the form
. The values of
and
and the proportion of variability (
) in the data explained by the lines are shown within each panel. In general the fits are quite good, explaining, on average, about 97% of the eccentricity-dependent variability in both
and
μE/
μ0. The y-axis in
Figure 4 is labeled “Magnification Factor” because the curves represent the degree to which the functions shift up and to the right with eccentricity.
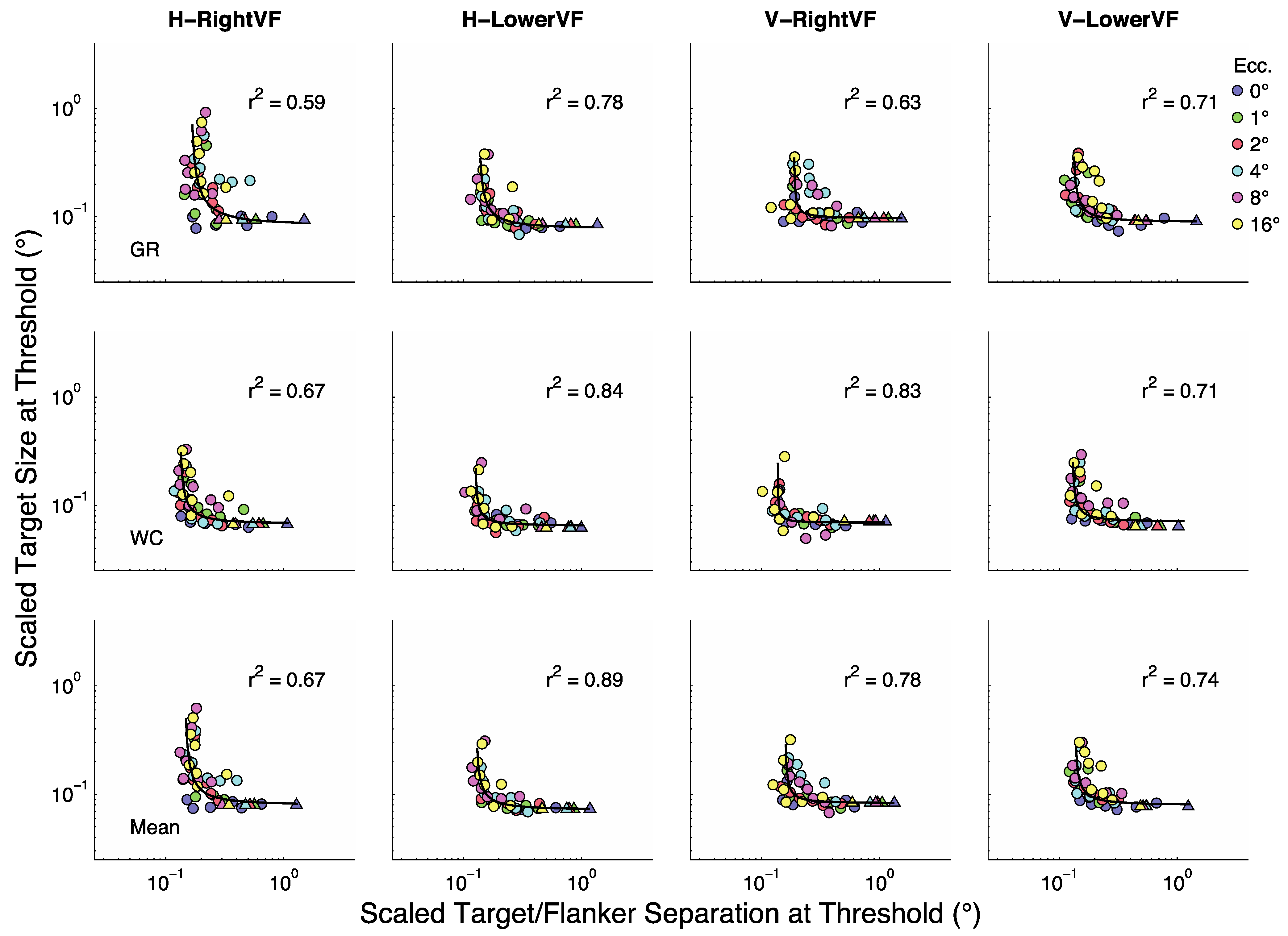
In the large literature on size scaling attempts are made to shift data from all eccentricities onto a single curve in order to provide a concise characterization of the data. For the present data set it is clear that the data would have to be shifted both down and to the left. We used three shifting methods in an attempt to find a simple characterization of the data. First, for each data point in
Figure 3 we divided separation at threshold by
μE/μ0 and the corresponding size at threshold by
. Once the data were shifted downward and leftward in this way, we found the best fitting rectangular parabola to the scaled data.
Figure 5 shows the shifted data along with the best-fitting rectangular parabola. Also shown is the proportion of variability that the parabola explains. (Please note that our measure of fit is based on the diagonal distance from data point to curve in the log-log space shown [
31]). Generally the fits were reasonably good and explained 72% of the variability in the data on average.
A second analysis was performed to see if a more concise characterization of the data could be achieved by replacing μE/μ0 with and with . Once the data were scaled we found the best fitting rectangular parabola. The results are not shown but they explained only 55% of the variability in the data on average, and are therefore substantially worse than the first analysis, which used μE/μ0 and to scale the data. Therefore, in spite of the seemingly nice fits of to μE/μ0 and to (r2 ≈ .97) these approximations leave a great deal of residual variability about the rectangular parabola fit to the scaled data.
A third analysis was performed to see if a better fit could be achieved by replacing linear magnification functions and with non-linear magnification functions M = 1+EαH/βH and M = 1+EαV/βV respectively. We thought these non-linear functions might better fit μE/μ0 and , respectively, and thus produce scaled data that are a better fit to a rectangular parabola. The obtained results explained 63% of the variability in the data and are thus substantially better than the second fit, yet still worse than for the original “raw fits”.
From these analyses we can conclude that the most effective scaling method shifts the curves down and to the left by dividing size at threshold by and separation at threshold by μE/μ0. It is disappointing that we are unable to find low-parameter fits to these scaling factors that are equally effective. It should be kept in mind, however, that the values reported above do not include variability attributable to eccentricity. That is, the values account only for the variability of the scaled data about the best fitting function. If the fits to the scaled data were “unscaled” by multiplying scaled size at threshold by and scaled separation at threshold by μE/μ0, the resulting fits to the original data would have produced values that exceed .97, on average, in all cases.
Finally, a number of reports in the crowding literature have shown that crowding is most effective when targets and flankers are aligned with the line that connects them with the point of fixation [
23,
32];
i.e., horizontally positioned flankers in the RightVF and vertically positioned flankers in the LowerVF. We were able to assess this trend in our data by comparing size at threshold in cases of flankers and targets forming a line connected to fixation to those forming a line perpendicular to this line. In the RightVF this means dividing size thresholds obtained with horizontally oriented flankers by the size thresholds obtained with vertically oriented flankers and in the LowerVF, dividing size thresholds obtained with vertically oriented flankers by the size thresholds obtained with horizontally oriented flankers. These results are shown in
Figure 6 for the averaged data. Threshold ratios are shown for each separation and eccentricity in the RightVF and LowerVF. It is notable that in almost all cases the ratios exceed 1, meaning that flankers are more effective when aligned with fixation.
4. Discussion
In this study we asked how flanking items affect the discrimination of symmetry across the visual field. To do this we measured, simultaneously, stimulus size- and target/flanker separation-at threshold across a range of eccentricities in the RightVF and LowerVF. Size thresholds for isolated (unflanked) patches of symmetry (triangles in
Figure 3) increased linearly with eccentricity at a relatively modest rate.
Figure 4 shows
values ranging from 0.51 to 0.93, with a mean of
= .79 (SEM = .053). These results are generally consistent with several previous studies that asked the same question with somewhat different methods [
12,
14,
15,
16]. A report by Barrett
et al. [
14] found average values of
= 1.21, but within this average one subject had
= 0.97 and the other had
= 1.62. A study by Saarinen [
15] found that
= 1.29 accounted for the data and a study by Sally and Gurnsey [
16] found that
= 0.88 averaged over conditions and subjects; the range was 0.49 to 1.14. As noted, some of this variability may be attributable to subject differences. However, some of this variability may be attributable to the nature of symmetry itself. In most studies of symmetry, symmetrical patches are generated randomly on each trial. Therefore, subjects cannot perform the task through a template-matching strategy. The downside of this stochastic stimulus generation is that two nominally identical stimuli (e.g., stimuli of identical size) might actually differ in “perceptual salience”. This inherent variability makes it difficult to estimate thresholds reliably. In fact, the data reported in
Figure 3, for example, are more variable than comparable data we have obtained [
25] in similar tasks (letter identification, and grating [≡ ⫴] and “T” [⊤ ⊢ ⊥ ⊣] orientation discrimination) that do not have this stochastic characteristic.
Changes with eccentricity in
μsize (or
μsep) for which
μsize/
μsep = 1, reflect the changes in the maximum crowding effects with eccentricity;
i.e., greatest extent of the crowding region or greatest elevation of target size at threshold.
Figure 4 (green circles) shows that the magnitude of crowding increases with eccentricity at a faster rate than changes in size thresholds in the unflanked conditions. Using the best-fitting linear fits to these data gives a mean
= 2.21 (SEM = 0.16), which is about 2.8 times greater than
= 0.79. Therefore, in the case of symmetry discrimination we find that crowding effects increase with eccentricity at a greater rate than unflanked size-thresholds, as found in several previous studies employing different stimuli and tasks [
22,
23,
25]. Clearly, symmetry is not exempt from crowding effects.
We noted earlier that the evidence shows clearly that bilateral symmetry does not pop-out of complex displays [
10,
11]. In addition, evidence suggests that symmetry does not provide an alerting signal when surrounded by non-symmetrical noise [
12] in peripheral vision. The present study extends what is known about sensitivity to symmetry in the visual periphery by showing that symmetry is susceptible to interference from flanking items in peripheral vision. We have recently employed the methodology used here to examine crowding effects for letter identification, and grating and “T” orientation discrimination tasks. In all these cases the growth of interference regions with eccentricity (the horizontal shifts of the curves in
Figure 3; green symbols in
Figure 4) is much faster than the loss of resolution with eccentricity (the vertical shifts of the curves in
Figure 3; blue symbols in
Figure 4). However, tasks differ with respect to the relative rates at which the extent of interference zones and resolution limits change with eccentricity. For example, the horizontal magnification factor (
μE/μ0) for grating and T orientation discrimination tasks reach levels of 54 and 58, respectively at 16° in the LowerVF, whereas for letter discrimination this ratio reaches only 33 at 16°. In the present tasks, averaged over observers and conditions we find that the
μE/μ0 reaches 38 at 16°. Interestingly, we found
= 0.79 in the present symmetry task, 0.759 in the letter discrimination task and 0.54 and 0.57 in the grating and T orientation tasks. Therefore, in terms of these metrics the symmetry tasks studied here have more in common with the letter identification task than the grating and T orientation discrimination tasks.
The similarity of the symmetry results and the letter task (and their difference from the grating and T orientation tasks) may have something to do with the complexity of the decision to be made. We might have expected the symmetry tasks to have more in common with the grating and T orientation tasks, than the letter identification task. After all, we asked subjects to judge the orientation of the axis of symmetry. On the other hand, the perceptual complexity of the symmetrical stimuli may be more akin to the perceptual complexity of the letters. It may be that orientation discrimination for simple figures is a relatively simple task and hence the relatively slower rates of eccentricity-dependent sensitivity loss whereas the more demanding tasks (letter and symmetry discrimination) produce faster rates of eccentricity dependent sensitivity loss.
We found little evidence of crowding at fixation, however, there was some evidence of crowding at the smallest two relative separations (1.25, 1.70), consistent with previous findings that suggest the extent of spatial interaction extends over a very small range [
22,
24,
32]. The smallest relative separation (1.25) translates to a centre-to-centre spacing of 0.15° on average at fixation. As the crowding effect at fovea is modest at best and only for the two smallest distances between target and flanker it could be the result of internal blur given the complexity of our stimulus pattern [
24]. A consistent issue in the crowding literature is the near impossibility of making the stimuli at fixation (e.g., target and flankers) smaller than the internal blur while still being able to measure performance. As a result crowding and masking may get confused in the fovea because of the effects of blur, so that what looks like crowding is actually partly masking [
33].
Finally,
Figure 6 shows that in both cases target to fixation size thresholds are higher when the flankers are aligned (parallel) with the axis. This is in keeping with literature that suggests that crowding zones are elliptical and oriented to the fovea [
23,
34].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}