
Modelling EEG Dynamics with Brain Sources



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Brain Micro-States
1.2. Brain Sources
1.3. Dynamics Determined by Brain Source
2. Modeling EEG Dynamics with Brain Sources
2.1. Formulation of the Model
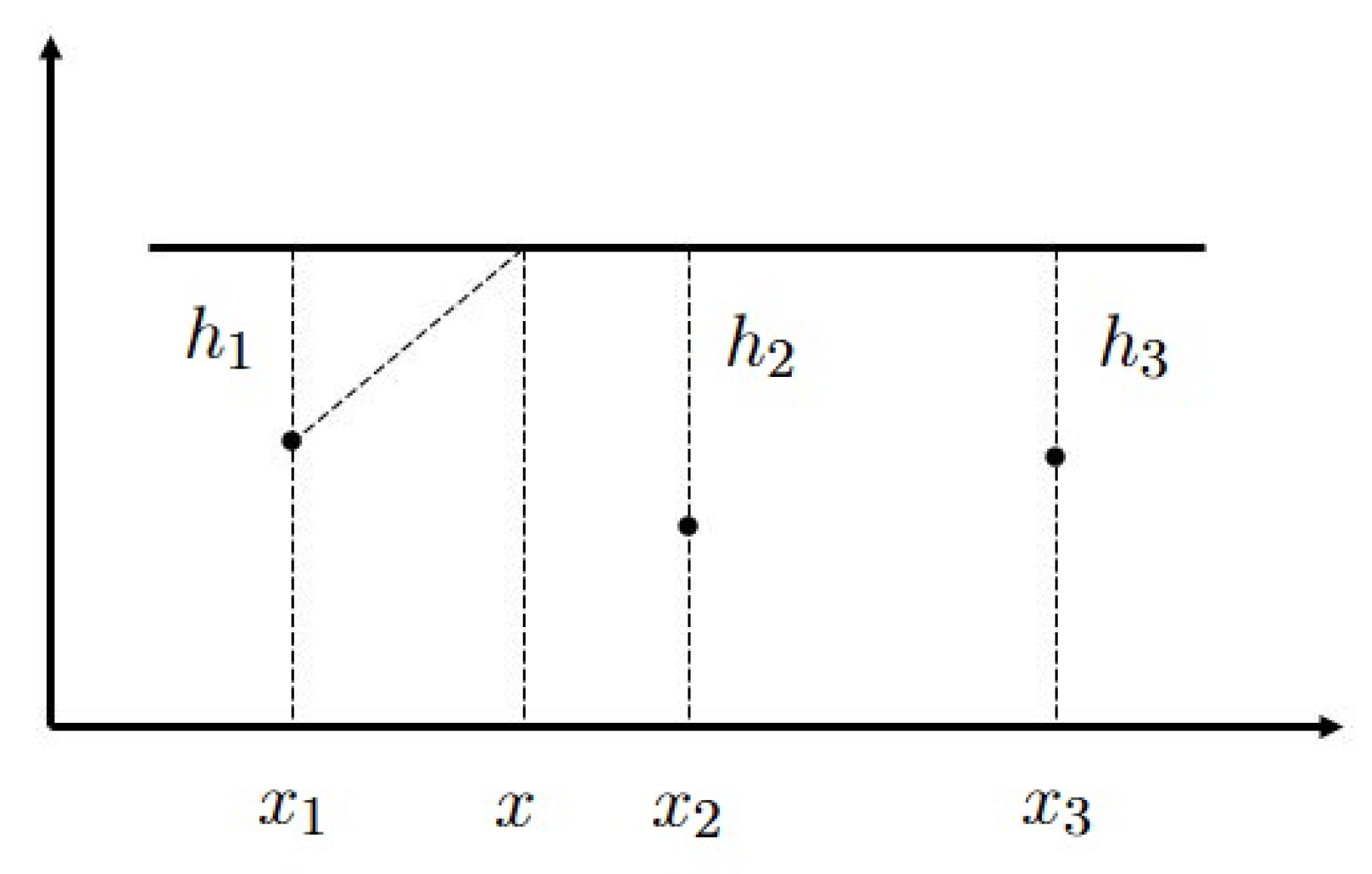
2.2. Analytical Approximation
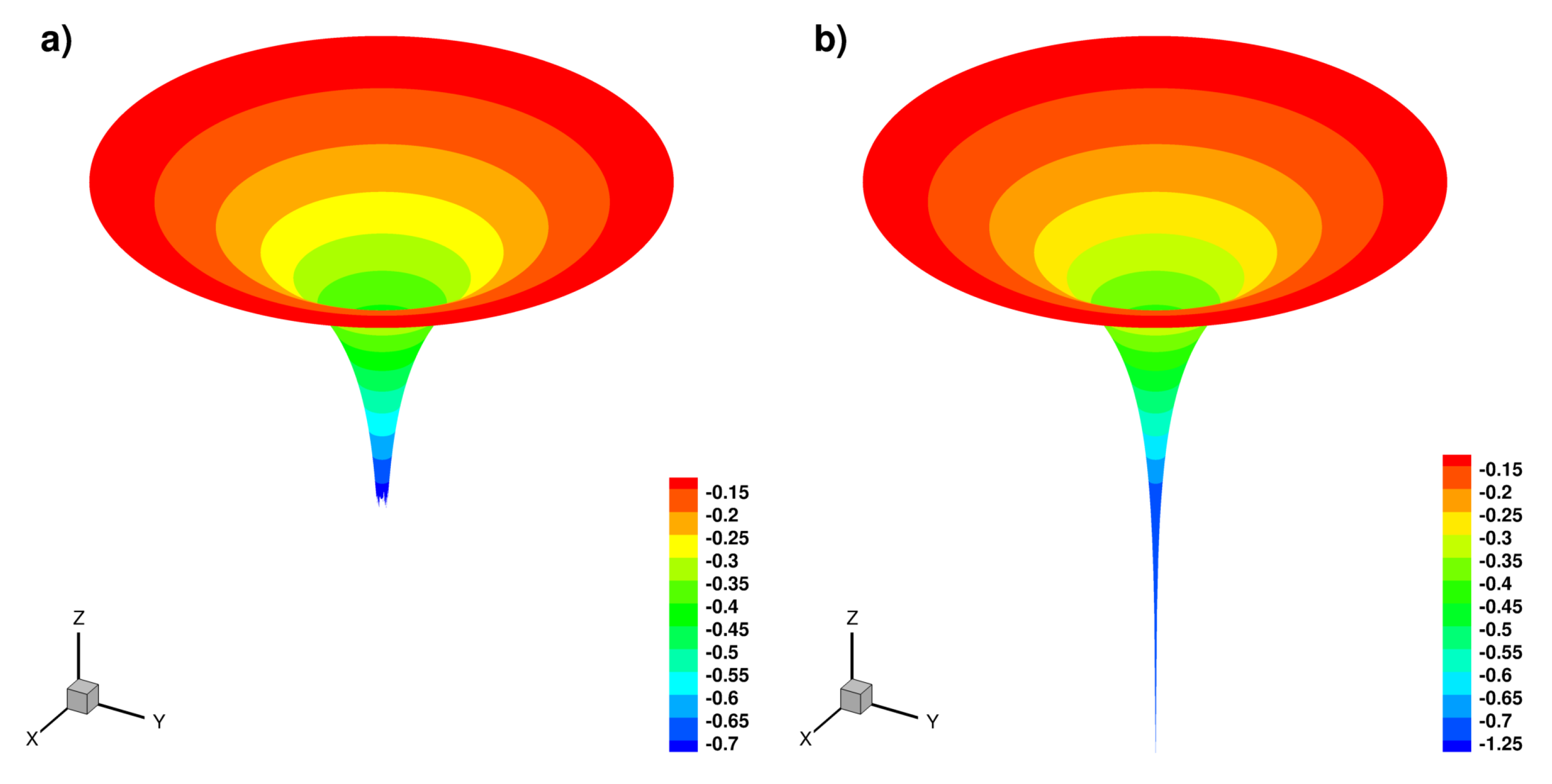
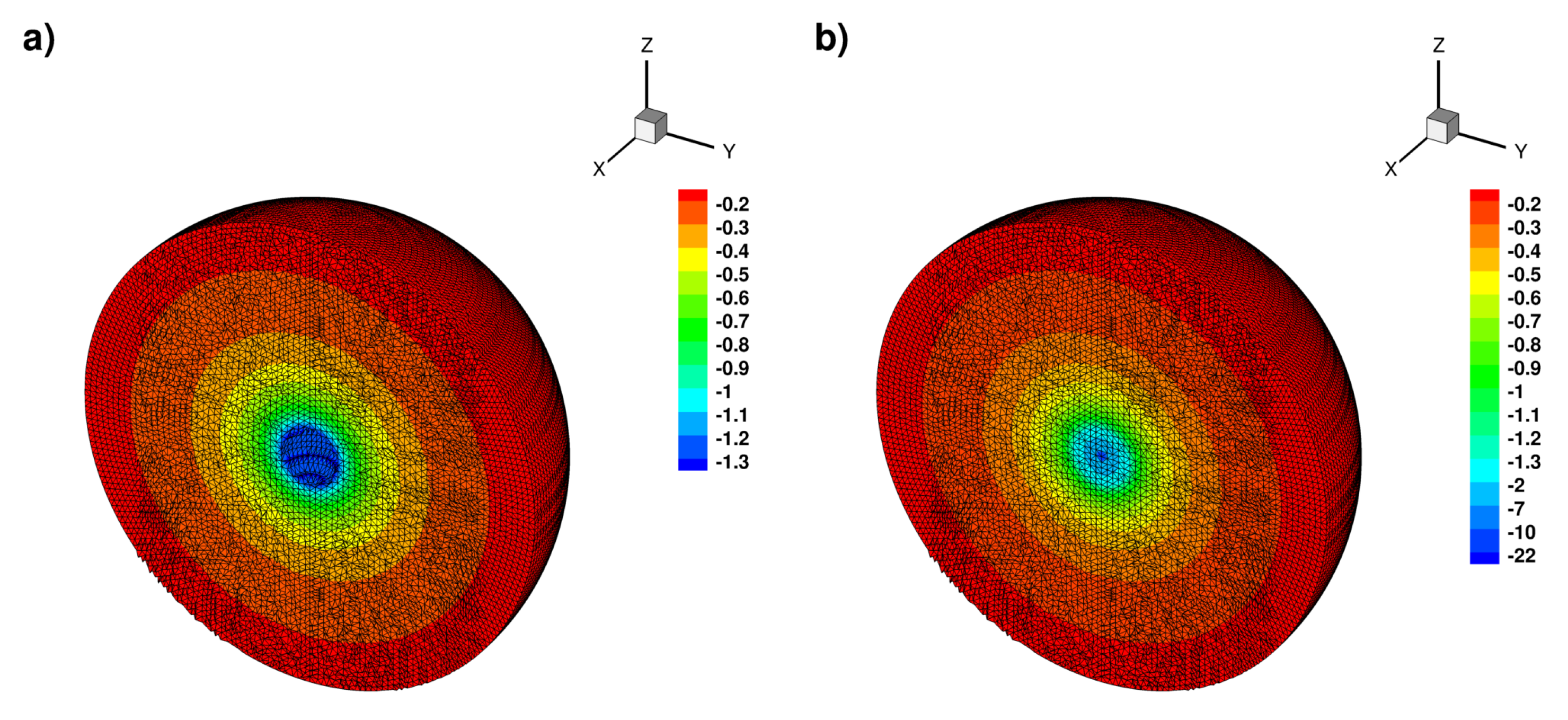
2.3. Dynamics on a Sphere
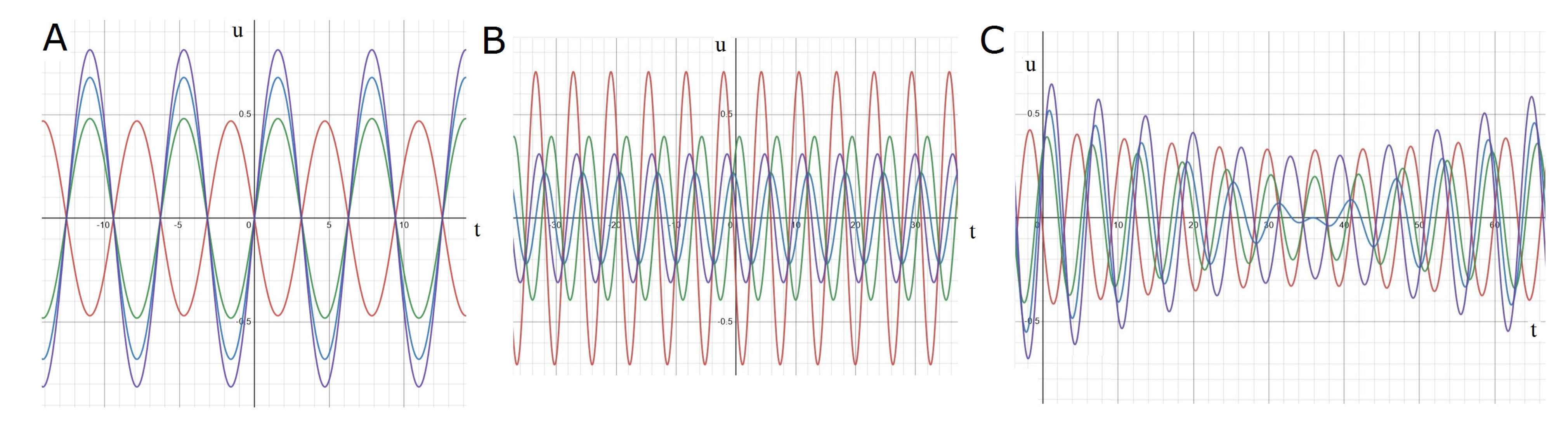
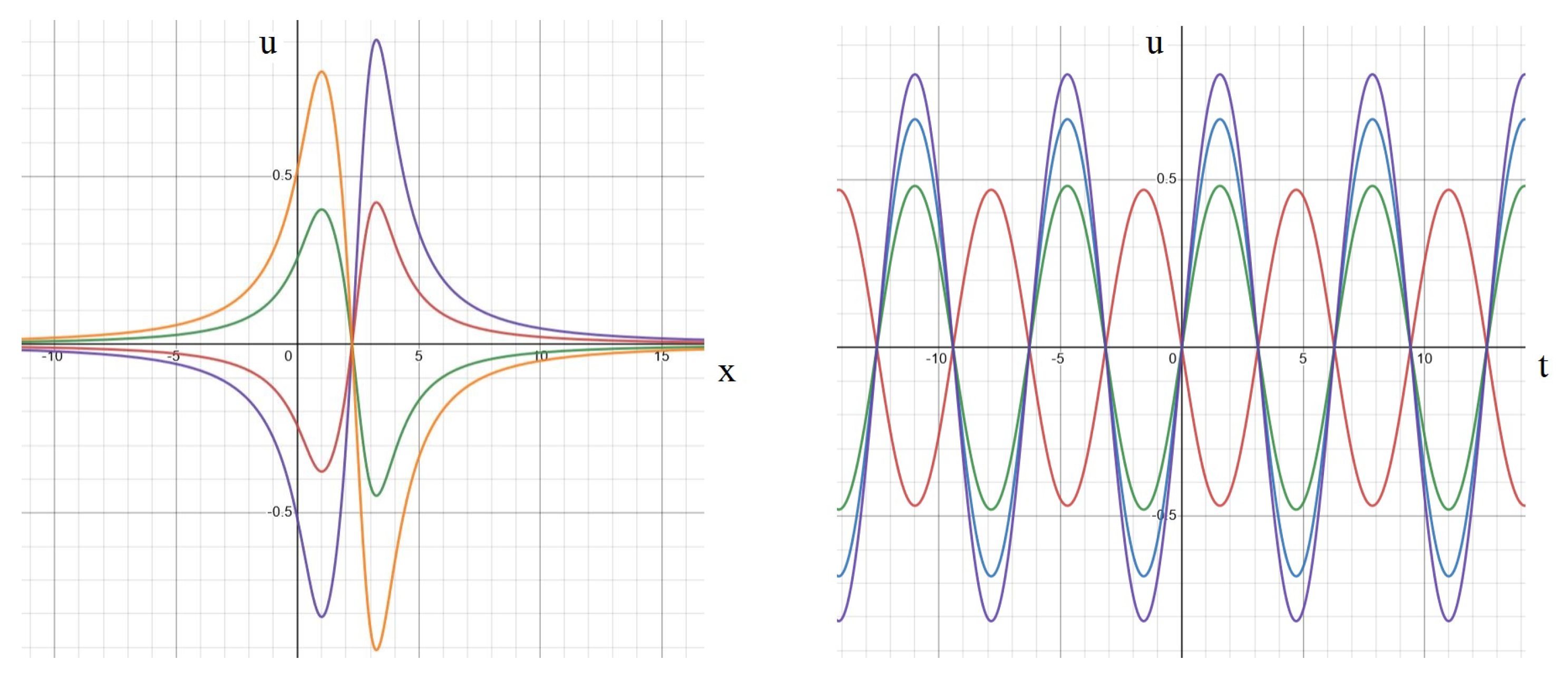
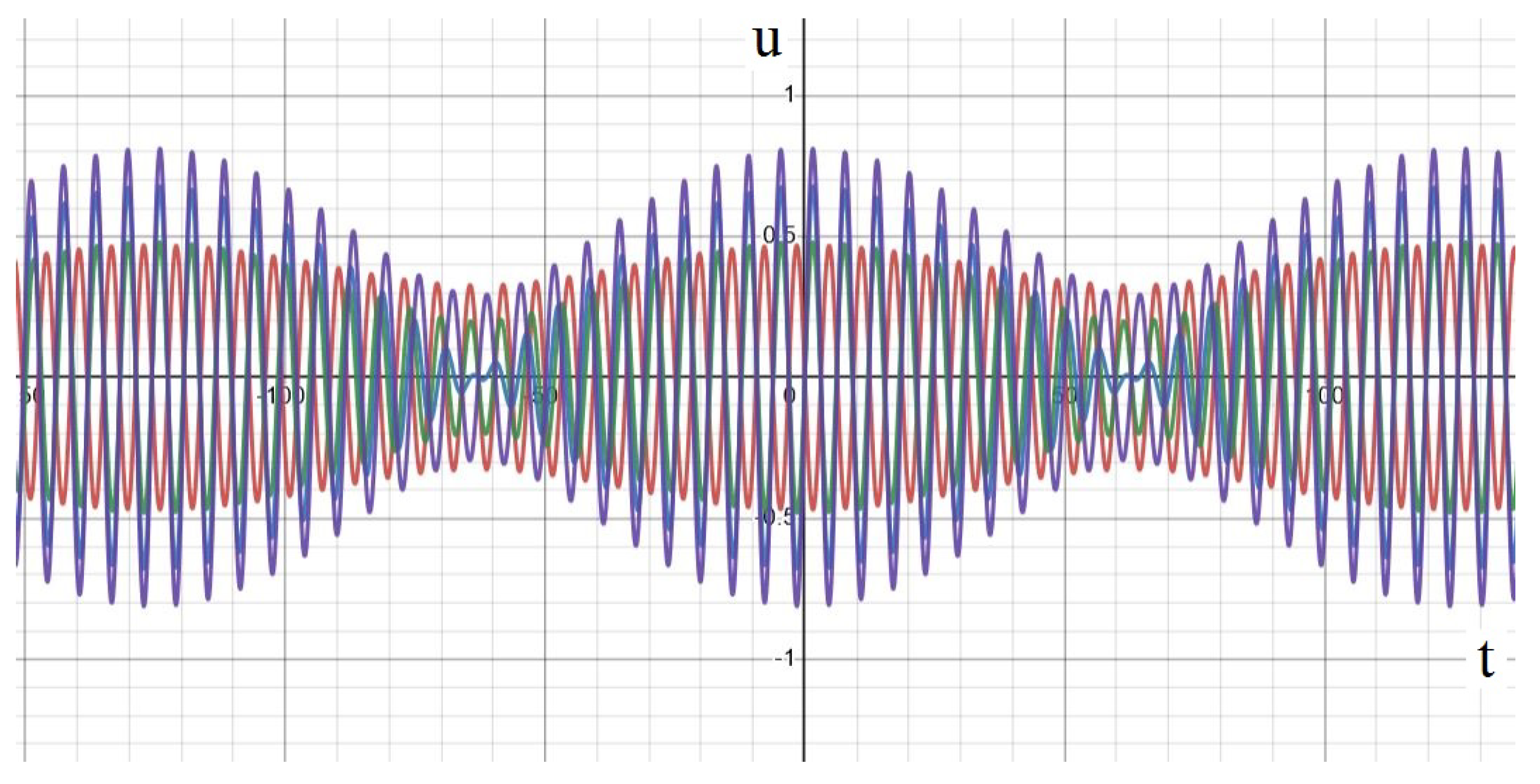
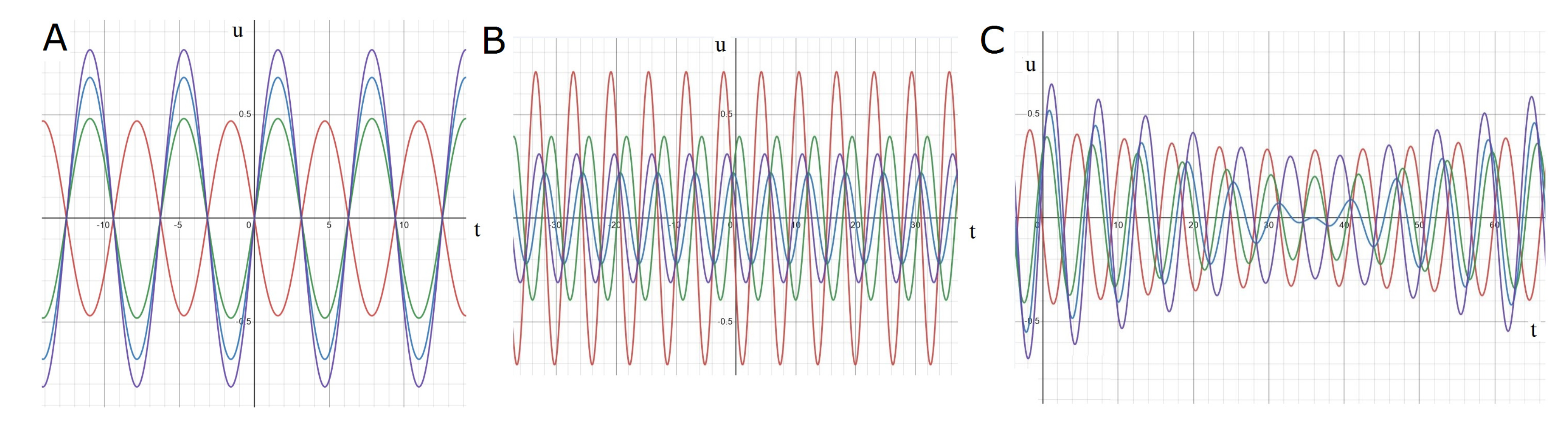
2.3.1. Standing Waves
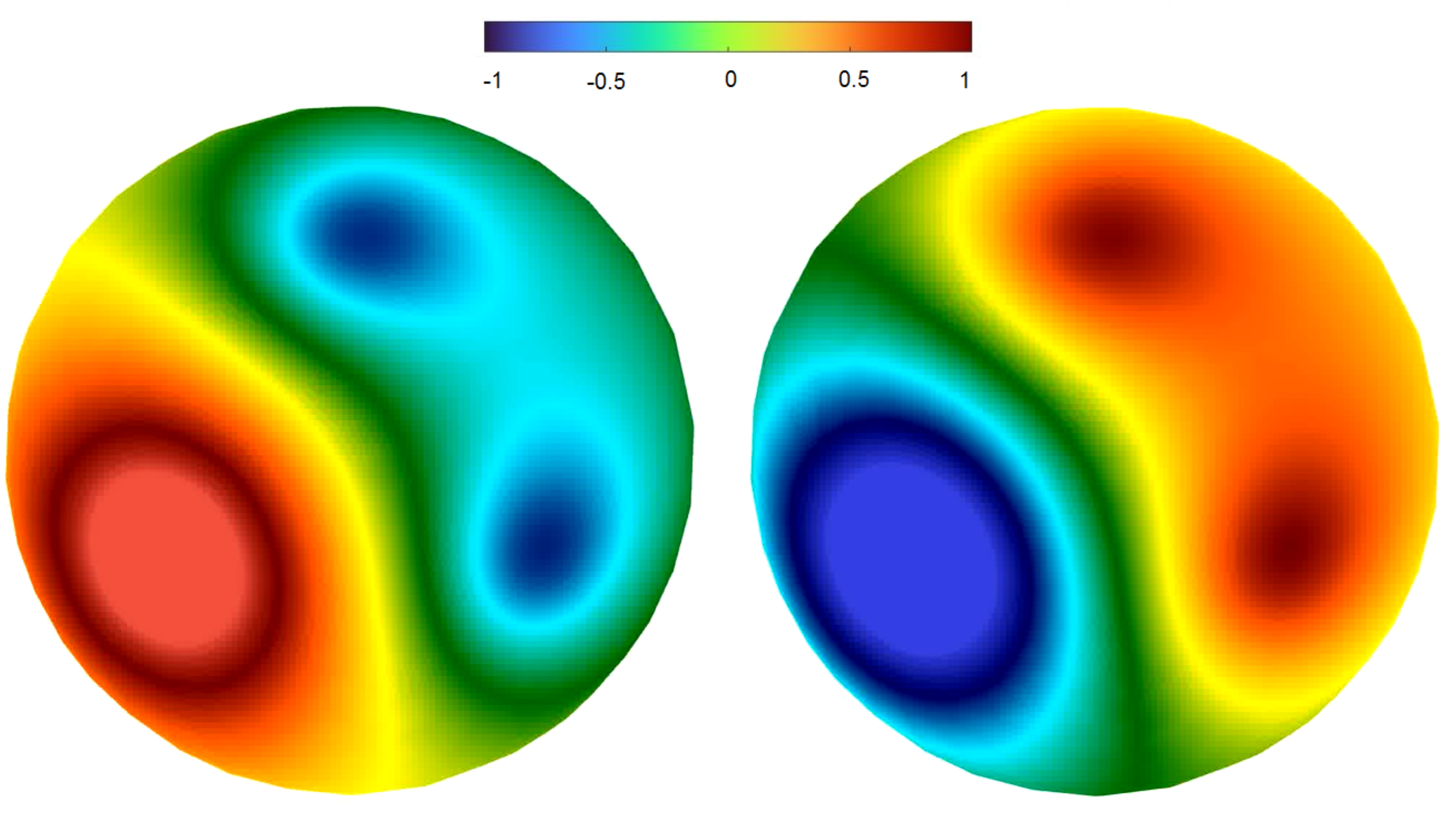
2.3.2. Rotating Regime
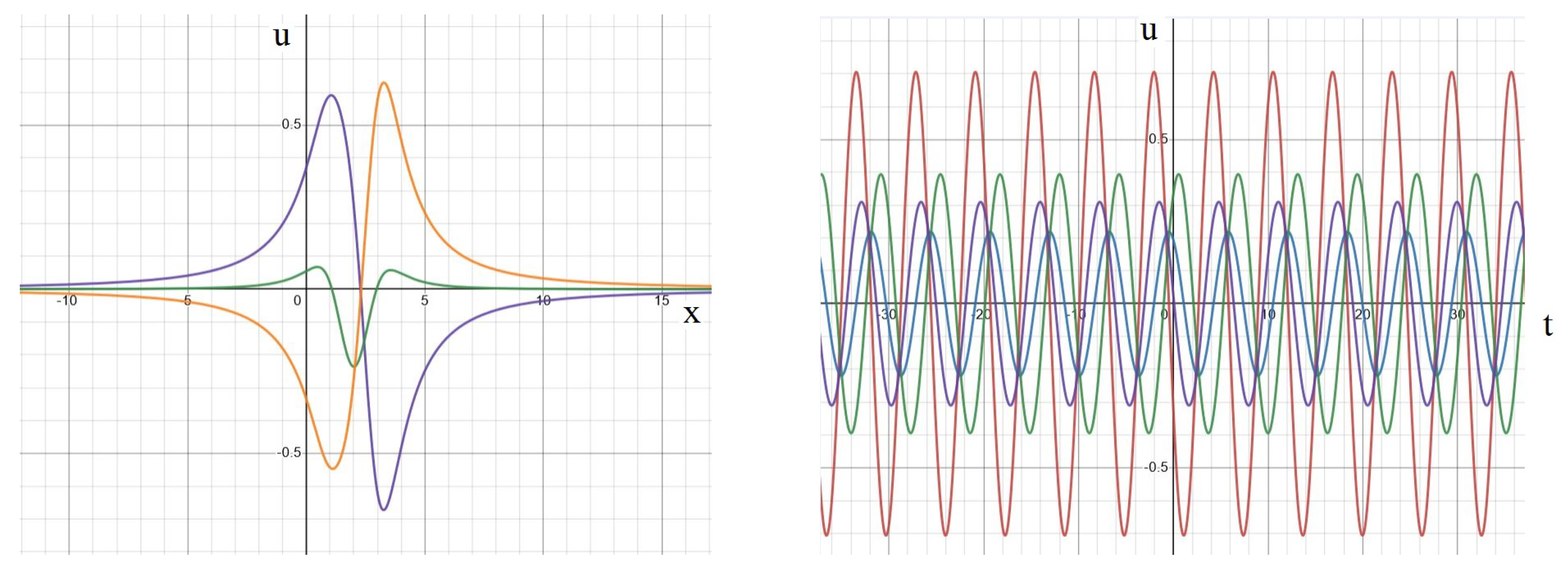
2.3.3. Symmetric Regime
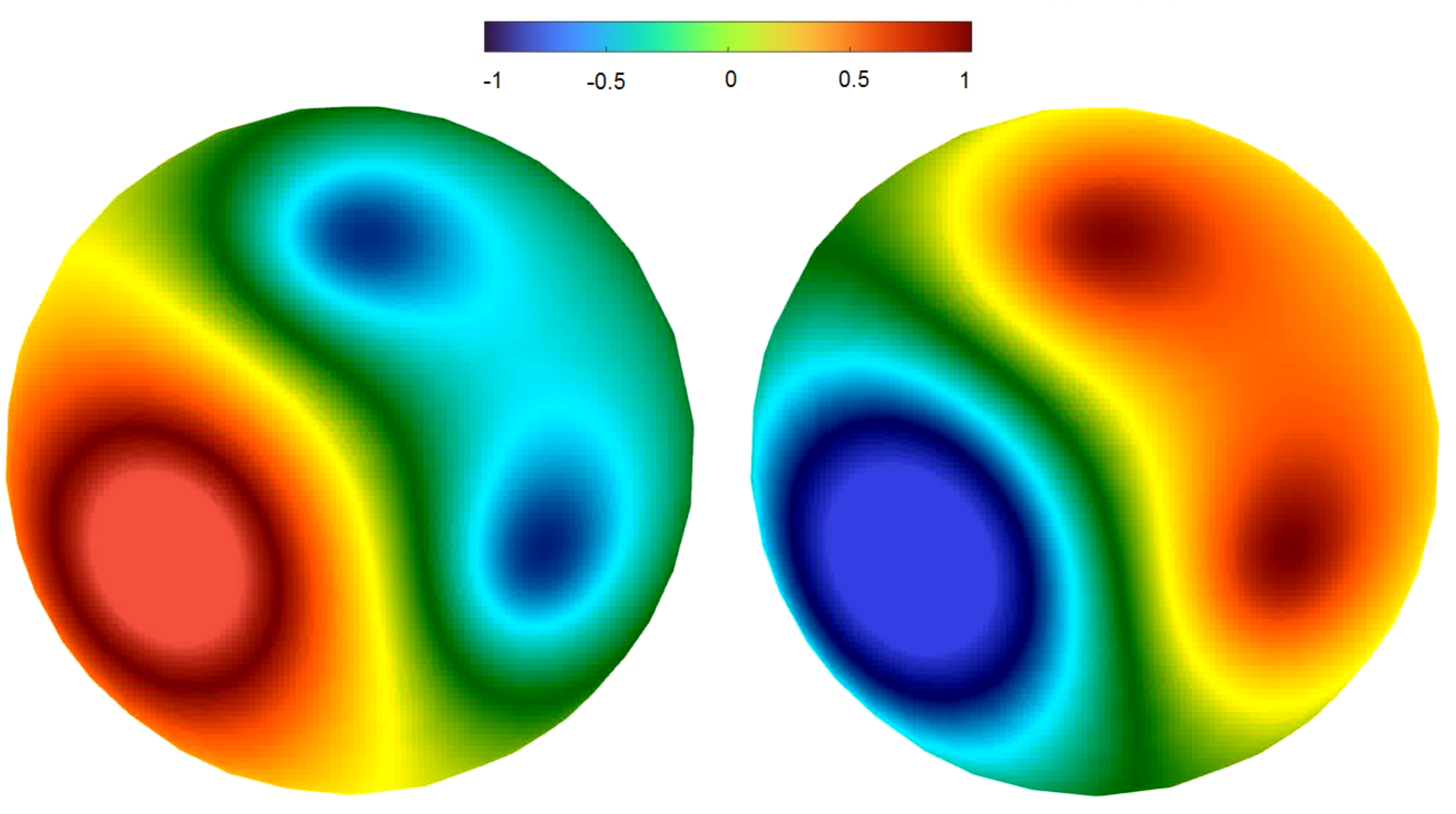
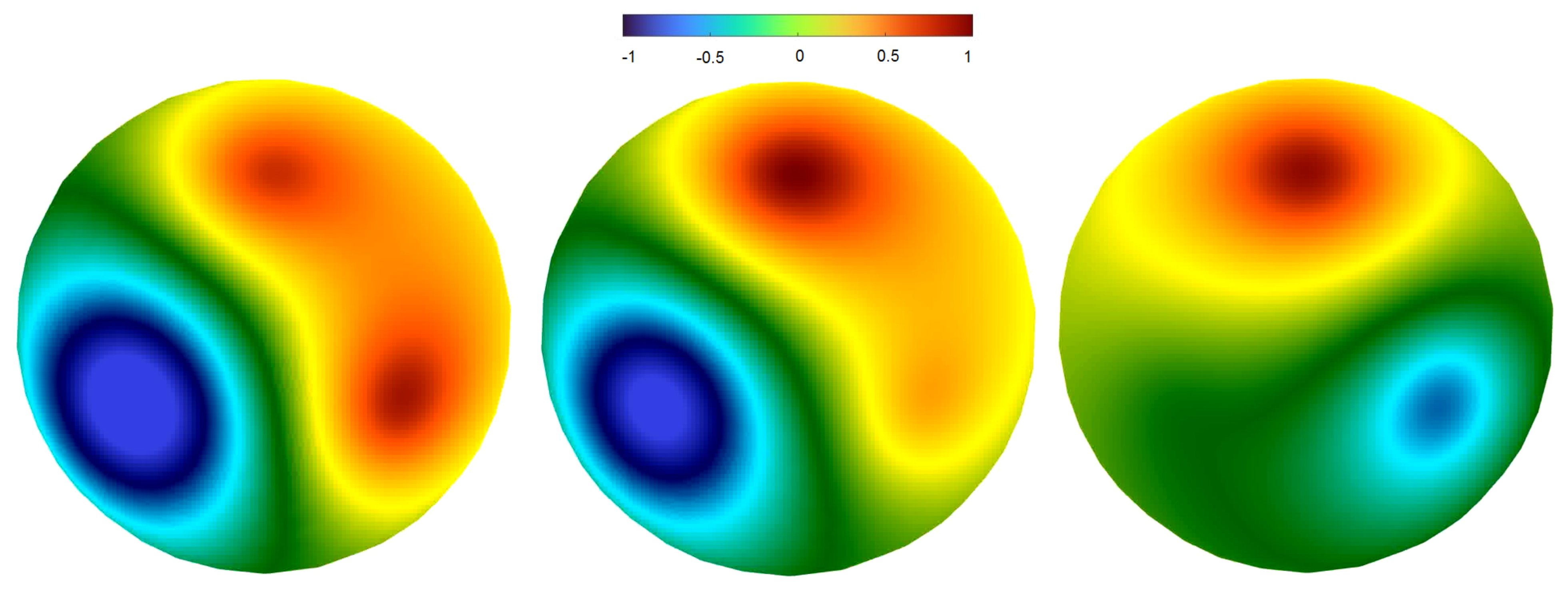
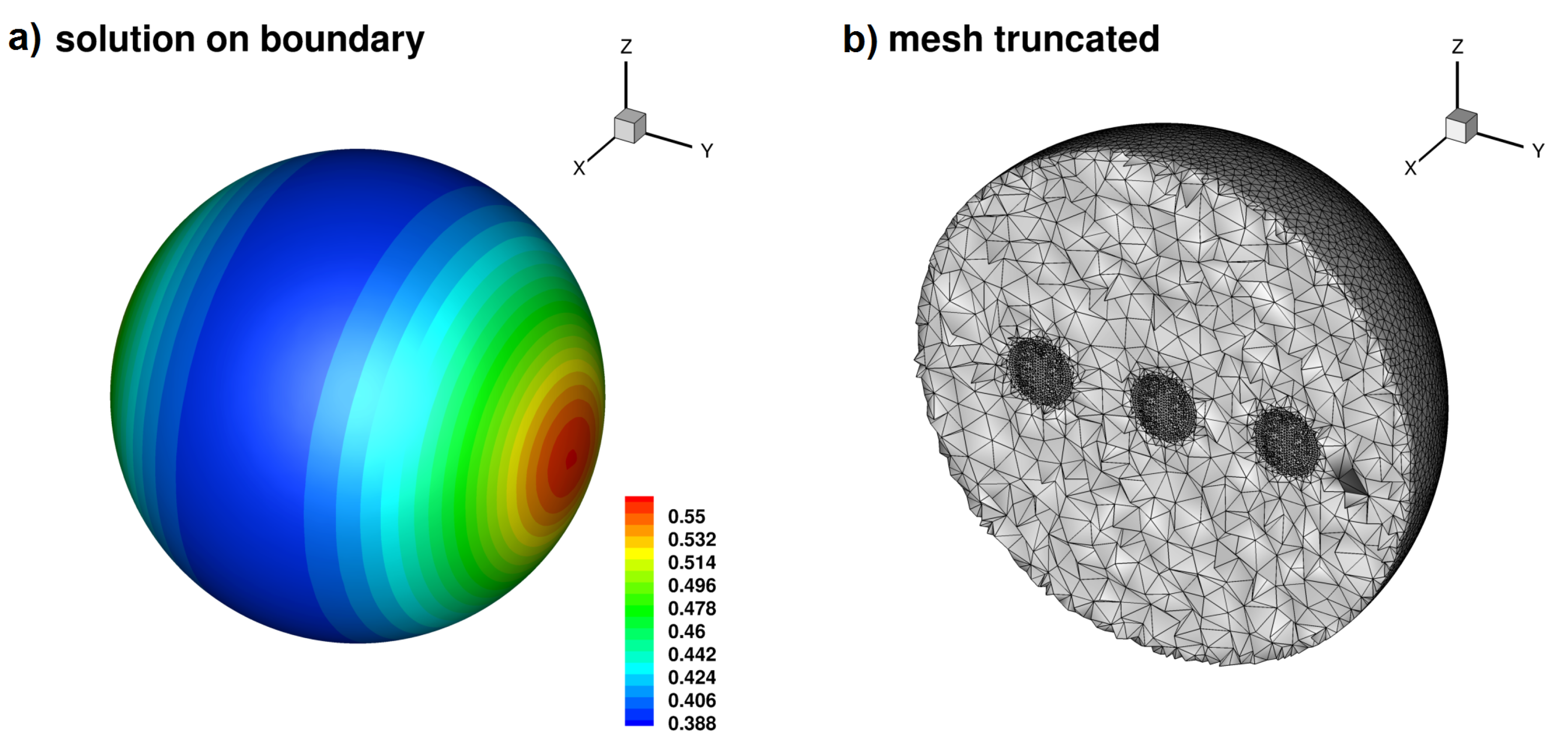
2.4. Dynamics on the Brain Surface (SimNIBS Software)
2.4.1. Software
2.4.2. Regimes of Spatiotemporal Dynamics
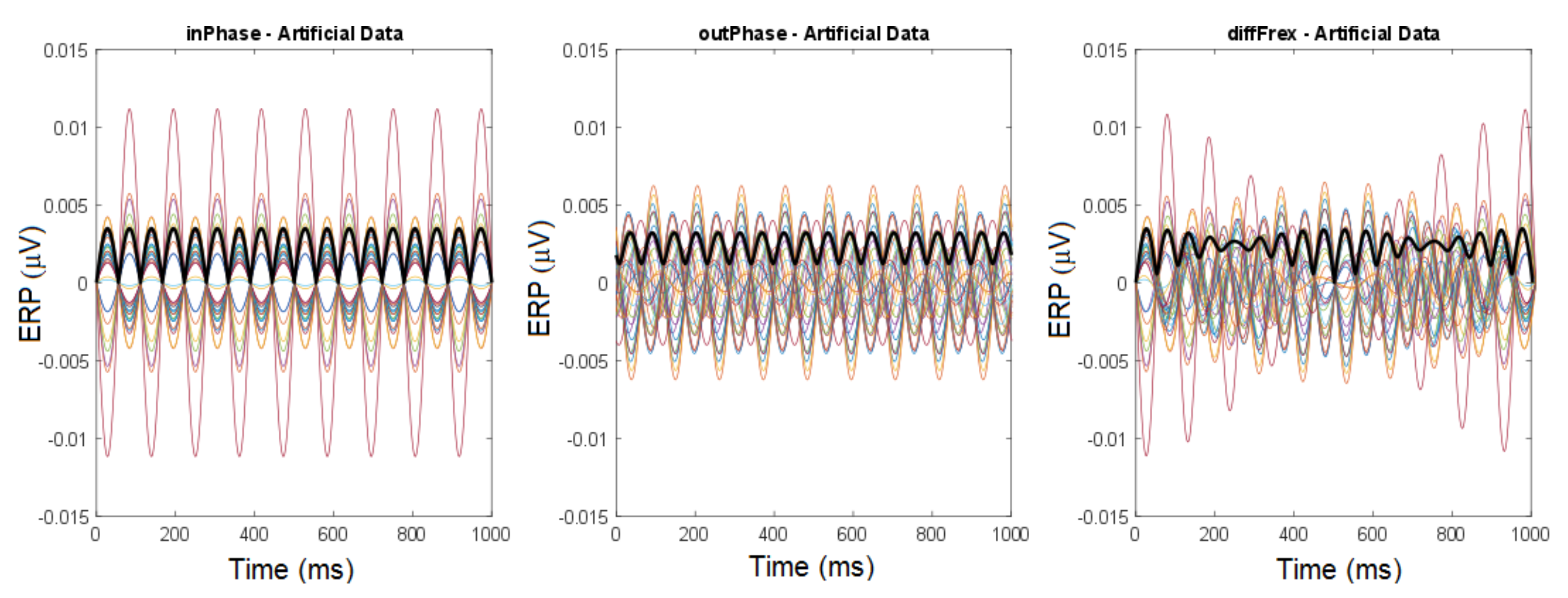
2.4.3. Global Field Power (GFP)
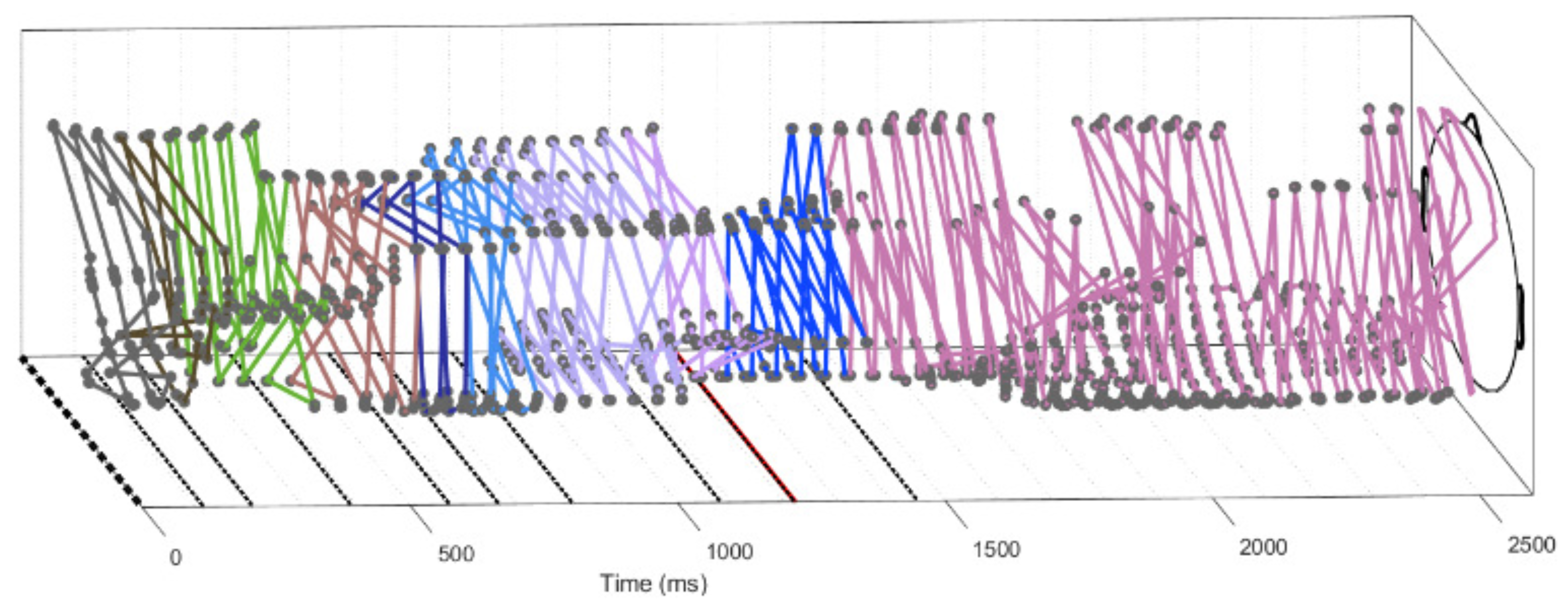
2.4.4. Trajectories
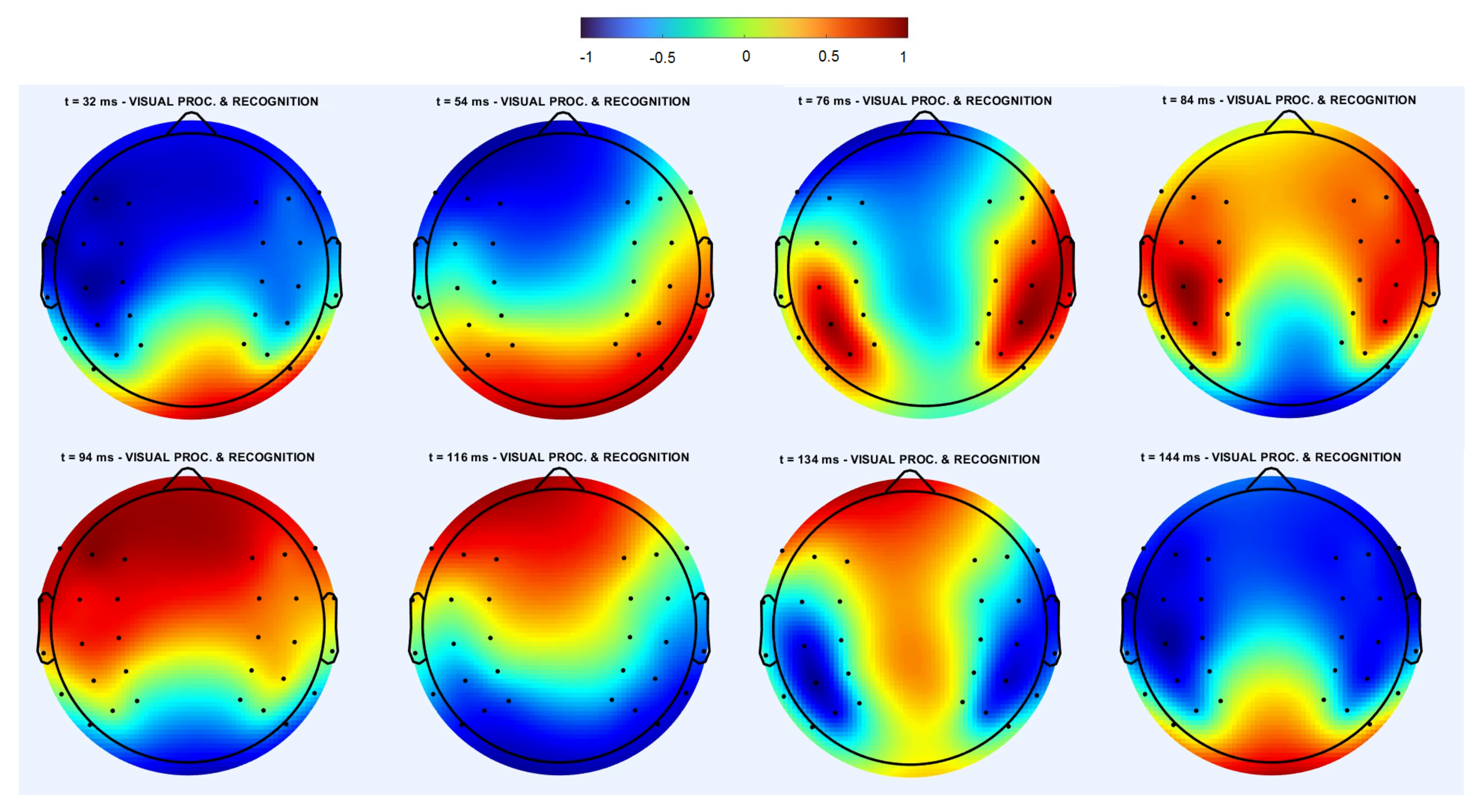
3. Spatiotemporal Dynamics in EEG Data
3.1. Data Acquisition and Analysis
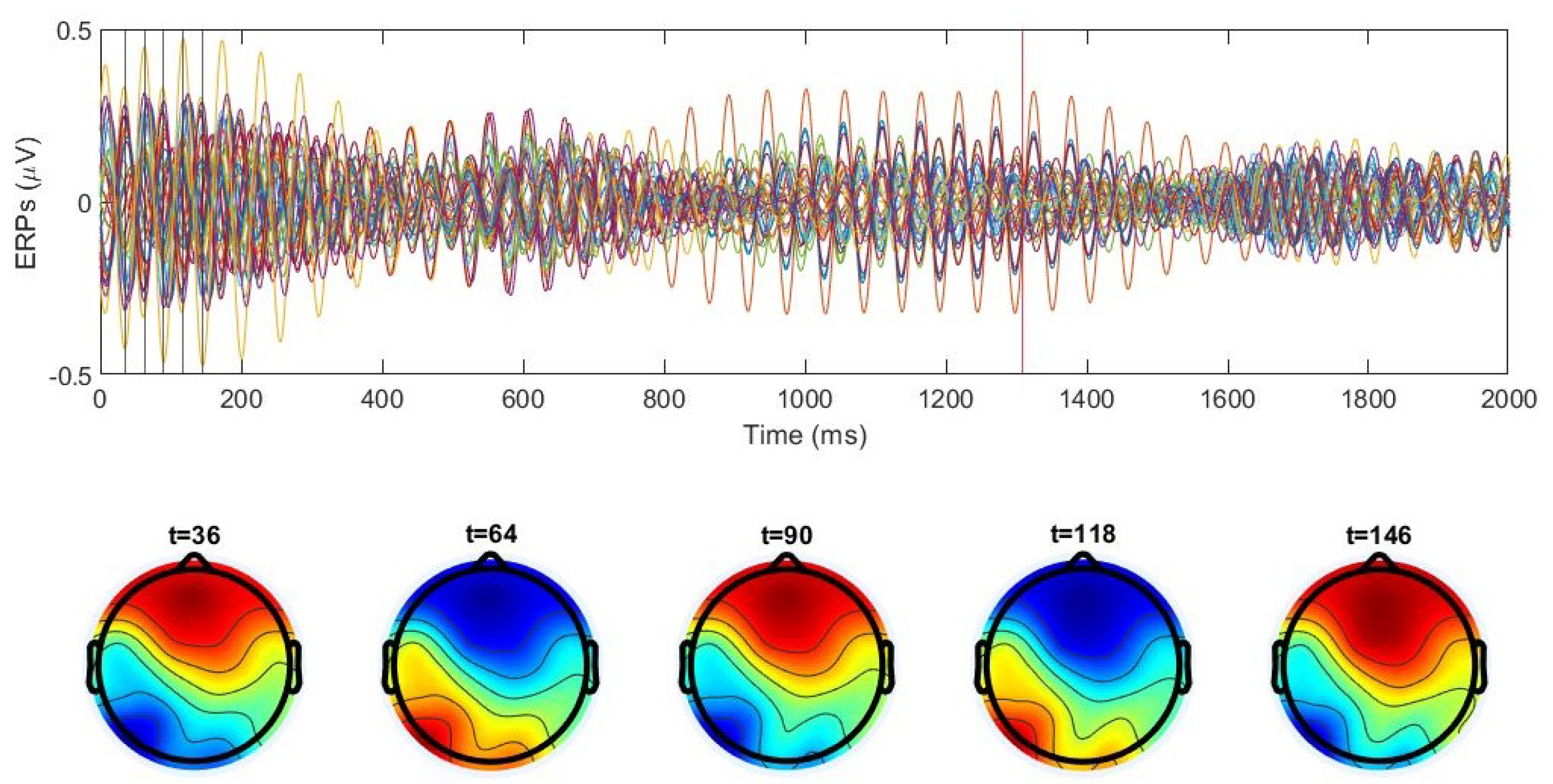
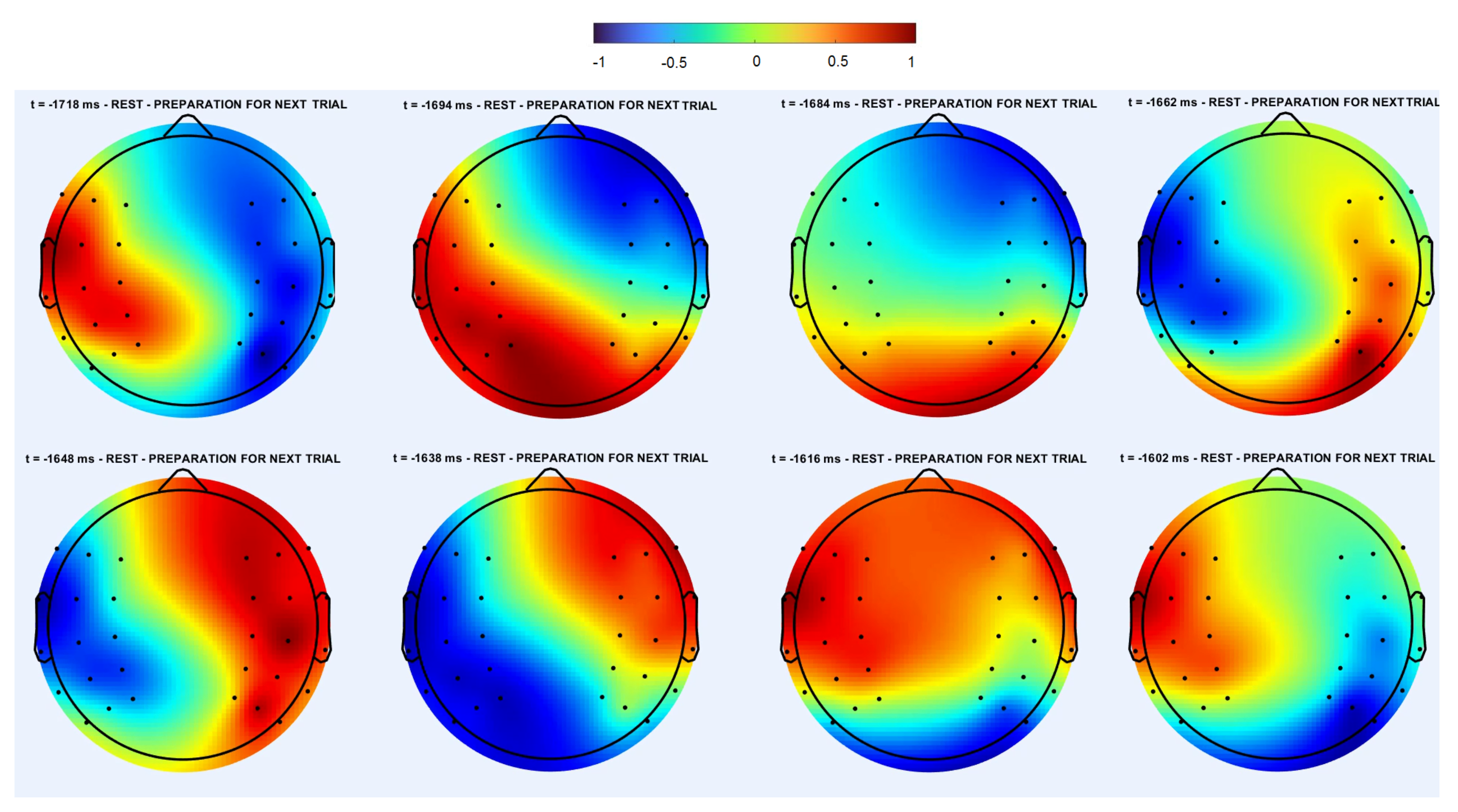
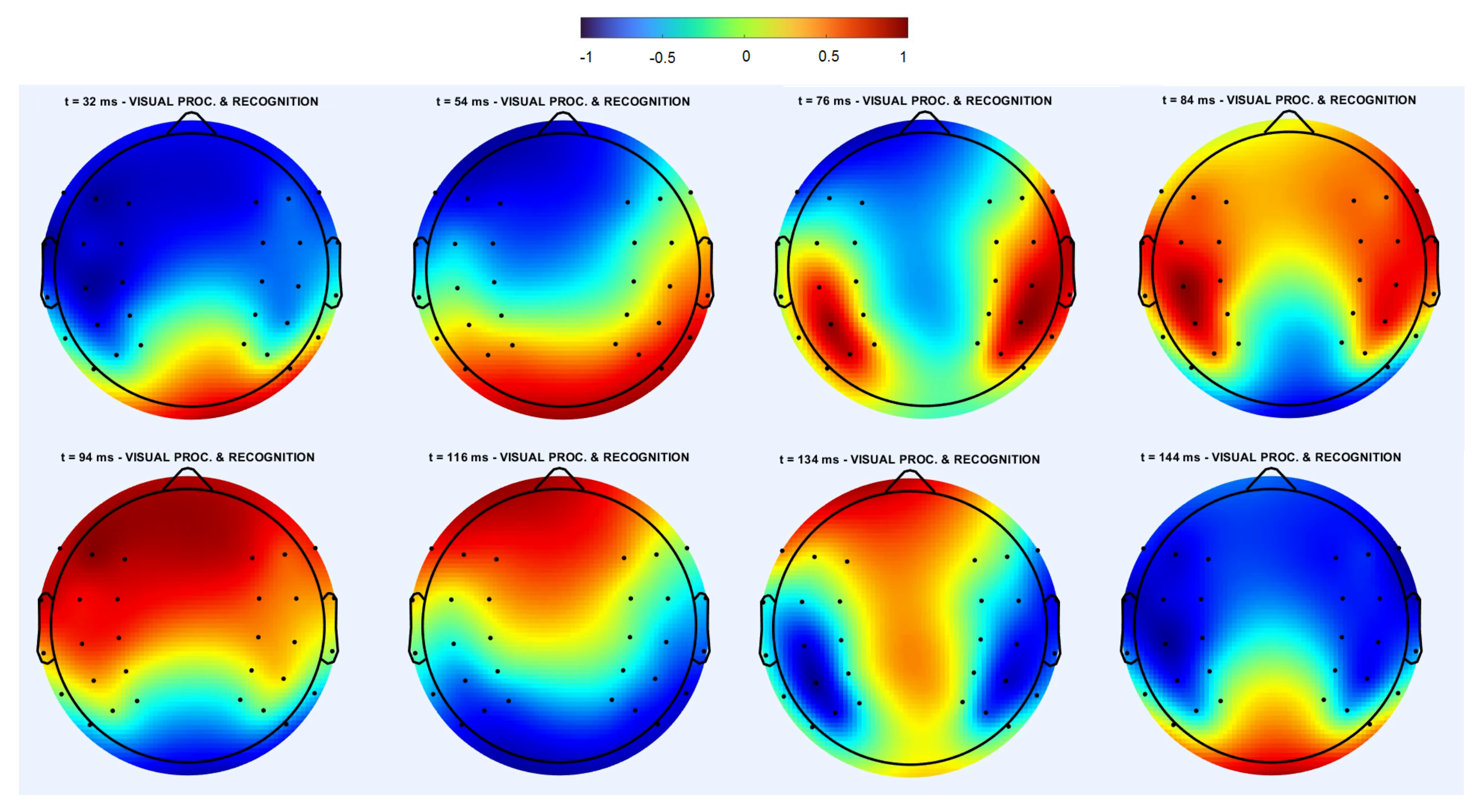
3.1.1. Cross-Trial Analysis
3.1.2. Individual Trial Analysis
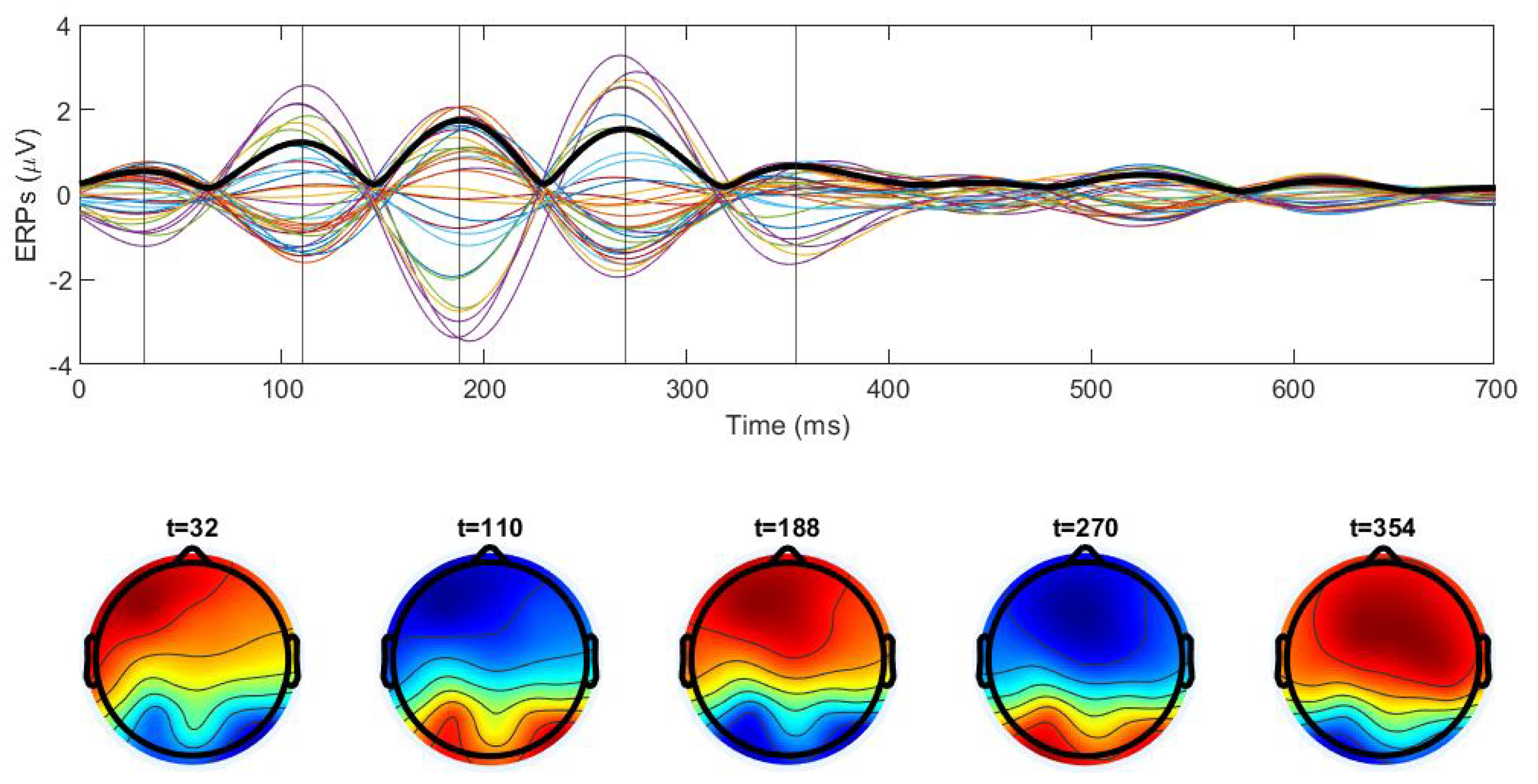
3.2. Averaged Cross-Trial Dynamics
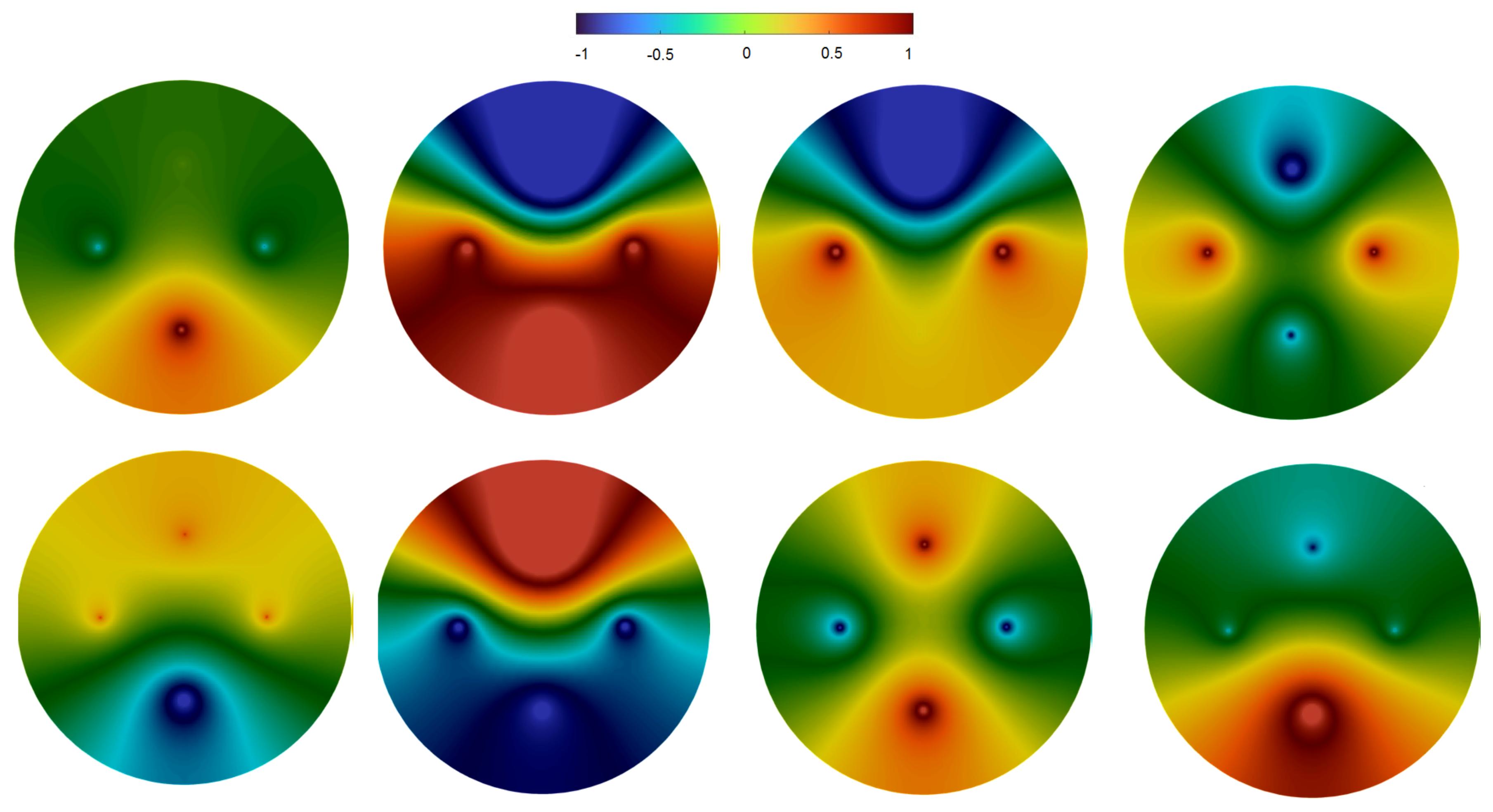
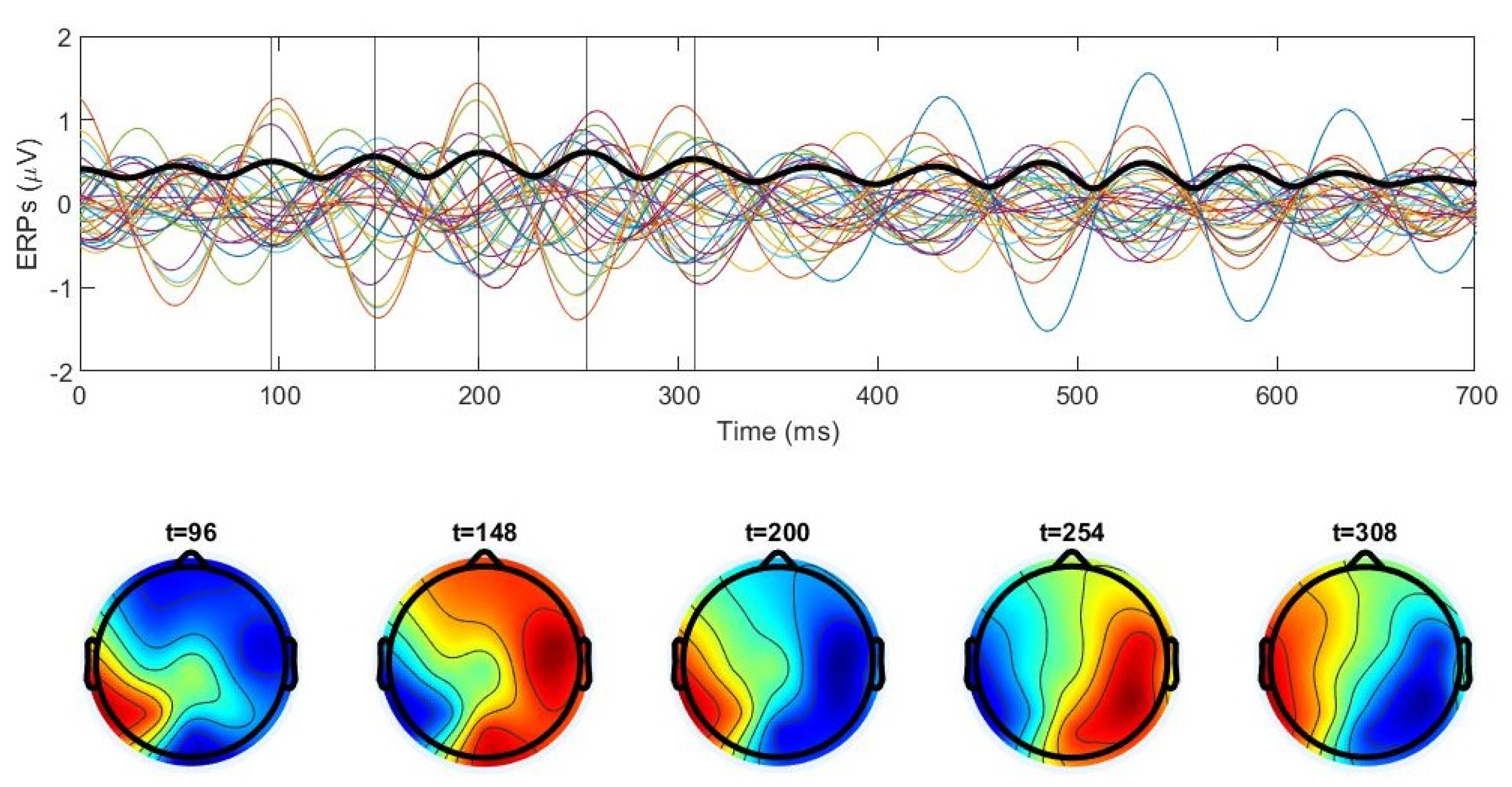
3.3. Spatiotemporal Regimes in Individual Trial Dynamics
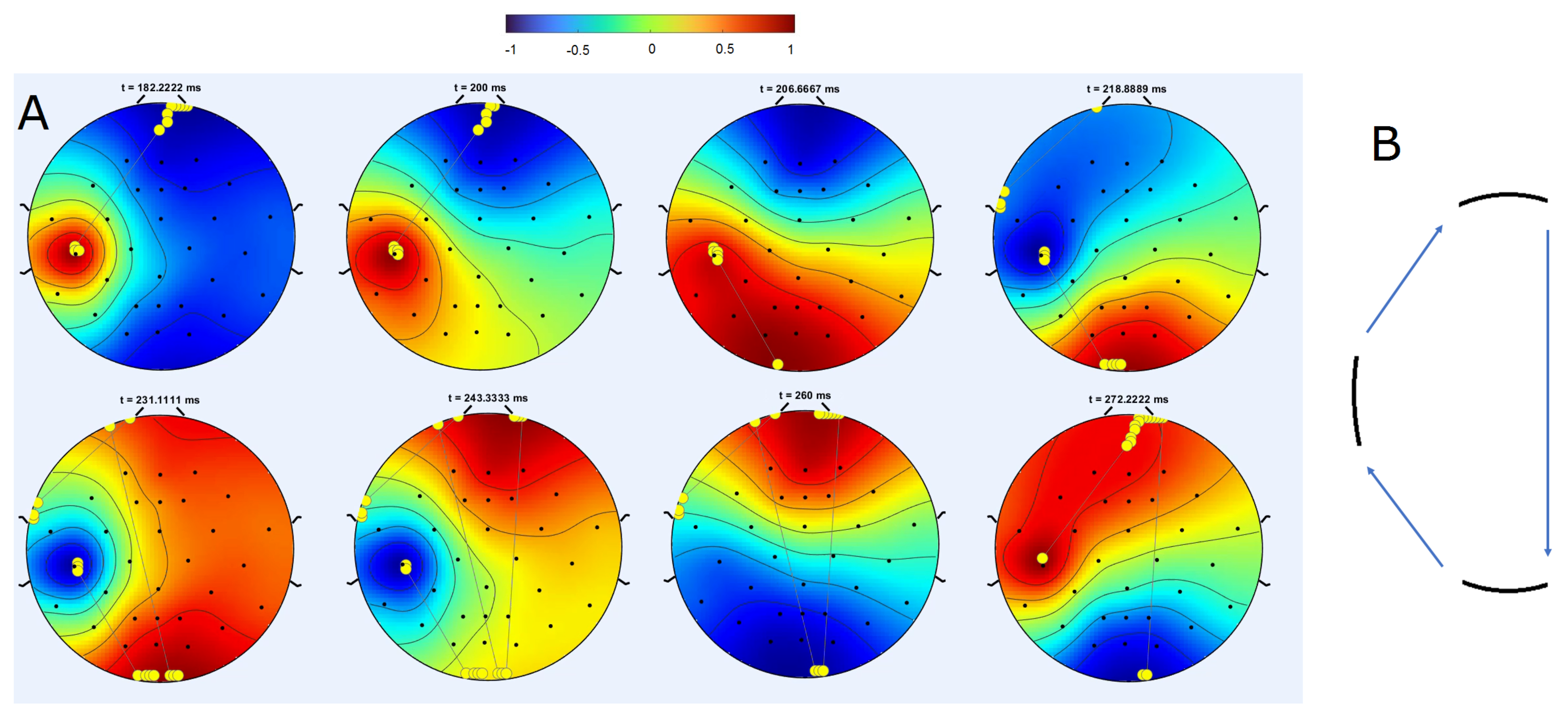
3.3.1. Rotating Regime
3.3.2. Symmetric Regime
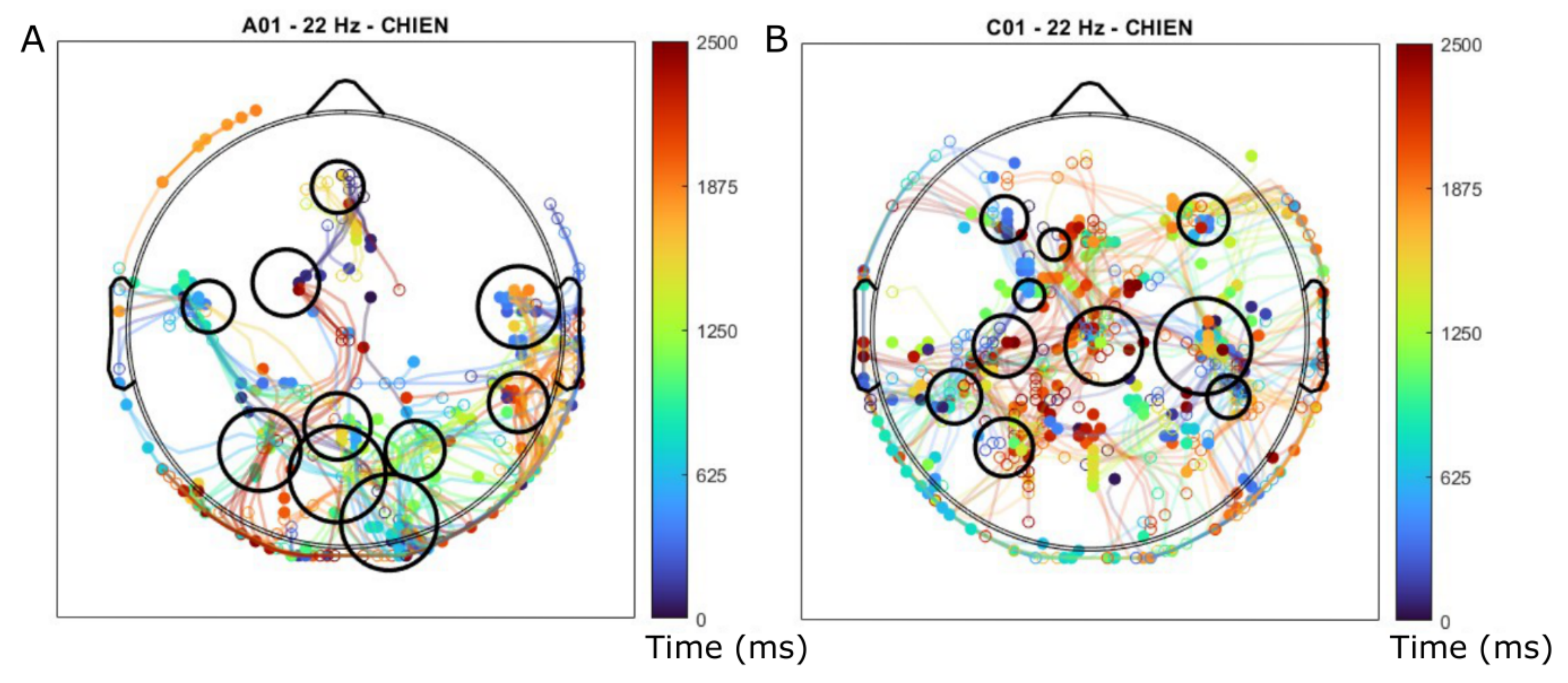
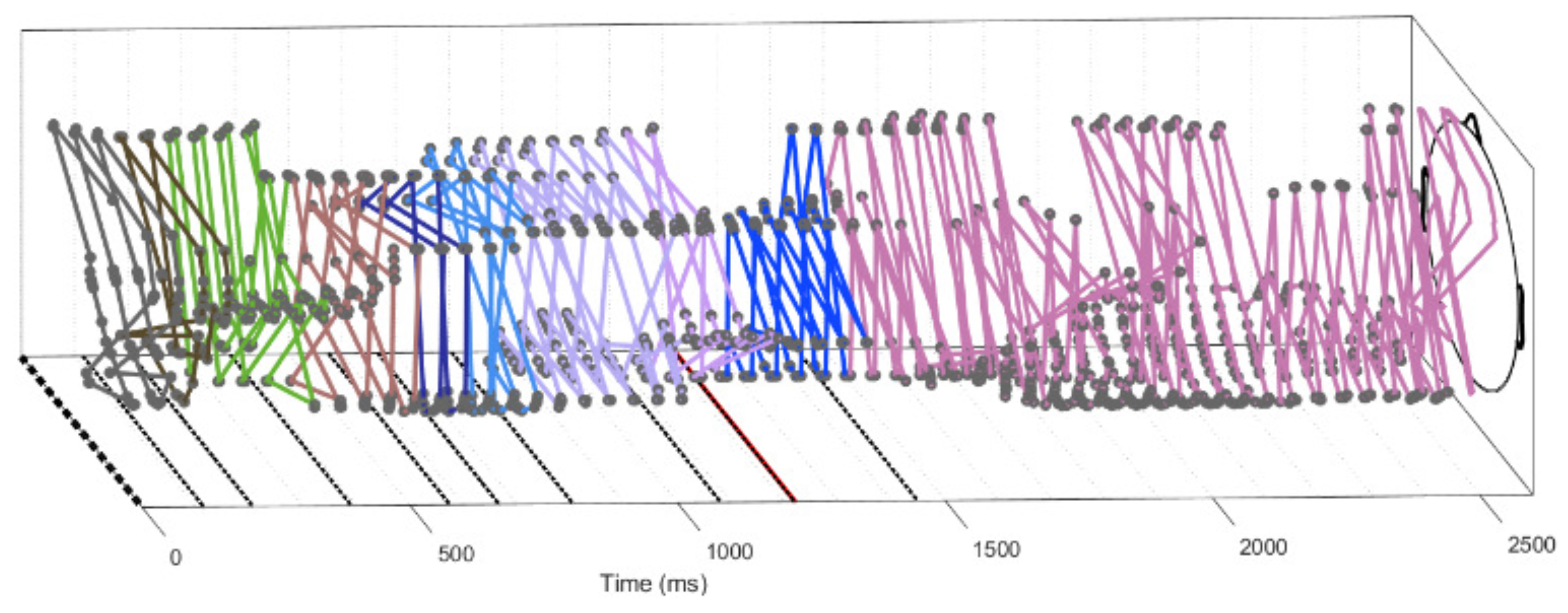
3.4. Trajectories
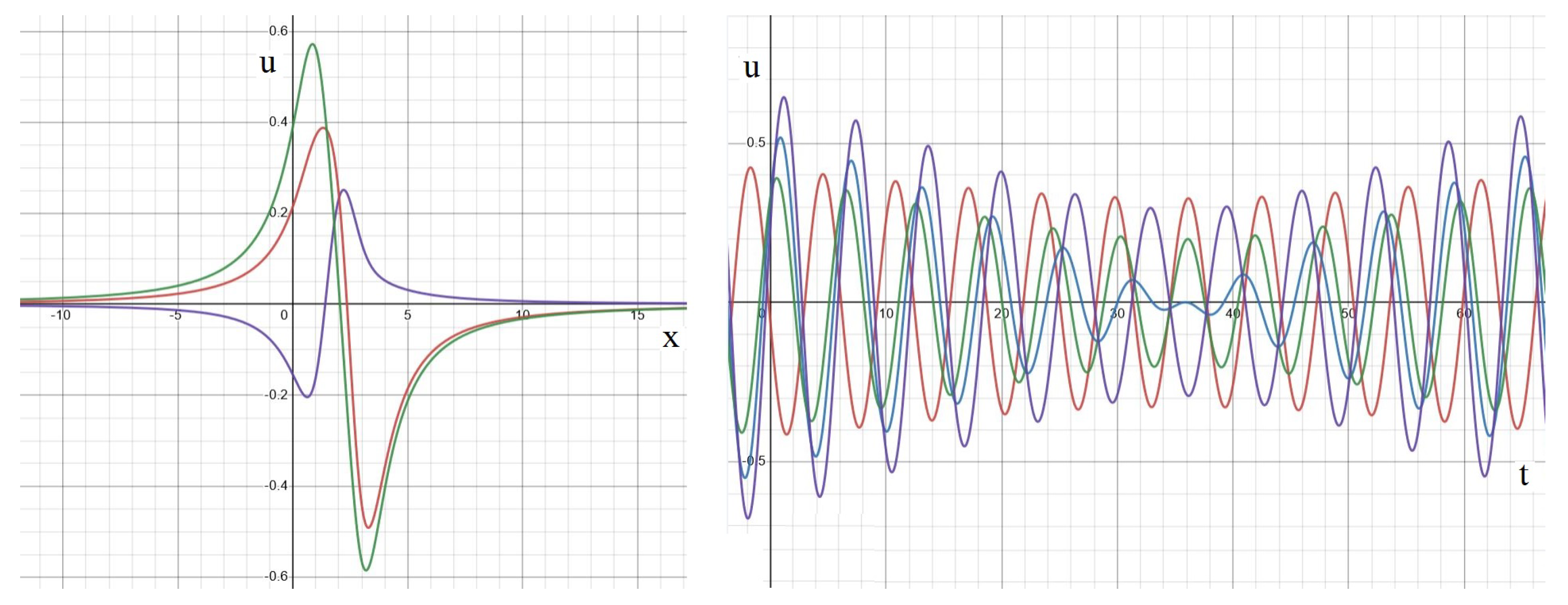
3.5. Moving Waves
4. Discussion
4.1. Mathematical Model
4.2. Dynamics in the Brain Source Model
4.3. Traveling and Moving Waves
4.4. Limitations and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
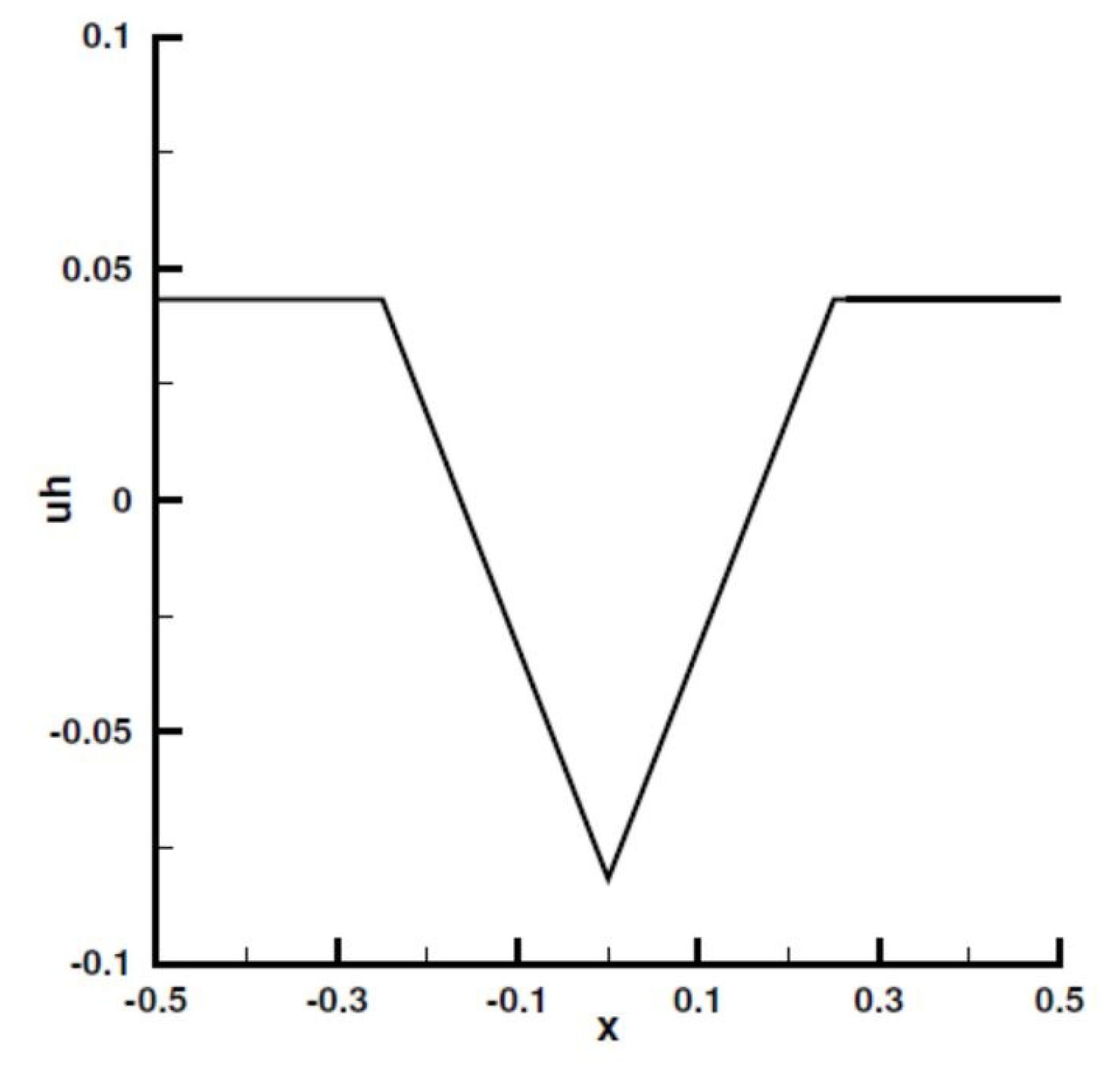
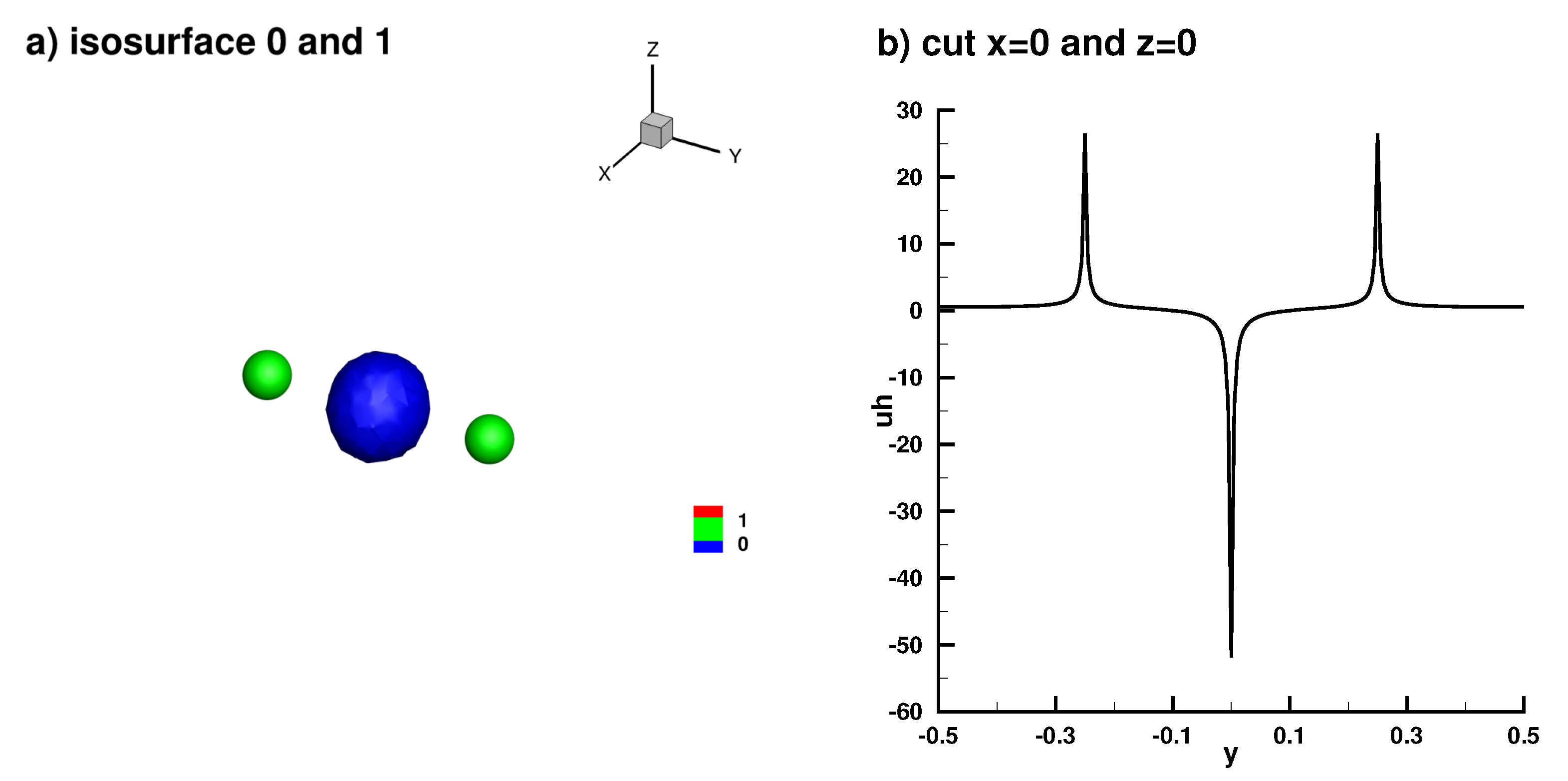
Appendix A. Analytical Solution

Standing Waves and Other Dynamics
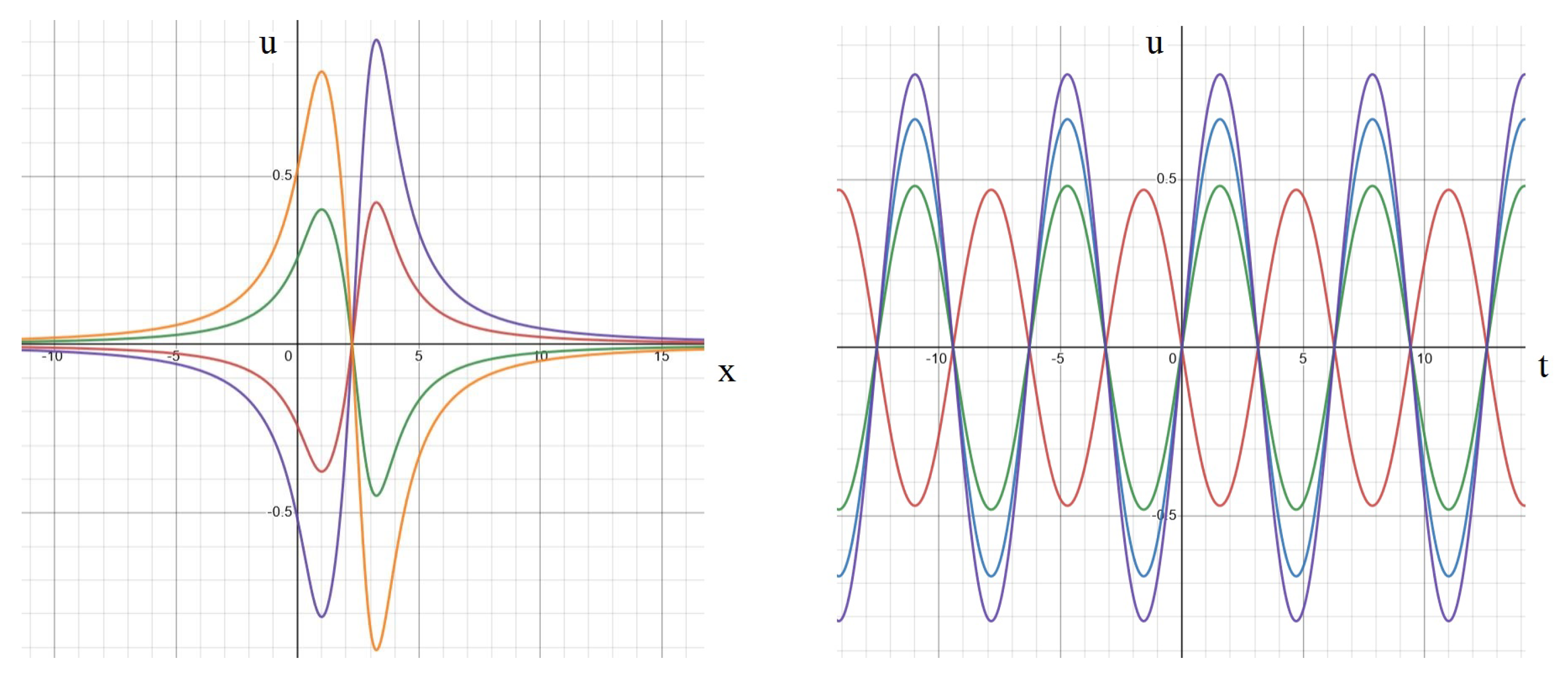
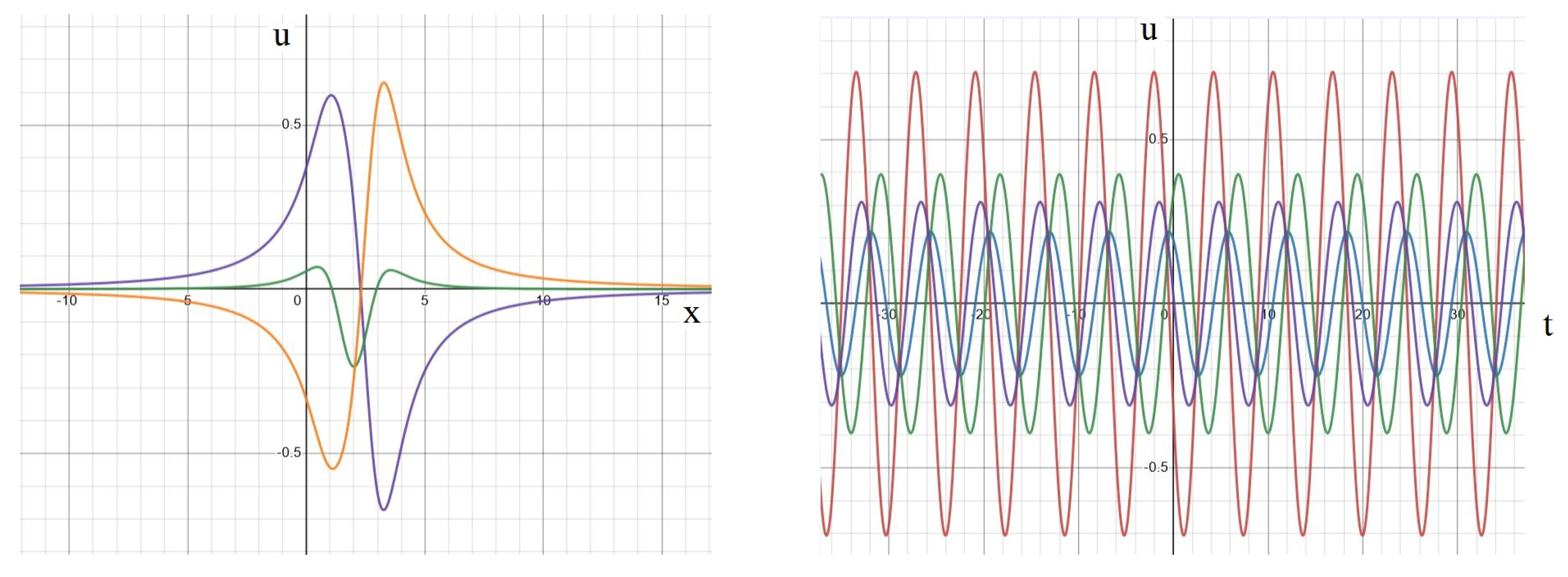

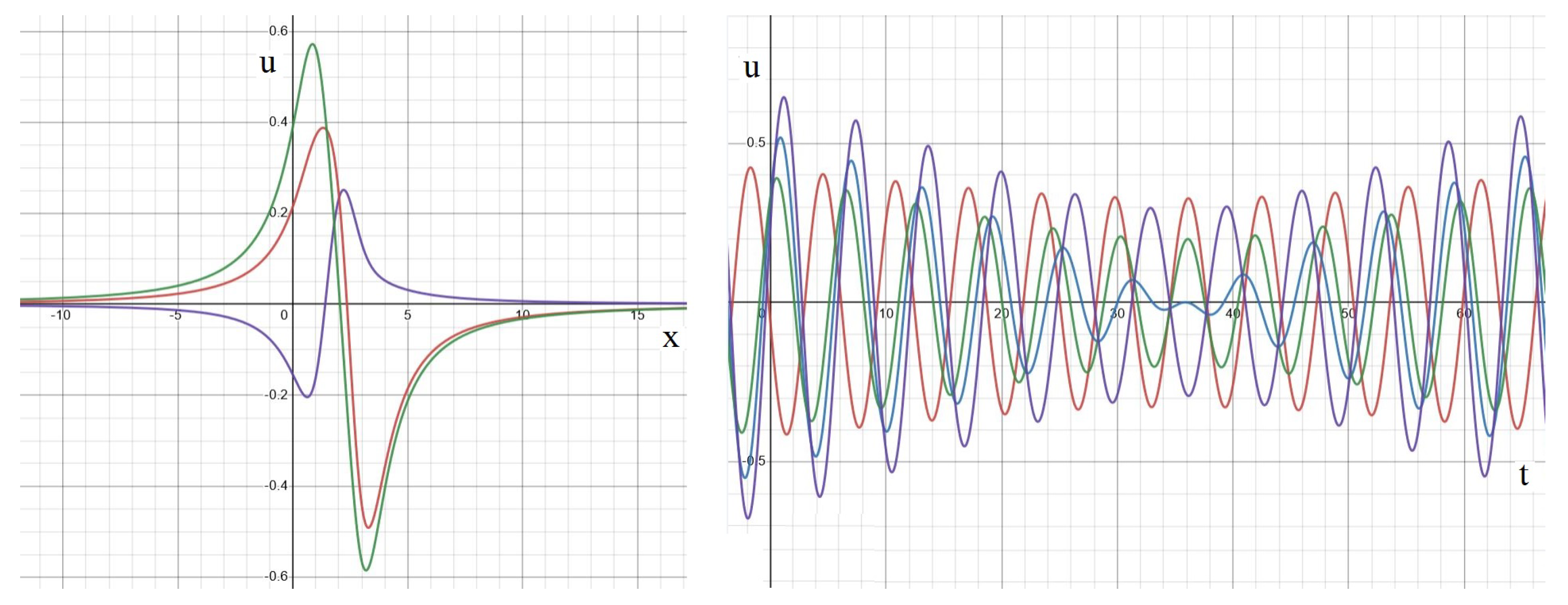
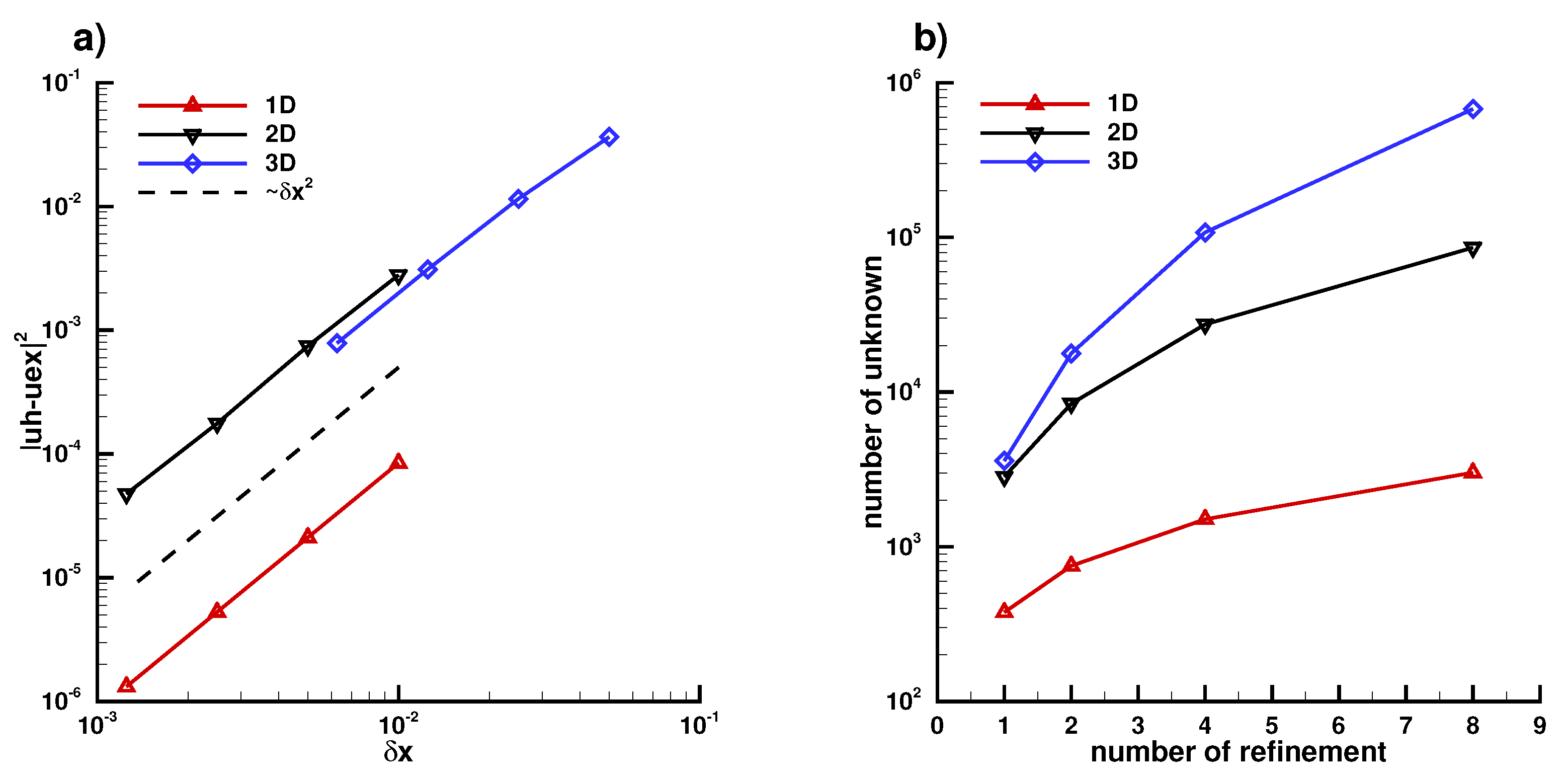
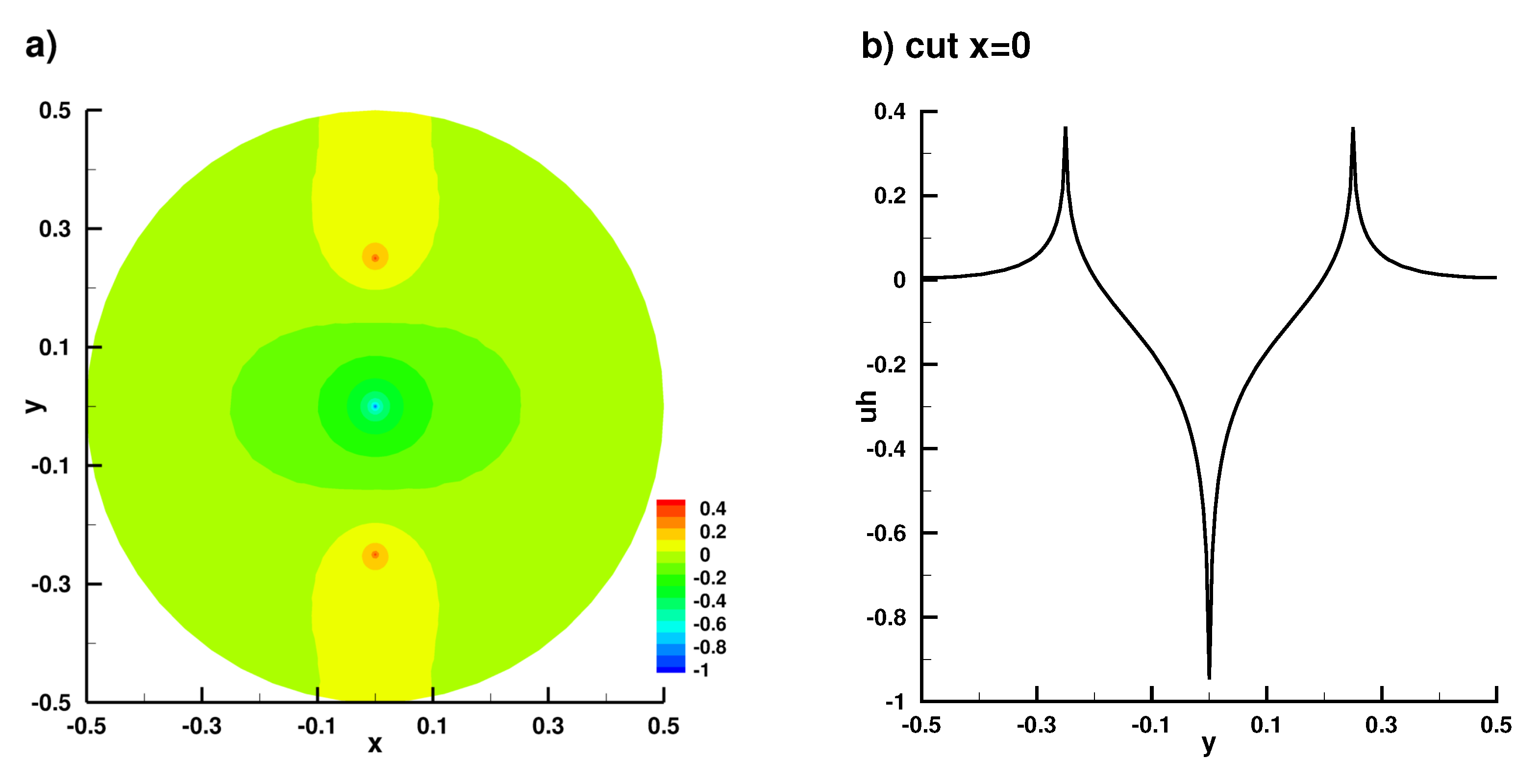
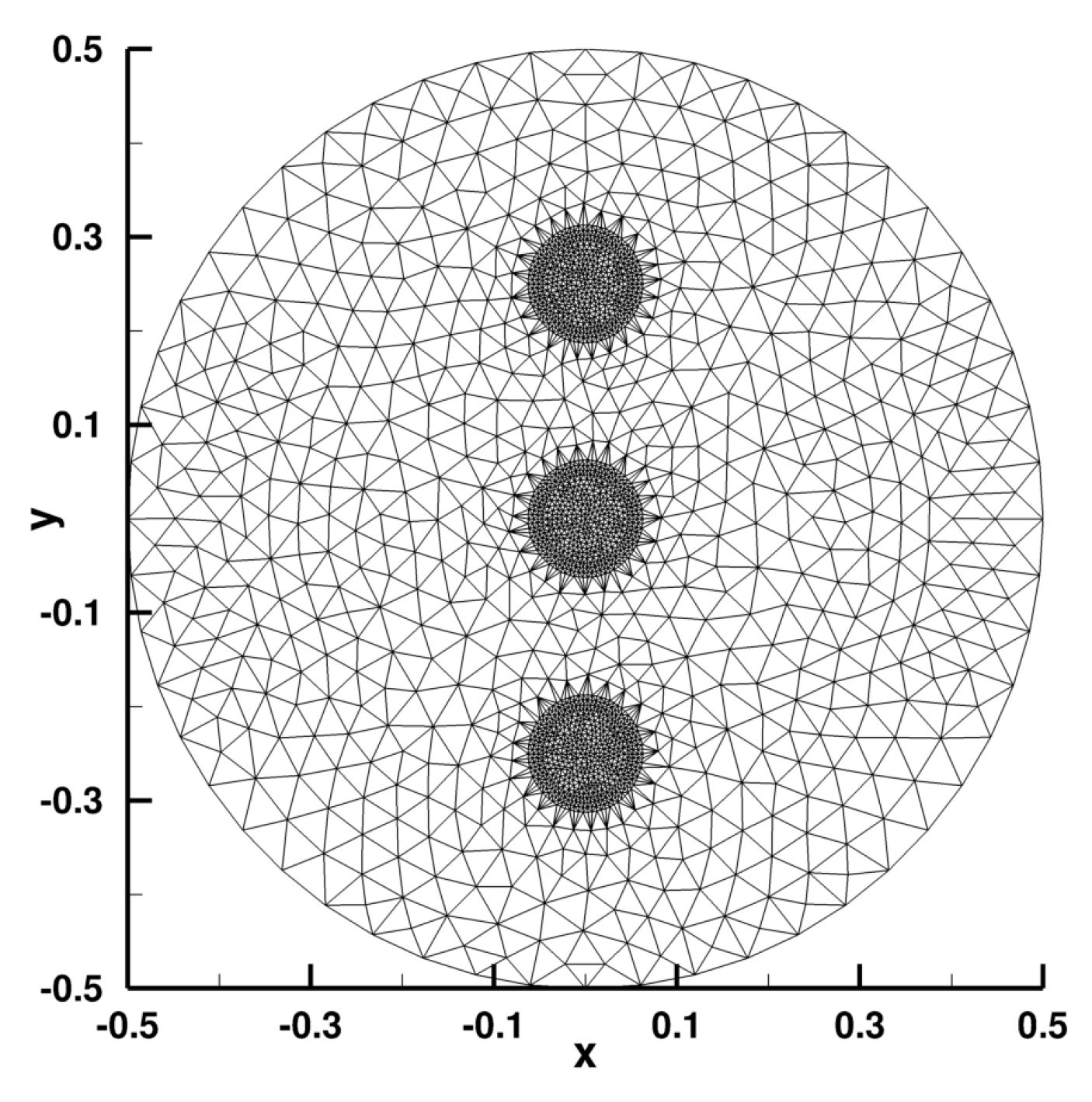
Appendix B. Numerical Implementation
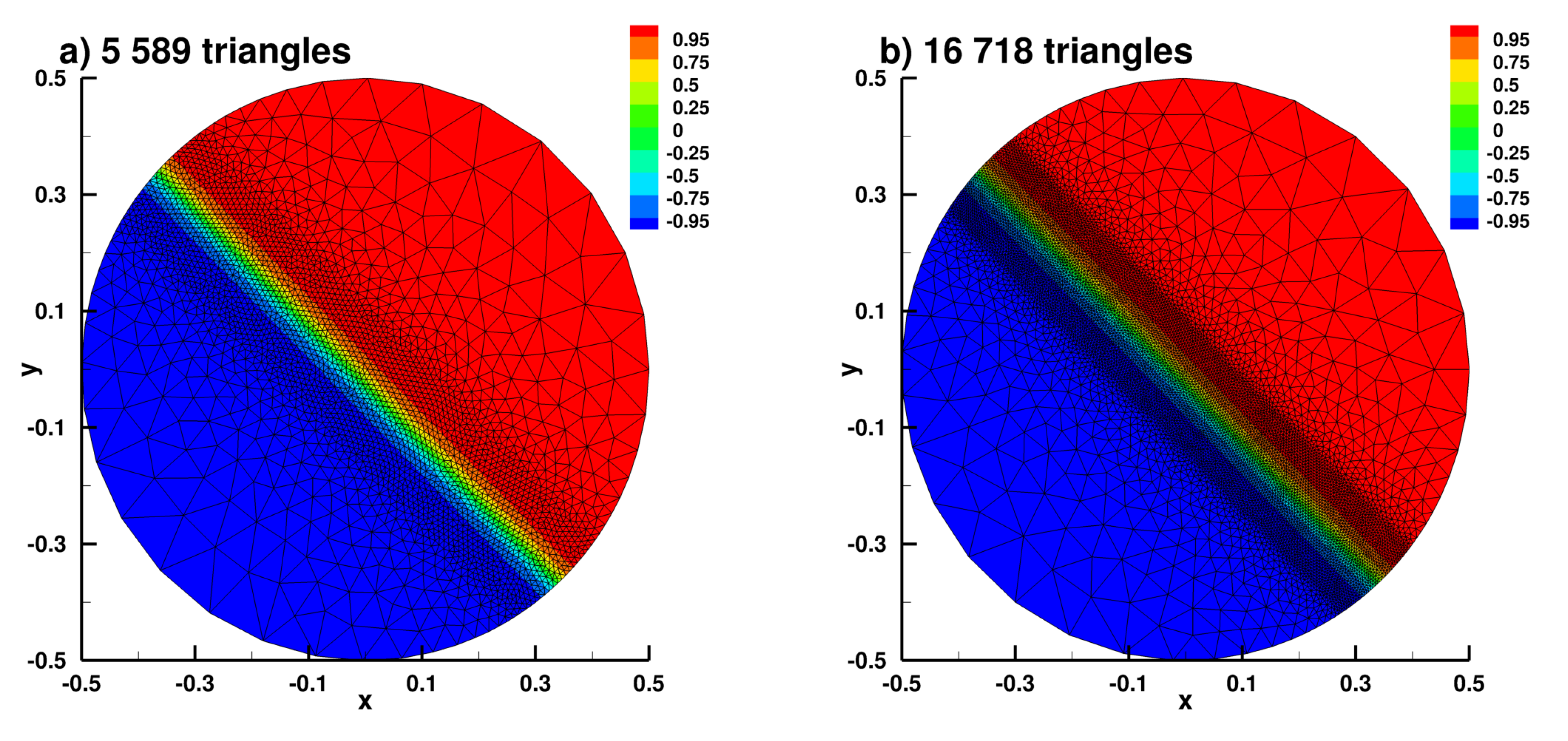
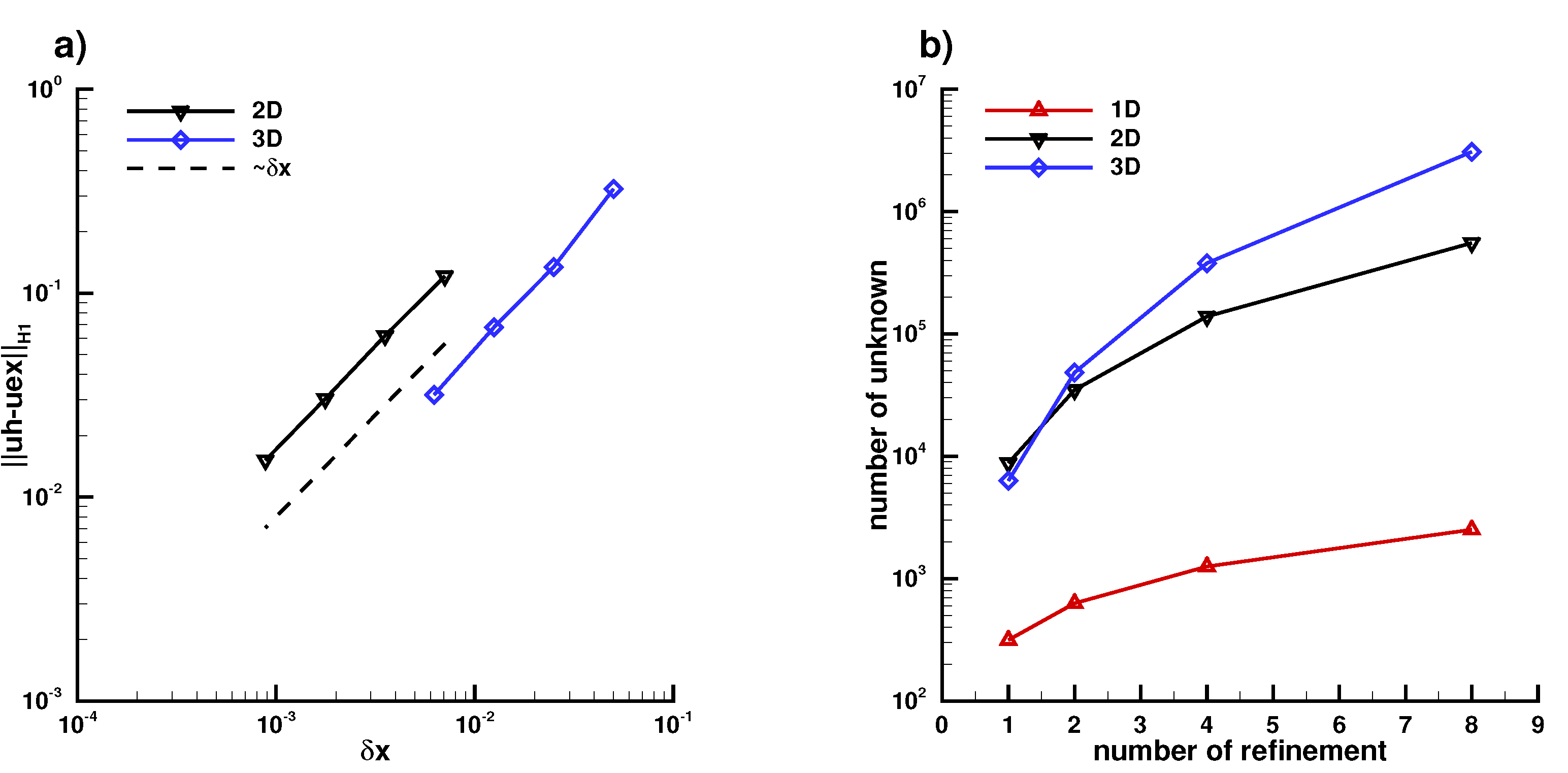
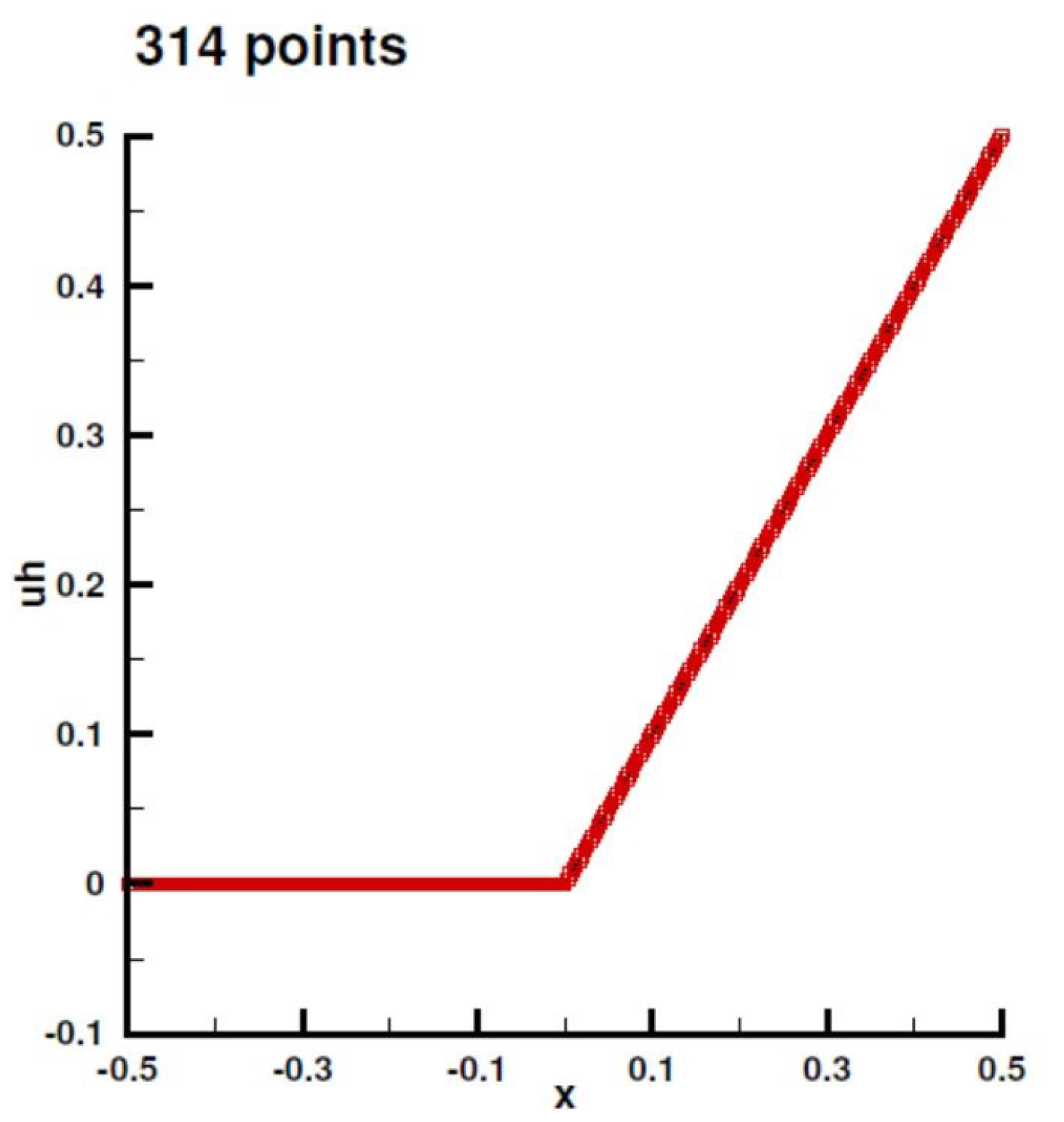
Appendix B.1. Laplace Equation Convergence Rate

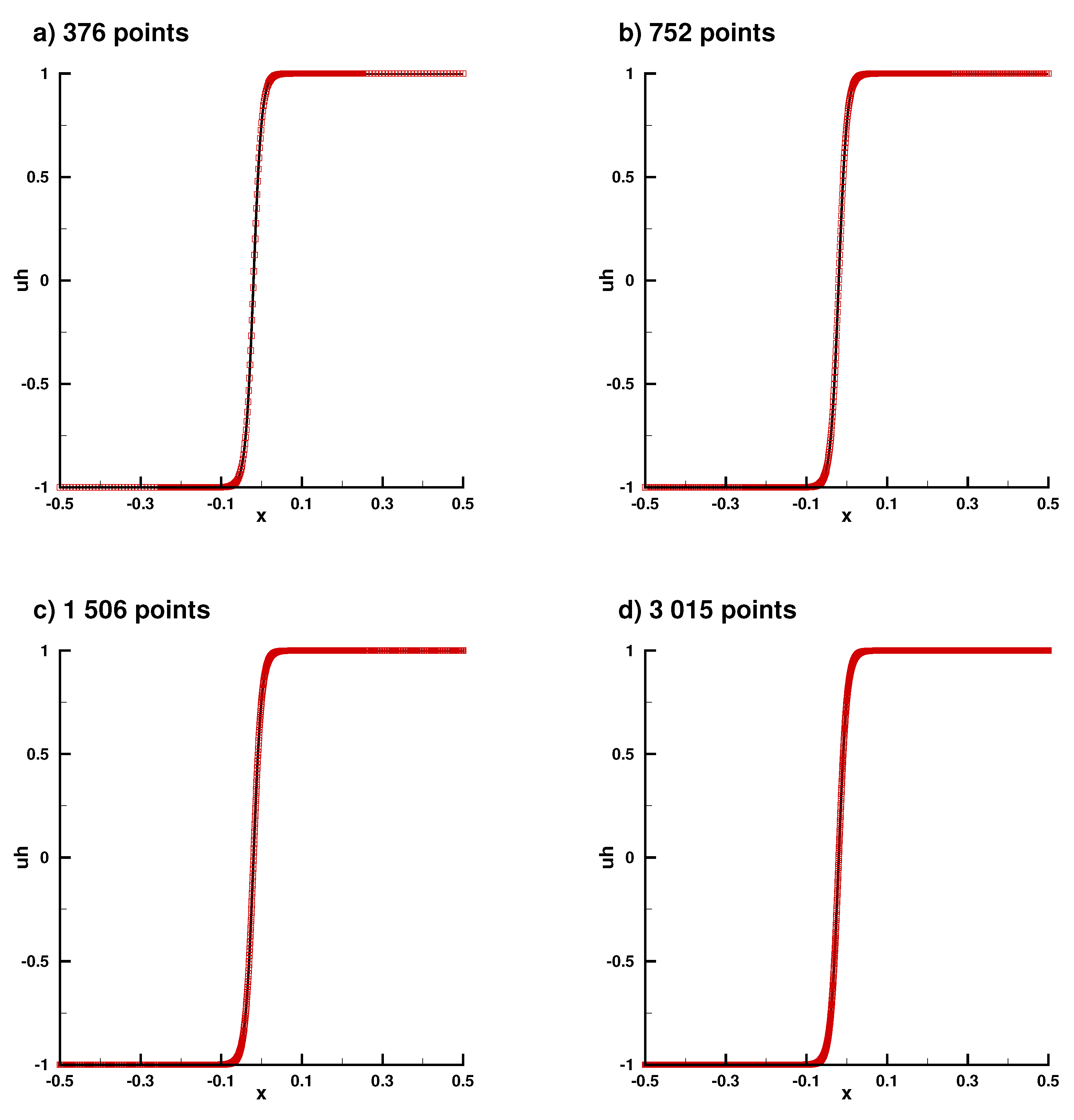
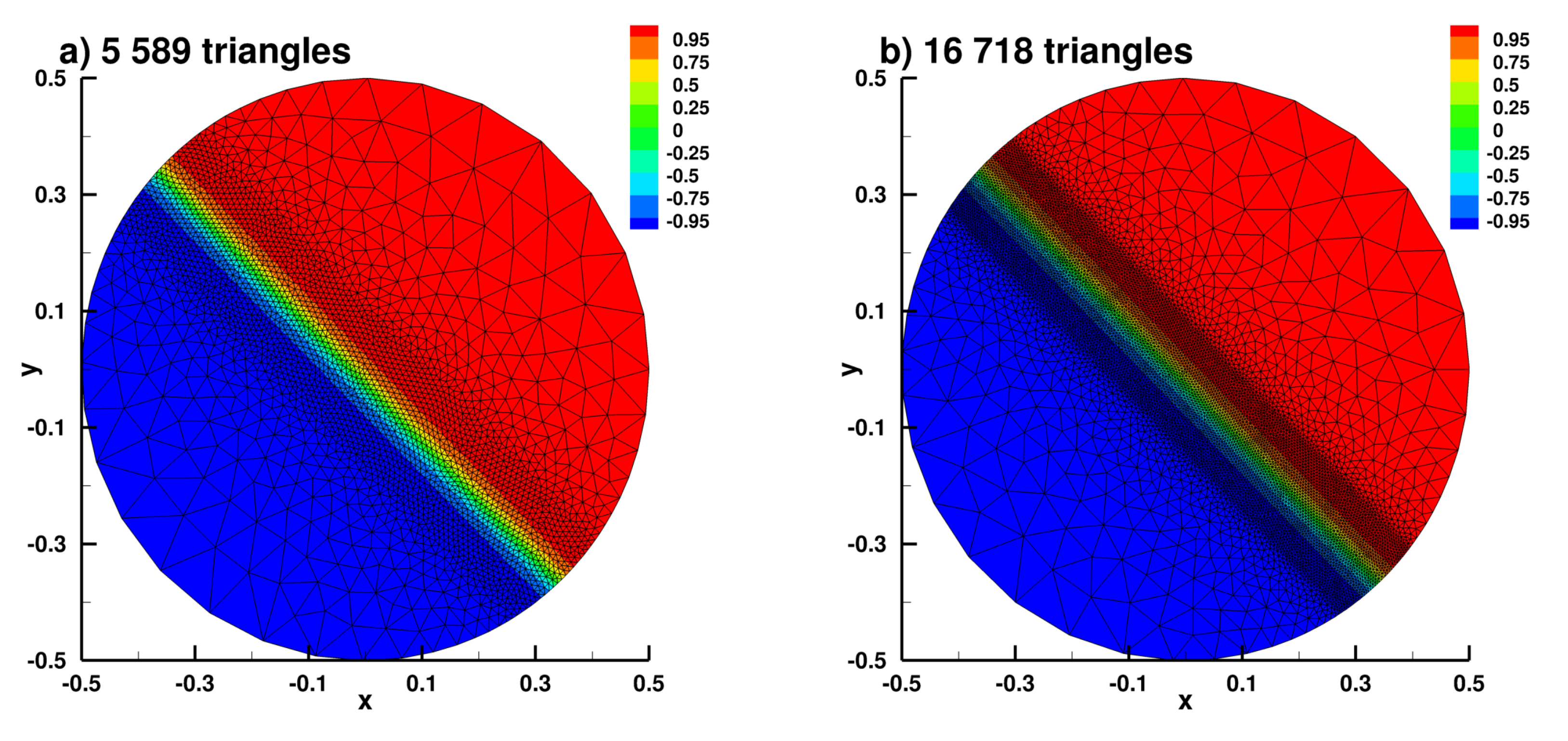
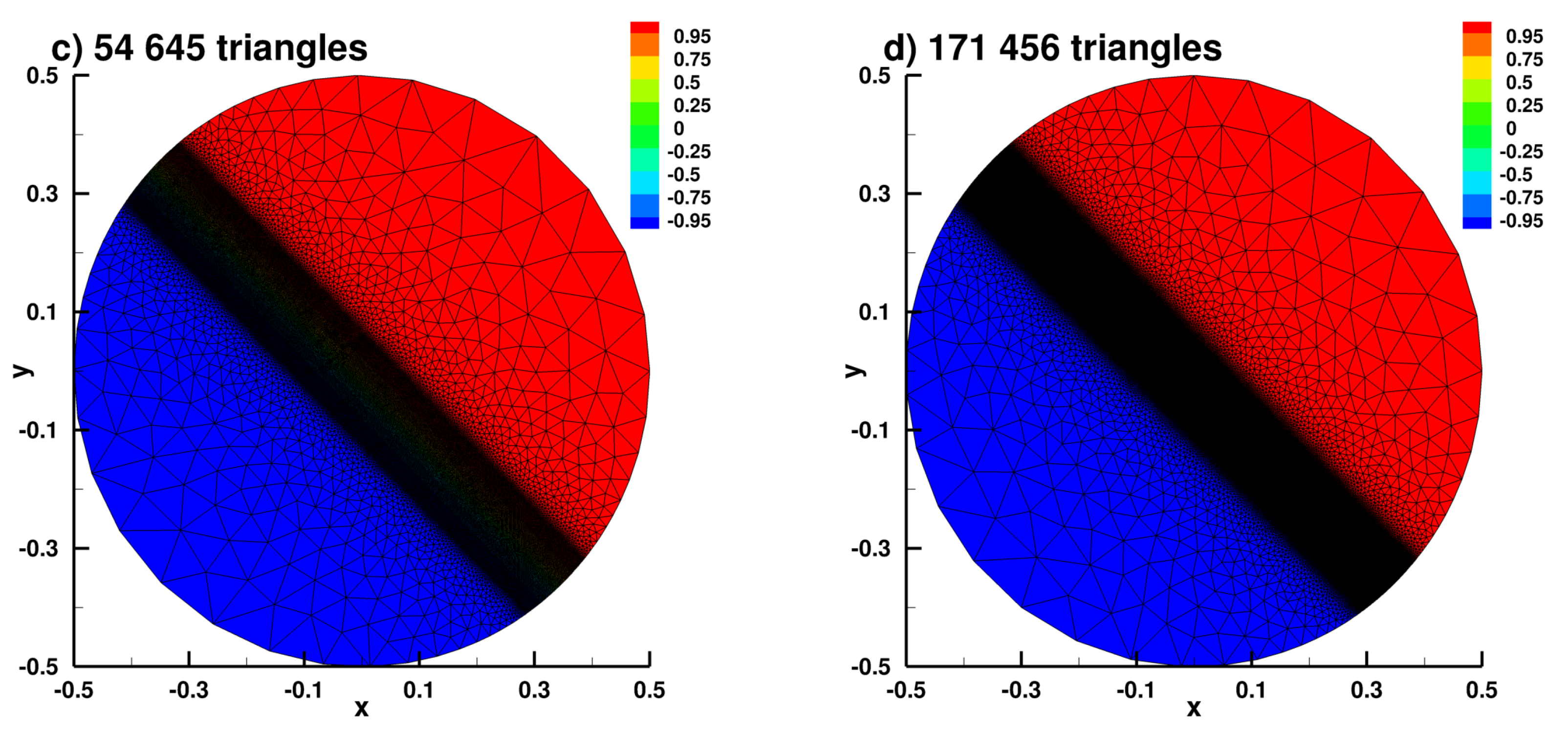

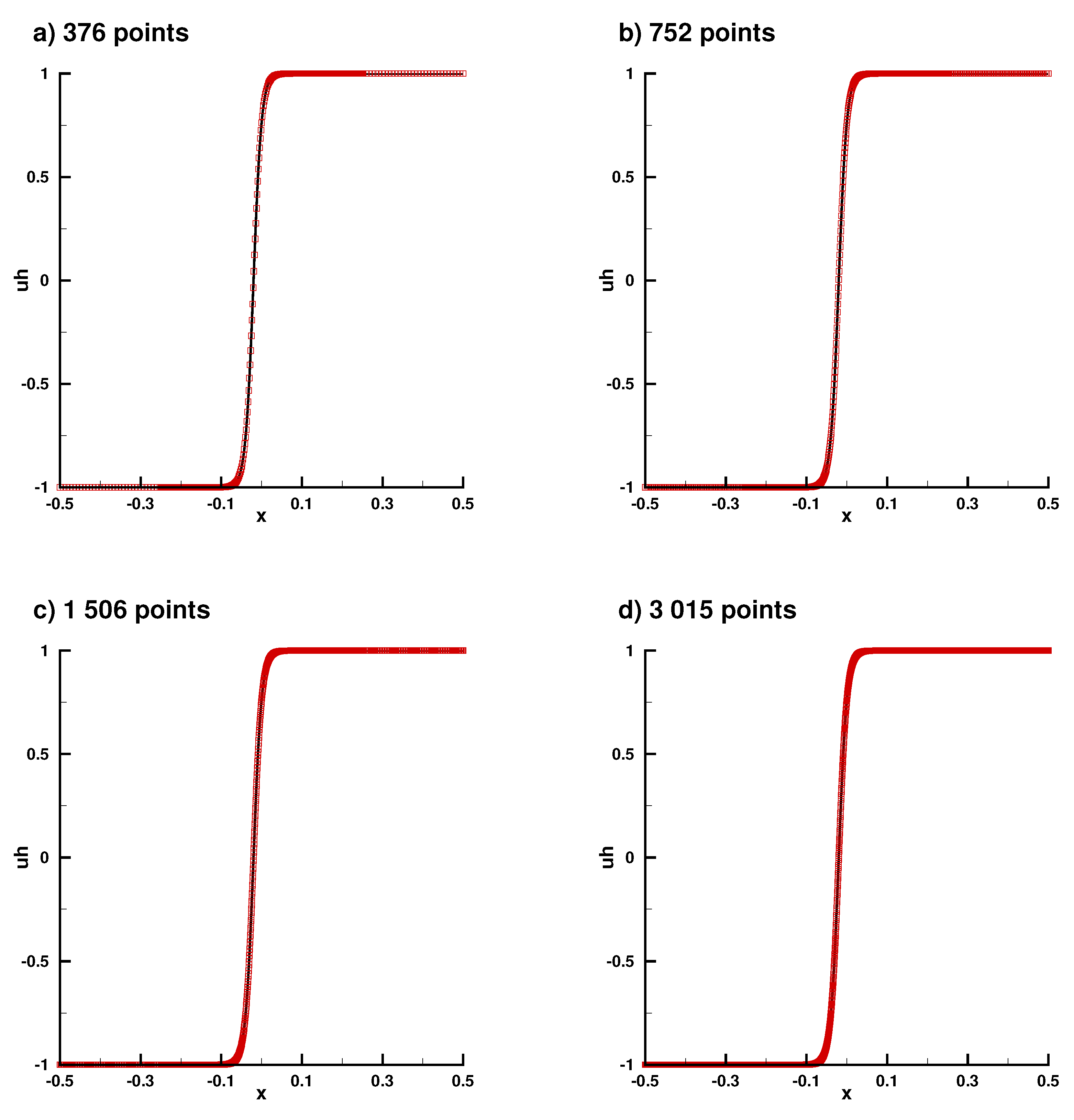
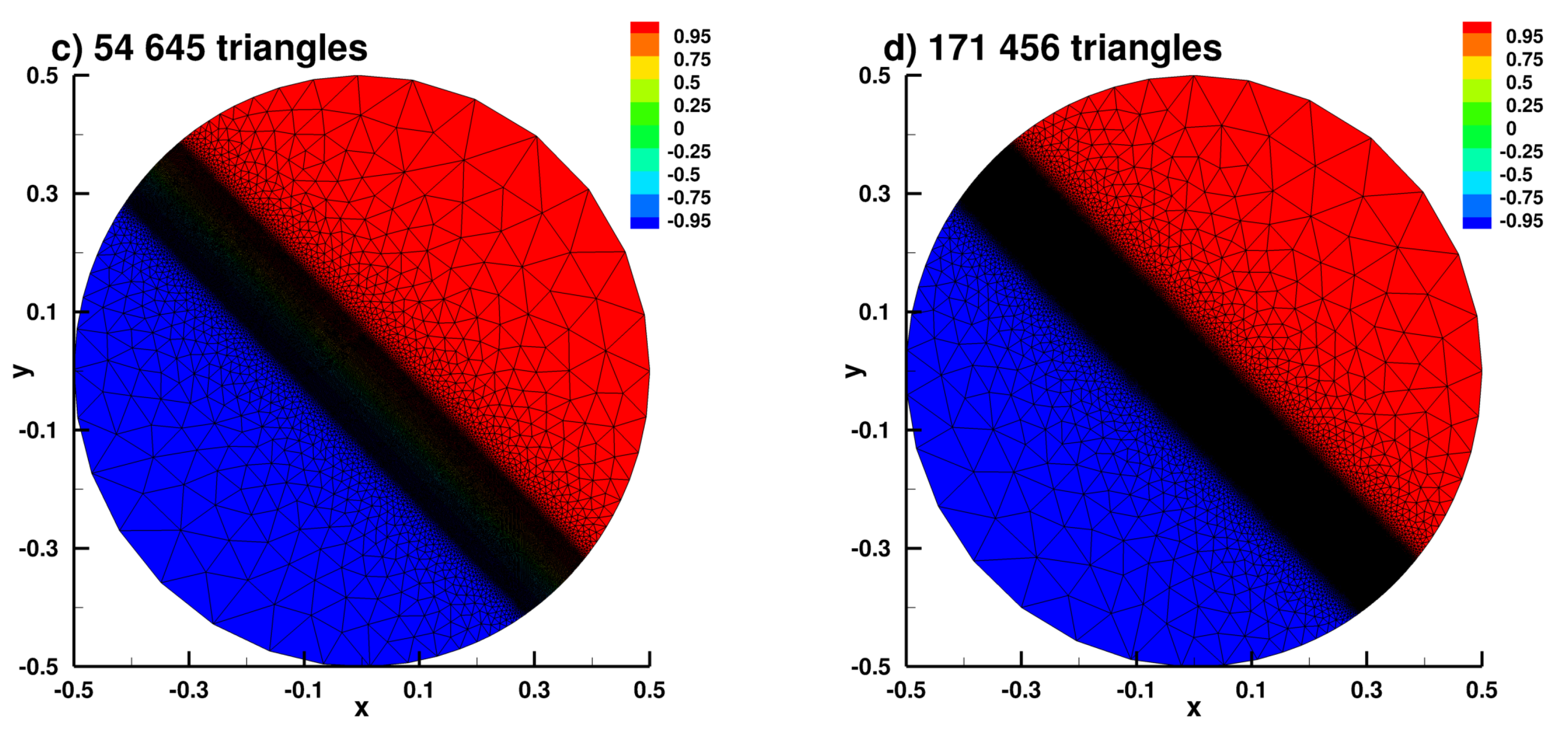
Appendix B.2. Laplace Equation Convergence Rate with Dirac Right Hand Side
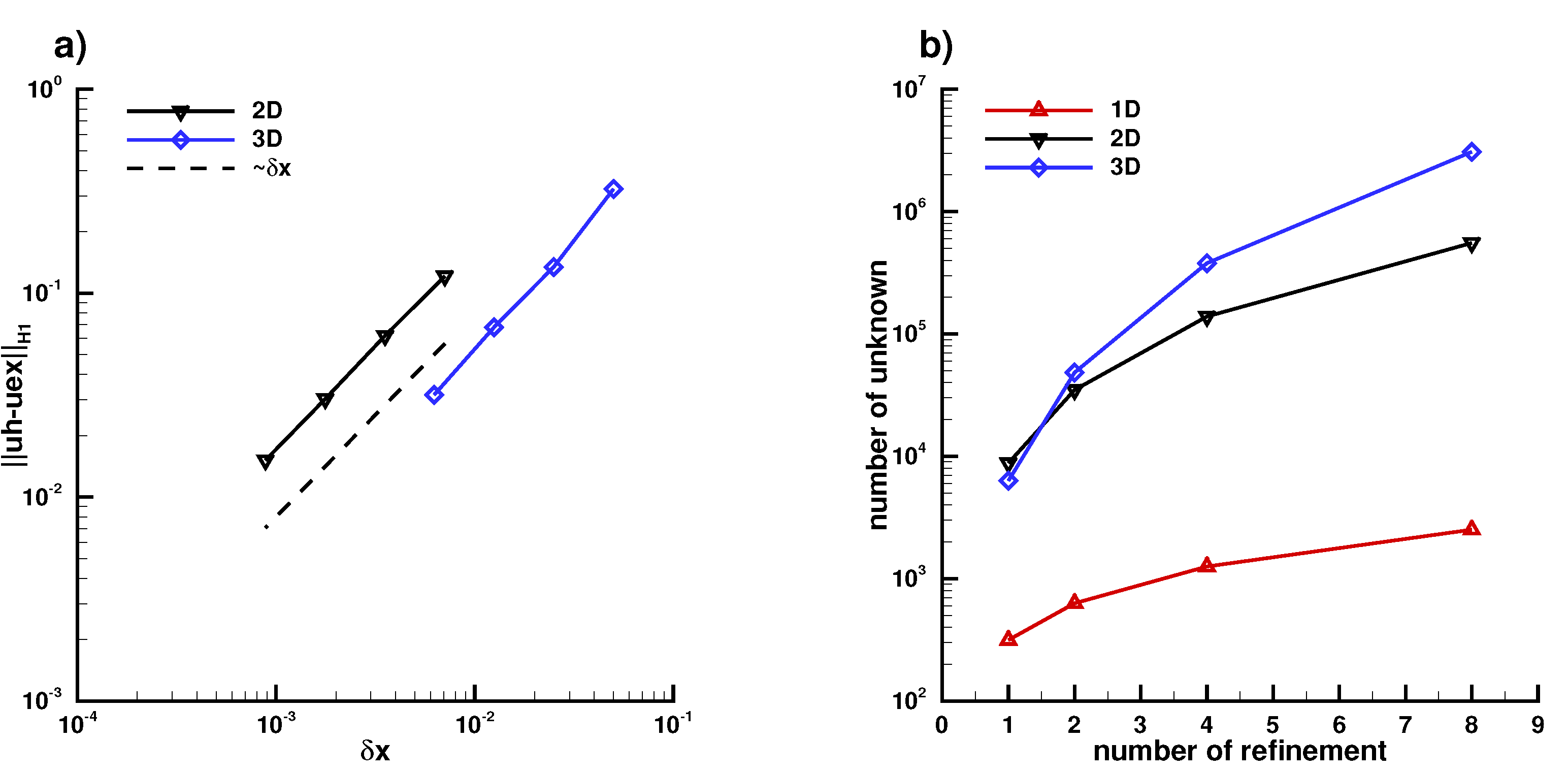


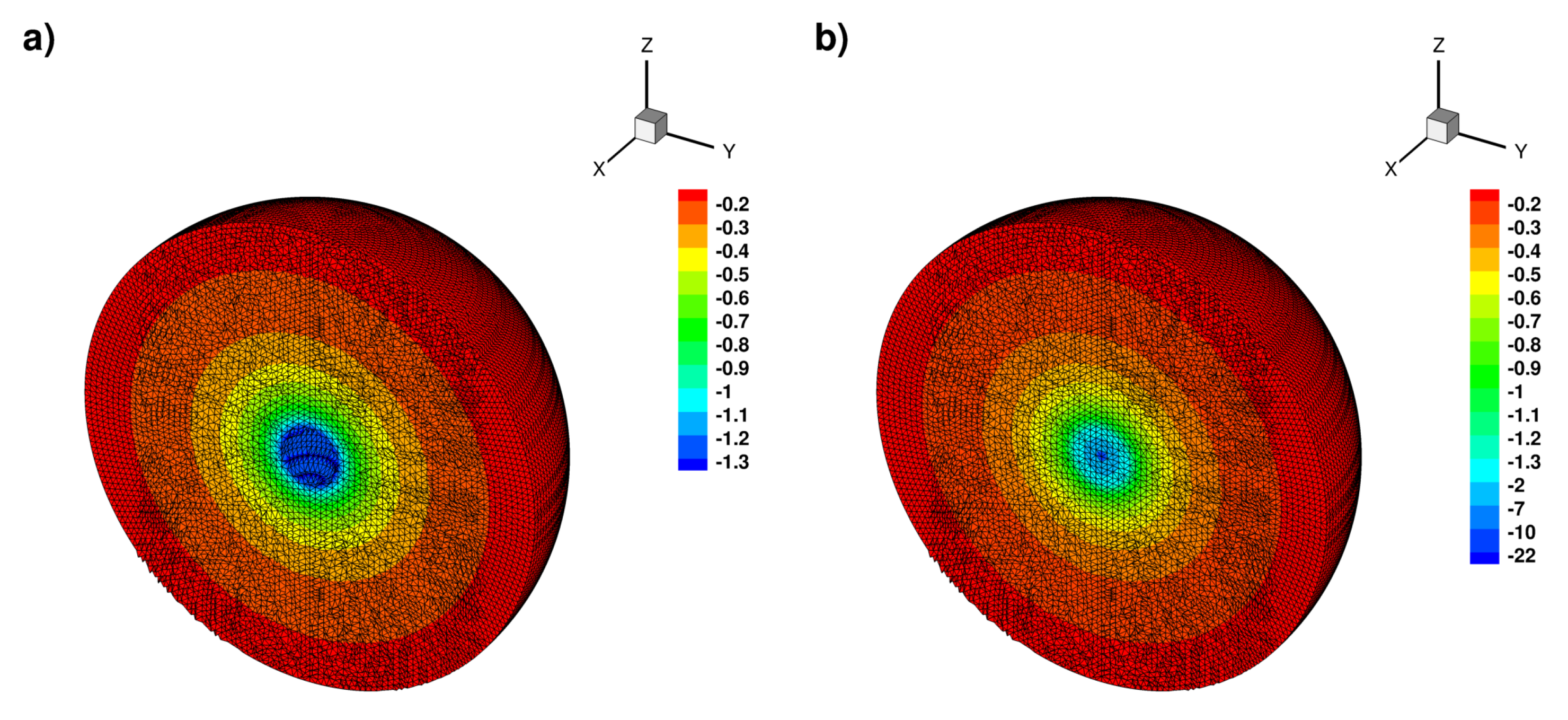
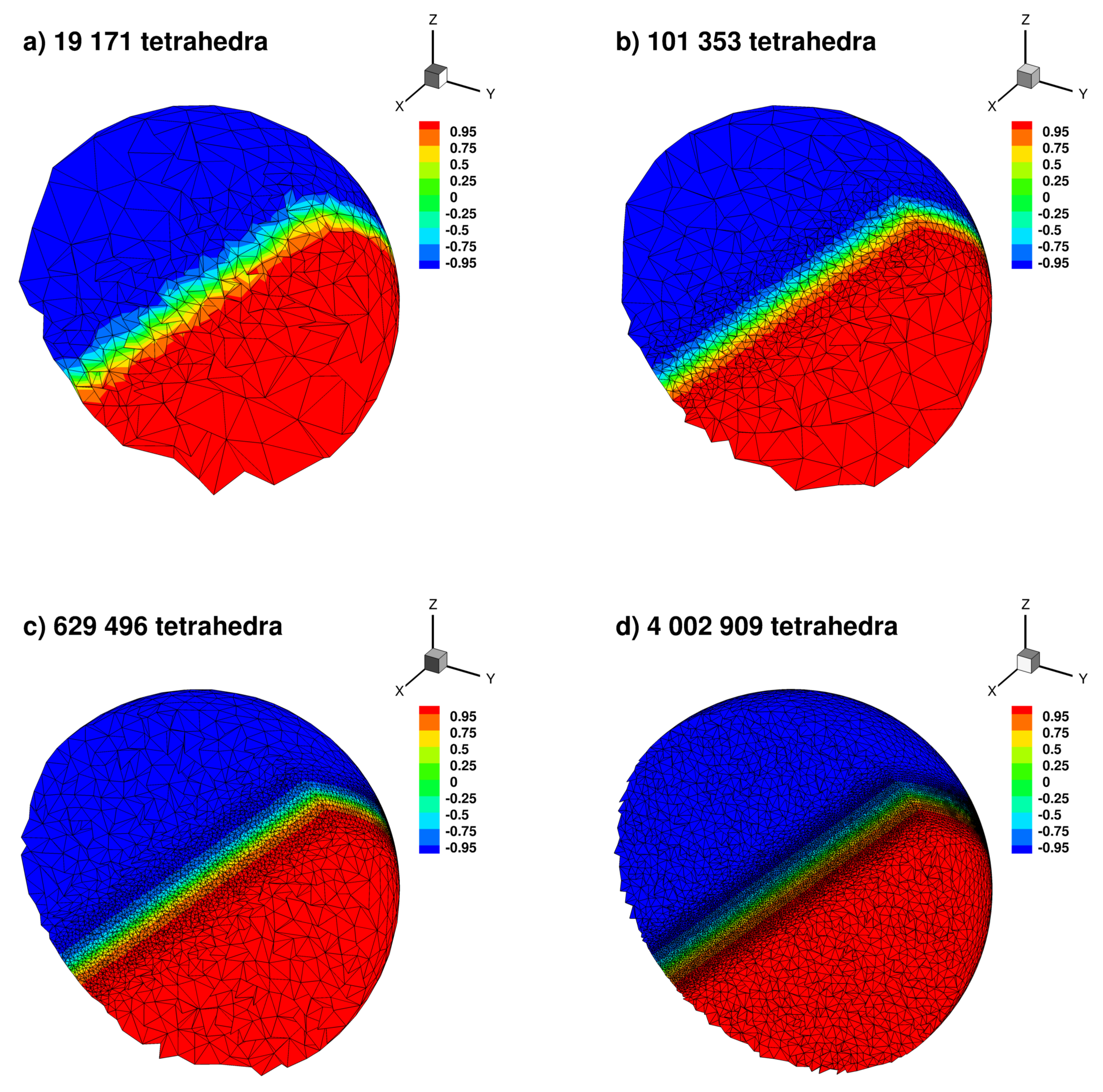
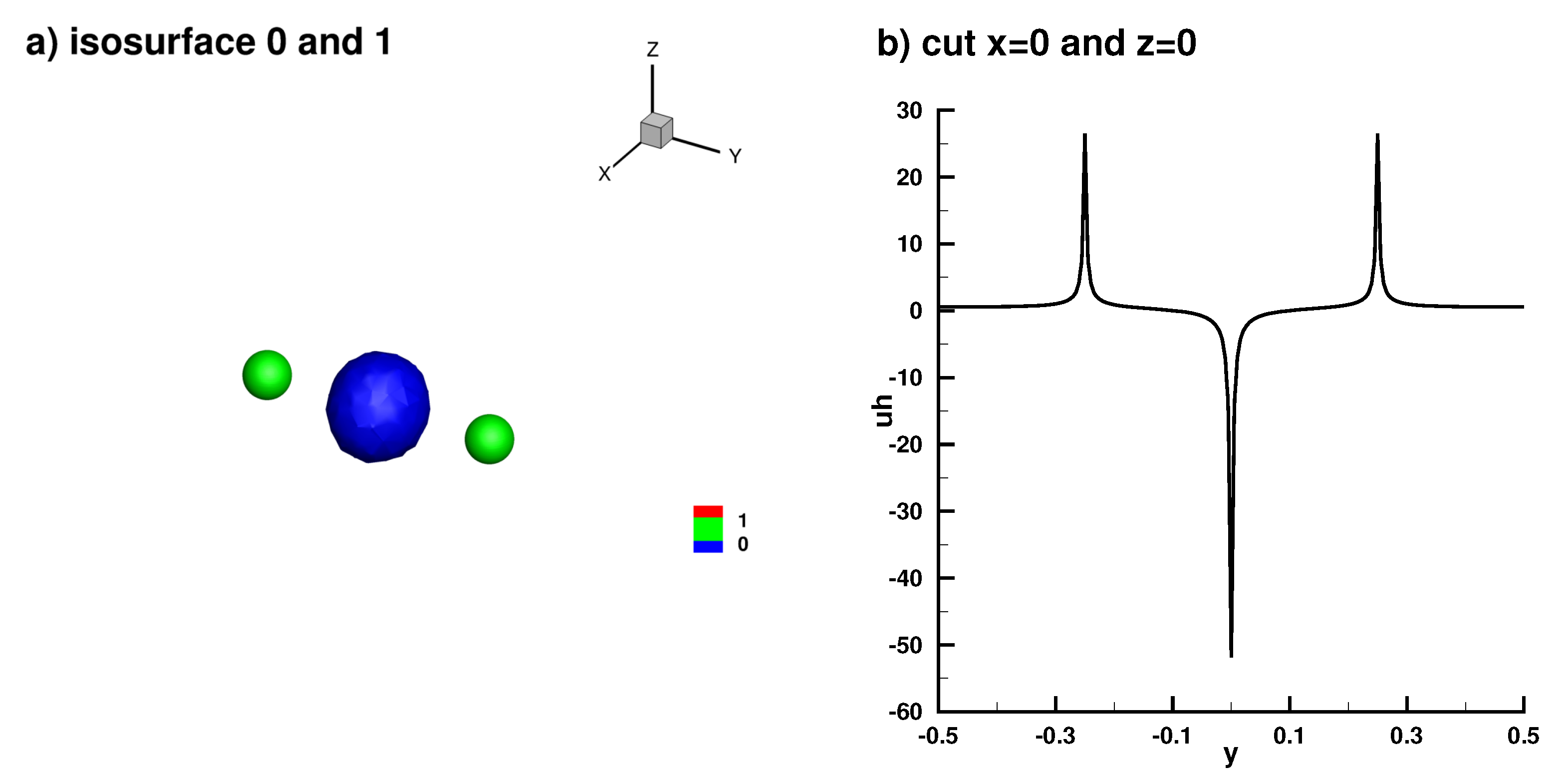
Appendix B.3. Laplace Equation with Dirac Right Hand Side





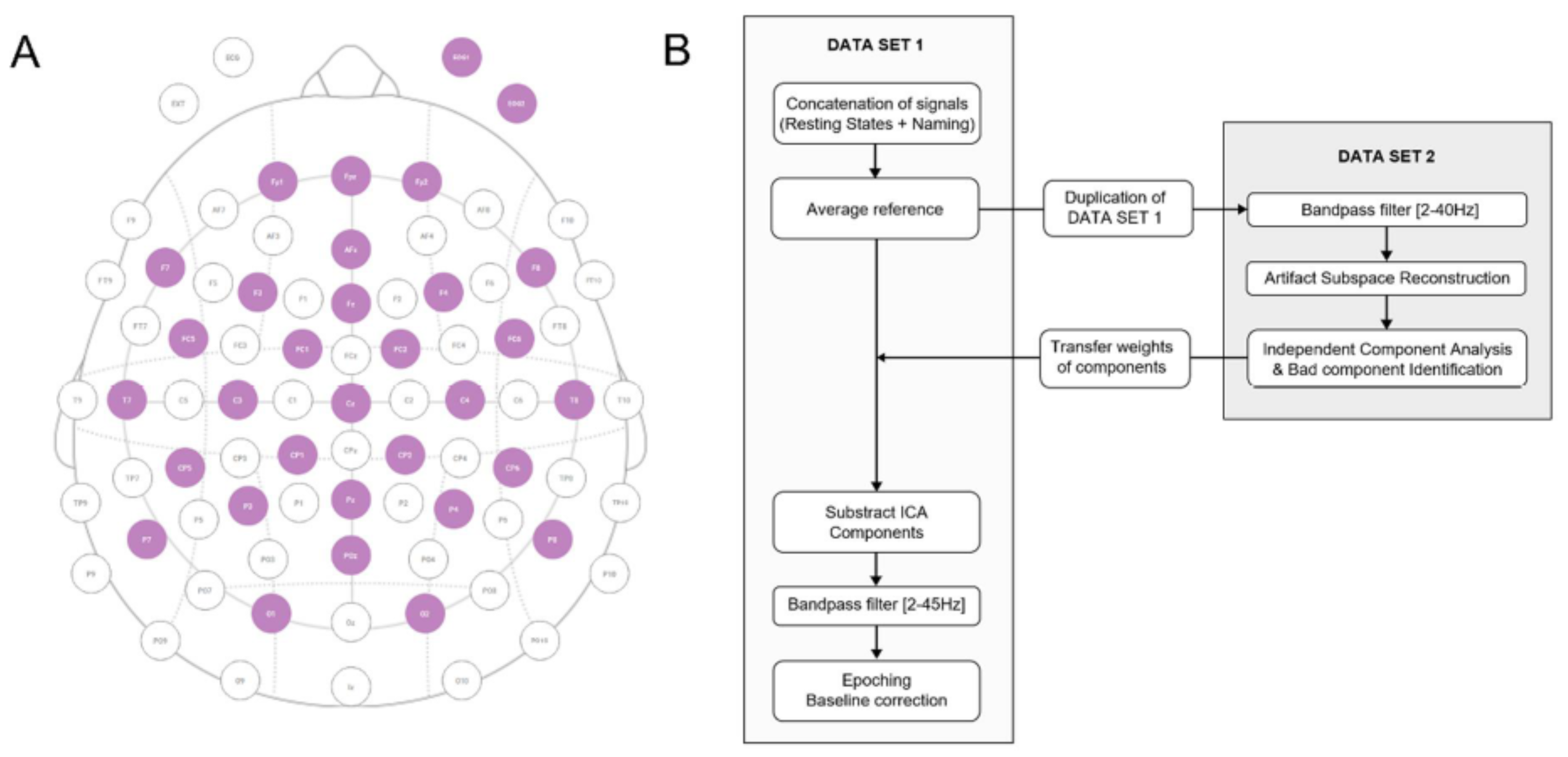
Appendix C. Data Preprocessing
References
- Adrian, E.D.; Yamagiwa, K. The origin of the Berger rhythm. Brain 1935, 58, 323–351. [Google Scholar] [CrossRef]
- Muller, L.; Chavane, F.; Reynolds, J.; Sejnowsk, T.J. Cortical travelling waves: Mechanisms and computational principles. Nat. Rev. Neurosci. 2018, 19, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Baese, L.; Watters, H.; Keilholz, S. Spatiotemporal patterns of spontaneous brain activity: A mini-review. Neurophotonics 2022, 9, 032209. [Google Scholar] [CrossRef]
- Klimesch, W.; Hanslmayr, S.; Sauseng, P.; Gruber, W.R.; Doppelmayr, M. P1 and Traveling Alpha Waves: Evidence for Evoked Oscillations. J. Neurophysiol. 2007, 97, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Patten, T.M.; Rennie, C.J.; Robinson, P.A.; Gong, P. Human Cortical Traveling Waves: Dynamical Properties and Correlations with Responses. PLoS ONE 2012, 7, e38392. [Google Scholar] [CrossRef] [PubMed]
- Massimini, M.; Huber, R.; Ferrarelli, F.; Hill, S.; Tononi, G. The Sleep Slow Oscillation as a Traveling Wave. J. Neurosci. 2004, 24, 6862–6870. [Google Scholar] [CrossRef] [PubMed]
- Alamia, A.; VanRullen, R. Alpha oscillations and traveling waves: Signatures of predictive coding? PLoS Biol. 2019, 17, e3000487. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.K.; Nauhaus, I.; Carandini, M. Traveling Waves in Visual Cortex Neuron 2012, 75, 218–229. 75.
- Zauner, A.; Gruber, W.; Himmelstoß, N.A.; Lechinger, J.; Klimesch, W. Lexical access and evoked traveling alpha waves. NeuroImage 2014, 91, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Burraco, A.; Murphy, E. Why Brain Oscillations Are Improving Our Understanding of Language. Front. Behav. Neurosci. 2019, 13, 190. [Google Scholar] [CrossRef]
- Alexander, D.M.; Jurica, P.; Trengove, C.; Nikolaev, A.R.; Gepshtein, S.; Zvyagintsev, M.; Mathiak, K.; Schulze-Bonhage, A.; Ruscher, J.; Ball, T.; et al. Traveling waves and trial averaging: The nature of single-trial and averaged brain responses in large-scale cortical signals. NeuroImage 2013, 73, 95–112. [Google Scholar] [CrossRef]
- Nir, Y.; Staba, R.J.; Andrillon, T.; Vyazovskiy, V.V.; Cirelli, C.; Fried, I.; Tononi, G. Regional Slow Waves and Spindles in Human Sleep. Neuron 2011, 70, 153–169. [Google Scholar] [CrossRef]
- Muller, L.; Piantoni, G.; Koller, D.; Cash, S.S.; Halgren, E.; Sejnowski, T.J. Rotating waves during human sleep spindles organize global patterns of activity that repeat precisely through the night. eLife 2016, 5, e17267. [Google Scholar] [CrossRef]
- Huang, X.; Xu, W.; Liang, J.; Takagaki, K.; Gao, X.; Wu, J. Spiral Wave Dynamics in Neocortex. Neuron 2010, 68, 978–990. [Google Scholar] [CrossRef]
- Lehmann, D.; Ozaki, H.; Pal, I. EEG alpha map series: Brain micro-states by space-oriented adaptive segmentation. Electroencephalogr. Clin. Neurophysiol. 1987, 67, 271–288. [Google Scholar] [CrossRef]
- Koenig, T.; Lehmann, D.; Merlo, M.C.G.; Kochi, K.; Hell, D.; Koukkou, M. A deviant EEG brain microstate in acute, neuroleptic-naive schizophrenics at rest. Eur. Arch. Psychiatry Clin. Neurosci. 1999, 249, 205–211. [Google Scholar] [CrossRef] [PubMed]
- De Menendez, R.G.; Murray, M.M.; Michel, C.M.; Martuzzi, R.; Andino, S.L.G. Electrical neuroimaging based on biophysical constraints. NeuroImage 2004, 21, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zotev, V.; Phillips, R.; Drevets, W.C.; Bodurka, J. Spatiotemporal dynamics of the brain at rest—Exploring EEG microstates as electrophysiological signatures of BOLD resting state networks. NeuroImage 2012, 60, 2062–2072. [Google Scholar] [CrossRef] [PubMed]
- Brechet, L.; Brunet, D.; Birot, G.; Gruetter, R.; Michel, C.M.; Jorge, J. Capturing the spatiotemporal dynamics of self-generated, task-initiated thoughts with EEG and fMRI. NeuroImage 2019, 194, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Benquet, P.; Biraben, A.; Berrou, C.; Dufor, O.; Wendling, F. Dynamic reorganization of functional brain networks during picture naming. Cortex 2015, 73, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Grappe, A.; Sarma, S.V.; Sacre, P.; Gonzalez-Martınez, J.; Liegeois-Chauvel, C.; Alario, F.-X. An intracerebral exploration of functional connectivity during word production. J. Comput. Neurosci. 2018, 46, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Hartwigsen, G.; Stockert, A.; Charpentier, L.; Wawrzyniak, M.; Klingbeil, J.; Wrede, K.; Obrig, H.; Saur, D. Short-term modulation of the lesioned language network. eLife 2020, 9, e54277. [Google Scholar] [CrossRef]
- Laganaro, M.; Python, G.; Toepel, U. Dynamics of phonological–phonetic encoding in word production: Evidence from diverging ERPs between stroke patients and controls. Brain Lang. 2013, 126, 123–132. [Google Scholar] [CrossRef]
- Mheich, A.; Dufor, O.; Yassine, S.; Kabbara, A.; Biraben, A.; Wendling, F.; Hassan, M. HD-EEG for tracking sub-second brain dynamics during cognitive tasks. Sci. Data 2021, 8, 32. [Google Scholar] [CrossRef]
- Hooi, L.S.; Nisar, H.; Voon, Y.V. Tracking of EEG Activity using Topographic Maps. In Proceedings of the 2015 IEEE International Conference on Signal and Image Processing Applications (ICSIPA), Kuala Lumpur, Malaysia, 19–21 October 2015. [Google Scholar]
- Wackermann, J.; Lehmann, D.; Michel, C.M.; Strik, W.K. Adaptive segmentation of spontaneous EEG map series into spatially defined microstates. Int. J. Psychophysiol. 1993, 14, 269–283. [Google Scholar] [CrossRef]
- Buzsaki, G.; Anastassiou, C.A.; Koch, C. The origin of extracellular fields and currents—EEG, ECoG, LFP and spikes. Nat. Rev. Neurosci. 2016, 13, 407–420. [Google Scholar] [CrossRef]
- Jatoi, M.A.; Kamel, N. Brain Source Localization Using EEG Signal Analysis; Taylor & Francis Group: Boca Raton, FL, USA, 2018. [Google Scholar]
- Saturnino, G.B.; Siebner, H.R.; Thielscher, A.; Madsen, K.H. Accessibility of cortical regions to focal tes: Dependence on spatial position, safety, and practical constraints. NeuroImage 2019, 203, 116183. [Google Scholar] [CrossRef] [PubMed]
- Saturnino, G.B.; Madsen, K.H.; Siebner, H.R.; Thielscher, A. How to target inter-regional phase synchronization with dual-site Transcranial Alternating Current Stimulation. NeuroImage 2017, 163, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Saturnino, G.B.; Puonti, O.; Nielsen, J.D.; Antonenko, D.; Madsen, K.H.; Thielscher, A. Simnibs 2.1: A comprehensive pipeline for individualized electric field modelling for transcranial brain stimulation. In Brain and Human Body Modeling; Makarov, S., Horner, M., Noetscher, G., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Volpert, V.; Xu, B.; Tchechmedjiev, A.; Harispe, S.; Aksenov, A.; Mesnildrey, Q.; Beuter, A. Characterization of spatiotemporal dynamics in EEG data during picture naming with optical flow patterns. Math. Biosci. Eng. 2023, 20, 11429–11463. [Google Scholar] [CrossRef] [PubMed]
- Aksenov, A.; Renaud-D’Ambra, M.; Volpert, V.; Beuter, A. Phase-shifted tACS can modulate cortical alpha waves in human subjects. Cogn. Neurodynamics 2023, in press. [Google Scholar] [CrossRef]
- Lehmann, D.; Skrandies, W. Reference-free identification of components of checkerboardevoked multichannel potential fields. Electroencephalogr. Clin. Neurophysiol. 1980, 48, 609–621. [Google Scholar] [CrossRef]
- Lehmann, D.; Skrandies, W. Spatial Analysis of Evoked Potentials in Man—A review. Prog. Neurobiol. 1984, 23, 227–250. [Google Scholar] [CrossRef]
- Murray, M.M.; Brunet, D.; Michel, C.M. Topographic ERP analyses: A step-by-step tutorial review. Brain Topogr. 2008, 20, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Chantsoulis, M.; Polrola, P.; Goral-Polrola, J.; Hajdukiewicz, A.; Supinski, J.; Kropotov, J.D.; Pachalska, M. Application of ERPs neuromarkers for assessment and treatment of a patient with chronic crossed aphasia after severe TBI and long-term coma—Case report. Ann. Agric. Environ. Med. 2017, 24, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Laganaro, M. Inter-study and inter-Individual Consistency and Variability of EEG/ERP Microstate Sequences in Referential Word Production. Brain Topogr. 2017, 30, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Ermentrout, G.B.; Kleinfeld, D. Traveling Electrical Waves in Cortex: Insights from Phase Dynamics and Speculation on a Computational Role the results. Neuron 2001, 29, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Watrous, A.; Patel, A.; Jacobs, J. Theta and Alpha Oscillations Are Traveling Waves in the Human Neocortex. Neuron 2018, 98, 1269–1281. [Google Scholar] [CrossRef] [PubMed]
- Mheich, A.; Hassan, M.; Khalil, M.; Berrou, C.; Wendling, F. A new algorithm for spatiotemporal analysis of brain functional connectivity. J. Neurosci. Methods 2015, 242, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Hecht, F. New developments in Freefem++. Journal of Numerical Mathematics 2012, 20, 251–266. [Google Scholar] [CrossRef]
- Dapogny, C.; Dobrzynski, C.; Frey, P. Three-dimensional adaptive domain remeshing, implicit domain meshing, and applications to free and moving boundary problems. J. Comput. Phys. 2014, 262, 358–378. [Google Scholar] [CrossRef]
- Scott, R. Finite element convergence for singular data. J. Numer. Math. 1973, 21, 317–327. [Google Scholar] [CrossRef]
- Bradji, A.; Holzbecher, E. On the Convergence Order in Sobolev Norms of COMSOL Solutions; WIAS: Berlin, Germany, 2008. [Google Scholar]
- Brainard, D.H. The Psychophysics Toolbox; Brill: Leiden, The Netherlands, 1997. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed]
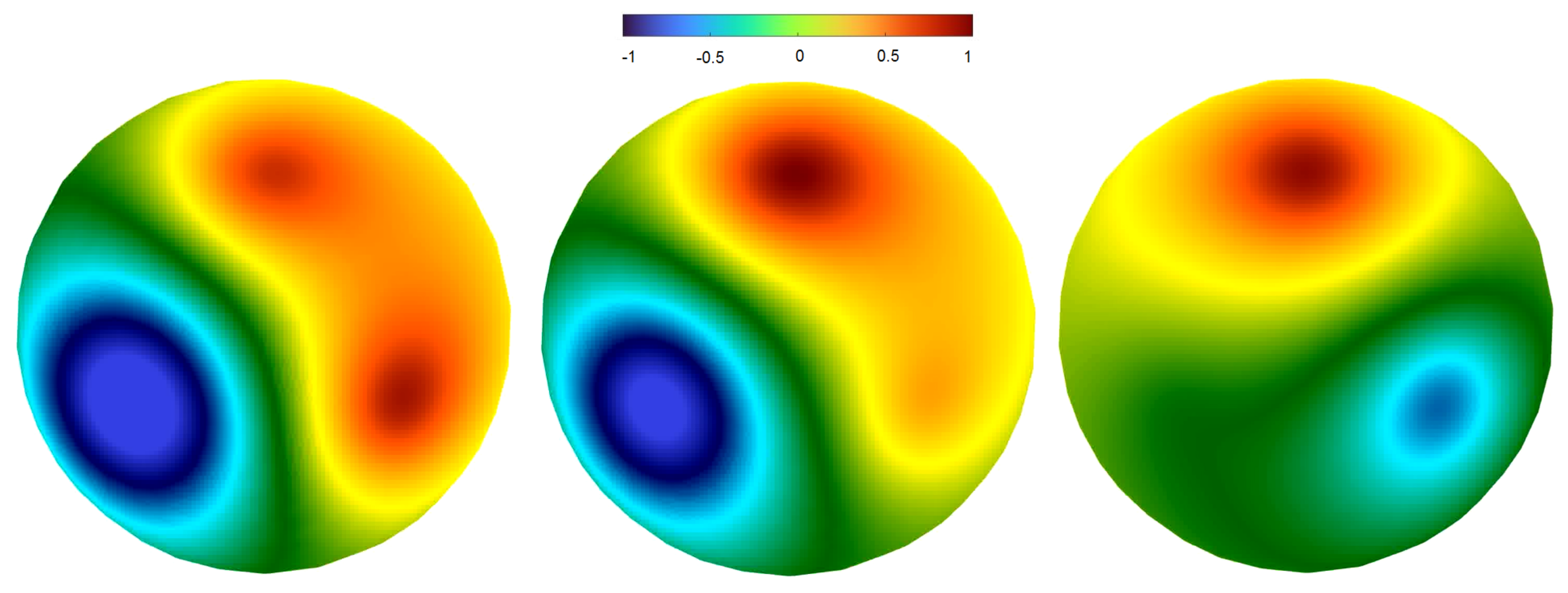

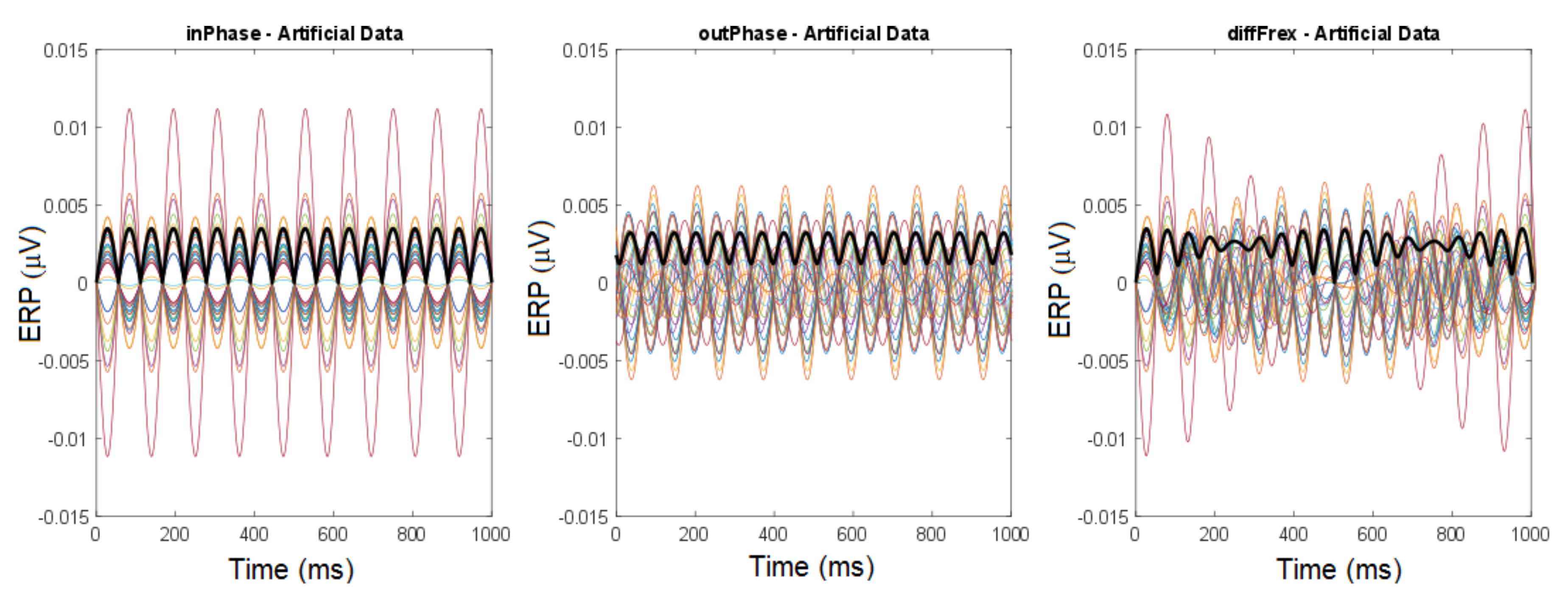
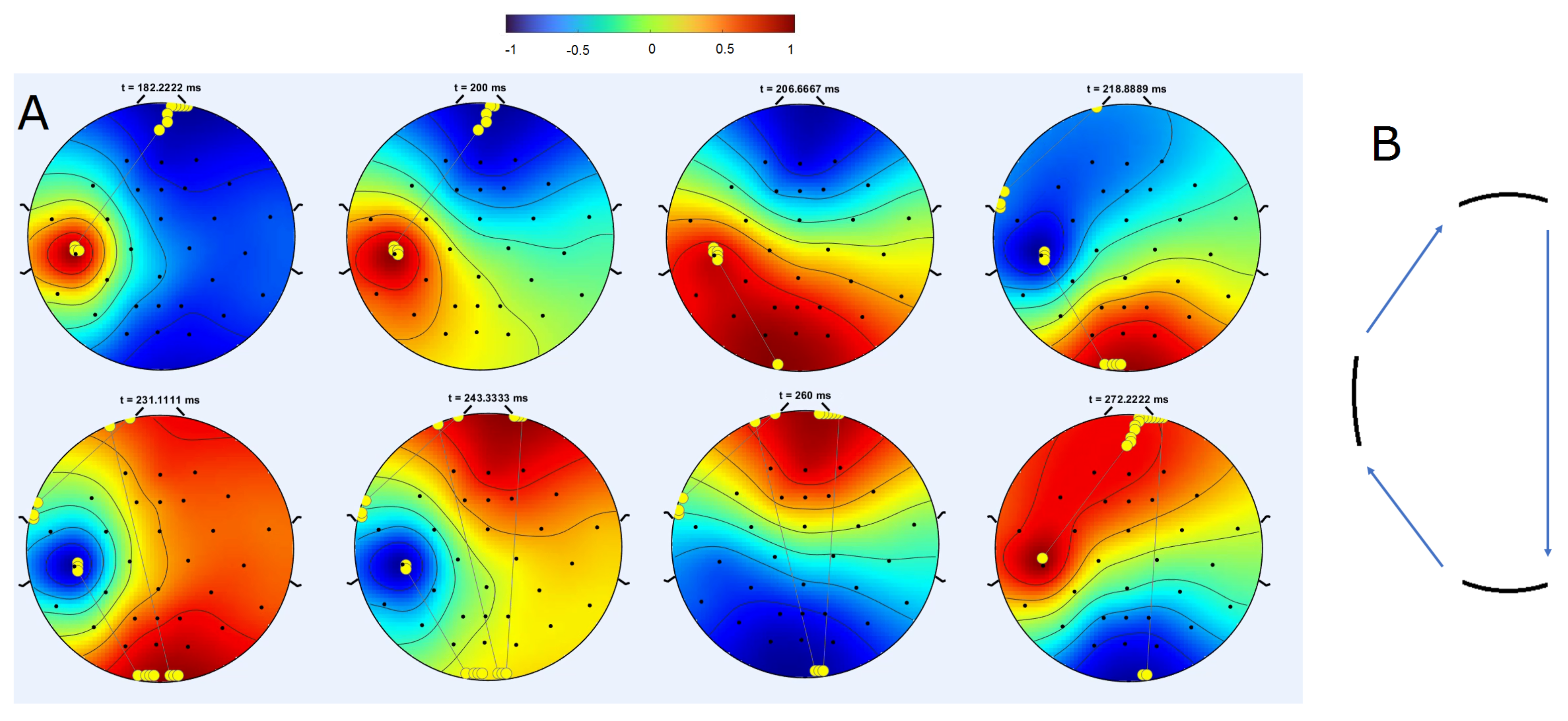
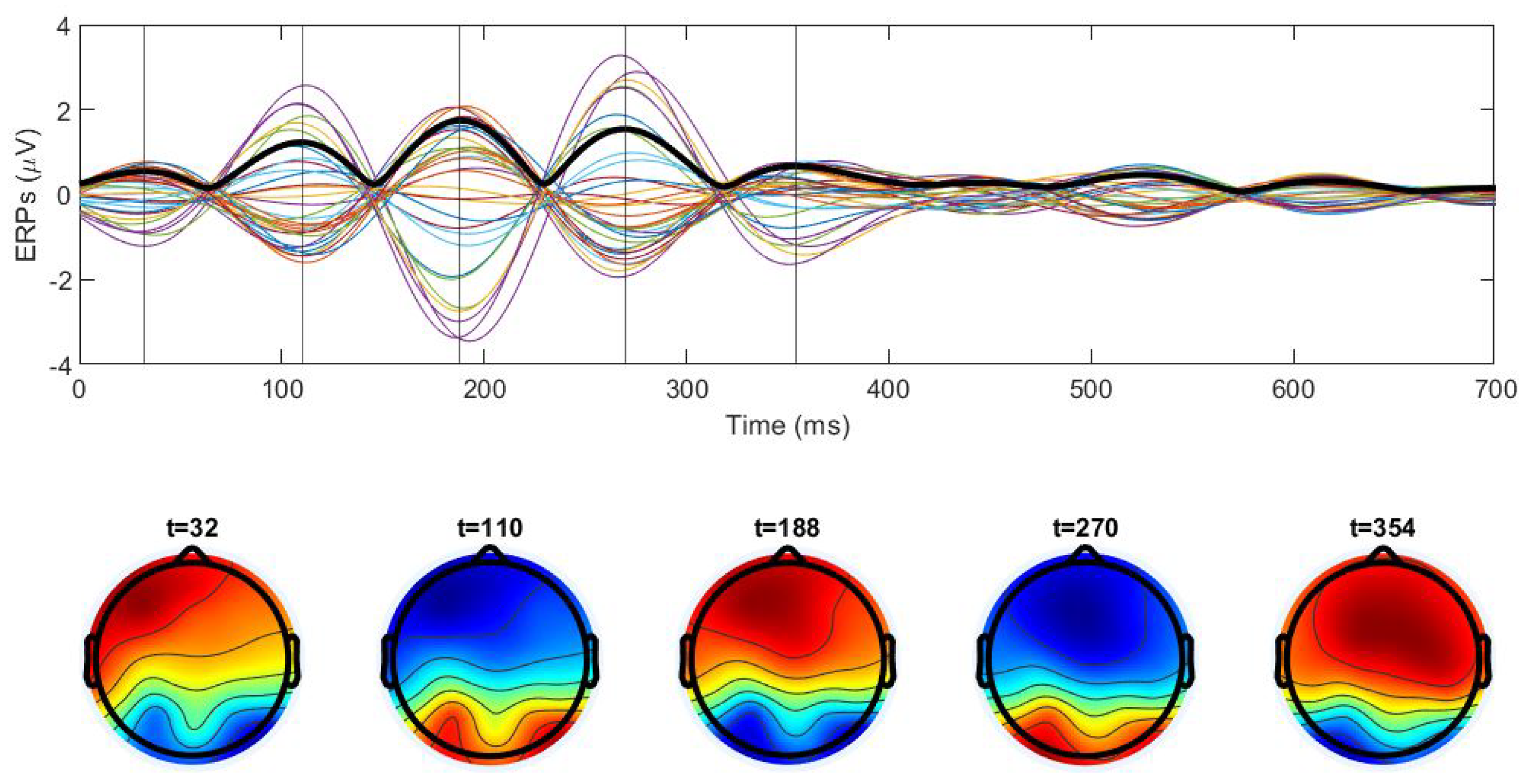




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Volpert, V.; Sadaka, G.; Mesnildrey, Q.; Beuter, A. Modelling EEG Dynamics with Brain Sources. Symmetry 2024, 16, 189. https://doi.org/10.3390/sym16020189
Volpert V, Sadaka G, Mesnildrey Q, Beuter A. Modelling EEG Dynamics with Brain Sources. Symmetry. 2024; 16(2):189. https://doi.org/10.3390/sym16020189
Chicago/Turabian StyleVolpert, Vitaly, Georges Sadaka, Quentin Mesnildrey, and Anne Beuter. 2024. "Modelling EEG Dynamics with Brain Sources" Symmetry 16, no. 2: 189. https://doi.org/10.3390/sym16020189
APA StyleVolpert, V., Sadaka, G., Mesnildrey, Q., & Beuter, A. (2024). Modelling EEG Dynamics with Brain Sources. Symmetry, 16(2), 189. https://doi.org/10.3390/sym16020189