
Wastewater Treatment Using Photosynthetic Microorganisms



Abstract
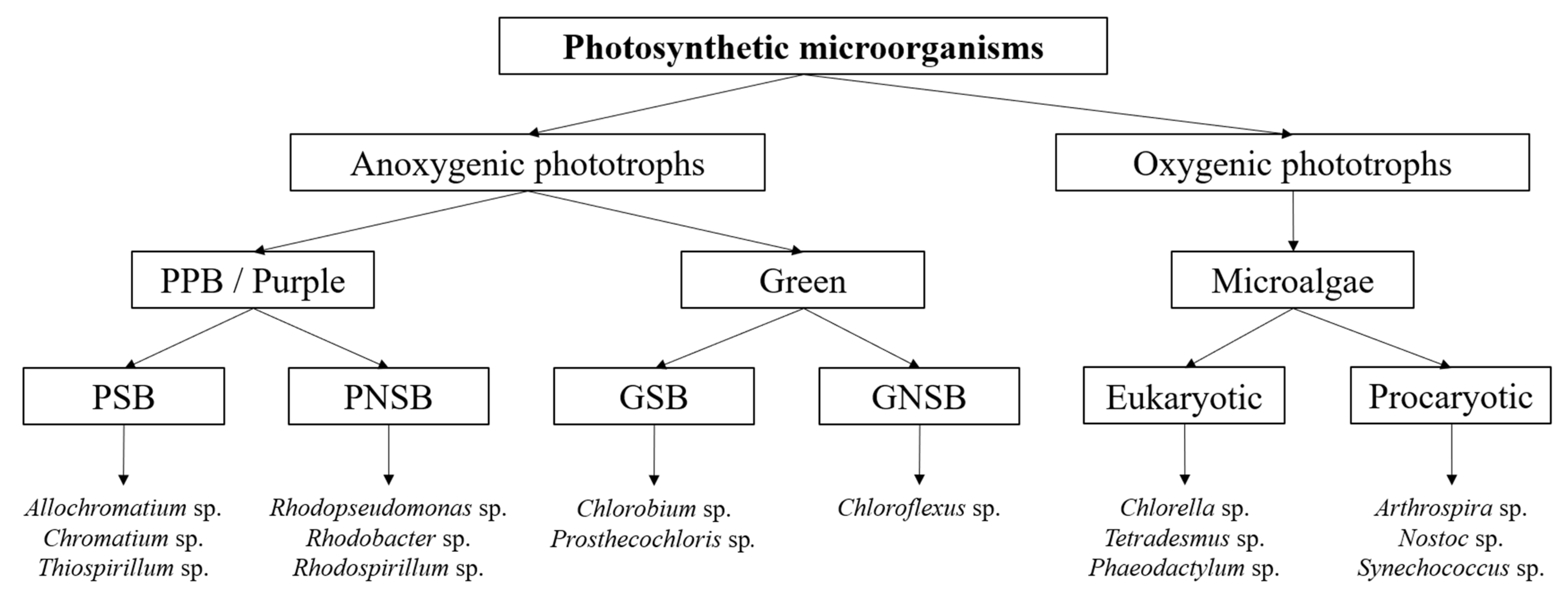
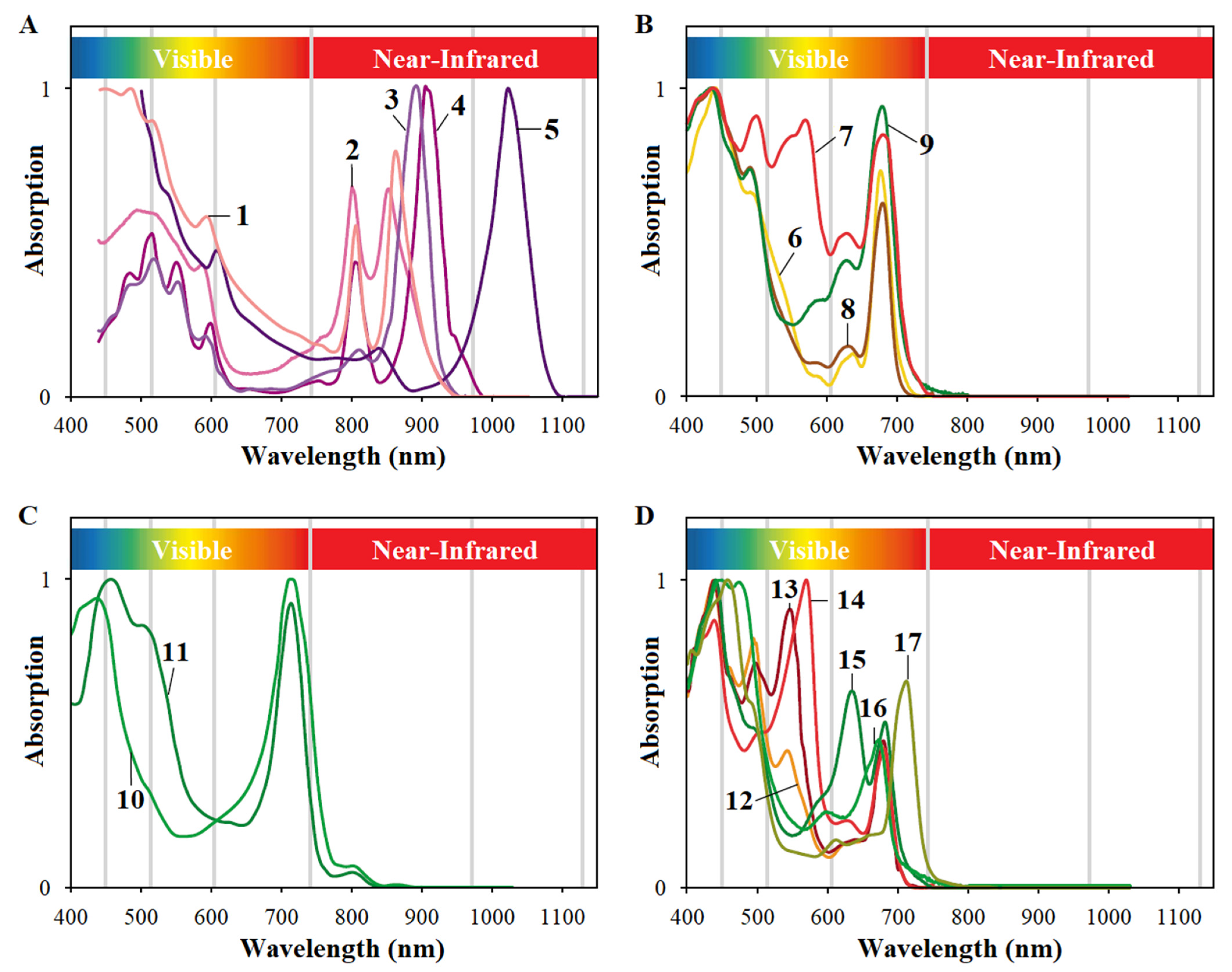
1. Introduction
2. Piggery Wastewater

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Farm | COD (mg L−1) | TN (mg L−1) | TP (mg L−1) | TSS (mg L−1) | Reference |
|---|---|---|---|---|---|
| Daejeon (South Korea) | 18,700 | 810 | 290 | - | [18] |
| Barcelona (Spain) | 7450 | 785 | 120 | 3100 | [19] |
| Yokohama (Japan) | 5300 | 1270 | - | - | [20] |
| Castilla y León (Spain) | 54,000 | 5000 | 1500 | - | [8] |
| Seosan-si (South Korea) | 8420 | 1150 | 34 | - | [21] |
| Santiago (Chile) | 18,400 | 1085 | 172 | - | [22] |
| Queensland (Australia) | 4130 | 1160 | 160 | 2420 | [3] |
3. Conventional Piggery Wastewater Treatment Technologies
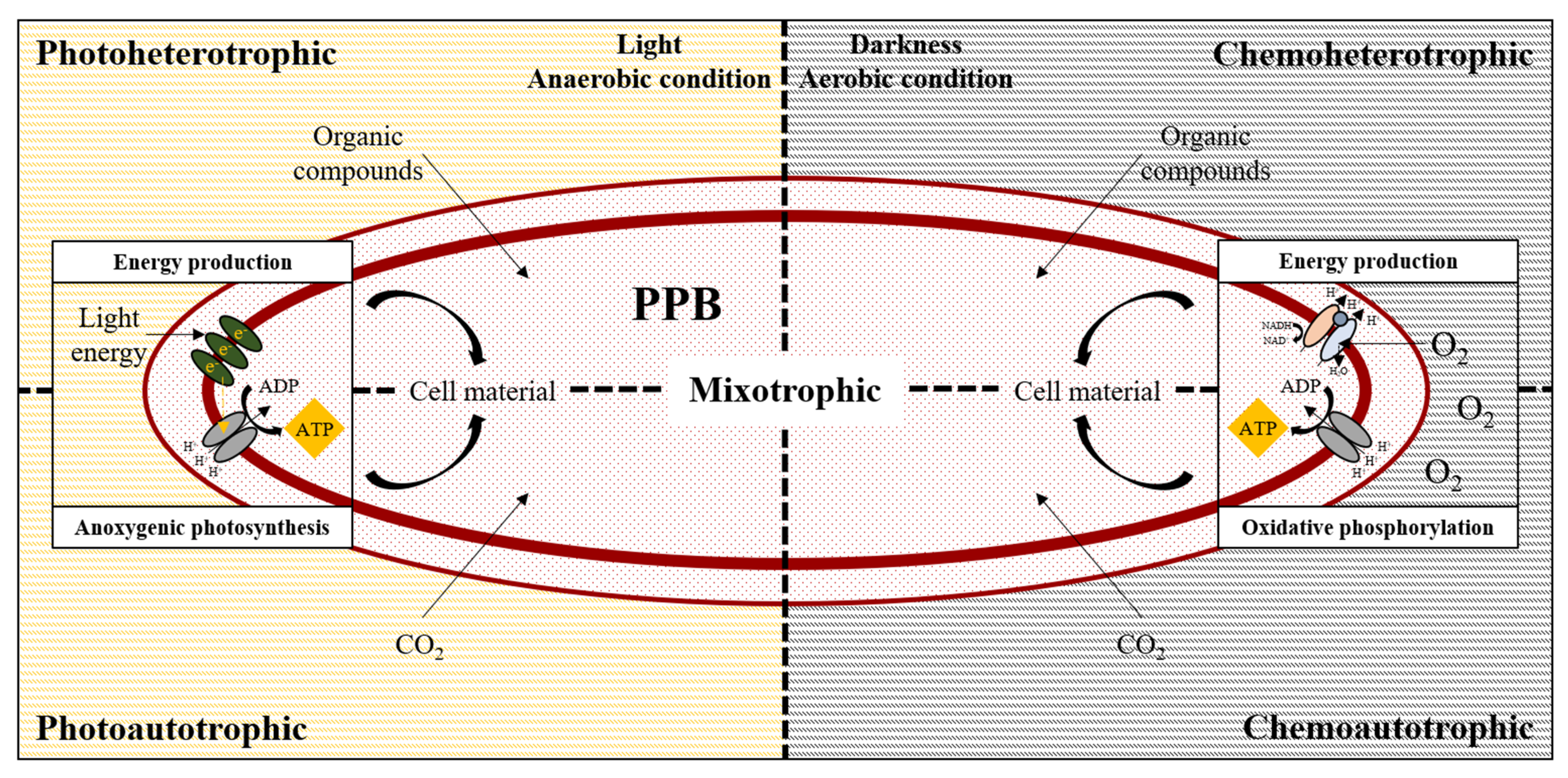
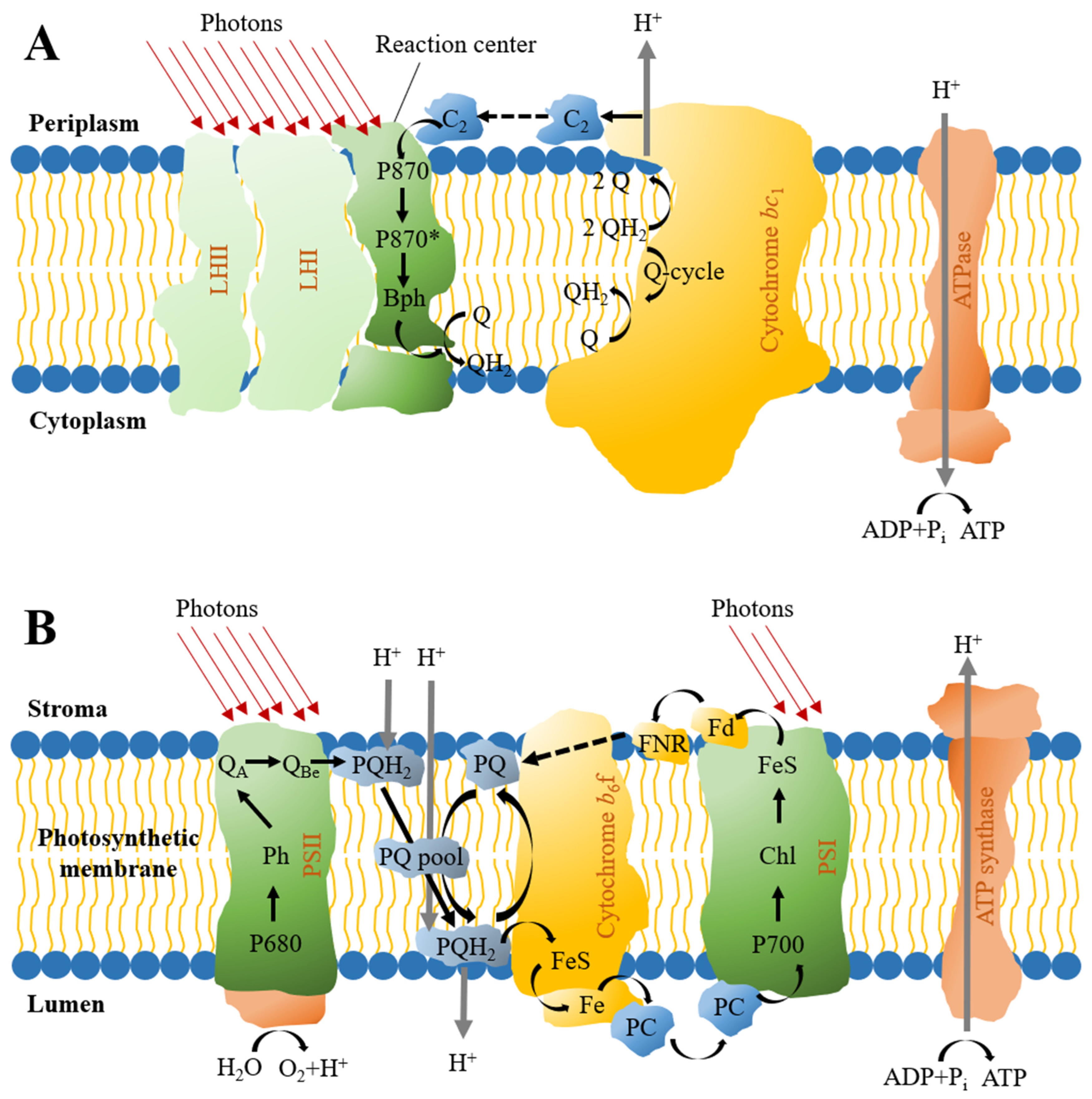
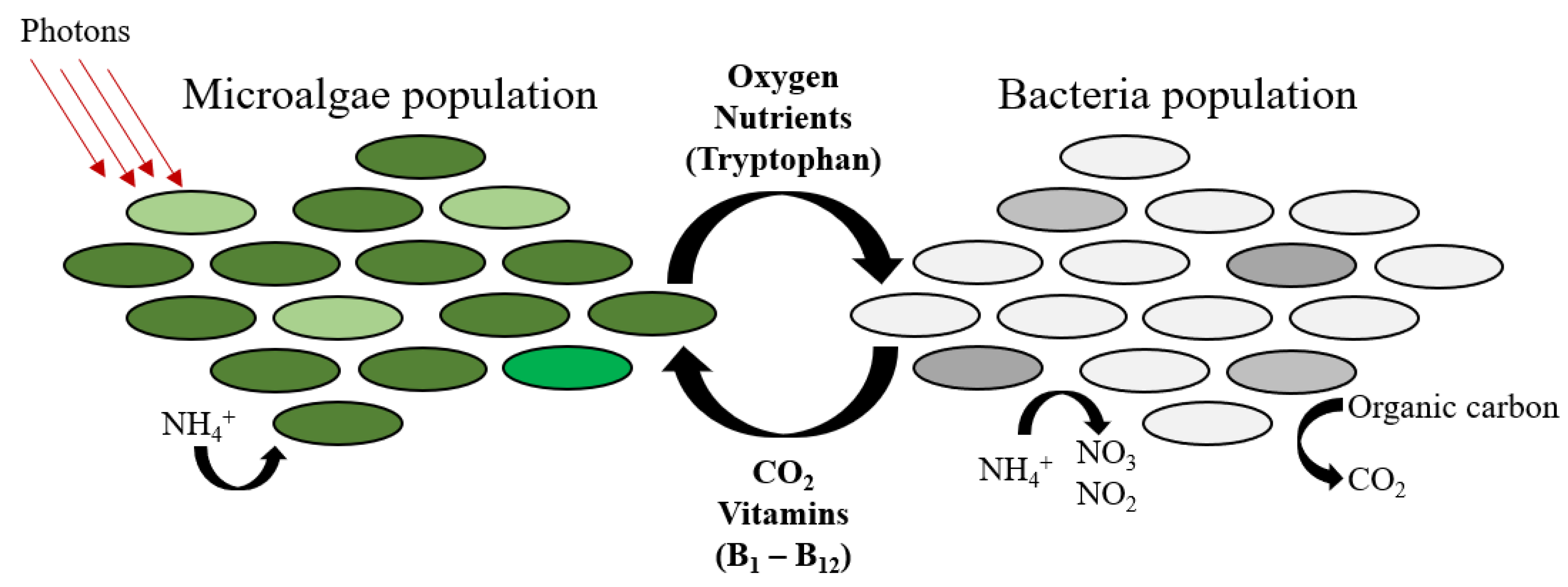
4. Photosynthetic Piggery Wastewater Treatment
4.1. PPB-Based Treatment
4.2. Microalgae-Based Treatment
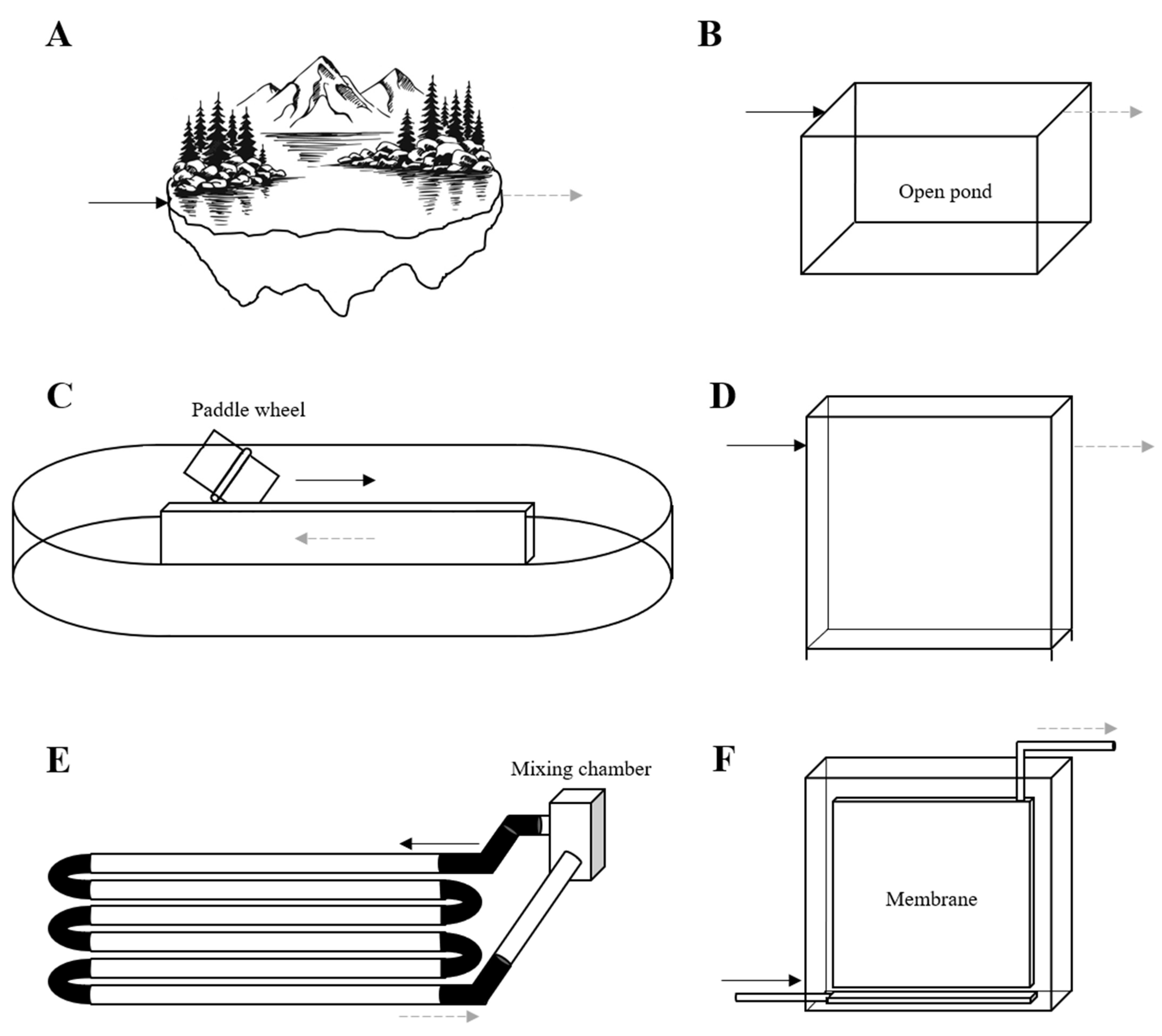
5. Photobioreactors for Wastewater Treatment
5.1. Open Photobioreactors
5.2. Closed Photobioreactors
6. Biomass Valorisation
7. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects 2019: Highlights; United Nations: New York, NY, USA, 2019; ISBN 978-92-1-148316-1. [Google Scholar]
- Cai, T.; Park, S.Y.; Li, Y. Nutrient Recovery from Wastewater Streams by Microalgae: Status and Prospects. Renew. Sustain. Energy Rev. 2013, 19, 360–369. [Google Scholar] [CrossRef]
- Hülsen, T.; Stegman, S.; Batstone, D.J.; Capson-Tojo, G. Naturally Illuminated Photobioreactors for Resource Recovery from Piggery and Chicken-Processing Wastewaters Utilising Purple Phototrophic Bacteria. Water Res. 2022, 214, 118194. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of Anaerobic Digestion Process: A Review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef] [PubMed]
- Aditya, L.; Mahlia, T.M.I.; Nguyen, L.N.; Vu, H.P.; Nghiem, L.D. Microalgae-Bacteria Consortium for Wastewater Treatment and Biomass Production. Sci. Total Environ. 2022, 838, 155871. [Google Scholar] [CrossRef] [PubMed]
- Hülsen, T.; Batstone, D.J.; Keller, J. Phototrophic Bacteria for Nutrient Recovery from Domestic Wastewater. Water Res. 2014, 50, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Zhang, G.; Dai, X.; Schideman, L.; Zhang, Y.; Li, B.; Wang, H. A Novel Wastewater Treatment and Biomass Cultivation System Combining Photosynthetic Bacteria and Membrane Bioreactor Technology. Desalination 2013, 322, 176–181. [Google Scholar] [CrossRef]
- Godos, I.d; Blanco, S.; García-Encina, P.A.; Becares, E.; Muñoz, R. Long-Term Operation of High Rate Algal Ponds for the Bioremediation of Piggery Wastewaters at High Loading Rates. Bioresour. Technol. 2009, 100, 4332–4339. [Google Scholar] [CrossRef]
- Sepúlveda-Muñoz, C.A.; Torres-Franco, A.F.; de Godos, I.; Muñoz, R. Exploring the Metabolic Capabilities of Purple Phototrophic Bacteria during Piggery Wastewater Treatment. J. Water Process Eng. 2022, 50, 103317. [Google Scholar] [CrossRef]
- Jiménez-Ruiz, S.; Laguna, E.; Vicente, J.; García-Bocanegra, I.; Martínez-Guijosa, J.; Cano-Terriza, D.; Risalde, M.A.; Acevedo, P. Characterization and Management of Interaction Risks between Livestock and Wild Ungulates on Outdoor Pig Farms in Spain. Porc. Health Manag. 2022, 8, 2. [Google Scholar] [CrossRef]
- Gilbert, M.; Nicolas, G.; Cinardi, G.; Van Boeckel, T.P.; Vanwambeke, S.O.; Wint, G.R.W.; Robinson, T.P. Global Distribution Data for Cattle, Buffaloes, Horses, Sheep, Goats, Pigs, Chickens and Ducks in 2010. Sci. Data 2018, 5, 180227. [Google Scholar] [CrossRef]
- Godos, I.d; Vargas, V.A.; Blanco, S.; González, M.C.G.; Soto, R.; García-Encina, P.A.; Becares, E.; Muñoz, R. A Comparative Evaluation of Microalgae for the Degradation of Piggery Wastewater under Photosynthetic Oxygenation. Bioresour. Technol. 2010, 101, 5150–5158. [Google Scholar] [CrossRef]
- Chen, G.; Huang, J.; Tian, X.; Chu, Q.; Zhao, Y.; Zhao, H. Effects of Influent Loads on Performance and Microbial Community Dynamics of Aerobic Granular Sludge Treating Piggery Wastewater. J. Chem. Technol. Biotechnol. 2018, 93, 1443–1452. [Google Scholar] [CrossRef]
- Sepúlveda-Muñoz, C.A.; Ángeles, R.; de Godos, I.; Muñoz, R. Comparative Evaluation of Continuous Piggery Wastewater Treatment in Open and Closed Purple Phototrophic Bacteria-Based Photobioreactors. J. Water Process Eng. 2020, 38, 101608. [Google Scholar] [CrossRef]
- Portela-Monge, C.; Bolado, S.; López-Serna, R.; Jiménez, J.J. Determination of Contaminants of Emerging Concern in Raw Pig Manure as a Whole: Difference with the Analysis of Solid and Liquid Phases Separately. Environ. Sci. Process. Impacts 2022, 87, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.L.; Yang, L.; Huang, K.X.; Chen, D.Z.; Gao, F. Mechanisms and Application of Microalgae on Removing Emerging Contaminants from Wastewater: A Review. Bioresour. Technol. 2022, 364, 128049. [Google Scholar] [CrossRef] [PubMed]
- López-Serna, R.; García, D.; Bolado, S.; Jiménez, J.J.; Lai, F.Y.; Golovko, O.; Gago-Ferrero, P.; Ahrens, L.; Wiberg, K.; Muñoz, R. Photobioreactors Based on Microalgae-Bacteria and Purple Phototrophic Bacteria Consortia: A Promising Technology to Reduce the Load of Veterinary Drugs from Piggery Wastewater. Sci. Total Environ. 2019, 692, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Myung, K.K.; Choi, K.M.; Yin, C.R.; Lee, K.Y.; Im, W.T.; Ju, H.L.; Lee, S.T. Odorous Swine Wastewater Treatment by Purple Non-Sulfur Bacteria, Rhodopseudomonas palustris, Isolated from Eutrophicated Ponds. Biotechnol. Lett. 2004, 26, 819–822. [Google Scholar] [CrossRef]
- Obaja, D.; MacÉ, S.; Mata-Alvarez, J. Biological Nutrient Removal by a Sequencing Batch Reactor (SBR) Using an Internal Organic Carbon Source in Digested Piggery Wastewater. Bioresour. Technol. 2005, 96, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Joo, H.S.; Hirai, M.; Shoda, M. Piggery Wastewater Treatment Using Alcaligenes faecalis Strain No. 4 with Heterotrophic Nitrification and Aerobic Denitrification. Water Res. 2006, 40, 3029–3036. [Google Scholar] [CrossRef]
- Lee, S.A.; Lee, N.; Oh, H.M.; Ahn, C.Y. Stepwise Treatment of Undiluted Raw Piggery Wastewater, Using Three Microalgal Species Adapted to High Ammonia. Chemosphere 2021, 263, 127934. [Google Scholar] [CrossRef]
- Palominos, N.; Castillo, A.; Guerrero, L.; Borja, R.; Huiliñir, C. Coupling of Anaerobic Digestion and Struvite Precipitation in the Same Reactor: Effect of Zeolite and Bischofite as Mg2+ Source. Front. Environ. Sci. 2021, 9, 581. [Google Scholar] [CrossRef]
- Montefiore, L.R.; Nelson, N.G.; Dean, A.; Sharara, M. Reconstructing the Historical Expansion of Industrial Swine Production from Landsat Imagery. Sci. Rep. 2022, 12, 1736. [Google Scholar] [CrossRef]
- Campagnolo, E.R.; Johnson, K.R.; Karpati, A.; Rubin, C.S.; Kolpin, D.W.; Meyer, M.T.; Esteban, J.E.; Currier, R.W.; Smith, K.; Thu, K.M.; et al. Antimicrobial Residues in Animal Waste and Water Resources Proximal to Large-Scale Swine and Poultry Feeding Operations. Sci. Total Environ. 2002, 299, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Manyi-Loh, C.E.; Mamphweli, S.N.; Meyer, E.L.; Okoh, A.I.; Makaka, G.; Simon, M. Microbial Anaerobic Digestion (Bio-Digesters) as an Approach to the Decontamination of Animal Wastes in Pollution Control and the Generation of Renewable Energy. Int. J. Environ. Res. Public Health 2013, 10, 4390–4417. [Google Scholar] [CrossRef] [PubMed]
- Ángeles, R.; Arnaiz, E.; Gutiérrez, J.; Sepúlveda-Muñoz, C.A.; Fernández-Ramos, O.; Muñoz, R.; Lebrero, R. Optimization of Photosynthetic Biogas Upgrading in Closed Photobioreactors Combined with Algal Biomass Production. J. Water Process Eng. 2020, 38, 101554. [Google Scholar] [CrossRef]
- Struk, M.; Sepúlveda-Muñoz, C.A.; Kushkevych, I.; Muñoz, R. Photoautotrophic Removal of Hydrogen Sulfide from Biogas Using Purple and Green Sulfur Bacteria. J. Hazard. Mater. 2023, 443, 130337. [Google Scholar] [CrossRef]
- Montes, N.; Otero, M.; Coimbra, R.N.; Méndez, R.; Martín-Villacorta, J. Removal of Tetracyclines from Swine Manure at Full-Scale Activated Sludge Treatment Plants. Environ. Technol. 2015, 36, 1966–1973. [Google Scholar] [CrossRef]
- Suzuki, K.; Waki, M.; Yasuda, T.; Fukumoto, Y.; Kuroda, K.; Sakai, T.; Suzuki, N.; Suzuki, R.; Matsuba, K. Distribution of Phosphorus, Copper and Zinc in Activated Sludge Treatment Process of Swine Wastewater. Bioresour. Technol. 2010, 101, 9399–9404. [Google Scholar] [CrossRef]
- García, D.; de Godos, I.; Domínguez, C.; Turiel, S.; Bolado, S.; Muñoz, R. A Systematic Comparison of the Potential of Microalgae-Bacteria and Purple Phototrophic Bacteria Consortia for the Treatment of Piggery Wastewater. Bioresour. Technol. 2019, 276, 18–27. [Google Scholar] [CrossRef]
- Marín, D.; Posadas, E.; García, D.; Puyol, D.; Lebrero, R.; Muñoz, R. Assessing the Potential of Purple Phototrophic Bacteria for the Simultaneous Treatment of Piggery Wastewater and Upgrading of Biogas. Bioresour. Technol. 2019, 281, 10–17. [Google Scholar] [CrossRef]
- Hülsen, T.; Hsieh, K.; Tait, S.; Barry, E.M.; Puyol, D.; Batstone, D.J. White and Infrared Light Continuous Photobioreactors for Resource Recovery from Poultry Processing Wastewater—A Comparison. Water Res. 2018, 144, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda-Muñoz, C.A.; Hontiyuelo, G.; Blanco, S.; Torres-Franco, A.F.; Muñoz, R. Photosynthetic Treatment of Piggery Wastewater in Sequential Purple Phototrophic Bacteria and Microalgae-Bacteria Photobioreactors. J. Water Process Eng. 2022, 47, 102825. [Google Scholar] [CrossRef]
- Capson-Tojo, G.; Batstone, D.J.; Grassino, M.; Vlaeminck, S.E.; Puyol, D.; Verstraete, W.; Kleerebezem, R.; Oehmen, A.; Ghimire, A.; Pikaar, I.; et al. Purple Phototrophic Bacteria for Resource Recovery: Challenges and Opportunities. Biotechnol. Adv. 2020, 43, 107567. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.N.; Daldal, F.; Thurnauer, M.C.; Beatty, J.T. The Purple Phototrophic Bacteria; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2009; Volume 28, ISBN 978-1-4020-8814-8. [Google Scholar]
- Chan, S.S.; Low, S.S.; Chew, K.W.; Ling, T.C.; Rinklebe, J.; Juan, J.C.; Ng, E.P.; Show, P.L. Prospects and Environmental Sustainability of Phyconanotechnology: A Review on Algae-Mediated Metal Nanoparticles Synthesis and Mechanism. Environ. Res. 2022, 212, 113140. [Google Scholar] [CrossRef]
- Medlin, L.K. Evolution of the Diatoms: Major Steps in Their Evolution and a Review of the Supporting Molecular and Morphological Evidence. Phycologia 2016, 55, 79–103. [Google Scholar] [CrossRef]
- Hallenbeck, P.C. Modern Topics in the Phototrophic Prokaryotes: Environmental and Applied Aspects; Springer International Publishing: Cham, Switzerland, 2017; ISBN 978-3-319-46259-2. [Google Scholar]
- Madigan, M.T.; Bender, K.S.; Buckley, D.H.; Sattley, W.M.; Stahl, D.A. Brock Biology of Microorganisms, 16th ed.; Pearson Education: Harlow, UK, 2021; ISBN 978-1-292-40479-0. [Google Scholar]
- Puyol, D.; Hülsen, T.; Padrino, B.; Batstone, D.J.; Martinez, F.; Melero, J.A. Exploring the Inhibition Boundaries of Mixed Cultures of Purple Phototrophic Bacteria for Wastewater Treatment in Anaerobic Conditions. Water Res. 2020, 183, 116057. [Google Scholar] [CrossRef]
- Larimer, F.W.; Chain, P.; Hauser, L.; Lamerdin, J.; Malfatti, S.; Do, L.; Land, M.L.; Pelletier, D.A.; Beatty, J.T.; Lang, A.S.; et al. Complete Genome Sequence of the Metabolically Versatile Photosynthetic Bacterium Rhodopseudomonas palustris. Nat. Biotechnol. 2004, 22, 55–61. [Google Scholar] [CrossRef]
- Touloupakis, E.; Poloniataki, E.G.; Casciana, M.; Ghanotakis, D.F.; Carlozzi, P. Poly-β-Hydroxybutyrate Production by Rhodopseudomonas sp. Grown in Semi-Continuous Mode in a 4 L Photobioreactor. Symmetry 2021, 13, 1609. [Google Scholar] [CrossRef]
- Hülsen, T.; Hsieh, K.; Batstone, D.J. Saline Wastewater Treatment with Purple Phototrophic Bacteria. Water Res. 2019, 160, 259–267. [Google Scholar] [CrossRef]
- Dalaei, P.; Ho, D.; Nakhla, G.; Santoro, D. Low Temperature Nutrient Removal from Municipal Wastewater by Purple Phototrophic Bacteria (PPB). Bioresour. Technol. 2019, 288, 121566. [Google Scholar] [CrossRef]
- Hülsen, T.; Barry, E.M.; Lu, Y.; Puyol, D.; Batstone, D.J. Low Temperature Treatment of Domestic Wastewater by Purple Phototrophic Bacteria: Performance, Activity, and Community. Water Res. 2016, 100, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Méndez, L.; Sepúlveda-Muñoz, C.A.; del Rosario Rodero, M.; de Godos, I.; Muñoz, R. Decarbonization Potentials Using Photobiological Systems. In Pathways to Water Sector Decarbonization, Carbon Capture and Utilization; IWA Publishing: London, UK, 2022; pp. 143–170. ISBN 9781789061789. [Google Scholar]
- Cogdell, R.J.; Isaacs, N.W.; Howard, T.D.; McLuskey, K.; Fraser, N.J.; Prince, S.M. How Photosynthetic Bacteria Harvest Solar Energy. J. Bacteriol. 1999, 181, 3869–3879. [Google Scholar] [CrossRef] [PubMed]
- Stomp, M.; Huisman, J.; Stal, L.J.; Matthijs, H.C.P. Colorful Niches of Phototrophic Microorganisms Shaped by Vibrations of the Water Molecule. ISME J. 2007, 1, 271–282. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, G.; Wan, T.; Lu, Y. Influences of Light and Oxygen Conditions on Photosynthetic Bacteria Macromolecule Degradation: Different Metabolic Pathways. Bioresour. Technol. 2011, 102, 9503–9508. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, G.; Dong, S. Quantitative Study of PNSB Energy Metabolism in Degrading Pollutants under Weak Light-Micro Oxygen Condition. Bioresour. Technol. 2011, 102, 4968–4973. [Google Scholar] [CrossRef] [PubMed]
- Izu, K.; Nakajima, F.; Yamamoto, K.; Kurisu, F. Aeration Conditions Affecting Growth of Purple Nonsulfur Bacteria in an Organic Wastewater Treatment Process. Syst. Appl. Microbiol. 2001, 24, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Navid, A.; Jiao, Y.; Wong, S.E.; Pett-Ridge, J. System-Level Analysis of Metabolic Trade-Offs during Anaerobic Photoheterotrophic Growth in Rhodopseudomonas palustris. BMC Bioinform. 2019, 20, 233. [Google Scholar] [CrossRef]
- Lu, H.; Zhang, G.; Zheng, Z.; Meng, F.; Du, T.; He, S. Bio-Conversion of Photosynthetic Bacteria from Non-Toxic Wastewater to Realize Wastewater Treatment and Bioresource Recovery: A Review. Bioresour. Technol. 2019, 278, 383–399. [Google Scholar] [CrossRef]
- Dalaei, P.; Bahreini, G.; Nakhla, G.; Santoro, D.; Batstone, D.; Hülsen, T. Municipal Wastewater Treatment by Purple Phototropic Bacteria at Low Infrared Irradiances Using a Photo-Anaerobic Membrane Bioreactor. Water Res. 2020, 173, 115535. [Google Scholar] [CrossRef]
- Yang, A.; Zhao, W.; Peng, M.; Zhang, G.; Zhi, R.; Meng, F. A Special Light-Aerobic Condition for Photosynthetic Bacteria-Membrane Bioreactor Technology. Bioresour. Technol. 2018, 268, 820–823. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, G.; Zhang, J.; Li, X.; Li, J. Performance, Carotenoids Yield and Microbial Population Dynamics in a Photobioreactor System Treating Acidic Wastewater: Effect of Hydraulic Retention Time (HRT) and Organic Loading Rate (OLR). Bioresour. Technol. 2016, 200, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Daigger, G.T.; Kang, J.; Zhang, G. Effects of Light Intensity and Photoperiod on Pigments Production and Corresponding Key Gene Expression of Rhodopseudomonas palustris in a Photobioreactor System. Bioresour. Technol. 2019, 294, 122172. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Liu, H.; He, H.; Luo, L.; Li, X.; Zeng, G.; Zhou, Z.; Lou, W.; Yang, C. Treatment of Anaerobically Digested Swine Wastewater by Rhodobacter blasticus and Rhodobacter capsulatus. Bioresour. Technol. 2016, 222, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda-Muñoz, C.A.; de Godos, I.; Puyol, D.; Muñoz, R. A Systematic Optimization of Piggery Wastewater Treatment with Purple Phototrophic Bacteria. Chemosphere 2020, 253, 126621. [Google Scholar] [CrossRef] [PubMed]
- Alloul, A.; Wuyts, S.; Lebeer, S.; Vlaeminck, S.E. Volatile Fatty Acids Impacting Phototrophic Growth Kinetics of Purple Bacteria: Paving the Way for Protein Production on Fermented Wastewater. Water Res. 2019, 152, 138–147. [Google Scholar] [CrossRef]
- Fradinho, J.C.; Oehmen, A.; Reis, M.A.M. Photosynthetic Mixed Culture Polyhydroxyalkanoate (PHA) Production from Individual and Mixed Volatile Fatty Acids (VFAs): Substrate Preferences and Co-Substrate Uptake. J. Biotechnol. 2014, 185, 19–27. [Google Scholar] [CrossRef]
- Oswald, W.J. Micro-Algae and Waste-Water Treatment. In Micro-Algal Biotechnology; Borowitzka, M.B.L., Ed.; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Chisti, Y. Biodiesel from Microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Christenson, L.; Sims, R. Production and Harvesting of Microalgae for Wastewater Treatment, Biofuels, and Bioproducts. Biotechnol. Adv. 2011, 29, 686–702. [Google Scholar] [CrossRef]
- Muñoz, R.; Guieysse, B. Algal-Bacterial Processes for the Treatment of Hazardous Contaminants: A Review. Water Res. 2006, 40, 2799–2815. [Google Scholar] [CrossRef]
- Posadas, E.; Marín, D.; Blanco, S.; Lebrero, R.; Muñoz, R. Simultaneous Biogas Upgrading and Centrate Treatment in an Outdoors Pilot Scale High Rate Algal Pond. Bioresour. Technol. 2017, 232, 133–141. [Google Scholar] [CrossRef]
- Nagarajan, D.; Lee, D.-J.; Varjani, S.; Lam, S.S.; Allakhverdiev, S.I.; Chang, J.-S. Microalgae-Based Wastewater Treatment—Microalgae-Bacteria Consortia, Multi-Omics Approaches and Algal Stress Response. Sci. Total Environ. 2022, 845, 157110. [Google Scholar] [CrossRef] [PubMed]
- Senge, M.; Ryan, A.; Letchford, K.; MacGowan, S.; Mielke, T. Chlorophylls, Symmetry, Chirality, and Photosynthesis. Symmetry 2014, 6, 781–843. [Google Scholar] [CrossRef]
- Faraloni, C.; Lorenzo, T.D.; Bonetti, A. Impact of Light Stress on the Synthesis of Both Antioxidants Polyphenols and Carotenoids, as Fast Photoprotective Response in Chlamydomonas reinhardtii: New Prospective for Biotechnological Potential of This Microalga. Symmetry 2021, 13, 2220. [Google Scholar] [CrossRef]
- Mountourakis, F.; Papazi, A.; Kotzabasis, K. The Microalga Chlorella vulgaris as a Natural Bioenergetic System for Effective CO2 Mitigation-New Perspectives against Global Warming. Symmetry. 2021, 13, 997. [Google Scholar] [CrossRef]
- Silveira, C.F.; de Assis, L.R.; de Sousa Oliveira, A.P.S.; Calijuri, M.L. Valorization of Swine Wastewater in a Circular Economy Approach: Effects of Hydraulic Retention Time on Microalgae Cultivation. Sci. Total Environ. 2021, 789, 147861. [Google Scholar] [CrossRef]
- Perez-Garcia, O.; Escalante, F.M.E.; De-Bashan, L.E.; Bashan, Y. Heterotrophic Cultures of Microalgae: Metabolism and Potential Products. Water Res. 2011, 45, 11–36. [Google Scholar] [CrossRef]
- Chai, W.S.; Chew, C.H.; Munawaroh, H.S.H.; Ashokkumar, V.; Cheng, C.K.; Park, Y.K.; Show, P.L. Microalgae and Ammonia: A Review on Inter-Relationship. Fuel 2021, 303, 121303. [Google Scholar] [CrossRef]
- García, D.; Alcántara, C.; Blanco, S.; Pérez, R.; Bolado, S.; Muñoz, R. Enhanced Carbon, Nitrogen and Phosphorus Removal from Domestic Wastewater in a Novel Anoxic-Aerobic Photobioreactor Coupled with Biogas Upgrading. Chem. Eng. J. 2017, 313, 424–434. [Google Scholar] [CrossRef]
- Dhaouefi, Z.; Toledo-Cervantes, A.; García, D.; Bedoui, A.; Ghedira, K.; Chekir-Ghedira, L.; Muñoz, R. Assessing Textile Wastewater Treatment in an Anoxic-Aerobic Photobioreactor and the Potential of the Treated Water for Irrigation. Algal Res. 2018, 29, 170–178. [Google Scholar] [CrossRef]
- Torres-Franco, A.F.; Zuluaga, M.; Hernández-Roldán, D.; Leroy-Freitas, D.; Sepúlveda-Muñoz, C.A.; Blanco, S.; Mota, C.R.; Muñoz, R. Assessment of the Performance of an Anoxic-Aerobic Microalgal-Bacterial System Treating Digestate. Chemosphere 2021, 270, 129437. [Google Scholar] [CrossRef]
- García, D.; Posadas, E.; Grajeda, C.; Blanco, S.; Martínez-Páramo, S.; Acién, G.; García-Encina, P.; Bolado, S.; Muñoz, R. Comparative Evaluation of Piggery Wastewater Treatment in Algal-Bacterial Photobioreactors under Indoor and Outdoor Conditions. Bioresour. Technol. 2017, 245, 483–490. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Nguyen, T.P.; Pham, T.H.; Duong, T.T.; Do, M.V.; Trinh, T.V.; Nguyen, Q.T.X.; Trinh, V.M. Removal of Nutrients and COD in Wastewater from Vietnamese Piggery Farm by the Culture of Chlorella vulgaris in a Pilot-Scaled Membrane Photobioreactor. Water 2022, 14, 3645. [Google Scholar] [CrossRef]
- Zittelli, G.C.; Silva Benavides, A.M.; Silovic, T.; Ranglová, K.; Masojídek, J.; Cicchi, B.; Faraloni, C.; Touloupakis, E.; Torzillo, G. Productivity and Nutrient Removal by the Microalga Chlamydopodium fusiforme Grown Outdoors in BG-11 and Piggery Wastewater. Front. Mar. Sci. 2022, 9, 2406. [Google Scholar] [CrossRef]
- Madigan, M.T.; Jung, D.O.; Woese, C.R.; Achenbach, L.A. Rhodoferax antarcticus sp. Nov., a Moderately Psychrophilic Purple Nonsulfur Bacterium Isolated from an Antarctic Microbial Mat. Arch. Microbiol. 2000, 173, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Capson-Tojo, G.; Lin, S.; Batstone, D.J.; Hülsen, T. Purple Phototrophic Bacteria Are Outcompeted by Aerobic Heterotrophs in the Presence of Oxygen. Water Res. 2021, 194, 116941. [Google Scholar] [CrossRef] [PubMed]
- Siefert, E.; Irgens, R.L.; Pfennig, N. Phototrophic Purple and Green Bacteria in a Sewage Treatment Plant. Appl. Environ. Microbiol. 1978, 35, 38–44. [Google Scholar] [CrossRef]
- Manhaeghe, D.; Michels, S.; Rousseau, D.P.L.; Van Hulle, S.W.H. A Semi-Mechanistic Model Describing the Influence of Light and Temperature on the Respiration and Photosynthetic Growth of Chlorella vulgaris. Bioresour. Technol. 2019, 274, 361–370. [Google Scholar] [CrossRef]
- Rossi, S.; Sforza, E.; Pastore, M.; Bellucci, M.; Casagli, F.; Marazzi, F.; Ficara, E. Photo-Respirometry to Shed Light on Microalgae-Bacteria Consortia—A Review. Rev. Environ. Sci. Biotechnol. 2020, 19, 43–72. [Google Scholar] [CrossRef]
- Mainardis, M.; Buttazzoni, M.; Cottes, M.; Moretti, A.; Goi, D. Respirometry Tests in Wastewater Treatment: Why and How? A Critical Review. Sci. Total Environ. 2021, 793, 148607. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Meireles, L.A.; Malcata, F.X. Microalgal Reactors: A Review of Enclosed System Designs and Performances. Biotechnol. Prog. 2006, 22, 1490–1506. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for Biodiesel Production and Other Applications: A Review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Marín, D.; Carmona-Martínez, A.A.; Blanco, S.; Lebrero, R.; Muñoz, R. Innovative Operational Strategies in Photosynthetic Biogas Upgrading in an Outdoors Pilot Scale Algal-Bacterial Photobioreactor. Chemosphere 2021, 264, 128470. [Google Scholar] [CrossRef] [PubMed]
- Marín, D.; Posadas, E.; Cano, P.; Pérez, V.; Blanco, S.; Lebrero, R.; Muñoz, R. Seasonal Variation of Biogas Upgrading Coupled with Digestate Treatment in an Outdoors Pilot Scale Algal-Bacterial Photobioreactor. Bioresour. Technol. 2018, 263, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Alloul, A.; Cerruti, M.; Adamczyk, D.; Weissbrodt, D.G.; Vlaeminck, S.E. Operational Strategies to Selectively Produce Purple Bacteria for Microbial Protein in Raceway Reactors. Environ. Sci. Technol. 2021, 55, 8278–8286. [Google Scholar] [CrossRef]
- Sakarika, M.; Spanoghe, J.; Sui, Y.; Wambacq, E.; Grunert, O.; Haesaert, G.; Spiller, M.; Vlaeminck, S.E. Purple Non-sulphur Bacteria and Plant Production: Benefits for Fertilization, Stress Resistance and the Environment. Microb. Biotechnol. 2020, 13, 1336–1365. [Google Scholar] [CrossRef]
- Davis, R.; Aden, A.; Pienkos, P.T. Techno-Economic Analysis of Autotrophic Microalgae for Fuel Production. Appl. Energy 2011, 88, 3524–3531. [Google Scholar] [CrossRef]
- Hülsen, T.; Barry, E.M.; Lu, Y.; Puyol, D.; Keller, J.; Batstone, D.J. Domestic Wastewater Treatment with Purple Phototrophic Bacteria Using a Novel Continuous Photo Anaerobic Membrane Bioreactor. Water Res. 2016, 100, 486–495. [Google Scholar] [CrossRef]
- Chitapornpan, S.; Chiemchaisri, C.; Chiemchaisri, W.; Honda, R.; Yamamoto, K. Organic Carbon Recovery and Photosynthetic Bacteria Population in an Anaerobic Membrane Photo-Bioreactor Treating Food Processing Wastewater. Bioresour. Technol. 2013, 141, 65–74. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from Microalgae-A Review of Technologies for Production, Processing, and Extractions of Biofuels and Co-Products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Grassino, M.; Batstone, D.J.; Yong, K.W.L.; Capson-Tojo, G.; Hülsen, T. Method Development for PPB Culture Screening, Pigment Analysis with UPLC-UV-HRMS vs. Spectrophotometric Methods, and Spectral Decomposition-Based Analysis. Talanta 2022, 246, 123490. [Google Scholar] [CrossRef]
- Borowitzka, M.A. High-Value Products from Microalgae-Their Development and Commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Lorenz, R.T.; Cysewski, G.R. Commercial Potential for Haematococcus Microalgae as a Natural Source of Astaxanthin. Trends Biotechnol. 2000, 18, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhu, M.; Wang, K.; He, S.; Shi, X.; Yuan, B.; Dong, B.; Wang, Z. Preparation of Core-Shell Microcapsules Based on Microfluidic Technology for the Encapsulation, Protection and Controlled Delivery of Phycocyanin. J. Drug Deliv. Sci. Technol. 2022, 72, 103361. [Google Scholar] [CrossRef]
- Cao, K.; Zhi, R.; Zhang, G. Photosynthetic Bacteria Wastewater Treatment with the Production of Value-Added Products: A Review. Bioresour. Technol. 2020, 299, 122648. [Google Scholar] [CrossRef] [PubMed]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial Applications of Microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Soltanzadeh, M.; Ebrahimi, A.R.; Hamishehkar, H. Spirulina platensis Protein Hydrolysates: Techno-Functional, Nutritional and Antioxidant Properties. Algal Res. 2022, 65, 102739. [Google Scholar] [CrossRef]
- Lupatini, A.L.; Colla, L.M.; Canan, C.; Colla, E. Potential Application of Microalga Spirulina platensis as a Protein Source. J. Sci. Food Agric. 2017, 97, 724–732. [Google Scholar] [CrossRef]
- Hülsen, T.; Hsieh, K.; Lu, Y.; Tait, S.; Batstone, D.J. Simultaneous Treatment and Single Cell Protein Production from Agri-Industrial Wastewaters Using Purple Phototrophic Bacteria or Microalgae—A Comparison. Bioresour. Technol. 2018, 254, 214–223. [Google Scholar] [CrossRef]
- Zhu, Y.; Lu, W.; Ye, L.; Chen, Z.; Hu, W.; Wang, C.; Chen, J.; Yu, H. Enhanced Synthesis of Coenzyme Q10 by Reducing the Competitive Production of Carotenoids in Rhodobacter sphaeroides. Biochem. Eng. J. 2017, 125, 50–55. [Google Scholar] [CrossRef]
- Kumar, A.; Kaur, H.; Devi, P.; Mohan, V. Role of Coenzyme Q10 (CoQ10) in Cardiac Disease, Hypertension and Meniere-like Syndrome. Pharmacol. Ther. 2009, 124, 259–268. [Google Scholar] [CrossRef]
- He, S.; Lu, H.; Zhang, G.; Ren, Z. Production of Coenzyme Q10 by Purple Non-Sulfur Bacteria: Current Development and Future Prospect. J. Clean. Prod. 2021, 307, 127326. [Google Scholar] [CrossRef]
- Fradinho, J.C.; Oehmen, A.; Reis, M.A.M. Improving Polyhydroxyalkanoates Production in Phototrophic Mixed Cultures by Optimizing Accumulator Reactor Operating Conditions. Int. J. Biol. Macromol. 2019, 126, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Q.; Patel, M.K. Plastics Derived from Biological Sources: Present and Future: A Technical and Environmental Review. Chem. Rev. 2012, 112, 2082–2099. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.S.; Miranda, A.L.; de Morais, M.G.; Costa, J.A.V.; Druzian, J.I. Microalgae as Source of Polyhydroxyalkanoates (PHAs)—A Review. Int. J. Biol. Macromol. 2019, 131, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Adessi, A.; McKinlay, J.B.; Harwood, C.S.; De Philippis, R. A Rhodopseudomonas palustris NifA* Mutant Produces H2 from NH4+ -Containing Vegetable Wastes. Int. J. Hydrogen Energy 2012, 37, 15893–15900. [Google Scholar] [CrossRef]
- Fan, L.; Tu, Z.; Chan, S.H. Recent Development of Hydrogen and Fuel Cell Technologies: A Review. Energy Rep. 2021, 7, 8421–8446. [Google Scholar] [CrossRef]
- Adessi, A.; De Philippis, R. Photobioreactor Design and Illumination Systems for H2 Production with Anoxygenic Photosynthetic Bacteria: A Review. Int. J. Hydrog. Energy 2014, 39, 3127–3141. [Google Scholar] [CrossRef]
- Iqbal, K.; Saxena, A.; Pande, P.; Tiwari, A.; Chandra Joshi, N.; Varma, A.; Mishra, A. Microalgae-Bacterial Granular Consortium: Striding towards Sustainable Production of Biohydrogen Coupled with Wastewater Treatment. Bioresour. Technol. 2022, 354, 127203. [Google Scholar] [CrossRef]
- Arutselvan, C.; Seenivasan, H.K.; Lewis Oscar, F.; Ramya, G.; Thuy Lan Chi, N.; Pugazhendhi, A.; Thajuddin, N. Review on Wastewater Treatment by Microalgae in Different Cultivation Systems and Its Importance in Biodiesel Production. Fuel 2022, 324, 124623. [Google Scholar] [CrossRef]
- Szulczyk, K.R.; Tan, Y.M. Economic Feasibility and Sustainability of Commercial Bioethanol from Microalgal Biomass: The Case of Malaysia. Energy 2022, 253, 124151. [Google Scholar] [CrossRef]
- Ho, S.H.; Huang, S.W.; Chen, C.Y.; Hasunuma, T.; Kondo, A.; Chang, J.S. Bioethanol Production Using Carbohydrate-Rich Microalgae Biomass as Feedstock. Bioresour. Technol. 2013, 135, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Acebu, P.I.G.; de Luna, M.D.G.; Chen, C.Y.; Abarca, R.R.M.; Chen, J.H.; Chang, J.S. Bioethanol Production from Chlorella vulgaris ESP-31 Grown in Unsterilized Swine Wastewater. Bioresour. Technol. 2022, 352, 127086. [Google Scholar] [CrossRef] [PubMed]
- Appels, L.; Baeyens, J.; Degrève, J.; Dewil, R. Principles and Potential of the Anaerobic Digestion of Waste-Activated Sludge. Prog. Energy Combust. Sci. 2008, 34, 755–781. [Google Scholar] [CrossRef]
- Muñoz, R.; Meier, L.; Diaz, I.; Jeison, D. A Review on the State-of-the-Art of Physical/Chemical and Biological Technologies for Biogas Upgrading. Rev. Environ. Sci. Biotechnol. 2015, 14, 727–759. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, Y.; Zhao, G.; Zhang, H. Nutrient Removal and Biogas Upgrading by Integrating Freshwater Algae Cultivation with Piggery Anaerobic Digestate Liquid Treatment. Appl. Microbiol. Biotechnol. 2015, 99, 6493–6501. [Google Scholar] [CrossRef] [PubMed]
- Zabed, H.M.; Akter, S.; Yun, J.; Zhang, G.; Zhang, Y.; Qi, X. Biogas from Microalgae: Technologies, Challenges and Opportunities. Renew. Sustain. Energy Rev. 2020, 117, 109503. [Google Scholar] [CrossRef]
- Batool, K.; tuz Zahra, F.; Rehman, Y. Arsenic-Redox Transformation and Plant Growth Promotion by Purple Nonsulfur Bacteria Rhodopseudomonas palustris CS2 and Rhodopseudomonas faecalis SS5. Biomed. Res. Int. 2017, 2017, 6250327. [Google Scholar] [CrossRef]
- Coppens, J.; Grunert, O.; Van Den Hende, S.; Vanhoutte, I.; Boon, N.; Haesaert, G.; De Gelder, L. The Use of Microalgae as a High-Value Organic Slow-Release Fertilizer Results in Tomatoes with Increased Carotenoid and Sugar Levels. J. Appl. Phycol. 2016, 28, 2367–2377. [Google Scholar] [CrossRef]








| Composition | Domestic | Industrial 1 | Agro-Industrial 2 |
|---|---|---|---|
| COD (mg L−1) | 526 | 10,000 | 54,000 |
| TN (mg L−1) | 46 | 1 | 5000 |
| TP (mg L−1) | 6 | 3 | 1500 |
| pH | 6.8 | 7.0 | 7.7 |
| PWW Characteristics (mg L−1) | Reactor Type (Volume) | Dominant Strain | Pollutant Removal (%) | References | |
|---|---|---|---|---|---|
| Carbon | Nitrogen | ||||
| TOC: 574 | Open PBR | Rhodoplanes sp. | 87 | 83 | [30] |
| TN: 166 | (3 L) | ||||
| TOC: 10,318–1989 | Batch PBR | PPB mixed | 78 | 13 | [31] |
| TN: 2209–563 | (0.4 L) | ||||
| TOC: 15,775–1131 | Batch PBR | Rhodopseudomonas sp. | 75 | 39 | [59] |
| TN: 5028–366 | (0.5 L) | ||||
| TOC: 1180 | Open PBR | PPB mixed | 99 | 99 | [14] |
| TN: 380 | (3 and 1.5 L) | ||||
| COD: 4130 | Flat plate PBR | Rhodopseudomonas sp. | 71 | 22 | [3] |
| TKN: 1160 | (60, 80 and 100 L) | ||||
| TOC: 860 | Batch PBR | R. palustris | 79 | 42 | [9] |
| TN: 380 | (0.5 L) | ||||
| PWW Characteristics (mg L−1) | Reactor Type (Volume) | Dominant Strain | Pollutant Removal (%) | References | |
|---|---|---|---|---|---|
| Carbon | Nitrogen | ||||
| COD: 526–4346 | High-rate algal ponds | Microalgae mixed | 76 | 88 | [8] |
| TKN: 59–370 | (464 L) | ||||
| TOC: 963 | Open PBR | C. vulgaris | 94 | 56 | [77] |
| TN: 341 | (3 L) | ||||
| TOC: 574 | Open PBR | Microalgae mixed | 84 | 87 | [30] |
| TN: 166 | (3 L) | ||||
| COD: 8420–6870 | Sequential Batch PBR | Microalgae mixed | 92 | 90 | [21] |
| NH3-N: 850–1150 | (4 L) | ||||
| COD: 287 | Membrane PBR | C. vulgaris | 66 | 74 | [78] |
| NH4+: 184 | (50 L) | ||||
| COD: 1016 | Cylindrica PBR | Chlamydopodium fusiforme | 41 | 93 | [79] |
| N-NH4: 92 | (10 L) | ||||
| Open Photobioreactors | Closed Photobioreactors | Reference | |
|---|---|---|---|
| Capital investment | Low | High | [91] |
| Scalability | High | Variable | [92] |
| Culture control | Low | High | [64] |
| Culture contamination | High | Low | [64] |
| Evaporation rate | High | Low | [64] |
| Biomass productivity | Variable | High | [92] |
| Nutrient removal | High | High | [65] |
| Nutrient recovery in biomass | Variable | High | [92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepúlveda-Muñoz, C.A.; de Godos, I.; Muñoz, R. Wastewater Treatment Using Photosynthetic Microorganisms. Symmetry 2023, 15, 525. https://doi.org/10.3390/sym15020525
Sepúlveda-Muñoz CA, de Godos I, Muñoz R. Wastewater Treatment Using Photosynthetic Microorganisms. Symmetry. 2023; 15(2):525. https://doi.org/10.3390/sym15020525
Chicago/Turabian StyleSepúlveda-Muñoz, Cristian A., Ignacio de Godos, and Raúl Muñoz. 2023. "Wastewater Treatment Using Photosynthetic Microorganisms" Symmetry 15, no. 2: 525. https://doi.org/10.3390/sym15020525
APA StyleSepúlveda-Muñoz, C. A., de Godos, I., & Muñoz, R. (2023). Wastewater Treatment Using Photosynthetic Microorganisms. Symmetry, 15(2), 525. https://doi.org/10.3390/sym15020525