
Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis



Abstract
1. Introduction
2. Dynamics of Total Nitrogen Transformation
2.1. Equilibrium Points and Local Stability Analysis
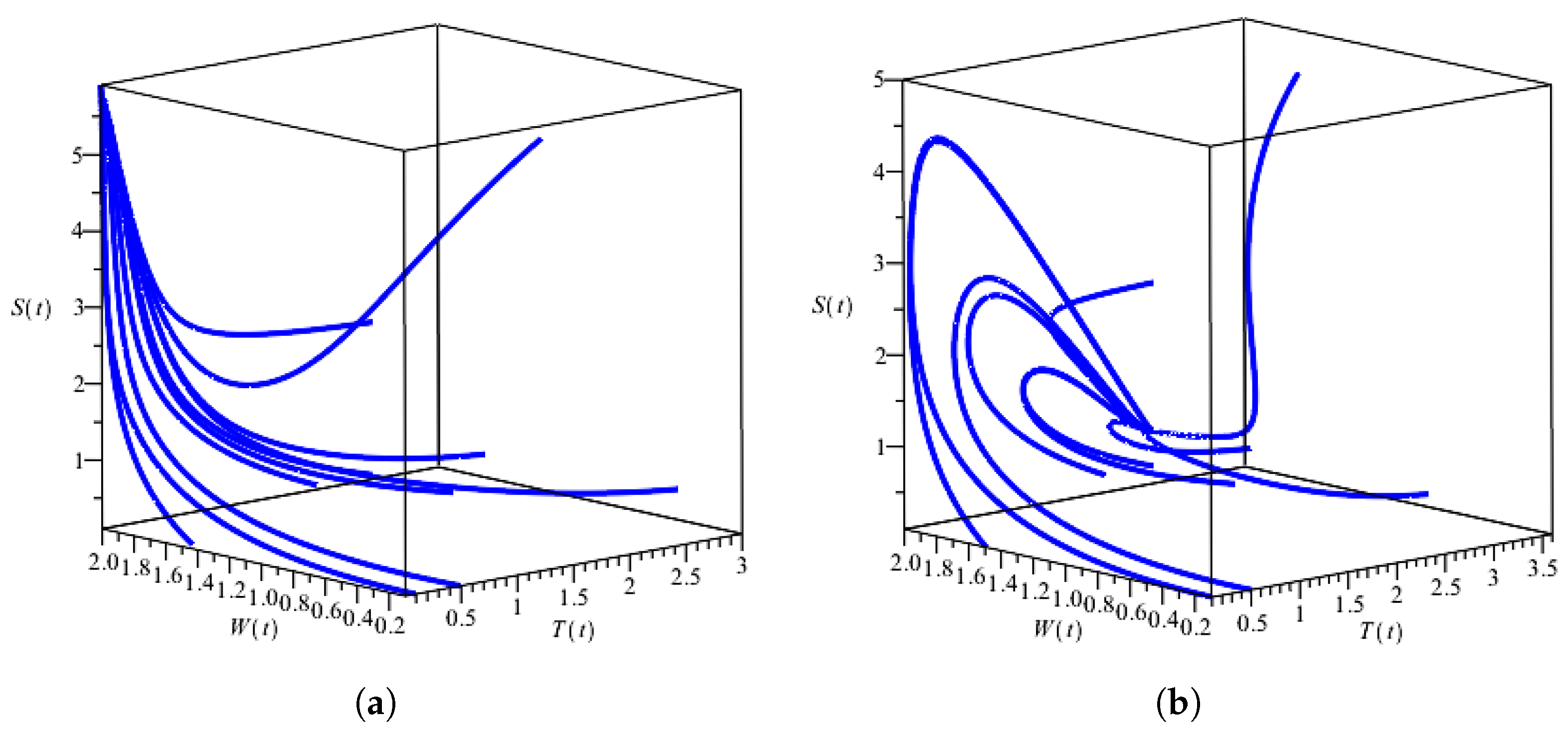
2.2. Numerical Solution
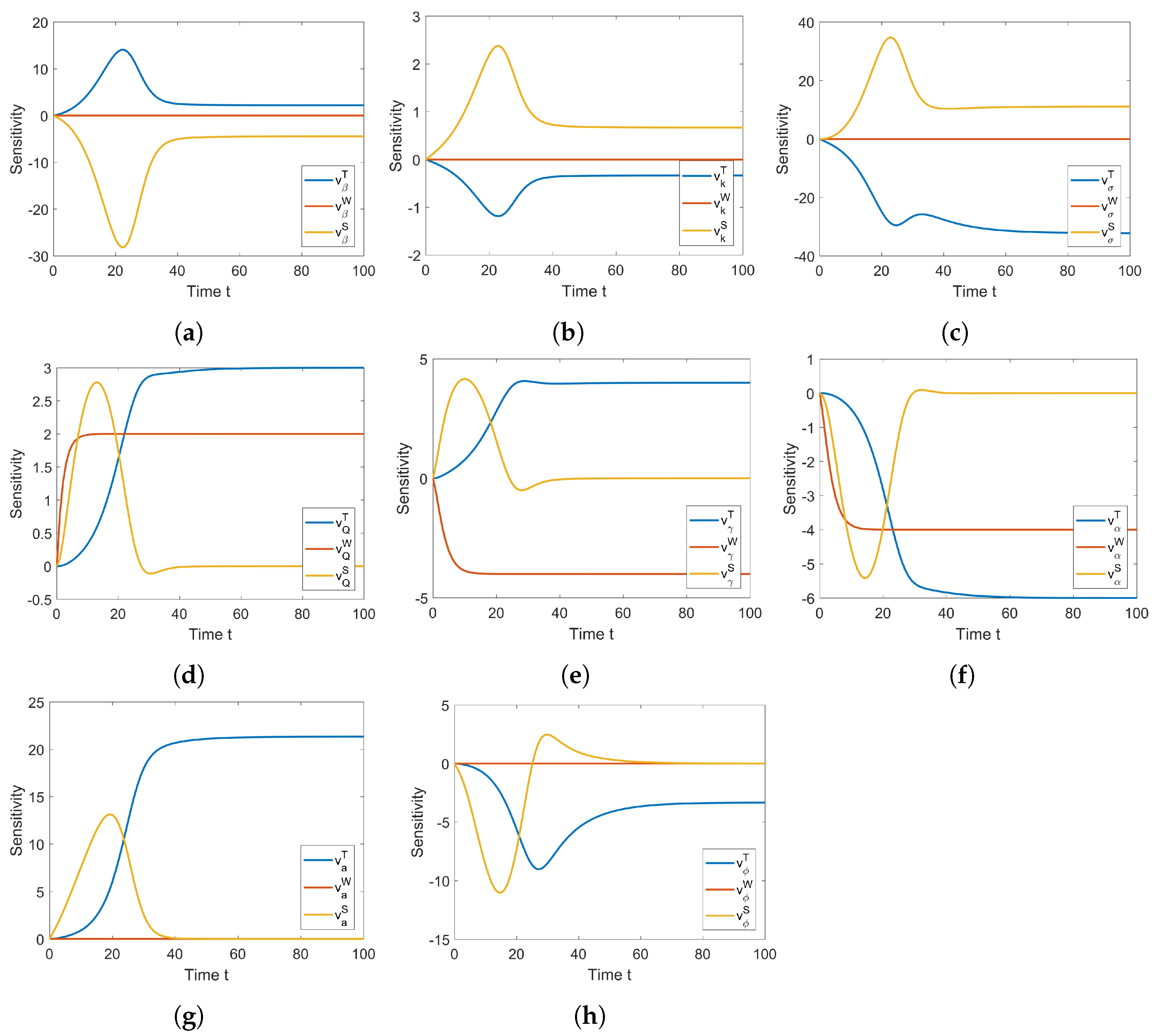
2.3. Sensitivity Analysis
3. Discrete Form Model
3.1. The Local Stability of Non-Zero Equilibrium
3.2. Numerical Simulations
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
Appendix B
References
- Choi, H.; Geronimo, F.K.; Jeon, M.; Kim, L.H. Evaluation of bacterial community in constructed wetlands treating different sources of wastewater. Ecol. Eng. 2022, 182, 106703. [Google Scholar] [CrossRef]
- Bunwong, K.; Sae-jie, W.; Lenbury, Y. Modelling nitrogen dynamics of a constructed wetland: Nutrient removal process with variable yield. Nonlinear Anal. Theory Methods Appl. 2009, 71, e1538–e1546. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, J.; Ngo, H.H.; Guo, W.; Hu, Z.; Liang, S.; Fan, J.; Liu, H. A review on the sustainability of constructed wetlands for wastewater treatment: Design and operation. Bioresour. Technol. 2015, 175, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Senzia, M.A.; Mashauri, D.A.; Mayo, A.W. Suitability of constructed wetlands and waste stabilisation ponds in wastewater treatment: Nitrogen transformation and removal. Phys. Chem. Earth Parts A/B/C 2003, 28, 1117–1124. [Google Scholar] [CrossRef]
- Han, Z.; Dong, J.; Shen, Z.; Mou, R.; Zhou, Y.; Chen, X.; Fu, X.; Yang, C. Nitrogen removal of anaerobically digested swine wastewater by pilot-scale tidal flow constructed wetland based on in-situ biological regeneration of zeolite. Chemosphere 2019, 217, 364–373. [Google Scholar] [CrossRef]
- Strigul, N.S.; Kravchenko, L.V. Mathematical modeling of PGPR inoculation into the rhizosphere. Environ. Model. Softw. 2006, 21, 1158–1171. [Google Scholar] [CrossRef]
- Jia, L.; Gou, E.; Liu, H.; Lu, S.; Wu, S.; Wu, H. Exploring utilization of recycled agricultural biomass in constructed wetlands: Characterization of the driving force for high-rate nitrogen removal. Environ. Sci. Technol. 2019, 53, 1258–1268. [Google Scholar] [CrossRef]
- Choi, H.; Geronimo, F.K.F.; Jeon, M.; Kim, L.H. Investigation of the Factors Affecting the Treatment Performance of a Stormwater Horizontal Subsurface Flow Constructed Wetland Treating Road and Parking lot Runoff. Water 2021, 13, 1242. [Google Scholar] [CrossRef]
- Rousseau, D.P.L.; Louage, F.; Wang, Q.; Zhang, R. Constructed Wetlands for Urban Wastewater Treatment: An Overview. In Encyclopedia of Inland Waters, 2nd ed.; Mehner, T., Tockner, K., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 272–284. [Google Scholar]
- Chen, Y.; Zhang, J.; Guo, Z.; Li, M.; Wu, H. Optimizing agricultural biomass application to enhance nitrogen removal in vertical flow constructed wetlands for treating low-carbon wastewater. Environ. Res. 2022, 209, 112867. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, M.; Dong, L.; Han, C.; Li, M.; Wu, H. Effects of biochar dosage on treatment performance, enzyme activity and microbial community in aerated constructed wetlands for treating low C/N domestic sewage. Environ. Technol. Innovat. 2021, 24, 101919. [Google Scholar] [CrossRef]
- Jia, L.; Li, C.; Zhang, Y.; Chen, Y.; Li, M.; Wu, S.; Wu, H. Microbial community responses to agricultural biomass addition in aerated constructed wetlands treating low carbon wastewater. J. Environ. Manag. 2020, 270, 110912. [Google Scholar] [CrossRef]
- Wu, Y.; Chung, A.; Tam, N.F.Y.; Pi, N.; Wong, M.H. Constructed mangrove wetland as secondary treatment system for municipal wastewater. Ecol. Eng. 2008, 34, 137–146. [Google Scholar] [CrossRef]
- Leung, J.Y.S.; Cai, Q.; Tam, N.F.Y. Comparing subsurface flow constructed wetlands with mangrove plants and freshwater wetland plants for removing nutrients and toxic pollutants. Ecol. Eng. 2016, 95, 129–137. [Google Scholar] [CrossRef]
- Lobry, J.R.; Flandrois, J.P.; Carret, G.; Pave, A. Monod’s bacterial growth model revisited. Bull. Math. Biol. 1992, 54, 117–122. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Lee, S.C.; Juang, H.T.; Hur, M.T.; Hwang, Y.H. Nitrogen nutritional status and fate of applied N in Mangrove soils. Bot. Bull. Acad. Sin. 1996, 37, 191–196. [Google Scholar]
- Pisman, T.I.; Pechurkin, N.S.; Mariasova, T.S.; Somova, L.A.; Sarangova, A.B. A mathematical model of “plants-microorganisms” interaction on complete mineral medium and under nitrogen limitation. Adv. Space Res. 1999, 24, 383–387. [Google Scholar] [CrossRef]
- Walker, R.L.; Burns, I.G.; Moorby, J. Responses of plant growth rate to nitrogen supply: A comparison of relative addition and N interruption treatments. J. Exp. Bot. 2001, 52, 309–317. [Google Scholar] [CrossRef]
- Pilyugin, S.S.; Waltman, P. Multiple limit cycles in the chemostat with variable yield. Math. Biosci. 2003, 182, 151–166. [Google Scholar] [CrossRef]
- Zhu, L.; Huang, X. Multiple limit cycles in a continuous culture vessel with variable yield. Nonlinear Anal. 2006, 64, 887–894. [Google Scholar] [CrossRef]
- Zhao, Z.; Pang, L.; Zhao, Z.; Luo, C. Impulsive State Feedback Control of the Rhizosphere Microbial Degradation in the Wetland Plant. Discret. Dyn. Nat. Soc. 2015, 2015, 612354. [Google Scholar] [CrossRef]
- Zhao, Z.; Song, Y.; Pang, L. Mathematical modeling of rhizosphere microbial degradation with impulsive diffusion on nutrient. Adv. Differ. Equ. 2016, 2016, 24. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, Q.; Chen, L. Effect of rhizosphere dispersal and impulsive input on the growth of wetland plant. Math. Comput. Simul. 2018, 152, 69–80. [Google Scholar] [CrossRef]
- Zhao, Z.; Yin, Q.; Li, Q.; Wu, X. Mathematical model for diffusion of the rhizosphere microbial degradation with impulsive feedback control. J. Biol. Dyn. 2020, 14, 566–577. [Google Scholar] [CrossRef]
- Suandi, D.; Ningrum, I.P.; Alifah, A.N.; Izzah, N.; Reza, M.P.; Muwahidah, I.K. Mathematical Modeling and Sensitivity Analysis of the Existence of Male Calico Cats Population Based on Cross Breeding of All Coat Colour Types. Commun. Biomath. Sci. 2019, 2, 96–104. [Google Scholar] [CrossRef][Green Version]
- Mapfumo, K.Z.; Pagan’a, J.C.; Juma, V.O.; Kavallaris, N.I.; Madzvamuse, A. A Model for the Proliferation–Quiescence Transition in Human Cells. Mathematics 2022, 10, 2426. [Google Scholar] [CrossRef]
- Rentzeperis, F.; Wallace, D. Local and global sensitivity analysis of spheroid and xenograft models of the acid-mediated development of tumor malignancy. Appl. Math. Model. 2022, 109, 629–650. [Google Scholar] [CrossRef]
- Alsahafi, S.; Woodcock, S. Exploring HIV Dynamics and an Optimal Control Strategy. Mathematics 2022, 10, 749. [Google Scholar] [CrossRef]
- Reinharz, V.; Churkin, A.; Dahari, H.; Barash, D. Advances in Parameter Estimation and Learning from Data for Mathematical Models of Hepatitis C Viral Kinetics. Mathematics 2022, 10, 2136. [Google Scholar] [CrossRef]
- Ansori, M.F.; Sumarti, N.; Sidarto, K.A.; Guandi, I. Analyzing a macroprudential instrument during the covid-19 pandemic using border collision bifurcation. Rev. Electron. Comun. Y Trab. ASEPUMA Rect. 2021, 22, 113–125. [Google Scholar] [CrossRef]
- Du, X.; Han, X.; Lei, C. Behavior Analysis of a Class of Discrete-Time Dynamical System with Capture Rate. Mathematics 2022, 10, 2410. [Google Scholar] [CrossRef]
- Saeed, T.; Djeddi, K.; Guirao, J.L.G.; Alsulami, H.H.; Alhodaly, M.S. A Discrete Dynamics Approach to a Tumor System. Mathematics 2022, 10, 1774. [Google Scholar] [CrossRef]
- Liu, B.; Wu, R. Bifurcation and Patterns Analysis for a Spatiotemporal Discrete Gierer-Meinhardt System. Mathematics 2022, 10, 243. [Google Scholar] [CrossRef]
- He, Z.Y.; Abbes, A.; Jahanshahi, H.; Alotaibi, N.D.; Wang, Y. Fractional-Order Discrete-Time SIR Epidemic Model with Vaccination: Chaos and Complexity. Mathematics 2022, 10, 165. [Google Scholar] [CrossRef]
- Gandolfo, G. Economic Dynamics: Methods and Models, 2nd ed.; Elsevier Science Publisher BV: Amsterdam, The Netherlands, 1985. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Notation | Description | Value | Unit |
|---|---|---|---|
| Maximum possible value of the plant growth rate at infinite total nitrogen concentration in the soil solution | 0.25 | ||
| k | Semi-saturation | 1 | |
| Level of garbage | 0.1 | ||
| Q | Input level of total nitrogen concentration | 1 | |
| Exchange rate of total nitrogen between wastewater and soil solution | 0.3 | ||
| Rate of total nitrogen loss in wastewater through runoff or evaporation | 0.2 | ||
| a | Conversion of nutrients consumed from biomass produced | 0.5 | Dimensionless |
| Rate of total nitrogen loss in the soil solution by leaching or denitrification | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sunarsih; Ansori, M.F.; Khabibah, S.; Sasongko, D.P. Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis. Symmetry 2022, 14, 1924. https://doi.org/10.3390/sym14091924
Sunarsih, Ansori MF, Khabibah S, Sasongko DP. Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis. Symmetry. 2022; 14(9):1924. https://doi.org/10.3390/sym14091924
Chicago/Turabian StyleSunarsih, Moch. Fandi Ansori, Siti Khabibah, and Dwi Purwantoro Sasongko. 2022. "Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis" Symmetry 14, no. 9: 1924. https://doi.org/10.3390/sym14091924
APA StyleSunarsih, Ansori, M. F., Khabibah, S., & Sasongko, D. P. (2022). Continuous and Discrete Dynamical Models of Total Nitrogen Transformation in a Constructed Wetland: Sensitivity and Bifurcation Analysis. Symmetry, 14(9), 1924. https://doi.org/10.3390/sym14091924