
A General Model for Describing the Ovate Leaf Shape

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
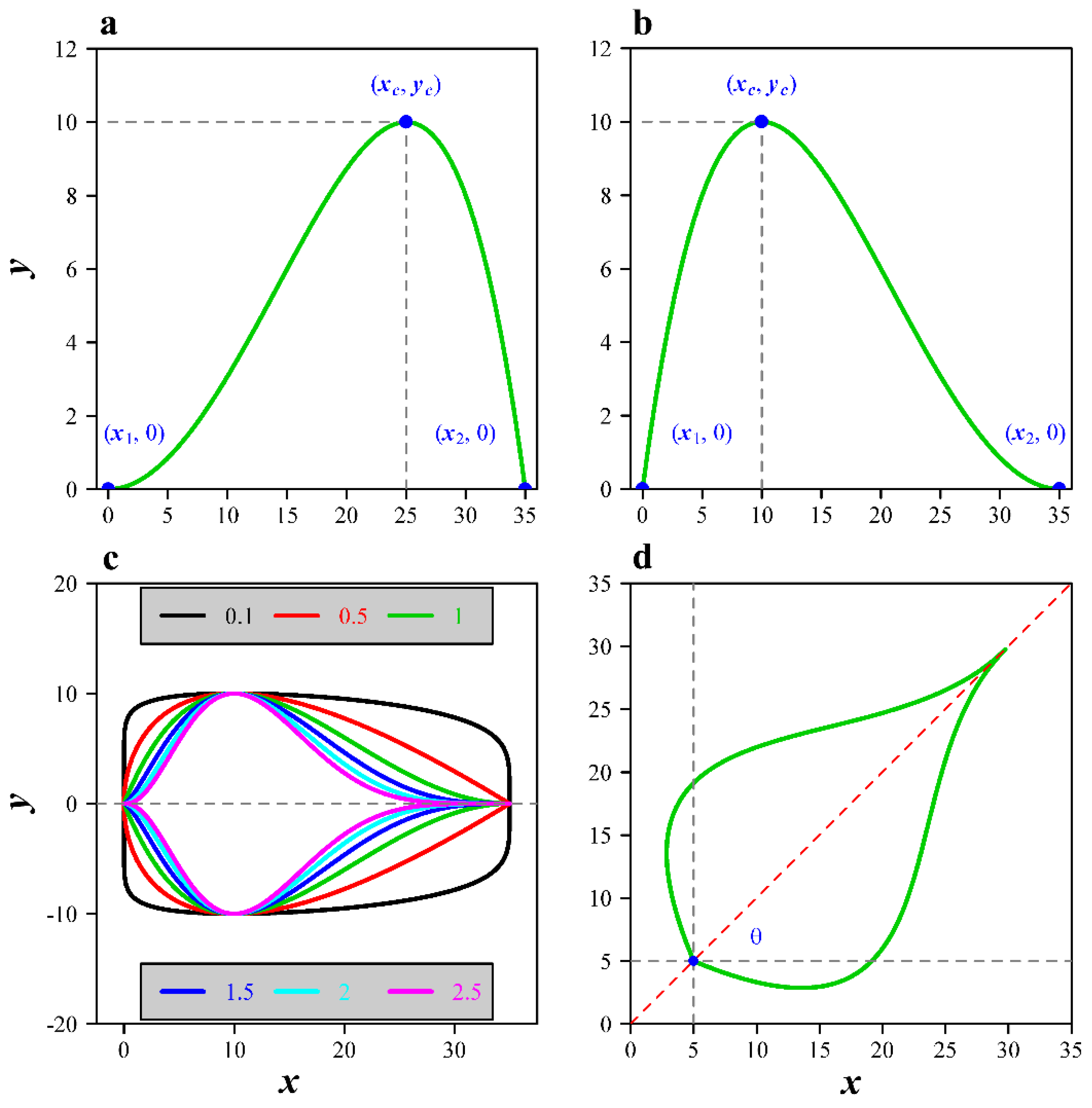
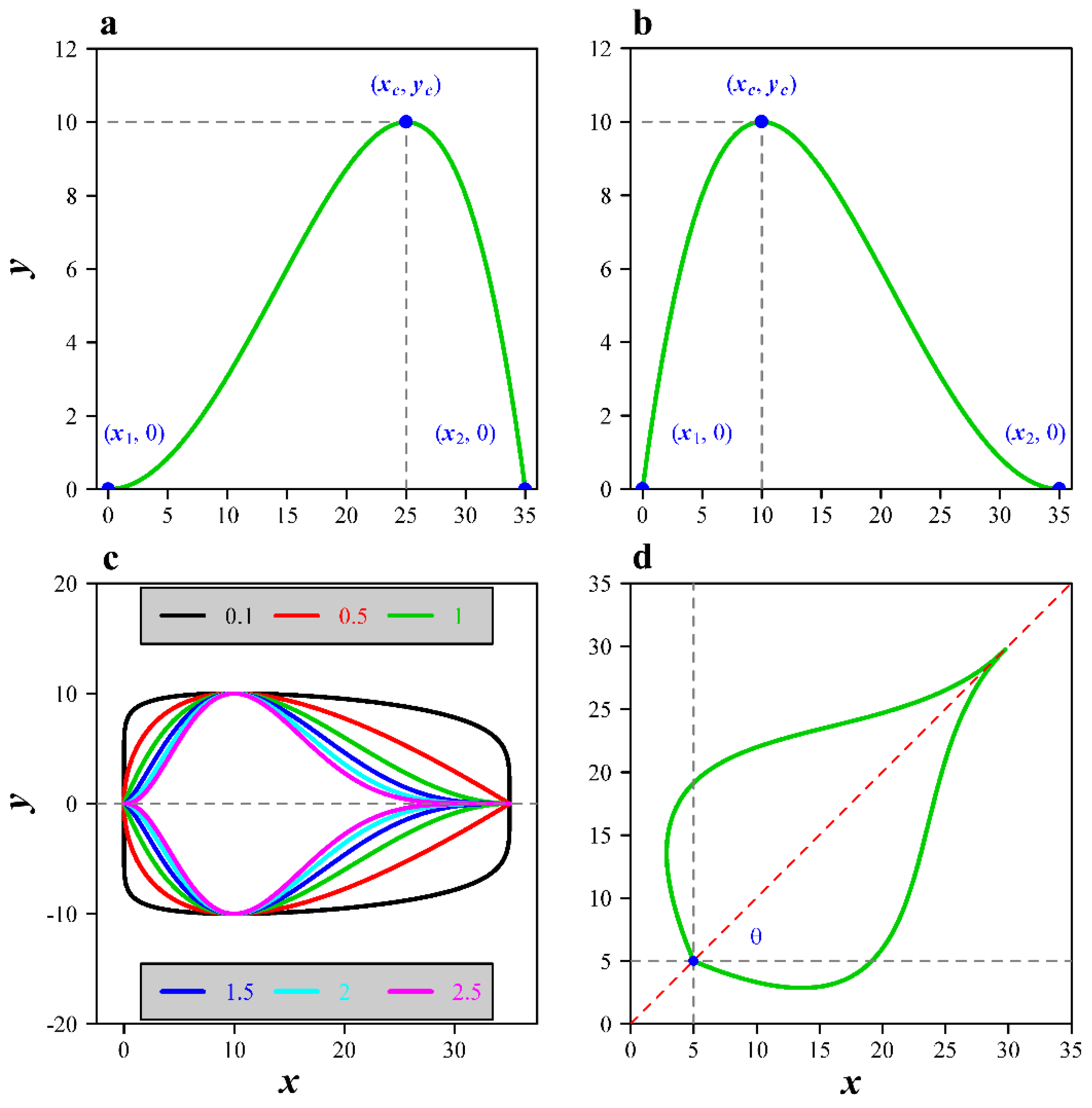
2.1. Models
2.2. Parameter Estimation for the Models
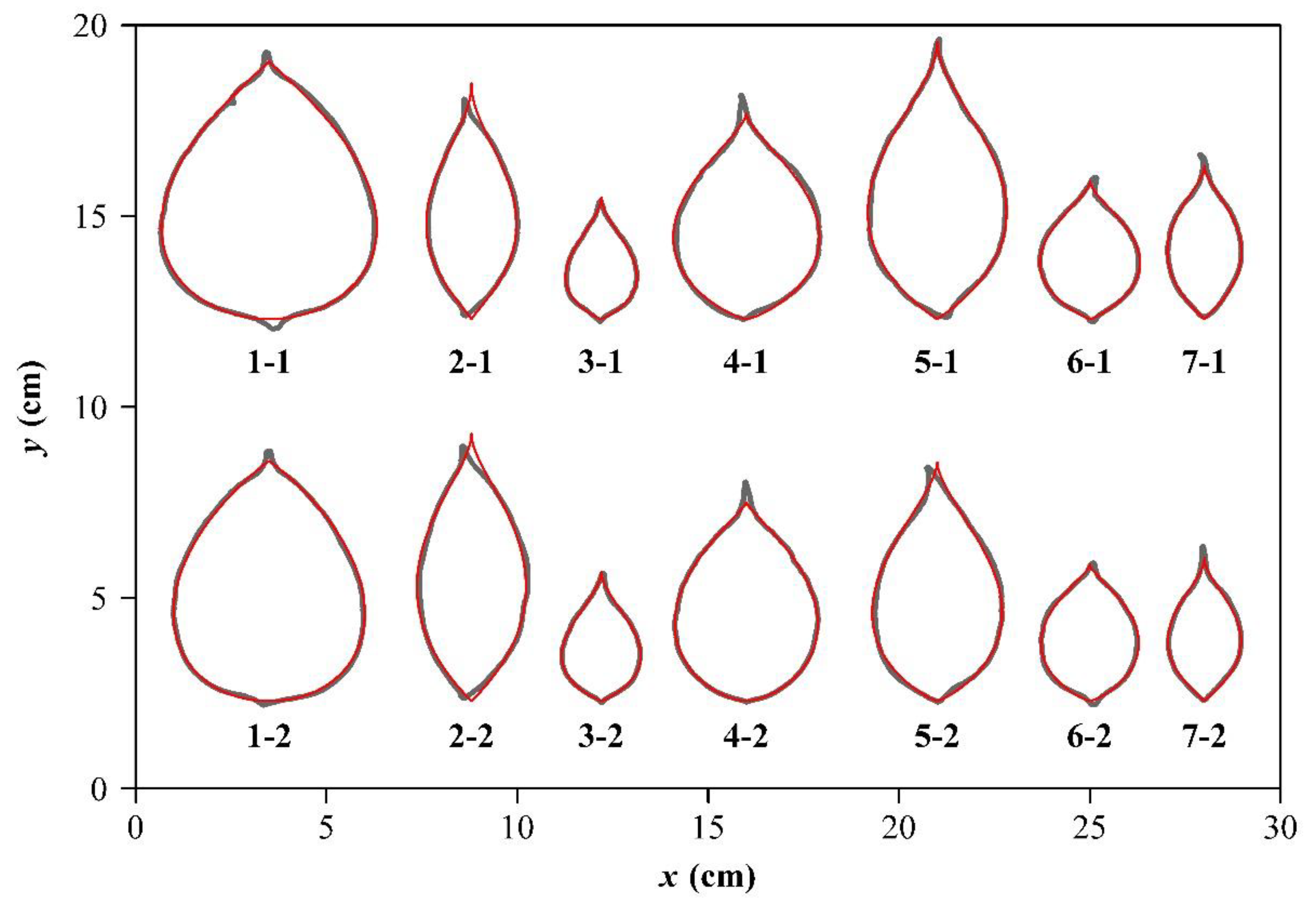
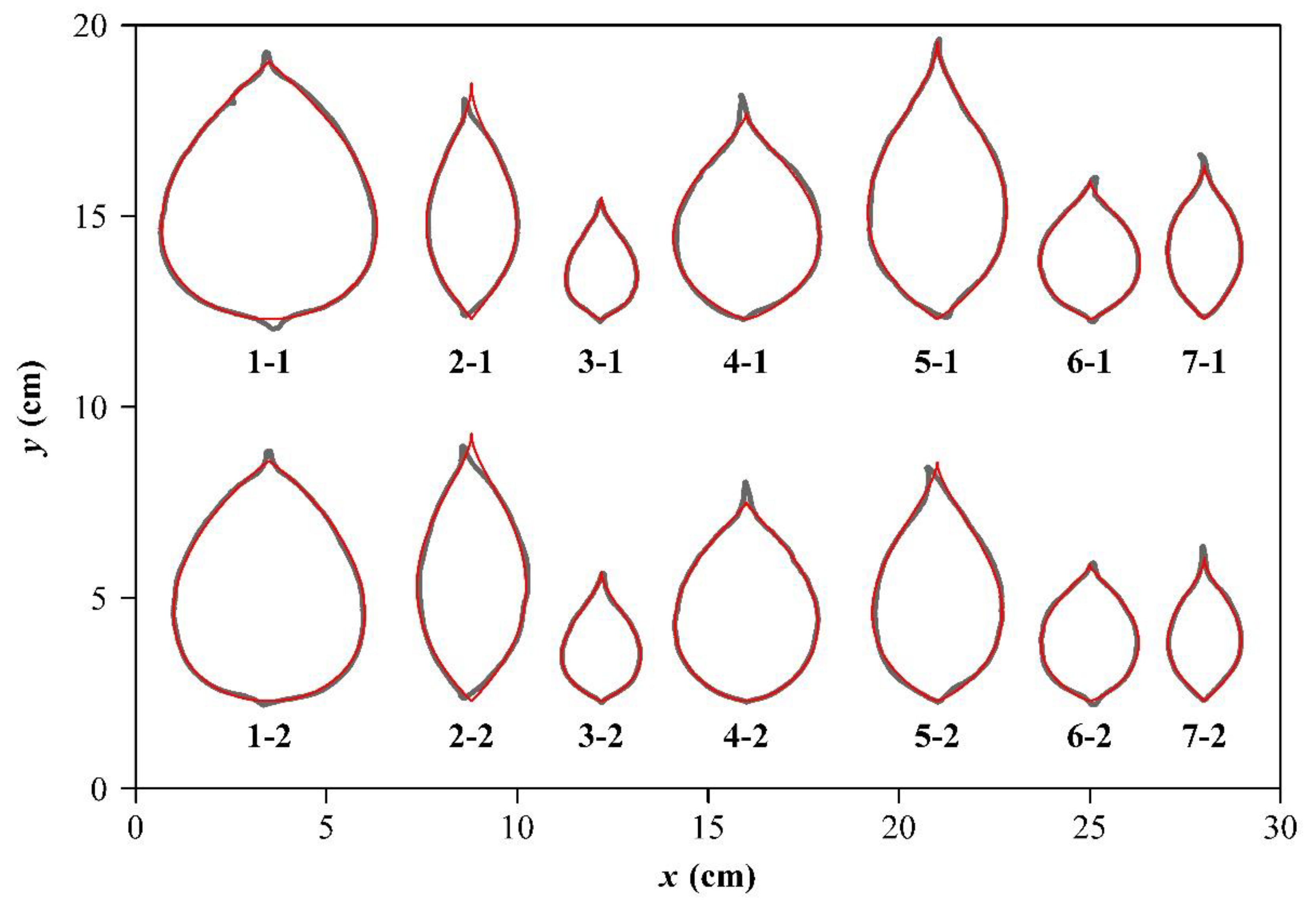
2.3. Samples of Representative Ovate Leaves
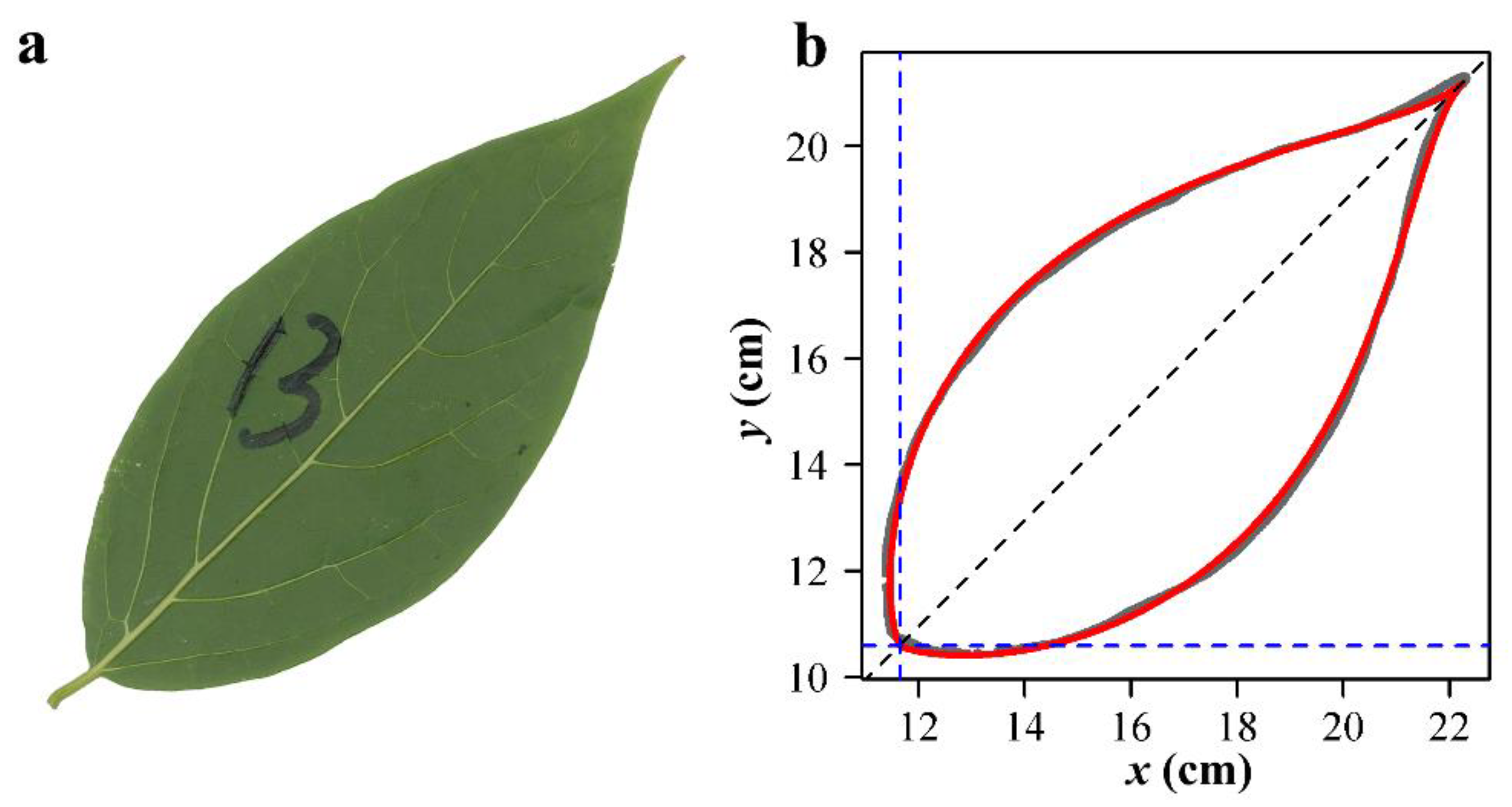
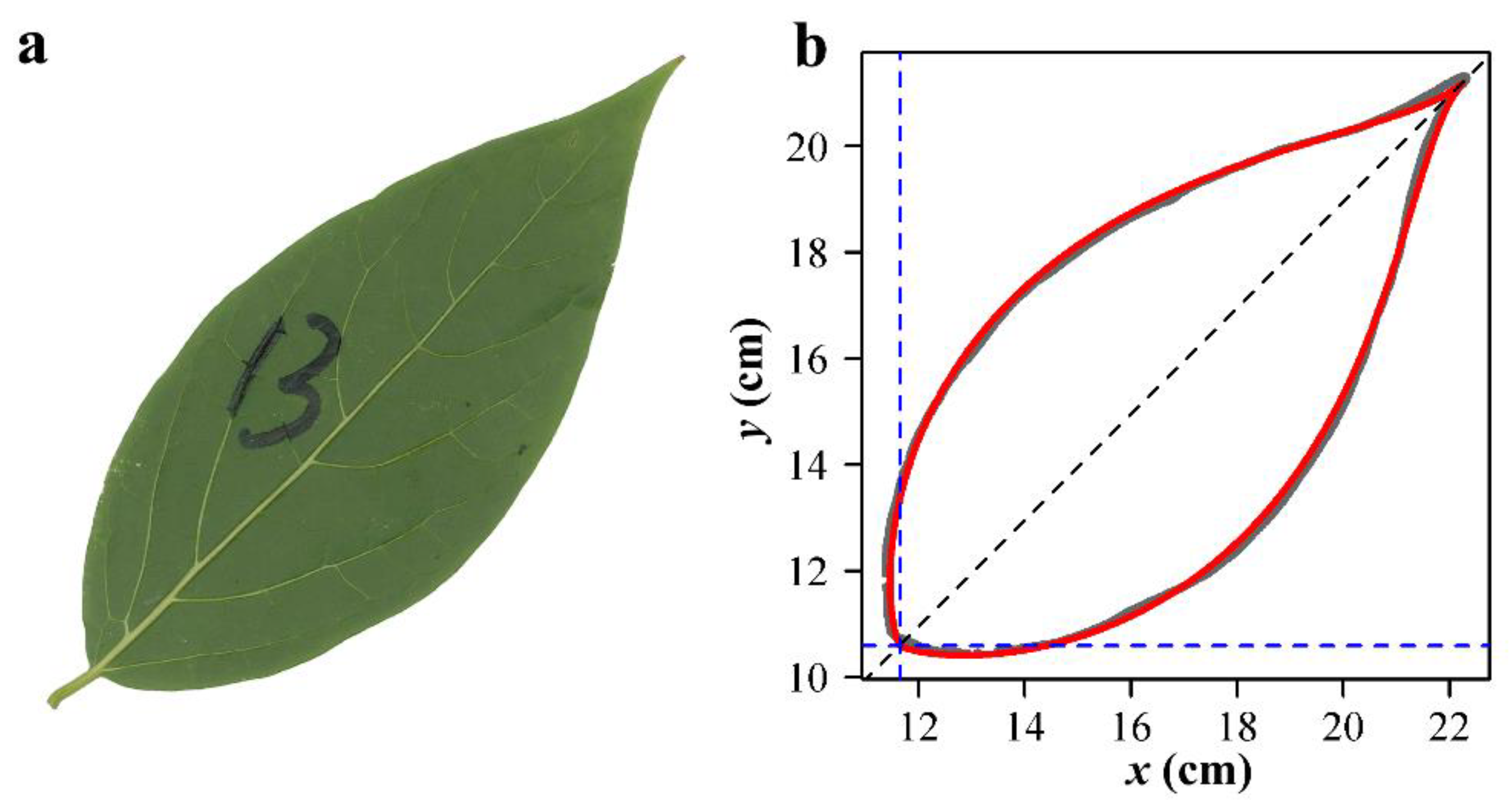
2.4. Leaf Image Processing and Data Acquisition
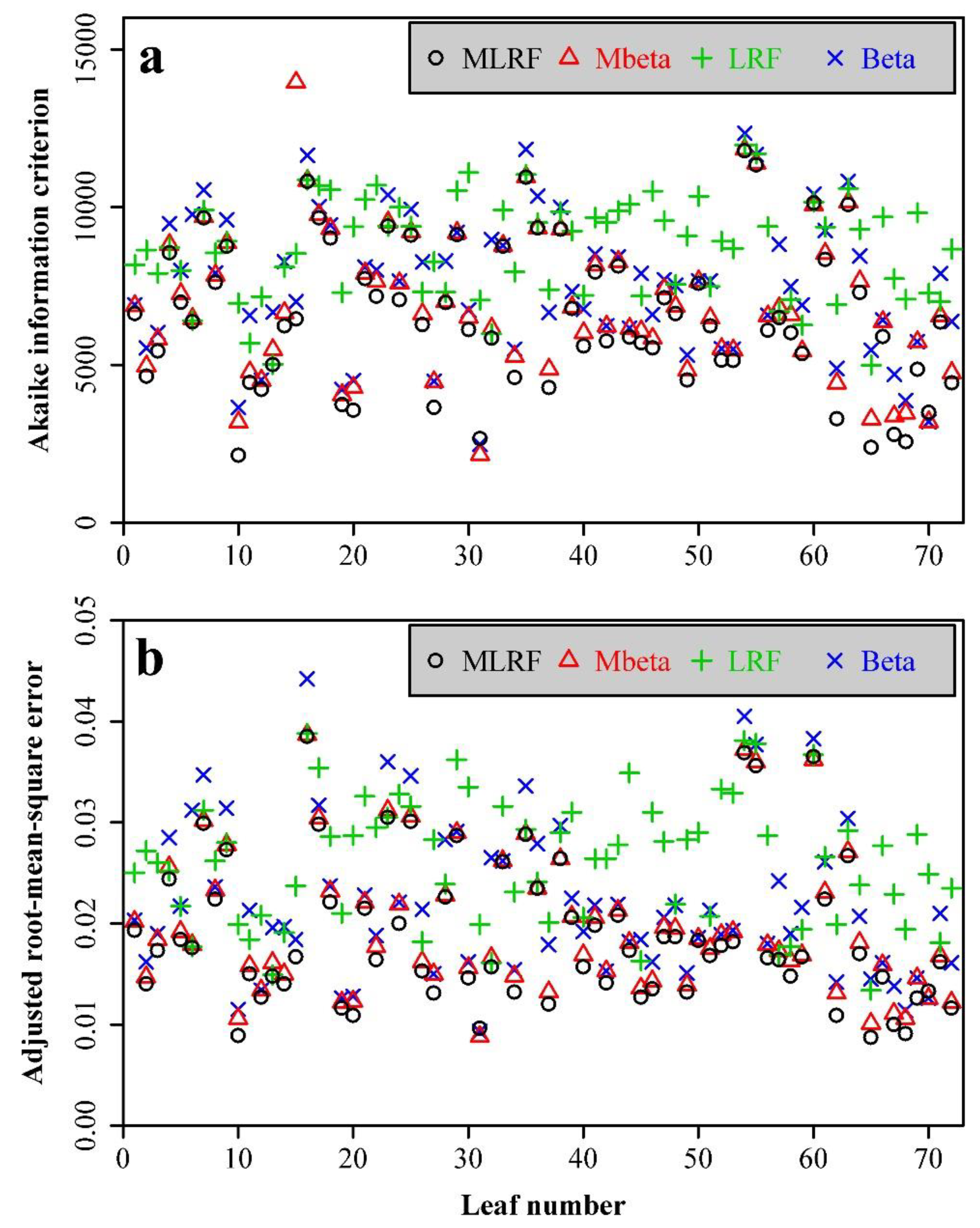
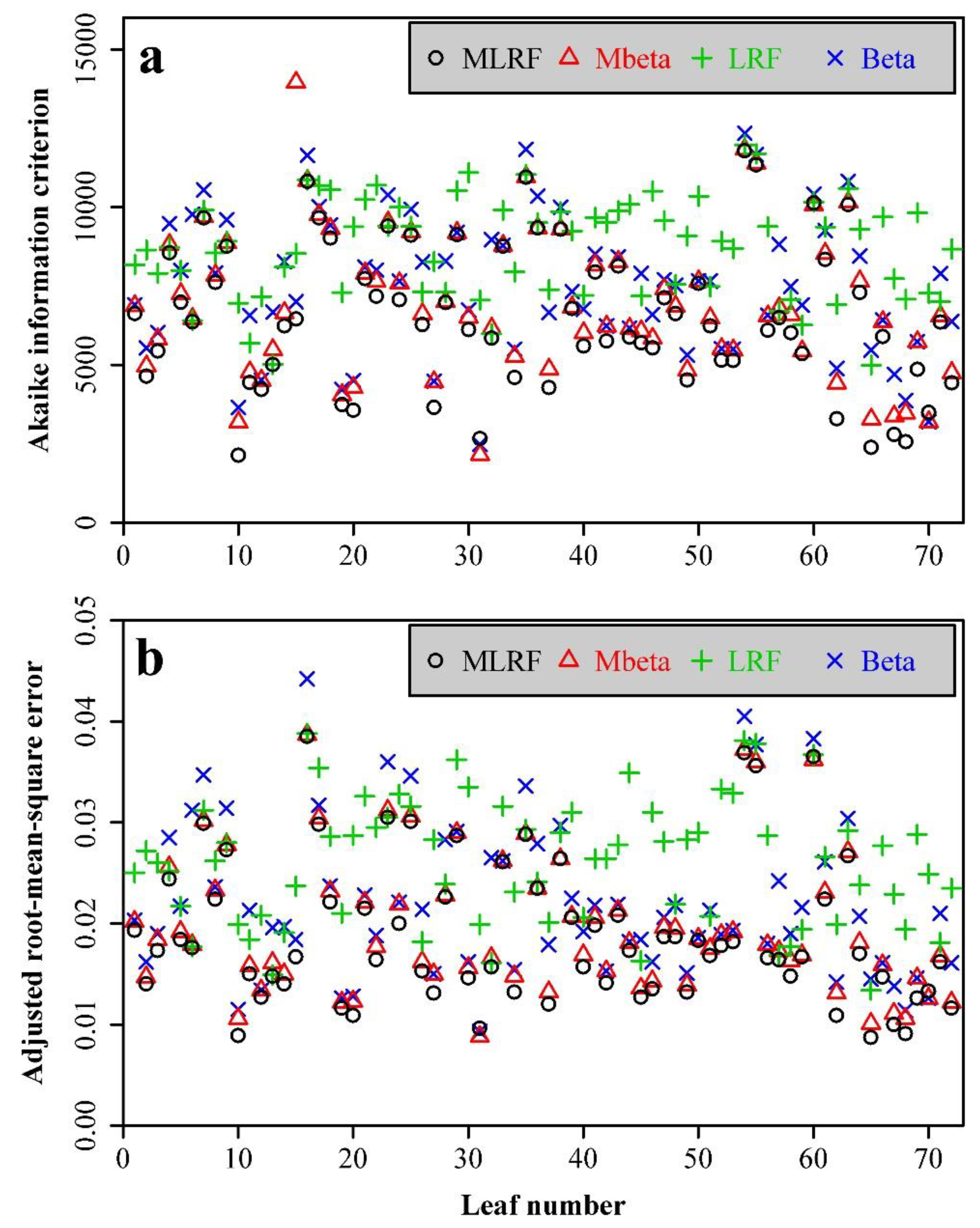
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, P.; Ratkowsky, D.A.; Li, Y.; Zhang, L.; Lin, S.; Gielis, J. General leaf-area geometric formula exists for plants—Evidence from the simplified Gielis equation. Forests 2018, 9, 714. [Google Scholar] [CrossRef] [Green Version]
- Gallaher, T.J.; Adams, D.C.; Attigala, L.; Burke, S.V.; Craine, J.M.; Duvall, M.R.; Klahs, P.C.; Sherratt, E.; Wysocki, W.P.; Clark, L.G. Leaf shape and size track habitat transitions across forest–grassland boundaries in the grass family (Poaceae). Evolution 2019, 73, 927–946. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.; Shi, P.; Royer, D.L.; Peppe, D.J.; Gallagher, R.V.; Li, Y.; Wang, R.; Wright, I.J. Leaf size estimation based on leaf length, width and shape. Ann. Bot. 2021, in press. [Google Scholar] [CrossRef]
- Gielis, J. A general geometric transformation that unifies a wide range of natural and abstract shapes. Am. J. Bot. 2003, 90, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.J.; Xu, Q.; Sandhu, H.S.; Gielis, J.; Ding, Y.L.; Li, H.R.; Dong, X.B. Comparison of dwarf bamboos (Indocalamus sp.) leaf parameters to determine relationship between spatial density of plants and total leaf area per plant. Ecol. Evol. 2015, 5, 4578–4589. [Google Scholar] [CrossRef]
- Lin, S.Y.; Zhang, L.; Reddy, G.V.P.; Hui, C.; Gielis, J.; Ding, Y.L.; Shi, P.J. A geometrical model for testing bilateral symmetry of bamboo leaf with a simplified Gielis equation. Ecol. Evol. 2016, 6, 6798–6806. [Google Scholar] [CrossRef]
- Su, J.; Niklas, K.J.; Huang, W.; Yu, X.; Yang, Y.; Shi, P. Lamina shape does not correlate with lamina surface area: An analysis based on the simplified Gielis equation. Glob. Ecol. Conserv. 2019, 19, e00666. [Google Scholar] [CrossRef]
- Yin, X.; Kropff, M.J.; McLaren, G.; Visperas, R.M. A nonlinear model for crop development as a function of temperature. Agric. For. Meteorol. 1995, 77, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lobry, J.R.; Rosso, L.; Flandrois, J.P. A FORTRAN subroutine for the determination of parameter confidence limits in non-linear models. Binary 1991, 3, 86–93. [Google Scholar]
- Rosso, L.; Lobry, J.R.; Flandrois, J.P. An unexpected correlation between cardinal temperatures of microbial growth highlighted by a new model. J. Theor. Biol. 1993, 162, 447–463. [Google Scholar] [CrossRef]
- Ratkowsky, D.A. Nonlinear Regression Modeling: A Unified Practical Approach; Marcel Dekker: New York, NY, USA, 1983. [Google Scholar]
- Ratkowsky, D.A.; Reddy, G.V.P. Empirical model with excellent statistical properties for describing temperature-dependent developmental rates of insects and mites. Ann. Entomol. Soc. Am. 2017, 110, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü.; Portsmuth, A.; Tobias, M. Leaf shape and venation pattern alter the support investments within leaf lamina in temperate species, a neglected source of leaf physiological differentiation. Funct. Ecol. 2007, 21, 28–40. [Google Scholar] [CrossRef]
- Shi, P.; Fan, M.; Ratkowsky, D.A.; Huang, J.; Wu, H.; Chen, L.; Fang, S.; Zhang, C. Comparison of two ontogenetic growth equations for animals and plants. Ecol. Model. 2017, 349, 1–10. [Google Scholar] [CrossRef]
- Dornbusch, T.; Watt, J.; Baccar, R.; Fournier, C.; Andrieu, B. A comparative analysis of leaf shape of wheat, barley and maize using an empirical shape model. Ann. Bot. 2011, 107, 865–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kincaid, D.T.; Schneider, R.B. Quantification of leaf shape with a microcomputer and Fourier transform. Can. J. Bot. 1983, 61, 2333–2342. [Google Scholar] [CrossRef]
- Thomas, S.C.; Bazzaz, F.A. Elevated CO2 and leaf shape: Are dandelions getting toothier? Am. J. Bot. 1996, 83, 106–111. [Google Scholar] [CrossRef]
- Niinemets, Ü. Adjustment of foliage structure and function to a canopy light gradient in two co-existing deciduous trees. Variability in leaf inclination angles in relation to petiole morphology. Trees Struct. Funct. 1998, 12, 446–451. [Google Scholar] [CrossRef]
- Santiago, L.S.; Kim, S.-C. Correlated evolution of leaf shape and physiology in the woody Sonchus alliance (Asteraceae: Sonchinae) in Macaronesia. Int. J. Plant Sci. 2009, 170, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Peppe, D.J.; Royer, D.L.; Gariglino, B.; Oliver, S.Y.; Newman, S.; Leight, E.; Enikolopov, G.; Fernandez-Burgos, M.; Herrera, F.; Adams, J.M.; et al. Sensitivity of leaf size and shape to climate: Global patterns and paleoclimatic applications. New Phytol. 2011, 190, 724–739. [Google Scholar] [CrossRef] [Green Version]
- Royer, D.L.; Wilf, P. Why do toothed leaves correlate with cold climates? Gas exchange at leaf margins provides new insights into a classic paleotemperature proxy. Int. J. Plant Sci. 2006, 167, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Fanourakis, D.; Kazakos, F.; Nektarios, P.A. Allometric individual leaf area estimation in Chrysanthemum. Agronomy 2021, 11, 795. [Google Scholar] [CrossRef]
- Spiess, A.-N.; Neumeyer, N. An evaluation of R squared as an inadequate measure for nonlinear models in pharmacological and biochemical research: A Monte Carlo approach. BMC Pharmacol. 2010, 10, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.L.; Li, X.M.; Huang, H. Leaf shape simulation of castor bean and its application in nondestructive leaf area estimation. Int. J. Agric. Biol. Eng. 2019, 12, 135–140. [Google Scholar] [CrossRef]
- Shi, P.; Ratkowsky, D.A.; Gielis, J. The generalized Gielis geometric equation and its application. Symmetry 2020, 12, 645. [Google Scholar] [CrossRef] [Green Version]
- Nelder, J.A.; Mead, R. A simplex algorithm for function minimization. Comput. J. 1965, 7, 308–313. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 1 January 2020).
- Wang, T.; Si, Y.; Dai, H.; Li, C.; Gao, C.; Dong, Z.; Jiang, L. Apex structures enhance water drainage on leaves. Proc. Natl. Acad. Sci. USA 2020, 117, 1890–1894. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, P.J.H.; DeMichele, D.W. Reaction kinetics of poikilotherm development. J. Theor. Biol. 1977, 64, 648–670. [Google Scholar] [CrossRef]
- Schoolfield, R.M.; Sharpe, P.J.H.; Magnuson, C.E. Non-linear regression of biological temperature-dependent rate models based on absolute reaction rate theory. J. Theor. Biol. 1981, 88, 719–731. [Google Scholar] [CrossRef]
- Ratkowsky, D.A.; Olley, J.; Ross, T. Unifying temperature effects on the growth rate of bacteria and the stability of globular proteins. J. Theor. Biol. 2005, 233, 351–362. [Google Scholar] [CrossRef]
- Voorend, W.; Lootens, P.; Nelissen, H.; Roldán-Ruiz, I.; Inzé, D.; Muylle, H. LEAF-E: A tool to analyze grass leaf growth using function fitting. Plant Methods 2014, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Camargo Neto, J.; Meyer, G.E.; Jones, D.D.; Samal, A.K. Plant species identification using Elliptic Fourier leaf shape analysis. Comput. Electron. Agric. 2006, 50, 121–134. [Google Scholar] [CrossRef]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning: Data Mining, Inference, and Prediction, 3rd ed.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Ratkowsky, D.A.; Lowry, R.K.; McMeekin, T.A.; Stokes, A.N.; Chandler, R.E. Model for bacterial culture growth rate throughout the entire biokinetic temperature range. J. Bacteriol. 1983, 154, 1222–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lactin, D.J.; Holliday, N.J.; Johnson, D.L.; Craigen, R. Improved rate model of temperature-dependent development by arthropods. Environ. Entomol. 1995, 24, 68–75. [Google Scholar] [CrossRef]
- Briére, J.-F.; Pracros, P.; Le Roux, A.-Y.; Pierre, J.-S. A novel rate model of temperature-dependent development for arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Coussement, J.R.; Steppe, K.; Lootens, P.; Roldán-Ruiz, I.; De Swaef, T. A flexible geometric model for leaf shape descriptions with high accuracy. Silva Fennica 2018, 52, 7740. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Latin Name | Locality | Longitude (°), Latitude (°) | Sampling Date |
|---|---|---|---|---|
| 1 | Neocinnamomum lecomtei H. Liu | Hainan, China | 107.8394, 22.4641 | 2017.03.09 |
| 2 | Neocinnamomum complanifructum S.K. Lee et F.N. Wei | Yunnan, China | 104.0189, 22.4669 | 2017.10.09 |
| 3 | Neocinnamomum fargesii (Lecomte) Kosterm. | Sichuan, China | 110.2984, 31.3034 | 2017.02.20 |
| 4 | Neocinnamomum delavayi (Lecomte) H. Liu | Sichuan, China | 103.9540, 28.5396 | 2017.09.04 |
| 5 | Neocinnamomum caudatum (Nees) Merr. | Hainan, China | 98.2949, 24.8070 | 2017.05.08 |
| 6 | Neocinnamomum caudatum (Nees) Merr. var. macrocarpum W.B. Xu et B.S. Xia | Guangxi, China | 106.3458, 23.5951 | 2017.11.17 |
| 7 | Neocinnamomum mekongense (Hand.-Mazz.) Kosterm. | Yunnan, China | 99.9285, 24.5804 | 2017.05.08 |
| Leaf No. | Beta | Mbeta | LRF | MLRF | ||||
|---|---|---|---|---|---|---|---|---|
| AIC | RMSEadj | AIC | RMSEadj | AIC | RMSEadj | AIC | RMSEadj | |
| 1-1 | 6158 | 0.0288 | 6119 | 0.0286 | 9288 | 0.0522 | 6240 | 0.0292 |
| 1-2 | 4947 | 0.0279 | 4467 | 0.0252 | 8623 | 0.0599 | 4552 | 0.0256 |
| 2-1 | 2508 | 0.0375 | 1968 | 0.0320 | 1998 | 0.0323 | 1940 | 0.0318 |
| 2-2 | 4013 | 0.0384 | 3774 | 0.0362 | 3712 | 0.0357 | 3712 | 0.0357 |
| 3-1 | 277 | 0.0372 | −653 | 0.0232 | −607 | 0.0238 | −648 | 0.0232 |
| 3-2 | 349 | 0.0342 | −702 | 0.0209 | −621 | 0.0218 | −723 | 0.0207 |
| 4-4 | 4587 | 0.0448 | 4425 | 0.0429 | 4664 | 0.0457 | 4380 | 0.0424 |
| 4-2 | 1037 | 0.0179 | 1035 | 0.0179 | 3816 | 0.0364 | 675 | 0.0163 |
| 5-1 | 4302 | 0.0309 | 2647 | 0.0215 | 2617 | 0.0214 | 2597 | 0.0213 |
| 5-2 | 3933 | 0.0378 | 3490 | 0.0337 | 3678 | 0.0354 | 3423 | 0.0331 |
| 6-1 | 1211 | 0.0379 | 999 | 0.0348 | 973 | 0.0345 | 936 | 0.0339 |
| 6-2 | 1189 | 0.0376 | 1165 | 0.0372 | 1293 | 0.0392 | 1136 | 0.0367 |
| 7-1 | 86 | 0.0265 | −294 | 0.0229 | −481 | 0.0213 | −483 | 0.0213 |
| 7-2 | 489 | 0.0333 | 53 | 0.0280 | 7 | 0.0275 | −8 | 0.0273 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, P.; Yu, K.; Niklas, K.J.; Schrader, J.; Song, Y.; Zhu, R.; Li, Y.; Wei, H.; Ratkowsky, D.A. A General Model for Describing the Ovate Leaf Shape. Symmetry 2021, 13, 1524. https://doi.org/10.3390/sym13081524
Shi P, Yu K, Niklas KJ, Schrader J, Song Y, Zhu R, Li Y, Wei H, Ratkowsky DA. A General Model for Describing the Ovate Leaf Shape. Symmetry. 2021; 13(8):1524. https://doi.org/10.3390/sym13081524
Chicago/Turabian StyleShi, Peijian, Kexin Yu, Karl J. Niklas, Julian Schrader, Yu Song, Renbin Zhu, Yang Li, Hailin Wei, and David A. Ratkowsky. 2021. "A General Model for Describing the Ovate Leaf Shape" Symmetry 13, no. 8: 1524. https://doi.org/10.3390/sym13081524
APA StyleShi, P., Yu, K., Niklas, K. J., Schrader, J., Song, Y., Zhu, R., Li, Y., Wei, H., & Ratkowsky, D. A. (2021). A General Model for Describing the Ovate Leaf Shape. Symmetry, 13(8), 1524. https://doi.org/10.3390/sym13081524