
Non-Coding RNAs and Reactive Oxygen Species–Symmetric Players of the Pathogenesis Associated with Bacterial and Viral Infections



 ,
, 
Abstract
1. Introduction
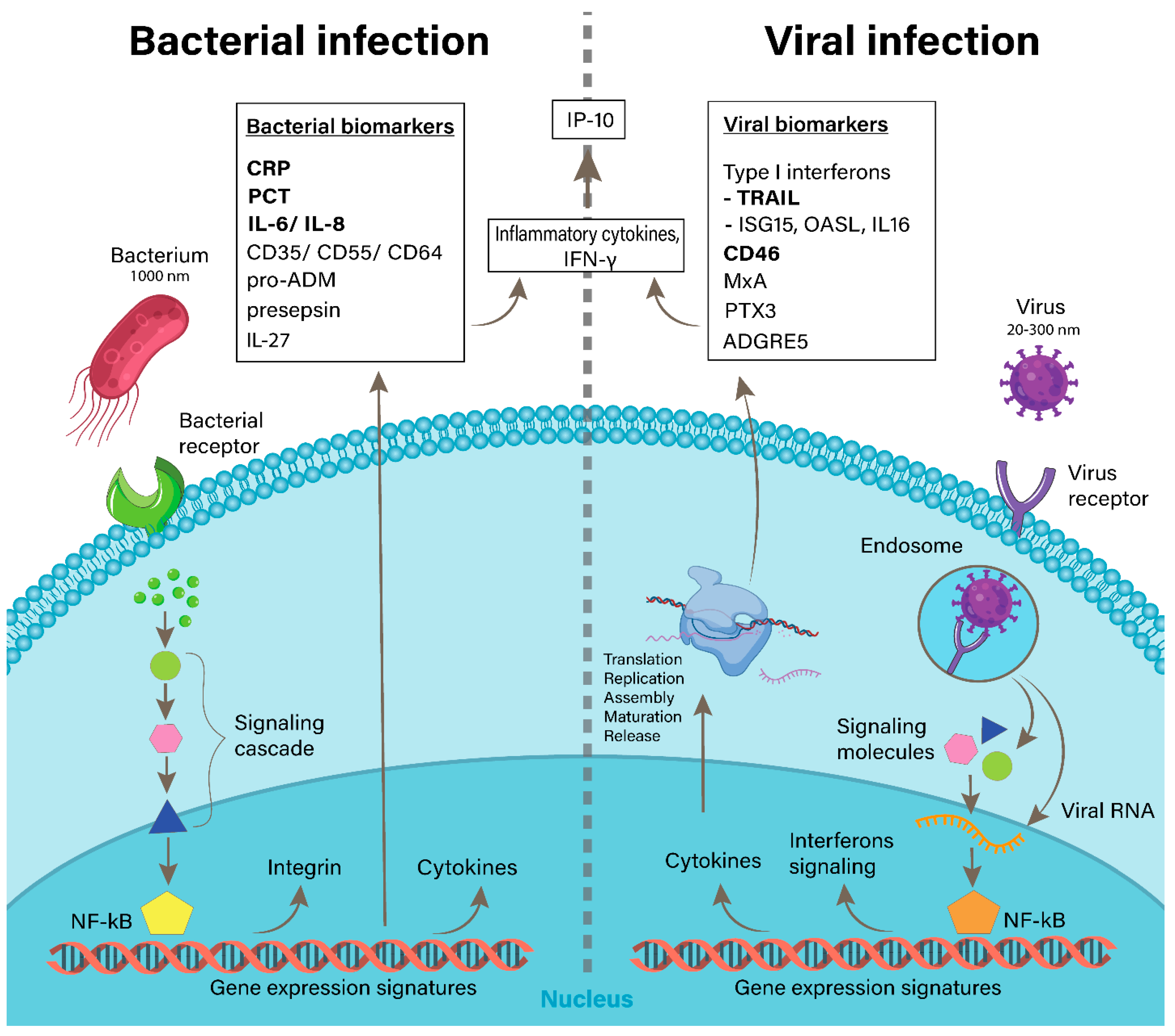
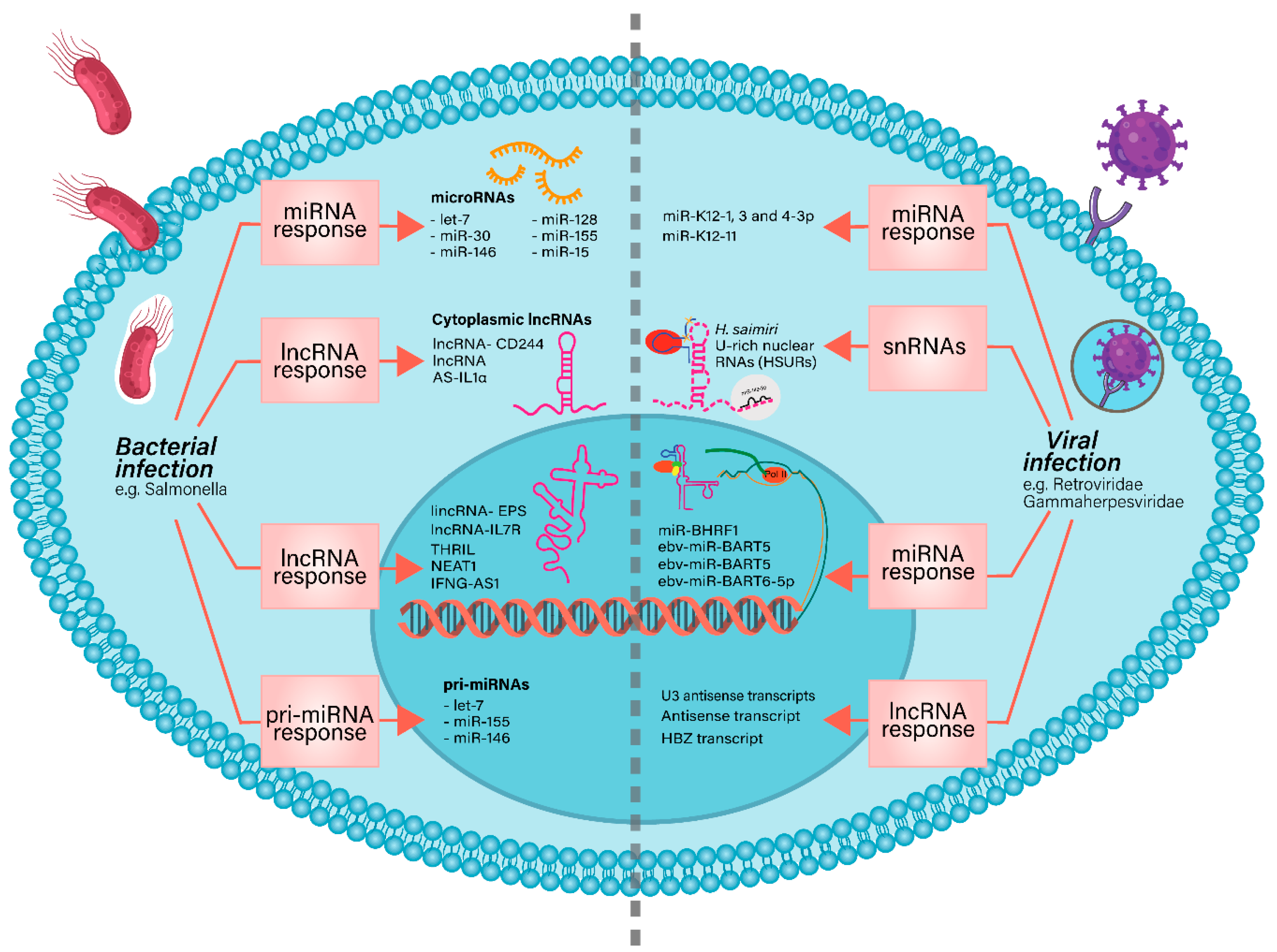
2. Non-Coding RNAs in Bacterial and Viral Infections
2.1. NcRNAs Involved in Bacterial Infections
2.2. NcRNAs in Viral Infections
2.3. Other ncRNAs Involved in Bacterial or Viral Infections
3. Reactive Oxygen Species in Bacterial and Viral Infections
4. The Interplay between ncRNAs and ROS in Bacterial and Viral Infections
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kaufmann, S.H.E.; Dorhoi, A.; Hotchkiss, R.S.; Bartenschlager, R. Host-Directed Therapies for Bacterial and Viral Infections. Nat. Rev. Drug Discov. 2018, 17, 35–56. [Google Scholar] [CrossRef]
- Smith, K.M.; Machalaba, C.C.; Seifman, R.; Feferholtz, Y.; Karesh, W.B. Infectious Disease and Economics: The Case for Considering Multi-Sectoral Impacts. One Health 2019, 7, 10080. [Google Scholar] [CrossRef] [PubMed]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate Point-Prevalence Survey of Health Care–Associated Infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef]
- Zimlichman, E.; Henderson, D.; Tamir, O.; Franz, C.; Song, P.; Yamin, C.K.; Keohane, C.; Denham, C.R.; Bates, D.W. Health Care-Associated Infections: AMeta-Analysis of Costs and Financial Impact on the US Health Care System. JAMA Intern. Med. 2013, 173, 2039–2046. [Google Scholar] [CrossRef] [PubMed]
- Riley, L.W.; Blanton, R.E. Advances in Molecular Epidemiology of Infectious Diseases: Definitions, Approaches, and Scope of the Field. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Tobin, N.H.; Campbell, A.J.P.; Zerr, D.M.; Melvin, A.J. Life-Threatening Viral Diseases and Their Treatment. In Pediatric Critical Care; Elsevier Inc.: Amsterdam, The Netherlands, 2011; pp. 1324–1335. [Google Scholar]
- Aliberti, S.; Kaye, K.S. The Changing Microbiologic Epidemiology of Community-Acquired Pneumonia. Postgrad. Med. 2013, 125, 31–42. [Google Scholar] [CrossRef]
- Self, W.H.; Balk, R.A.; Grijalva, C.G.; Williams, D.J.; Zhu, Y.; Anderson, E.J.; Waterer, G.W.; Courtney, D.M.; Bramley, A.M.; Trabue, C.; et al. Procalcitonin as a Marker of Etiology in Adults Hospitalized with Community-Acquired Pneumonia. Clin. Infect. Dis. 2017, 65, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Cawcutt, K.; Kalil, A.C. Pneumonia with Bacterial and Viral Coinfection. Curr. Opin. Crit. Care 2017, 23, 385–390. [Google Scholar] [CrossRef]
- Ruuskanen, O.; Järvinen, A. What Is the Real Role of Respiratory Viruses in Severe Community-Acquired Pneumonia? In Clinical Infectious Diseases; Oxford University Press: Oxford, UK, 2014; pp. 71–73. [Google Scholar]
- Kang, Y.; Xu, S. Comprehensive Overview of COVID-19 Based on Current Evidence. Dermatol. Ther. 2020, 33, e13525. [Google Scholar] [CrossRef] [PubMed]
- Fischer Walker, C.L.; Rudan, I.; Liu, L.; Nair, H.; Theodoratou, E.; Bhutta, Z.A.; O’Brien, K.L.; Campbell, H.; Black, R.E. Global Burden of Childhood Pneumonia and Diarrhoea. Lancet 2013, 381, 1405–1416. [Google Scholar] [CrossRef]
- Qureshi, A.; Niazi, J.H. Biosensors for Detecting Viral and Bacterial Infections Using Host Biomarkers: A Review. Analyst 2020, 145, 7825–7848. [Google Scholar] [CrossRef]
- Tsao, Y.-T.; Tsai, Y.-H.; Liao, W.-T.; Shen, C.-J.; Shen, C.-F.; Cheng, C.-M. Differential Markers of Bacterial and Viral Infections in Children for Point-of-Care Testing. Trends Mol. Med. 2020, 26, 1118–1132. [Google Scholar] [CrossRef]
- Chang, Z.; Wang, Y.; Zhou, X.; Long, J.E. STAT3 Roles in Viral Infection: Antiviral or Proviral? Future Virol. 2018, 13, 557–574. [Google Scholar] [CrossRef]
- Sampson, D.L.; Fox, B.A.; Yager, T.D.; Bhide, S.; Cermelli, S.; McHugh, L.C.; Seldon, T.A.; Brandon, R.A.; Sullivan, E.; Zimmerman, J.J.; et al. A Four-Biomarker Blood Signature Discriminates Systemic Inflammation Due to Viral Infection versus Other Etiologies. Sci. Rep. 2017, 7, 2914. [Google Scholar] [CrossRef]
- Esteller, M. Non-Coding RNAs in Human Disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Lekka, E.; Hall, J. Noncoding RNAs in Disease. FEBS Lett. 2018, 592, 2884–2900. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.F.; Lee, E.S. Non-Coding RNA: What Is Functional and What Is Junk? Front. Genet. 2015, 5, 2. [Google Scholar] [CrossRef]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent Deletions and Down-Regulation of Micro-RNA Genes MiR15 and MiR16 at 13q14 in Chronic Lymphocytic Leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- Choudhuri, S. Small Noncoding RNAs: Biogenesis, Function, and Emerging Significance in Toxicology. J. Biochem. Mol. Toxicol. 2010, 24, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2017; Volume 1509, pp. 1–10. [Google Scholar]
- Hombach, S.; Kretz, M. Non-Coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar]
- Yang, B.; Chen, Y.; Shi, J. Reactive Oxygen Species (ROS)-Based Nanomedicine. Chem. Rev. 2019, 119, 4881–4985. [Google Scholar] [CrossRef]
- Sun, J.; Wang, J.; Li, L.; Wu, Z.; Chen, X.; Yuan, J. ROS induced by spring viraemia of carp virus activate the inflammatory response via the MAPK/AP-1 and PI3K signaling pathways. Fish Shellfish Immunol. 2020, 101, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Ware, H.H.; Kulkarni, V.V.; Wang, Y.; Pantaleón García, J.; Leiva Juarez, M.; Kirkpatrick, C.T.; Wali, S.; Syed, S.; Kontoyiannis, A.D.; Sikkema, W.K.A.; et al. Inducible lung epithelial resistance requires multisource reactive oxygen species generation to protect against bacterial infections. PLoS ONE 2019, 14, e0208216. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of MicroRNA Biogenesis and Its Crosstalk with Other Cellular Pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long Non-Coding RNA: Classification, Biogenesis and Functions in Blood Cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Quinn, J.J.; Chang, H.Y. Unique Features of Long Non-Coding RNA Biogenesis and Function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Chen, H.; Luo, F.; Zhou, H.; Li, Z. Roles of Long Noncoding RNAs in Bacterial Infection. Life Sci. 2020, 263, 118579. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Li, N.; Gu, J.; Zhuang, Y.; Li, Q.; Wang, H.G.; Fang, Y.; Yu, B.; Zhang, J.Y.; Xie, Q.H.; et al. Compromised Autophagy by MIR30Bbenefits the Intracellular Survival of Helicobacter pylori. Autophagy 2012, 8, 1045–1057. [Google Scholar] [CrossRef]
- Lind, E.F.; Elford, A.R.; Ohashi, P.S. Micro-RNA 155 Is Required for Optimal CD8 + T Cell Responses to Acute Viral and Intracellular Bacterial Challenges. J. Immunol. 2013, 190, 1210–1216. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Xu, S.; Liu, X.; Zhang, Q.; Xu, X.; Liu, M.; Hua, M.; Li, N.; Yao, H.; Cao, X. The MicroRNA MiR-29 Controls Innate and Adaptive Immune Responses to Intracellular Bacterial Infection by Targeting Interferon-γ. Nat. Immunol. 2011, 12, 861–869. [Google Scholar] [CrossRef]
- Wang, Y.; Zhong, H.; Xie, X.; Chen, C.Y.; Huang, D.; Shen, L.; Zhang, H.; Chen, Z.W.; Zeng, G. Long Noncoding RNA Derived from CD244 Signaling Epigenetically Controls CD8+ T-Cell Immune Responses in Tuberculosis Infection. Proc. Natl. Acad. Sci. USA 2015, 112, E3883–E3892. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Huang, Z.; Luo, Q.; Qing, C. The Expression of LncRNA NEAT1 in Human Tuberculosis and Its Antituberculosis Effect. BioMed Res. Int. 2018, 2018, 9529072. [Google Scholar] [CrossRef] [PubMed]
- Penkala, I.; Wang, J.; Syrett, C.M.; Goetzl, L.; López, C.B.; Anguera, M.C. LncRHOXF1, a Long Noncoding RNA from the X Chromosome That Suppresses Viral Response Genes during Development of the Early Human Placenta. Mol. Cell. Biol. 2016, 36, 1764–1775. [Google Scholar] [CrossRef]
- Reshi, M.L.; Su, Y.C.; Hong, J.R. RNA Viruses: ROS-Mediated Cell Death. Int. J. Cell Biol. 2014, 2014, 467452. [Google Scholar] [CrossRef]
- Paiva, C.N.; Bozza, M.T. Are reactive oxygen species always detrimental to pathogens? Antioxid. Redox Signal. 2014, 20, 1000–1037. [Google Scholar] [CrossRef]
- Aryal, B.; Suárez, Y. Non-Coding RNA Regulation of Endothelial and Macrophage Functions during Atherosclerosis. Vasc. Pharmacol. 2019, 114, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.J.; Tay, Y. Noncoding RNA: RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef]
- Sharma, N.; Singh, S.K. Implications of Non-Coding RNAs in Viral Infections. Rev. Med. Virol. 2016, 26, 356–368. [Google Scholar] [CrossRef]
- Duval, M.; Cossart, P.; Lebreton, A. Mammalian MicroRNAs and Long Noncoding RNAs in the Host-Bacterial Pathogen Crosstalk. Semin. Cell Dev. Biol. 2017, 65, 11–19. [Google Scholar] [CrossRef]
- Peterson, S.M.; Thompson, J.A.; Ufkin, M.L.; Sathyanarayana, P.; Liaw, L.; Congdon, C.B. Common Features of MicroRNA Target Prediction Tools. Front. Genet. 2014, 5, 23. [Google Scholar] [CrossRef]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-ΚB-Dependent Induction of MicroRNA MiR-146, an Inhibitor Targeted to Signaling Proteins of Innate Immune Responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Ibeagha-Awemu, E.M.; Liang, G.; Beaudoin, F.; Zhao, X.; Guan, L.L. Transcriptome MicroRNA Profiling of Bovine Mammary Epithelial Cells Challenged with Escherichia coli or Staphylococcus aureus Bacteria Reveals Pathogen Directed MicroRNA Expression Profiles. BMC Genom. 2014, 15, 181. [Google Scholar] [CrossRef]
- Zeng, F.R.; Tang, L.J.; He, Y.; Garcia, R.C. An Update on the Role of MiRNA-155 in Pathogenic Microbial Infections. Microbes Infect. 2015, 17, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Maudet, C.; Mano, M.; Eulalio, A. MicroRNAs in the Interaction between Host and Bacterial Pathogens. FEBS Lett. 2014, 588, 4140–4147. [Google Scholar] [CrossRef]
- Josset, L.; Tisoncik-Go, J.; Katze, M.G. Noncoding RNA Expression during Viral Infection: The Long and the Short of It. In MicroRNAs and Other Non-Coding RNAs in Inflammation; Springer International Publishing: Cham, Switzerland, 2015; pp. 107–137. [Google Scholar]
- Munschauer, M.; Vogel, J. Nuclear Lnc RNA Stabilization in the Host Response to Bacterial Infection. EMBO J. 2018, 37, e99875. [Google Scholar] [CrossRef] [PubMed]
- Zur Bruegge, J.; Einspanier, R.; Sharbati, S. A Long Journey Ahead: Long Non-Coding RNAs in Bacterial Infections. Front. Cell. Infect. Microbiol. 2017, 7, 95. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Mohapatra, G.; Ahmad, S.M.; Rana, S.; Jain, S.; Khalsa, J.K.; Srikanth, C.V. Salmonella Engages Host MicroRNAs To Modulate SUMOylation: A New Arsenal for Intracellular Survival. Mol. Cell. Biol. 2015, 35, 2932–2946. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, M.V.S.; Ni, B.; Morris, J.D.; Brooks, M.N.; Carlson, T.K.; Bakthavachalu, B.; Schoenberg, D.R.; Torrelles, J.B.; Schlesinger, L.S. Mycobacterium tuberculosis Lipomannan Blocks TNF Biosynthesis by Regulating Macrophage MAPK-Activated Protein Kinase 2 (MK2) and MicroRNA MiR-125b. Proc. Natl. Acad. Sci. USA 2011, 108, 17408–17413. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Kaul, V.; Mehra, A.; Chatterjee, S.; Tousif, S.; Dwivedi, V.P.; Suar, M.; Van Kaer, L.; Bishai, W.R.; Das, G. Mycobacterium tuberculosis Controls MicroRNA-99b (MiR-99b) Expression in Infected Murine Dendritic Cells to Modulate Host Immunity. J. Biol. Chem. 2013, 288, 5056–5061. [Google Scholar] [CrossRef]
- Kumar, R.; Halder, P.; Sahu, S.K.; Kumar, M.; Kumari, M.; Jana, K.; Ghosh, Z.; Sharma, P.; Kundu, M.; Basu, J. Identification of a Novel Role of ESAT-6-Dependent MiR-155 Induction during Infection of Macrophages with Mycobacterium tuberculosis. Cell. Microbiol. 2012, 14, 1620–1631. [Google Scholar] [CrossRef]
- Kumar, M.; Sahu, S.K.; Kumar, R.; Subuddhi, A.; Maji, R.K.; Jana, K.; Gupta, P.; Raffetseder, J.; Lerm, M.; Ghosh, Z.; et al. MicroRNA Let-7 Modulates the Immune Response to Mycobacterium tuberculosis Infection via Control of A20, an Inhibitor of the NF-ΚB Pathway. Cell Host Microbe 2015, 17, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Clare, S.; John, V.; Walker, A.W.; Hill, J.L.; Abreu-Goodger, C.; Hale, C.; Goulding, D.; Lawley, T.D.; Mastroeni, P.; Frankel, G.; et al. Enhanced Susceptibility to Citrobacter rodentium Infection in MicroRNA-155-Deficient Mice. Infect. Immun. 2013, 81, 723–732. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ro, Y.T.; Jo, G.H.; Jung, S.A.; Lee, E.H.; Shin, J.; Lee, J.H. Salmonella-Induced MiR-155 Enhances Necroptotic Death in Macrophage Cells via Targeting RIP1/3. Mol. Med. Rep. 2018, 18, 5133–5140. [Google Scholar] [CrossRef]
- Cremer, T.J.; Ravneberg, D.H.; Clay, C.D.; Piper-Hunter, M.G.; Marsh, C.B.; Elton, T.S.; Gunn, J.S.; Amer, A.; Kanneganti, T.D.; Schlesinger, L.S.; et al. MiR-155 Induction by F. Novicida but Not the Virulent F. Tularensis Results in SHIP down-Regulation and Enhanced pro-Inflammatory Cytokine Response. PLoS ONE 2009, 4, e8508. [Google Scholar] [CrossRef]
- Li, M.; Wang, J.; Fang, Y.; Gong, S.; Li, M.; Wu, M.; Lai, X.; Zeng, G.; Wang, Y.; Yang, K.; et al. MicroRNA-146a Promotes Mycobacterial Survival in Macrophages through Suppressing Nitric Oxide Production. Sci. Rep. 2016, 6, 23351. [Google Scholar] [CrossRef]
- Zhang, T.; Yu, J.; Zhang, Y.; Li, L.; Chen, Y.; Li, D.; Liu, F.; Zhang, C.Y.; Gu, H.; Zen, K. Salmonella enterica Serovar Enteritidis Modulates Intestinal Epithelial Mir-128 Levels to Decrease Macrophage Recruitment via Macrophage Colony-Stimulating Factor. J. Infect. Dis. 2014, 209, 2000–2011. [Google Scholar] [CrossRef]
- Gomez, J.A.; Wapinski, O.L.; Yang, Y.W.; Bureau, J.F.; Gopinath, S.; Monack, D.M.; Chang, H.Y.; Brahic, M.; Kirkegaard, K. The NeST Long NcRNA Controls Microbial Susceptibility and Epigenetic Activation of the Interferon-γ Locus. Cell 2013, 152, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, C.Y.; Yedavalli, V.S.R.K.; Jeang, K.T. NEAT1 Long Noncoding RNA and Paraspeckle Bodies Modulate HIV-1 Posttranscriptional Expression. mBio 2013, 4, e00596-12. [Google Scholar] [CrossRef] [PubMed]
- Imamura, K.; Imamachi, N.; Akizuki, G.; Kumakura, M.; Kawaguchi, A.; Nagata, K.; Kato, A.; Kawaguchi, Y.; Sato, H.; Yoneda, M.; et al. Long Noncoding RNA NEAT1-Dependent SFPQ Relocation from Promoter Region to Paraspeckle Mediates IL8 Expression upon Immune Stimuli. Mol. Cell 2014, 53, 393–406. [Google Scholar] [CrossRef]
- Laha, S.; Saha, C.; Dutta, S.; Basu, M.; Chatterjee, R.; Ghosh, S.; Bhattacharyya, N.P. In Silico Analysis of Altered Expression of Long Non-Coding RNA in SARS-CoV-2 Infected Cells and Their Possible Regulation by STAT1, STAT3 and Interferon Regulatory Factors. Heliyon 2021, 7, e06395. [Google Scholar] [CrossRef]
- Li, C.; Hu, X.; Li, L.; Li, J.H. Differential MicroRNA Expression in the Peripheral Blood from Human Patients with COVID-19. J. Clin. Lab. Anal. 2020, 34, e23590. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.R.; Abd-Aziz, N.; Affendi, S.; Poh, C.L. Role of MicroRNAs in Antiviral Responses to Dengue Infection. J. Biomed. Sci. 2020, 27, 4. [Google Scholar] [CrossRef]
- Castrillón-Betancur, J.C.; Urcuqui-Inchima, S. Overexpression of MiR-484 and MiR-744 in Vero Cells Alters Dengue Virus Replication. Memórias Inst. Oswaldo Cruz 2017, 112, 281–291. [Google Scholar] [CrossRef]
- Castillo, J.A.; Castrillón, J.C.; Diosa-Toro, M.; Betancur, J.G.; St Laurent, G.; Smit, J.M.; Urcuqui-Inchima, S. Complex Interaction between Dengue Virus Replication and Expression of MiRNA-133a. BMC Infect. Dis. 2016, 16, 29. [Google Scholar] [CrossRef]
- Kambara, H.; Niazi, F.; Kostadinova, L.; Moonka, D.K.; Siegel, C.T.; Post, A.B.; Carnero, E.; Barriocanal, M.; Fortes, P.; Anthony, D.D.; et al. Negative Regulation of the Interferon Response by an Interferon-Induced Long Non-Coding RNA. Nucleic Acids Res. 2014, 42, 10668–10681. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Xu, C.; Zhao, P.; Qi, Z. Long Non-Coding RNA GAS5 Inhibited Hepatitis C Virus Replication by Binding Viral NS3 Protein. Virology 2016, 492, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lowey, B.; Sodroski, C.; Krishnamurthy, S.; Alao, H.; Cha, H.; Chiu, S.; El-Diwany, R.; Ghany, M.G.; Liang, T.J. Cellular MicroRNA Networks Regulate Host Dependency of Hepatitis C Virus Infection. Nat. Commun. 2017, 8, 1789. [Google Scholar] [CrossRef]
- Girardi, E.; López, P.; Pfeffer, S. On the Importance of Host MicroRNAs during Viral Infection. Front. Genet. 2018, 9, 439. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Hu, J.; Cao, P.; Yan, Q.; Zhang, S.; Dang, W.; Lu, J. Long Noncoding RNAs Involvement in Epstein-Barr Virus Infection and Tumorigenesis. Virol. J. 2020, 17, 51. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.C.; Yu, S.L.; Chen, J.J.W.; Chang, S.Y.; Yan, B.S.; Hong, Q.S.; Singh, S.; Kao, C.L.; Chen, H.Y.; Su, K.Y.; et al. Enterovirus-Induced MiR-141 Contributes to Shutoff of Host Protein Translation by Targeting the Translation Initiation Factor EIF4E. Cell Host Microbe 2011, 9, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Ingle, H.; Kumar, S.; Raut, A.A.; Mishra, A.; Kulkarni, D.D.; Kameyama, T.; Takaoka, A.; Akira, S.; Kumar, H. The MicroRNA MiR-485 Targets Host and Influenza Virus Transcripts to Regulate Antiviral Immunity and Restrict Viral Replication. Sci. Signal. 2015, 8, ra126. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, Q.; Guo, Y.; Liu, S.; Song, R.; Gao, X.; Dai, L.; Li, B.; Zhang, D.; Cheng, J. Microarray Analysis of MicroRNA Expression in Peripheral Blood Mononuclear Cells of Critically Ill Patients with Influenza A (H1N1). BMC Infect. Dis. 2013, 13, 257. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Liu, X.; Su, Z.Z.; Hsu, A.C.Y.; Foster, P.S.; Yang, M. Potential Role of MicroRNAs in the Regulation of Antiviral Responses to Influenza Infection. Front. Immunol. 2018, 9, 1541. [Google Scholar] [CrossRef]
- Peng, F.; He, J.; Loo, J.F.C.; Yao, J.; Shi, L.; Liu, C.; Zhao, C.; Xie, W.; Shao, Y.; Kong, S.K.; et al. Identification of MicroRNAs in Throat Swab as the Biomarkers for Diagnosis of Influenza. Int. J. Med. Sci. 2016, 13, 77–84. [Google Scholar] [CrossRef]
- Makkoch, J.; Poomipak, W.; Saengchoowong, S.; Khongnomnan, K.; Praianantathavorn, K.; Jinato, T.; Poovorawan, Y.; Payungporn, S. Human MicroRNAs Profiling in Response to Influenza A Viruses (Subtypes PH1N1, H3N2, and H5N1). Exp. Biol. Med. 2016, 241, 409–420. [Google Scholar] [CrossRef]
- Qu, D.; Sun, W.W.; Li, L.; Ma, L.; Sun, L.; Jin, X.; Li, T.; Hou, W.; Wang, J.H. Long Noncoding RNA MALAT1 Releases Epigenetic Silencing of HIV-1 Replication by Displacing the Polycomb Repressive Complex 2 from Binding to the LTR Promoter. Nucleic Acids Res. 2019, 47, 3013–3027. [Google Scholar] [CrossRef]
- Ray, R.M.; Morris, K.V. Long Non-Coding RNAs Mechanisms of Action in HIV-1 Modulation and the Identification of Novel Therapeutic Targets. Non-Coding RNA 2020, 6, 12. [Google Scholar] [CrossRef]
- Chen, L.; Chen, L.; Zuo, L.; Gao, Z.; Shi, Y.; Yuan, P.; Han, S.; Yin, J.; Peng, B.; He, X.; et al. Short Communication: Long Noncoding RNA Gas5 Inhibits HIV-1 Replication through Interaction with MiR-873. AIDS Res. Hum. Retrovir. 2018, 34, 544–549. [Google Scholar] [CrossRef]
- Imam, H.; Bano, A.S.; Patel, P.; Holla, P.; Jameel, S. The LncRNA NRON Modulates HIV-1 Replication in a NFAT-Dependent Manner and Is Differentially Regulated by Early and Late Viral Proteins. Sci. Rep. 2015, 5, 8639. [Google Scholar] [CrossRef]
- Chao, T.C.; Zhang, Q.; Li, Z.; Tiwari, S.K.; Qin, Y.; Yau, E.; Sanchez, A.; Singh, G.; Chang, K.; Kaul, M.; et al. The Long Noncoding RNA HEAL Regulates HIV-1 Replication through Epigenetic Regulation of the HIV-1 Promoter. mBio 2019, 10, e020616-19. [Google Scholar] [CrossRef] [PubMed]
- Postler, T.S.; Pantry, S.N.; Desrosiers, R.C.; Ghosh, S. Identification and Characterization of a Long Non-Coding RNA up-Regulated during HIV-1 Infection. Virology 2017, 511, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Jeffers, L.K.; Duan, K.; Ellies, L.G.; Seaman, W.T.; Burger-Calderon, R.A.; Diatchenko, L.B.; Webster-Cyriaque, J. Correlation of Transcription of MALAT-1, a Novel Noncoding RNA, with Deregulated Expression of Tumor Suppressor P53 in Small DNA Tumor Virus Models. J. Cancer Ther. 2013, 4, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Vrati, S. The Malat1 Long Non-Coding RNA Is Upregulated by Signalling through the PERK Axis of Unfolded Protein Response during Flavivirus Infection. Sci. Rep. 2015, 5, 17794. [Google Scholar] [CrossRef]
- Fang, H.; Shuang, D.; Yi, Z.; Sheng, H.; Liu, Y. Up-Regulated MicroRNA-155 Expression Is Associated with Poor Prognosis in Cervical Cancer Patients. Biomed. Pharmacother. 2016, 83, 64–69. [Google Scholar] [CrossRef]
- Santos, J.M.O.; da Silva, S.P.; Costa, N.R.; Gil da Costa, R.M.; Medeiros, R. The Role of MicroRNAs in the Metastatic Process of High-Risk HPV-Induced Cancers. Cancers 2018, 10, 493. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Kinoshita, T.; Nohata, N.; Itesako, T.; Yoshino, H.; Enokida, H.; Nakagawa, M.; Shozu, M.; Seki, N. Tumor Suppressive MicroRNA-218 Inhibits Cancer Cell Migration and Invasion by Targeting Focal Adhesion Pathways in Cervical Squamous Cell Carcinoma. Int. J. Oncol. 2013, 42, 1523–1532. [Google Scholar] [CrossRef]
- Zhong, J.; Yuan, H.; Xu, X.; Kong, S. MicroRNA-195 Inhibits Cell Proliferation, Migration and Invasion by Targeting Defective in Cullin Neddylation 1 Domain Containing 1 in Cervical Cancer. Int. J. Mol. Med. 2018, 42, 779–788. [Google Scholar]
- Wang, F.; Li, Y.; Zhou, J.; Xu, J.; Peng, C.; Ye, F.; Shen, Y.; Lu, W.; Wan, X.; Xie, X. MiR-375 Is down-Regulated in Squamous Cervical Cancer and Inhibits Cell Migration and Invasion via Targeting Transcription Factor SP1. Am. J. Pathol. 2011, 179, 2580–2588. [Google Scholar] [CrossRef]
- Au Yeung, C.L.; Tsang, T.Y.; Yau, P.L.; Kwok, T.T. Human Papillomavirus Type 16 E6 Induces Cervical Cancer Cell Migration through the P53/MicroRNA-23b/Urokinase-Type Plasminogen Activator Pathway. Oncogene 2011, 30, 2401–2410. [Google Scholar] [CrossRef]
- Zhou, X.; Li, X.; Wu, M. MiRNAs Reshape Immunity and Inflammatory Responses in Bacterial Infection. Signal Transduct. Target. Ther. 2018, 3, 14. [Google Scholar] [CrossRef]
- Reed, M.B.; Domenech, P.; Manca, C.; Su, H.; Barczak, A.K.; Kreiswirth, B.N.; Kaplan, G. A Glycolipid of Hypervirulent Tuberculosis Strains That Inhibits the Innate Immune Response. Nature 2004, 431, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Zhu, C.; Yao, Y.; Wang, X.; Song, J.; Zhai, J. MicroRNA-155-Enhanced Autophagy in Human Gastric Epithelial Cell in Response to Helicobacter pylori. Saudi J. Gastroenterol. Assoc. 2016, 22, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Liu, Z.; Li, B.S.; Tang, B.; Li, W.; Guo, G.; Shi, Y.; Wang, F.; Wu, Y.; Tong, W.D.; et al. Induction of MicroRNA-155 during Helicobacter pylori Infection and Its Negative Regulatory Role in the Inflammatory Response. J. Infect. Dis. 2009, 200, 916–925. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef]
- Rau, C.S.; Wu, S.C.; Yang, J.C.S.; Lu, T.H.; Wu, Y.C.; Chen, Y.C.; Tzeng, S.L.; Wu, C.J.; Hsieh, C.H. Profiling the Circulating MiRNAs in Mice Exposed to Gram-Positive and Gram-Negative Bacteria by Illumina Small RNA Deep Sequencing. J. Biomed. Sci. 2015, 22, 1. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, M.; Geng, G.; Liu, B.; Huang, Z.; Luo, H.; Zhou, J.; Guo, X.; Cai, W.; Zhang, H. A Novel HIV-1-Encoded MicroRNA Enhances Its Viral Replication by Targeting the TATA Box Region. Retrovirology 2014, 11, 23. [Google Scholar] [CrossRef]
- Nukui, M.; Mori, Y.; Murphy, E.A. A Human Herpesvirus 6A-Encoded MicroRNA: Role in Viral Lytic Replication. J. Virol. 2015, 89, 2615–2627. [Google Scholar] [CrossRef]
- Kincaid, R.P.; Sullivan, C.S. Virus-Encoded MicroRNAs: An Overview and a Look to the Future. PLoS Pathog. 2012, 8, e1003018. [Google Scholar] [CrossRef]
- Jopling, C.L.; Yi, M.K.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Molecular Biology: Modulation of Hepatitis C Virus RNA Abundance by a Liver-Specific MicroRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, X.; Ding, S.W. Viral Suppressors of RNA-Based Viral Immunity: Host Targets. Cell Host Microbe 2010, 8, 12–15. [Google Scholar] [CrossRef] [PubMed]
- McCaskill, J.; Praihirunkit, P.; Sharp, P.M.; Buck, A.H. RNA-Mediated Degradation of MicroRNAs: A Widespread Viral Strategy? RNA Biol. 2015, 12, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Fani, M.; Zandi, M.; Ebrahimi, S.; Soltani, S.; Abbasi, S. The Role of MiRNAs in COVID-19 Disease. Future Virol. 2021, 16, 301–306. [Google Scholar] [CrossRef]
- Farr, R.; Cheng, A.; Kedzierska, K. Altered MicroRNA Expression in COVID-19 Patients Enables Identication of SARS-CoV-2 Infection. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Liu, W.; Ding, C. Roles of LncRNAs in Viral Infections. Front. Cell. Infect. Microbiol. 2017, 7, 205. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xu, J.; Wang, Y.; Cao, X. An Interferon-Independent LncRNA Promotes Viral Replication by Modulating Cellular Metabolism. Science 2017, 358, 1051–1055. [Google Scholar] [CrossRef]
- McAninch, D.; Roberts, C.T.; Bianco-Miotto, T. Mechanistic Insight into Long Noncoding RNAs and the Placenta. Int. J. Mol. Sci. 2017, 18, 1371. [Google Scholar] [CrossRef] [PubMed]
- Hum, C.; Loiselle, J.; Ahmed, N.; Shaw, T.A.; Toudic, C.; Pezacki, J.P. MicroRNA Mimics or Inhibitors as Antiviral Therapeutic Approaches Against COVID-19. Drugs 2021, 81, 517–531. [Google Scholar] [CrossRef]
- Morishita, A.; Oura, K.; Tadokoro, T.; Fujita, K.; Tani, J.; Masaki, T. MicroRNA Interference in Hepatic Host-Pathogen Interac tions. Int. J. Mol. Sci. 2021, 22, 3554. [Google Scholar] [CrossRef]
- Staedel, C.; Darfeuille, F. MicroRNAs and bacterial infection. Cell. Microbiol. 2013, 15, 1496–1507. [Google Scholar] [CrossRef]
- Ozata, D.M.; Gainetdinov, I.; Zoch, A.; O’Carroll, D.; Zamore, P.D. PIWI-interacting RNAs: Small RNAs with big functions. Nat. Rev. Genet. 2019, 20, 89–108. [Google Scholar] [CrossRef] [PubMed]
- Firmino, N.; Martinez, V.D.; Rowbotham, D.A.; Enfield, K.; Bennewith, K.L.; Lam, W.L. HPV status is associated with altered PIWI-interacting RNA expression pattern in head and neck cancer. Oral Oncol. 2016, 55, 43–48. [Google Scholar] [CrossRef]
- Zhang, X.; Liang, Z.; Zhang, Y.; Zhu, M.; Zhu, Y.; Li, S.; Zhao, W.; Hu, X.; Wang, J. Specific PIWI-interacting small noncoding RNA expression patterns in pulmonary tuberculosis patients. Epigenomics 2019, 11, 1779–1794. [Google Scholar] [CrossRef] [PubMed]
- de Araujo, L.S.; Ribeiro-Alves, M.; Leal-Calvo, T.; Leung, J.; Durán, V.; Samir, M.; Talbot, S.; Tallam, A.; Mello, F.; Geffers, R.; et al. Reprogramming of Small Noncoding RNA Populations in Peripheral Blood Reveals Host Biomarkers for Latent and Active Mycobacterium tuberculosis Infection. mBio 2019, 10, e01037-19. [Google Scholar] [CrossRef]
- Yan, L.; Chen, Y.G. Circular RNAs in Immune Response and Viral Infection. Trends Biochem. Sci. 2020, 45, 1022–1034. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Sun, H.; Mu, R.; Li, S.; Li, Y.; Yang, C.; Xu, M.; Duan, X.; Chen, L. The role of circular RNAs in viral infection and related diseases. Virus Res. 2021, 291, 198205. [Google Scholar] [CrossRef] [PubMed]
- Ojha, R.; Nandani, R.; Chatterjee, N.; Prajapati, V.K. Emerging Role of Circular RNAs as Potential Biomarkers for the Diagnosis of Human Diseases. Adv. Exp. Med. Biol. 2018, 1087, 141–157. [Google Scholar]
- Nunes, A.; Ribeiro, D.R.; Marques, M.; Santos, M.; Ribeiro, D.; Soares, A.R. Emerging Roles of tRNAs in RNA Virus Infections. Trends Biochem. Sci. 2020, 45, 794–805. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive Oxygen Species, Cellular Redox Systems, and Apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef]
- Forman, H.J.; Maiorino, M.; Ursini, F. Signaling Functions of Reactive Oxygen Species. Biochemistry 2010, 49, 835–842. [Google Scholar] [CrossRef]
- Kowaltowski, A.J.; de Souza-Pinto, N.C.; Castilho, R.F.; Vercesi, A.E. Mitochondria and Reactive Oxygen Species. Free Radic. Biol. Med. 2009, 47, 333–343. [Google Scholar] [CrossRef]
- Spooner, R.; Yilmaz, Ö. The Role of Reactive-Oxygen-Species in Microbial Persistence and Inflammation. Int. J. Mol. Sci. 2011, 12, 334–352. [Google Scholar] [CrossRef]
- Geng, J.; Sun, X.; Wang, P.; Zhang, S.; Wang, X.; Wu, H.; Hong, L.; Xie, C.; Li, X.; Zhao, H.; et al. Kinases Mst1 and Mst2 Positively Regulate Phagocytic Induction of Reactive Oxygen Species and Bactericidal Activity. Nat. Immunol. 2015, 16, 1142–1152. [Google Scholar] [CrossRef]
- Dryden, M. Reactive Oxygen Therapy: A Novel Therapy in Soft Tissue Infection. Curr. Opin. Infect. Dis. 2017, 30, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Unno, Y.; Miyazaki, C.; Ubagai, T.; Ono, Y. Multidrug-Resistant Acinetobacter baumannii Resists Reactive Oxygen Species and Survives in Macrophages. Sci. Rep. 2019, 9, 17462. [Google Scholar] [CrossRef] [PubMed]
- Silwal, P.; Kim, J.K.; Kim, Y.J.; Jo, E.K. Mitochondrial Reactive Oxygen Species: Double-Edged Weapon in Host Defense and Pathological Inflammation During Infection. Front. Immunol. 2020, 11, 1649. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Grimm, M.J.; Robert Vethanayagam, R.; Almyroudis, N.G.; Lewandowski, D.; Rall, N.; Blackwell, T.S.; Segal, B.H. Role of NADPH Oxidase in Host Defense against Aspergillosis. Med. Mycol. 2011, 49, S144–S149. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Holland, S.M. Chronic Granulomatous Disease. Clin. Rev. Allergy Immunol. 2010, 38, 3–10. [Google Scholar] [CrossRef]
- Pizzolla, A.; Hultqvist, M.; Nilson, B.; Grimm, M.J.; Eneljung, T.; Jonsson, I.-M.; Verdrengh, M.; Kelkka, T.; Gjertsson, I.; Segal, B.H.; et al. Reactive Oxygen Species Produced by the NADPH Oxidase 2 Complex in Monocytes Protect Mice from Bacterial Infections. J. Immunol. 2012, 188, 5003–5011. [Google Scholar] [CrossRef]
- van den Berg, J.M.; van Koppen, E.; Åhlin, A.; Belohradsky, B.H.; Bernatowska, E.; Corbeel, L.; Espanñol, T.; Fischer, A.; Kurenko-Deptuch, M.; Mouy, R.; et al. Chronic Granulomatous Disease: The European Experience. PLoS ONE 2009, 4, e5234. [Google Scholar] [CrossRef] [PubMed]
- Handa, O.; Naito, Y.; Yoshikawa, T. Helicobacter pylori: A ROS-Inducing Bacterial Species in the Stomach. Inflamm. Res. 2010, 59, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Sander, W.J.; Fourie, C.; Sabiu, S.; O’Neill, F.H.; Pohl, C.H.; O’Neill, H.G. Reactive Oxygen Species as Potential Antiviral Targets. Rev. Med. Virol. 2021. [Google Scholar] [CrossRef]
- Kirkpatrick, C.T.; Wang, Y.; Leiva Juarez, M.M.; Shivshankar, P.; Pantaleón García, J.; Plumer, A.K.; Kulkarni, V.V.; Ware, H.H.; Gulraiz, F.; Chavez Cavasos, M.A.; et al. Inducible Lung Epithelial Resistance Requires Multisource Reactive Oxygen Species Generation to Protect against Viral Infections. mBio 2018, 9, e00696-18. [Google Scholar] [CrossRef]
- Lee, C. Therapeutic Modulation of Virus-Induced Oxidative Stress via the Nrf2-Dependent Antioxidative Pathway. Oxid. Med. Cell. Longev. 2018, 2018, 6208067. [Google Scholar] [CrossRef] [PubMed]
- Qiang, W.; Kuang, X.; Liu, J.; Liu, N.; Scofield, V.L.; Reid, A.J.; Jiang, Y.; Stoica, G.; Lynn, W.S.; Wong, P.K.Y. Astrocytes Survive Chronic Infection and Cytopathic Effects of the Ts1 Mutant of the Retrovirus Moloney Murine Leukemia Virus by Upregulation of Antioxidant Defenses. J. Virol. 2006, 80, 3273–3284. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.V.B.; Agudelo, M.; Atluri, V.S.R.; Nair, M.P. Inhibition of Nuclear Factor Erythroid 2-Related Factor 2 Exacerbates HIV-1 Gp120-Induced Oxidative and Inflammatory Response: Role in HIV Associated Neurocognitive Disorder. Neurochem. Res. 2012, 37, 1697–1706. [Google Scholar] [CrossRef]
- Reddy, P.V.B.; Gandhi, N.; Samikkannu, T.; Saiyed, Z.; Agudelo, M.; Yndart, A.; Khatavkar, P.; Nair, M.P.N. HIV-1 Gp120 Induces Antioxidant Response Element-Mediated Expression in Primary Astrocytes: Role in HIV Associated Neurocognitive Disorder. Neurochem. Int. 2012, 61, 807–814. [Google Scholar] [CrossRef]
- Mastrantonio, R.; Cervelli, M.; Pietropaoli, S.; Mariottini, P.; Colasanti, M.; Persichini, T. HIV-Tat Induces the Nrf2/ARE Pathway through NMDA Receptor-Elicited Spermine Oxidase Activation in Human Neuroblastoma Cells. PLoS ONE 2016, 11, e0149802. [Google Scholar] [CrossRef]
- Zhang, H.S.; Wu, T.C.; Sang, W.W.; Ruan, Z. EGCG Inhibits Tat-Induced LTR Transactivation: Role of Nrf2, AKT, AMPK Signaling Pathway. Life Sci. 2012, 90, 747–754. [Google Scholar] [CrossRef]
- Zhang, H.S.; Li, H.Y.; Zhou, Y.; Wu, M.R.; Zhou, H. Sen. Nrf2 Is Involved in Inhibiting Tat-Induced HIV-1 Long Terminal Repeat Transactivation. Free Radic. Biol. Med. 2009, 47, 261–268. [Google Scholar] [CrossRef]
- Fan, X.; Staitieh, B.S.; Jensen, J.S.; Mould, K.J.; Greenberg, J.A.; Joshi, P.C.; Koval, M.; Guidot, D.M. Activating the Nrf2-Mediated Antioxidant Response Element Restores Barrier Function in the Alveolar Epithelium of HIV-1 Transgenic Rats. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, L267–L277. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Ichimura, Y.; Taguchi, K.; Suzuki, T.; Mizushima, T.; Takagi, K.; Hirose, Y.; Nagahashi, M.; Iso, T.; Fukutomi, T.; et al. P62/Sqstm1 Promotes Malignancy of HCV-Positive Hepatocellular Carcinoma through Nrf2-Dependent Metabolic Reprogramming. Nat. Commun. 2016, 7, 12030. [Google Scholar] [CrossRef]
- Kosmider, B.; Messier, E.M.; Janssen, W.J.; Nahreini, P.; Wang, J.; Hartshorn, K.L.; Mason, R.J. Nrf2 Protects Human Alveolar Epithelial Cells against Injury Induced by Influenza A Virus. Respir. Res. 2012, 13, 43. [Google Scholar] [CrossRef]
- Hosakote, Y.M.; Jantzi, P.D.; Esham, D.L.; Spratt, H.; Kurosky, A.; Casola, A.; Garofalo, R.P. Viral-Mediated Inhibition of Antioxidant Enzymes Contributes to the Pathogenesis of Severe Respiratory Syncytial Virus Bronchiolitis. Am. J. Respir. Crit. Care Med. 2011, 183, 1550–1560. [Google Scholar] [CrossRef]
- Schaedler, S.; Krause, J.; Himmelsbach, K.; Carvajal-Yepes, M.; Lieder, F.; Klingel, K.; Nassal, M.; Weiss, T.S.; Werner, S.; Hildt, E. Hepatitis B Virus Induces Expression of Antioxidant Response Element-Regulated Genes by Activation of Nrf2. J. Biol. Chem. 2010, 285, 41074–41086. [Google Scholar] [CrossRef] [PubMed]
- Schachtele, S.J.; Hu, S.; Lokensgard, J.R. Modulation of Experimental Herpes Encephalitis-Associated Neurotoxicity through Sulforaphane Treatment. PLoS ONE 2012, 7, e36216. [Google Scholar] [CrossRef]
- Tilton, C.; Clippinger, A.J.; Maguire, T.; Alwine, J.C. Human Cytomegalovirus Induces Multiple Means To Combat Reactive Oxygen Species. J. Virol. 2011, 85, 12585–12593. [Google Scholar] [CrossRef]
- Gjyshi, O.; Bottero, V.; Veettil, M.V.; Dutta, S.; Singh, V.V.; Chikoti, L.; Chandran, B. Kaposi’s Sarcoma-Associated Herpesvirus Induces Nrf2 during De Novo Infection of Endothelial Cells to Create a Microenvironment Conducive to Infection. PLoS Pathog. 2014, 10, e1004460. [Google Scholar] [CrossRef] [PubMed]
- Olagnier, D.; Peri, S.; Steel, C.; van Montfoort, N.; Chiang, C.; Beljanski, V.; Slifker, M.; He, Z.; Nichols, C.N.; Lin, R.; et al. Cellular Oxidative Stress Response Controls the Antiviral and Apoptotic Programs in Dengue Virus-Infected Dendritic Cells. PLoS Pathog. 2014, 10, e1004566. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Song, C.H. Effect of Reactive Oxygen Species on the Endoplasmic Reticulum and Mitochondria during Intracellular Pathogen Infection of Mammalian Cells. Antioxidants 2021, 10, 872. [Google Scholar] [CrossRef] [PubMed]
- Herb, M.; Schramm, M. Functions of ROS in Macrophages and Antimicrobial Immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Fu, P.; Ramchandran, R.; Sudhadevi, T.; Kumar, P.; Krishnan, Y.; Liu, Y.; Zhao, Y.; Parinandi, N.L.; Harijith, A.; Sadoshima, J.; et al. NOX4 Mediates Pseudomonas aeruginosa-Induced Nuclear Reactive Oxygen Species Generation and Chromatin Remodeling in Lung Epithelium. Antioxidants 2021, 10, 477. [Google Scholar] [CrossRef] [PubMed]
- Warnatsch, A.; Tsourouktsoglou, T.D.; Branzk, N.; Wang, Q.; Reincke, S.; Herbst, S.; Gutierrez, M.; Papayannopoulos, V. Reactive Oxygen Species Localization Programs Inflammation to Clear Microbes of Different Size. Immunity 2017, 46, 421–432. [Google Scholar] [CrossRef]
- Garaude, J.; Acín-Pérez, R.; Martínez-Cano, S.; Enamorado, M.; Ugolini, M.; Nistal-Villán, E.; Hervás-Stubbs, S.; Pelegrín, P.; Sander, L.E.; Enríquez, J.A.; et al. Mitochondrial respiratory-chain adaptations in macrophages contribute to antibacterial host defense. Nat. Immunol. 2016, 17, 1037–1045. [Google Scholar] [CrossRef]
- Jadhav, V.S.; Krause, K.H.; Singh, S.K. HIV-1 Tat C Modulates NOX2 and NOX4 Expressions through MiR-17 in a Human Microglial Cell Line. J. Neurochem. 2014, 131, 803–815. [Google Scholar] [CrossRef]
- Zhang, J.; Thakuri, B.K.C.; Zhao, J.; Nguyen, L.N.; Nguyen, L.N.T.; Cao, D.; Dang, X.; Khanal, S.; Schank, M.; Lu, Z.; et al. Long Noncoding RNA HOTAIRM1 Promotes Myeloid-Derived Suppressor Cell Expansion and Suppressive Functions through up-Regulating HOXA1 Expression during Latent HIV Infection. AIDS 2020, 34, 2211–2221. [Google Scholar] [CrossRef]
- Wang, J.; Wu, M.; Wen, J.; Yang, K.; Li, M.; Zhan, X.; Feng, L.; Li, M.; Huang, X. MicroRNA-155 Induction by Mycobacterium bovis BCG Enhances ROS Production through Targeting SHIP1. Mol. Immunol. 2014, 62, 29–36. [Google Scholar] [CrossRef]
- Singh, A.K.; Ghosh, M.; Kumar, V.; Aggarwal, S.; Patil, S.A. Interplay between MiRNAs and Mycobacterium tuberculosis: Diagnostic and Therapeutic Implications. Drug Discov. Today 2021, 26, 1245–1255. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, C.; Shan, S.; Liu, X.; Jiang, Z.; Ren, T. TLR4/ROS/MiRNA-21 Pathway Underlies Lipopolysaccharide Instructed Primary Tumor Outgrowth in Lung Cancer Patients. Oncotarget 2016, 7, 42172–42182. [Google Scholar] [CrossRef]
- Liang, S.; Song, Z.; Wu, Y.; Gao, Y.; Gao, M.; Liu, F.; Wang, F.; Zhang, Y. MicroRNA-27b Modulates Inflammatory Response and Apoptosis during Mycobacterium tuberculosis Infection. J. Immunol. 2018, 200, 3506–3518. [Google Scholar] [CrossRef]
- Huang, T.; Pu, Q.; Zhou, C.; Lin, P.; Gao, P.; Zhang, X.; Chu, Y.; Yue, B.; Wu, M. MicroRNA-302/367 Cluster Impacts Host Antimicrobial Defense via Regulation of Mitophagic Response Against Pseudomonas aeruginosa Infection. Front. Immunol. 2020, 11, 1. [Google Scholar] [CrossRef]
- Strickertsson, J.A.B.; Rasmussen, L.J.; Friis-Hansen, L. Enterococcus Faecalis Infection and Reactive Oxygen Species Down-Regulates the MiR-17-92 Cluster in Gastric Adenocarcinoma Cell Culture. Genes 2014, 5, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chan, E.Y.; Li, J.; Ni, C.; Peng, X.; Rosenzweig, E.; Tumpey, T.M.; Katze, M.G. MicroRNA Expression and Virulence in Pandemic Influenza Virus-Infected Mice. J. Virol. 2010, 84, 3023–3032. [Google Scholar] [CrossRef]
- Kim, S.M.; Hur, D.Y.; Hong, S.W.; Kim, J.H. EBV-Encoded EBNA1 Regulates Cell Viability by Modulating MiR34a-NOX2-ROS Signaling in Gastric Cancer Cells. Biochem. Biophys. Res. Commun. 2017, 494, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Q.; Gu, X.Y.; Hu, J.X.; Ping, Y.; Li, H.; Yan, J.Y.; Li, J.; Sun, R.; Yu, Z.J.; Zhang, Y. Hepatitis C Virus Core Protein Impairs Metabolic Disorder of Liver Cell via HOTAIR-Sirt1 Signalling. Biosci. Rep. 2016, 36, e00336. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yu, J.; Ma, C.; Gong, Z.; Wu, X.; Deng, G. Impact of Knockdown LincRNA-Cox2 on Apoptosis of Macrophage Infected with Bacillus Calmette-Guérin. Mol. Immunol. 2021, 130, 85–95. [Google Scholar] [CrossRef]
- Zuo, K.; Kong, L.; Xue, D.; Yang, Y.; Xie, L. The Expression and Role of LncRNA AX800134 in Hepatitis B Virus-Related Hepatocellular Carcinoma. Virus Genes 2018, 54, 475–483. [Google Scholar] [CrossRef]
- Sahin, U.; Karikó, K.; Türeci, Ö. MRNA-Based Therapeutics-Developing a New Class of Drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Zhong, Z.; Mc Cafferty, S.; Combes, F.; Huysmans, H.; De Temmerman, J.; Gitsels, A.; Vanrompay, D.; Portela Catani, J.; Sanders, N.N. MRNA Therapeutics Deliver a Hopeful Message. Nano Today 2018, 23, 16–39. [Google Scholar] [CrossRef]
- Zhang, C.; Maruggi, G.; Shan, H.; Li, J. Advances in MRNA Vaccines for Infectious Diseases. Front. Immunol. 2019, 10, 594. [Google Scholar] [CrossRef] [PubMed]
- Nadhan, R.; Patra, D.; Krishnan, N.; Rajan, A.; Gopala, S.; Ravi, D.; Srinivas, P. Perspectives on mechanistic implications of ROS inducers for targeting viral infections. Eur. J. Pharmacol. 2021, 890, 173621. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| ncRNA | Regulation | Target/Function | Pathogen | References |
|---|---|---|---|---|
| miR-1289 | ↑ | Gastric acidity | Helicobacter pylori | [33] |
| miR-30c miR-30e | ↑ | SUMOylation | Salmonella | [53] |
| miR-132 miR-26a | ↑ | Macrophage response to IFN-γ | Mycobacterium tuberculosis | [54] |
| miR-125b miR-99b miR-155 | ↑ ↑ ↓ | Production of TNF-α | Mycobacterium tuberculosis | [55,56] |
| Let-7f | ↓ | NF-κB pathway inhibition | Mycobacterium tuberculosis | [57] |
| miR-29 | ↓ | IFN-γ production | BCG Listeria monocytogenes | [35] |
| miR-155 | ↑ | Activation of the pro-survival Akt pathway | Listeria monocytogenes | [34] |
| ↓ | Reduction of humoral immune responses | Citrobacter rodentium | [58] | |
| ↑ | RIP1/3-related necroptosis and PARP-1-mediated necrosis | Salmonella | [59] | |
| ↑ | Down-regulation of SHIP | Francisella tularensis | [60] | |
| ↑ | Decreased the production of IL-8 and GRO-α | Helicobacter pylori | [33] | |
| miR-146a | ↑ | Suppression of nitric oxide production | Mycobacterium tuberculosis | [61] |
| miR-128 | ↑ | p53 knockdown | Salmonella | [62] |
| lncRNA-CD244 | ↑ | Inhibition of TNF-α and INF-γ expression | Mycobacterium tuberculosis | [36] |
| NeST | ↑ | IFN-γ transcription | Salmonella enterica | [63] |
| NEAT1 | ↑ | Decrease the expression of IL-6 | Mycobacterium tuberculosis | [37] |
| ↑ | Export of Rev-dependent instability element (INS) | Human immunodeficiency virus | [64] | |
| ↑ | IL-8 secretion | Influenza virus | [65] | |
| ↑ | Regulation by IRF1, IRF4, STAT1 and STAT3 | SARS-CoV-2 virus | [66] | |
| miR-618 | ↑ | Dysregulation of immune function | SARS-CoV-2 virus | [67] |
| lncRHOXF1 | ↑ | siRNA-mediated disruption | Sendai virus | [38] |
| miR-24-1-5p miR-512-5p miR-4640-3p | ↑ | Biomarkers for mild dengue forms | Dengue virus | [68] |
| miR-383 | ↑ | Biomarker for mild dengue form | ||
| hsa-miR-21-5p hsa-miR-146a-5p hsa-miR-590-5p hsa-miR-188-5p hsa-miR-152-3p | ↑ | non-invasive molecular markers for detecting DENV infection | ||
| miR-548g-3p | ↑ | Suppress DENV multiplication | [69] | |
| miR-133a miR-484 miR-744 | ↑ | Downregulate DENV replication | [70] | |
| miR-122 | ↑ | Stabilizes the HCV genome | Hepatitis C virus | [68] |
| lncRNA-CMPK2 | ↑ | Interferon response modulation | [71] | |
| GAS5 | ↑ | Decoy of HCV NS3 protein | [72] | |
| miR-25 miR-130a/b let-7a | ↓ | HCV reduces their levels offsetting their antiviral capacity | [73,74] | |
| BART BHLF1 | ↑ | Tumorigenesis | Epstein–Barr virus | [75] |
| miR-141 | ↑ | Pro-viral effect | Enterovirus 71 | [76] |
| miR-485-5p | ↑ | Suppression of the antiviral response and enhanced viral replication | Newcastle disease virus | [77] |
| miR-148 | ↑ | Valuable biomarkers for severe influenza virus infections | Influenza virus | [78] |
|
miR-31 miR-29a | ↓ | |||
| miR-34c-3p | ↑ | Biomarker | [79,80] | |
|
miR-29a-3p miR-30c-5p miR-181a-5p | ↓ | Biomarkers | [79,80] | |
|
miR-323 miR-491 miR-654 miR-146a | ↑ | Inhibition of influenza virus replication | [81] | |
| MALA1 LINC01426 (uc002yug. 2) | ↑ | Activate HIV-1 replication and reactivates HIV-1 from latency | Human immunodeficiency virus | [82,83] |
|
7SKRNA NEAT1 NRON | ↑ | Repress HIV-1 infection | ||
| GAS5 | ↓ | Inhibits HIV-1 replication | [83,84,85] | |
| HEAL | ↑ |
Regulates HIV-1 transcription | [83,86] | |
| LINC00173 | ↓ | Dysregulates the immune response | [83,87] | |
| MALAT-1 | ↑ | Cervical cancer cell growth | Human papilloma virus | [88] |
| ↑ |
Induction of unfolded protein response | Flaviviruses | [89] | |
| miR-155 | ↑ |
Biomarker for poor prognosis | Human papilloma virus | [90,91] |
| miR-218 | ↓ | Inhibition of cancer cell migration and invasion | [90,91] | |
| miR-195 | ↓ | Biomarker for advanced clinical stages | [91,92,93] | |
| miR-375 | ↓ |
Biomarker for poor prognosis | [91,94] | |
|
miR-34a miR-23b | ↓ |
Increased migration of cervical cancer cell lines | [91,95] |
| ncRNA | Target/Function | Pathogen Regulation | References | ||
|---|---|---|---|---|---|
| Viral | Bacterial | Viral | Bacterial | ||
| miR-146a | -Inhibition of influenza virus replication; -Unknown | Suppression of nitric oxide production | ↑ Influenza virus ↑↓ Hepatitis C virus | ↑Mycobacterium tuberculosis | [61,81] |
| miR-155 | Biomarker for poor prognosis, IFN-related molecule | -Production of TNF-α -Activation of the pro-survival Akt pathway -Reduction of humoral immune responses -RIP1/3-related necroptosis and PARP-1-mediated necrosis | ↑ Hepatitis C virus | ↓ Mycobacterium tuberculosis ↑Listeria monocytogenes ↓Citrobacter rodentium; ↑Salmonella | [55,61,90] |
| miR-16 | Increased production of ROS | Unknown | ↓SARS-CoV-2 | ↓C. trachomatis | [113,114] |
| miR-30c | Biomarker | SUMOylation | ↓ Influenza virus | ↑Salmonella | [53,79] |
| NEAT1 | Repress HIV-1 infection, export of Rev-dependent instability element (INS) | Decrease the expression of IL-6 | ↑ Human immunodeficiency virus | ↑Mycobacterium tuberculosis | [37,64] |
| miR-133a | Downregulate DENV replication | Biomarker-circulating miRNA | ↑ Dengue virus | Gram-positive bacterial (mir-133a-1-3p, mir-133a-2-3p) | [70,101] |
| let-7 | -HCV reduces their levels offsetting their antiviral capacity; -TLR4 and STAT3 signaling | -Induced acute inflammation characterized by neutrophil infiltration | ↓ Hepatitis C virus (let-7a) ↓ Hepatitis B virus (let-7) | ↓Helicobacter pylori | [73,74,114,115] |
| miR-125b | TLR2/MyD88 signaling | Production of TNF-α | ↓ Hepatitis C Virus | ↑Mycobacterium tuberculosis | [55,56,114] |
| miR-29 | Biomarker for severe influenza virus infections | -IFN-γ-production; -Unknown | ↓ Influenza virus (miR-29a) | ↓BCG Listeria monocytogenes ↓C. trachomatis | [35,78,114] |
| miR-372, miR-373 | NFIB-dependent signaling | Cell cycle and apoptosis | ↑ Hepatitis B Virus | ↓Helicobacter | [49,114] |
| mir-122 | Stabilizes the HCV genome | Unknown | ↑ Hepatitis C Virus | ↓Enterobacteriaceae spp. | [68,114] |
| miR-223 | Antiviral–target Indirect: STMN1 mRNA | Unknown | DENV-2 (strain TR1751) | ↑Enterobacteriaceae spp. | [68,114] |
| miR-125b | TLR2/MyD88 signaling/ Immune response | Directly targets mRNA of TNF | ↓ Hepatitis C Virus | ↑Mycobacteria | [49,114] |
| miR-21 | -STAT3 signaling; -Non-invasive molecular markers for detecting DENV infection. | Immune response | ↑ Hepatitis B virus ↑ Dengue virus hsa-miR-21-5p | ↑Salmonella | [49,68,114] |
| Virus | Disease | ROS | Nfr2 | Reference |
|---|---|---|---|---|
| MoMuLV ts1 | Neurodegenerative disease | H2O2↓ | ↑ | [140,141] |
| HIV | Neurodegenerative disease | ↑ | ↑ | [140,142,143] |
| Neurocognitive disorder | H2O2↑ | ↑ | [140,144] | |
| AIDS | ↑ | ↑ | [140,144,145,146] | |
| Respiratory disorder | ↑ | ↓ | [140,147] | |
| HCV | HCC | ND | ↑ | [140,148] |
| Influenza | Respiratory disease | ↑ | Nuclear localization ↑ | [140,149] |
| RSV | Respiratory disease | ↑ | ↓ | [140,150] |
| HBV | Viral hepatitis | ↓ | ↑ | [140] |
| HSV | Encephalitis neurotoxicity | ↑ | ↑ | [140,151,152] |
| HCMV | Congenital abnormalities | ↑ | ↑ | [140,153] |
| KSHV | Sarcoma | ↑ | Nuclear localization ↑ | [140,154] |
| DENV | Fever | ↑ | ↑ | [140,155] |
| ROS Generator | Effect of ↑ ROS | Pathogen Trigger | Reference | ||
|---|---|---|---|---|---|
| Virus | Bacteria | Virus | Bacteria | ||
| Mitochondrial ROS | Damage the mtDNA, membrane lipid permeability, release of cytochrome C and apoptosis | -Bactericidal; -Stimulation of TLR9 by CpG-containing DNA and subsequent ROS production | Hepatitis C virus | Salmonella typhimurium Staphylococcus aureus | [156] |
| ER stress |
Boosts proinflammatory cytokine production via MAPK-killing | both | [156] | ||
| NADPH oxidase (NOX) | Induce activation of Capsase-3 and apoptosis | Promote oxidative and nonoxidative mechanisms of microbe elimination | both | [39] | |
| NOX2-derived ROS in macrophages | Produce ROS in the phagosomal membrane, resulting in elimination of the pathogen | Influenza A virus | Escherichia coli L. mono- cytogenes | [156,157] | |
| NOX4 | Induction of superoxide and H2O2 in hepatocytes | Induction of superoxide and H2O2 in lung cell and nuclear ROS generation => histone acetylation and chromatin remodeling | HCV infection | P. aeruginosa | [156,158] |
| NF-κB pathway | Inducing ROS-upregulation of antiviral genes in lymphocytes | Adjust inflammation through neutrophils that modulating inflammation | Japanese encephalitis virus (JEV) | C. albicans | [156,159] |
| Activation of NLRP3 inflammasome | Antiviral/antibacterial activity | RNA viruses DNA viruses | bacterial RNA | [156,160] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milhem, Z.; Chiroi, P.; Nutu, A.; Ilea, M.; Lupse, M.; Zanoaga, O.; Berindan-Neagoe, I. Non-Coding RNAs and Reactive Oxygen Species–Symmetric Players of the Pathogenesis Associated with Bacterial and Viral Infections. Symmetry 2021, 13, 1307. https://doi.org/10.3390/sym13071307
Milhem Z, Chiroi P, Nutu A, Ilea M, Lupse M, Zanoaga O, Berindan-Neagoe I. Non-Coding RNAs and Reactive Oxygen Species–Symmetric Players of the Pathogenesis Associated with Bacterial and Viral Infections. Symmetry. 2021; 13(7):1307. https://doi.org/10.3390/sym13071307
Chicago/Turabian StyleMilhem, Zaki, Paul Chiroi, Andreea Nutu, Maximilian Ilea, Mihaela Lupse, Oana Zanoaga, and Ioana Berindan-Neagoe. 2021. "Non-Coding RNAs and Reactive Oxygen Species–Symmetric Players of the Pathogenesis Associated with Bacterial and Viral Infections" Symmetry 13, no. 7: 1307. https://doi.org/10.3390/sym13071307
APA StyleMilhem, Z., Chiroi, P., Nutu, A., Ilea, M., Lupse, M., Zanoaga, O., & Berindan-Neagoe, I. (2021). Non-Coding RNAs and Reactive Oxygen Species–Symmetric Players of the Pathogenesis Associated with Bacterial and Viral Infections. Symmetry, 13(7), 1307. https://doi.org/10.3390/sym13071307