
Recent Advances in Handedness Genetics


Abstract
:1. Is Handedness a Genetic Trait?
2. How to Measure Handedness
3. There Is No Handedness Gene
4. Resequencing the Genome
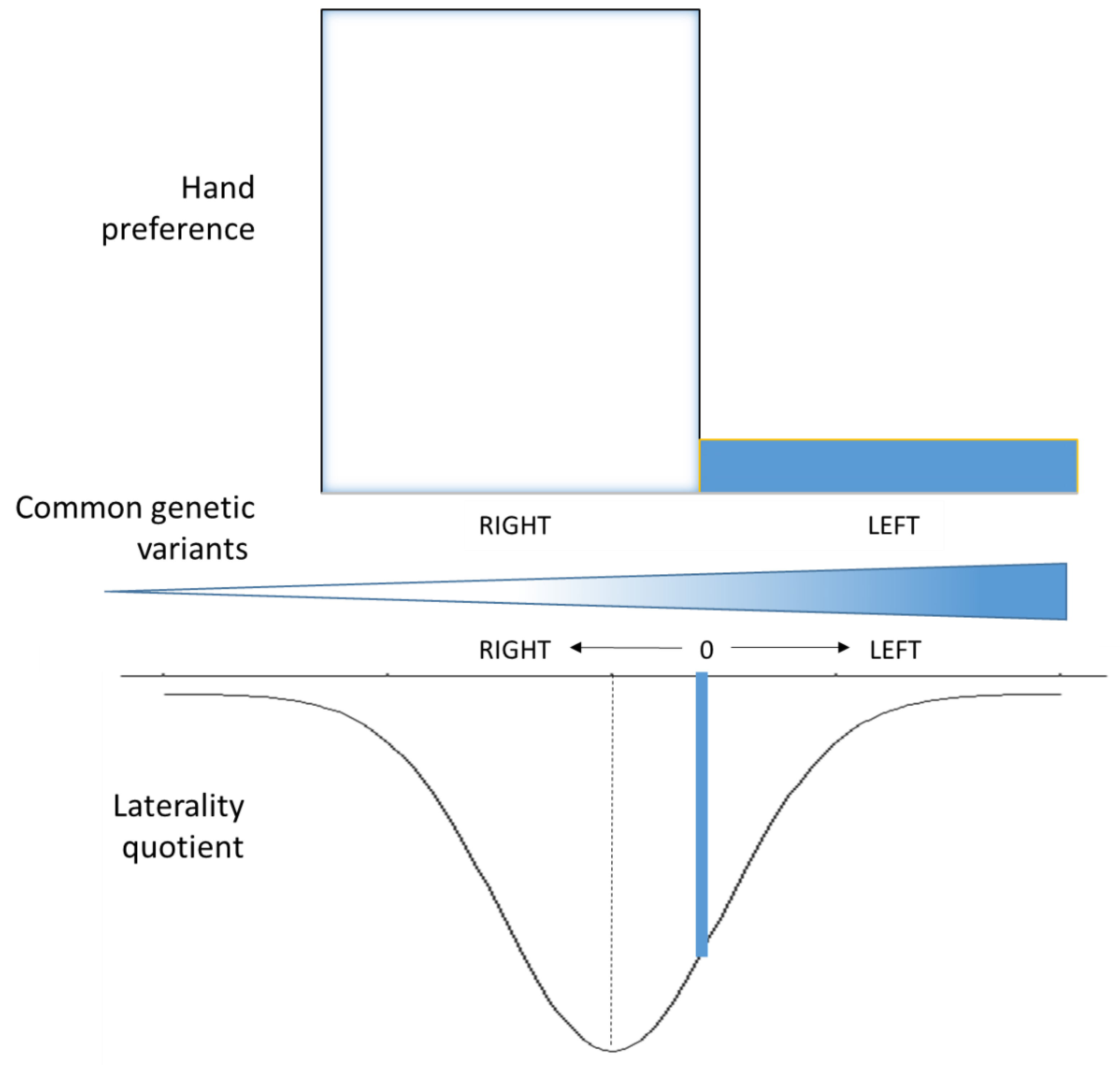
5. Handedness GWAS
6. Genetics, Handedness and Brain Asymmetries

{kind=link}
| Reference | N Participants | Cohorts | Handedness Phenotype | N Associated Genes |
|---|---|---|---|---|
| Eriksson et al. 2010 [60] | 9126 | 23andMe | Handedness questionnaire | none |
| Scerri et al. 2011 [61] | 744 | Dyslexia cohorts and ALSPAC | LQ from pegboard task | 1 |
| Brandler et al. 2013 [45] | 728 + 2666 | Dyslexia cohorts and ALSPAC | LQ from pegboard task | 1 |
| Wiberg et al. 2019 [44] | ~400,000 | UK Biobank | Hand preference | 4 |
| De Kovel et al. 2019 [56] | 331,037 | UK Biobank | Hand preference | 3 |
| Cuellar–Partida et al. 2021 [22] | 1,766,671 | UK Biobank, 23andMe, International Handedness Consortium | Hand preference | 48 |
7. Conclusions
Funding
Conflicts of Interest
References
- De Kovel, C.G.F.; Carrión-Castillo, A.; Francks, C. A large-scale population study of early life factors influencing left-handedness. Sci. Rep. 2019, 9, 584. [Google Scholar] [CrossRef] [Green Version]
- Papadatou-Pastou, M.; Ntolka, E.; Schmitz, J.; Martin, M.; Munafò, M.R.; Ocklenburg, S.; Paracchini, S. Human handedness: A meta-analysis. Psychol. Bull. 2020, 146, 481–524. [Google Scholar] [CrossRef] [PubMed]
- HI, K. Why are there (almost) no left-handers in China? Endeavour 2013, 37, 71–81. [Google Scholar]
- Zheng, M.; McBride, C.; Ho, C.S.-H.; Chan, J.K.-C.; Choy, K.W.; Paracchini, S. Prevalence and heritability of handedness in a Hong Kong Chinese twin and singleton sample. BMC Psychol. 2020, 8, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghirlanda, S.; Vallortigara, G. The evolution of brain lateralization: A game-theoretical analysis of population structure. Proc. R. Soc. B Biol. Sci. 2004, 271, 853–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faurie, C.; Raymond, M. The fighting hypothesis as an evolutionary explanation for the handedness polymorphism in humans: Where are we? Ann. N. Y. Acad. Sci. 2013, 1288, 110–113. [Google Scholar] [CrossRef]
- Faurie, C.; Raymond, M.; Uomini, N. Origins, Development, and Persistence of Laterality in Humans. In Laterality in Sports; Academic Press: Cambridge, MA, USA, 2016; pp. 11–30. [Google Scholar] [CrossRef]
- Mazoyer, B.; Zago, L.; Jobard, G.; Crivello, F.; Joliot, M.; Perchey, G.; Mellet, E.; Petit, L.; Tzourio-Mazoyer, N. Gaussian Mixture Modeling of Hemispheric Lateralization for Language in a Large Sample of Healthy Individuals Balanced for Handedness. PLoS ONE 2014, 9, e101165. [Google Scholar] [CrossRef] [Green Version]
- Corballis, M. How Asymmetries Evolved: Hearts, Brains, and Molecules. Symmetry 2021, 13, 914. [Google Scholar] [CrossRef]
- Vingerhoets, G.; Gerrits, R.; Verhelst, H. Atypical Brain Asymmetry in Human Situs Inversus: Gut Feeling or Real Evidence? Symmetry 2021, 13, 695. [Google Scholar] [CrossRef]
- Medland, S.E.; Duffy, D.L.; Wright, M.; Geffen, G.M.; Hay, D.A.; Levy, F.; Van-Beijsterveldt, C.E.; Willemsen, G.; Townsend, G.C.; White, V.; et al. Genetic influences on handedness: Data from 25,732 Australian and Dutch twin families. Neuropsychologia 2009, 47, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.J. Innate: How the Wiring of Our Brains Shapes Who We Are. Innate: How the Wiring of Our Brains Shapes Who We Are; Princeton University Press: Princeton, NJ, USA, 2020. [Google Scholar]
- McManus, C. Is any but a tiny fraction of handedness variance likely to be due to the external environment? Laterality 2021, 26, 310–314. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Berretz, G.; Packheiser, J.; Friedrich, P. Laterality 2020: Entering the next decade. Laterality 2021, 26, 265–297. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Berretz, G.; Packheiser, J.; Friedrich, P. Laterality 2020: Response to the article commentaries. Laterality 2021, 26, 348–357. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Annett, M. A classification of hand preference by association analysis. Br. J. Psychol. 1970, 61, 303–321. [Google Scholar] [CrossRef]
- Paracchini, S.; Scerri, T. Genetics of human handedness and laterality. In Lateralized Brain Functions; Rogers, L., Vallortigara, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 523–552. [Google Scholar]
- Boyd, A.; Golding, J.; Macleod, J.; Lawlor, D.A.; Fraser, A.; Henderson, J.; Molloy, L.; Ness, A.; Ring, S.; Davey Smith, G. Cohort Profile: The ‘children of the 90s’—The index offspring of the Avon Longitudinal Study of Parents and Children. Int. J. Epidemiol. 2013, 42, 111–127. [Google Scholar] [CrossRef] [Green Version]
- Castillo, C.B.; Lynch, A.G.; Paracchini, S. Different laterality indexes are poorly correlated with one another but consistently show the tendency of males and females to be more left- and right-lateralized, respectively. R. Soc. Open Sci. 2020, 7, 191700. [Google Scholar] [CrossRef] [Green Version]
- Bycroft, C.; Freeman, C.; Petkova, D.; Band, G.; Elliott, L.T.; Sharp, K.; Motyer, A.; Vukcevic, D.; Delaneau, O.; O’Connell, J.; et al. The UK Biobank resource with deep phenotyping and genomic data. Nat. Cell Biol. 2018, 562, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuellar-Partida, G.; Tung, J.Y.; Eriksson, N.; Albrecht, E.; Aliev, F.; Andreassen, O.A.; Barroso, I.; Beckmann, J.S.; Boks, M.P.; Boomsma, D.I.; et al. Genome-wide association study identifies 48 common genetic variants associated with handedness. Nat. Hum. Behav. 2021, 5, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Doust, C.; Fontanillas, P.; Eising, E.; Gordon, S.D.; Wang, Z.; Alagöz, G.; Molz, B.; St Pourcain, B.; Francks, C.; Marioni, R.E.; et al. Discovery of 42 Genome-Wide Significant Loci Associated with Dyslexia. medRxiv 2021. [Google Scholar] [CrossRef]
- Schmitz, J.; Zheng, M.; Lui, K.F.H.; McBride, C.; Ho, C.S.-H.; Paracchini, S. Quantitative multidimensional phenotypes improve genetic analysis of laterality traits. bioRxiv 2021. [Google Scholar] [CrossRef]
- Verhoef, E.; Shapland, C.Y.; Fisher, S.E.; Dale, P.S.; Pourcain, B.S. The developmental origins of genetic factors influencing language and literacy: Associations with early-childhood vocabulary. J. Child Psychol. Psychiatry 2021, 62, 728–738. [Google Scholar] [CrossRef]
- Suzuki, K.; Ando, J. Genetic and environmental structure of individual differences in hand, foot, and ear preferences: A twin study. Laterality 2013, 19, 113–128. [Google Scholar] [CrossRef]
- Warren, D.M.; Stern, M.; Duggirala, R.; Dyer, T.D.; Almasy, L. Heritability and linkage analysis of hand, foot, and eye preference in Mexican Americans. Laterality 2006, 11, 508–524. [Google Scholar] [CrossRef]
- Packheiser, J.; Schmitz, J.; Berretz, G.; Carey, D.P.; Paracchini, S.; Papadatou-Pastou, M.; Ocklenburg, S. Four meta-analyses across 164 studies on atypical footedness prevalence and its relation to handedness. Sci. Rep. 2020, 10, 14501. [Google Scholar] [CrossRef]
- Annett, M. Left, Right, Hand and Brain: The Right Shift Theory; Psychology Press: London, UK, 1985. [Google Scholar]
- McManus, I. Right- and left-hand skill: Failure of the right shift model. Br. J. Psychol. 1985, 76, 1–16. [Google Scholar] [CrossRef]
- Claussnitzer, M.; Cho, J.H.; Collins, R.; Cox, N.J.; Dermitzakis, E.T.; Hurles, M.E.; Kathiresan, S.; Kenny, E.E.; Lindgren, C.M.; MacArthur, D.G.; et al. A brief history of human disease genetics. Nat. Cell Biol. 2020, 577, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, C.F.; Fitzgerald, T.W.; Jones, W.D.; Clayton, S.; McRae, J.F.; van Kogelenberg, M.; King, D.A.; Ambridge, K.; Barrett, D.M.; Bayzetinova, T.; et al. Genetic diagnosis of developmental disorders in the DDD study: A scalable analysis of genome-wide research data. Lancet 2015, 385, 1305–1314. [Google Scholar] [CrossRef] [Green Version]
- Diquigiovanni, C.; Bergamini, C.; Diaz, R.; Liparulo, I.; Bianco, F.; Masin, L.; Baldassarro, V.A.; Rizzardi, N.; Tranchina, A.; Buscherini, F.; et al. A novel mutation in SPART gene causes a severe neurodevelopmental delay due to mitochondrial dysfunction with complex I impairments and altered pyruvate metabolism. FASEB J. 2019, 33, 11284–11302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postema, M.C.; Carrion-Castillo, A.; Fisher, S.E.; Vingerhoets, G.; Francks, C. The genetics of situs inversus without primary ciliary dyskinesia. Sci. Rep. 2020, 10, 3677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.S.L.; Fisher, S.; Hurst, J.A.; Vargha-Khadem, F.; Monaco, A. A forkhead-domain gene is mutated in a severe speech and language disorder. Nat. Cell Biol. 2001, 413, 519–523. [Google Scholar] [CrossRef]
- Martinelli, A.; Rice, M.L.; Talcott, J.B.; Diaz, R.; Smith, S.; Raza, M.H.; Snowling, M.J.; Hulme, C.; Stein, J.; Hayiou-Thomas, M.E.; et al. A rare missense variant in the ATP2C2 gene is associated with language impairment and related measures. Hum. Mol. Genet. 2021, 30, 1160–1171. [Google Scholar] [CrossRef]
- Ceroni, F.; Simpson, N.H.; Francks, C.; Baird, G.; Conti-Ramsden, G.; Clark, A.; Bolton, P.F.; Hennessy, E.R.; Donnelly, P.; Bentley, D.R.; et al. Homozygous microdeletion of exon 5 in ZNF277 in a girl with specific language impairment. Eur. J. Hum. Genet. 2014, 22, 1165–1171. [Google Scholar] [CrossRef] [Green Version]
- Ocklenburg, S.; Barutçuoğlu, C.; Özgören, A.Ö.; Özgören, M.; Erdal, E.; Moser, D.; Schmitz, J.; Kumsta, R.; Güntürkün, O. The Genetics of Asymmetry: Whole Exome Sequencing in a Consanguineous Turkish Family with an Overrepresentation of Left-Handedness. Symmetry 2017, 9, 66. [Google Scholar] [CrossRef] [Green Version]
- Kavaklioglu, T.; Ajmal, M.; Hameed, A.; Francks, C. Whole exome sequencing for handedness in a large and highly consanguineous family. Neuropsychologia 2016, 93, 342–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McManus, I.C.; Davison, A.; Armour, J. Multilocus genetic models of handedness closely resemble single-locus models in explaining family data and are compatible with genome-wide association studies. Ann. N. Y. Acad. Sci. 2013, 1288, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Del Castillo, U.; Norkett, R.; Gelfand, V.I. Unconventional Roles of Cytoskeletal Mitotic Machinery in Neurodevelopment. Trends Cell Biol. 2019, 29, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Lasser, M.; Tiber, J.; Lowery, L.A. The Role of the Microtubule Cytoskeleton in Neurodevelopmental Disorders. Front. Cell. Neurosci. 2018, 12, 165. [Google Scholar] [CrossRef] [Green Version]
- Hirnstein, M.; Hugdahl, K. Excess of non-right-handedness in schizophrenia: Meta-analysis of gender effects and potential biases in handedness assessment. Br. J. Psychiatry 2014, 205, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Wiberg, A.; Ng, M.; Al Omran, Y.; Alfaro-Almagro, F.; McCarthy, P.; Marchini, J.; Bennett, D.; Smith, S.; Douaud, G.; Furniss, D. Handedness, language areas and neuropsychiatric diseases: Insights from brain imaging and genetics. Brain 2019, 142, 2938–2947. [Google Scholar] [CrossRef] [Green Version]
- Brandler, W.M.; Morris, A.P.; Evans, D.M.; Scerri, T.S.; Kemp, J.P.; Timpson, N.J.; Pourcain, B.S.; Smith, G.D.; Ring, S.M.; Stein, J.; et al. Common Variants in Left/Right Asymmetry Genes and Pathways Are Associated with Relative Hand Skill. PLoS Genet. 2013, 9, e1003751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandler, W.M.; Paracchini, S. The genetic relationship between handedness and neurodevelopmental disorders. Trends Mol. Med. 2014, 20, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shore, R.; Covill, L.; Pettigrew, K.; Brandler, W.M.; Diaz, R.; Xu, Y.; Tello, J.A.; Talcott, J.B.; Newbury, D.F.; Stein, J.; et al. The handedness-associated PCSK6 locus spans an intronic promoter regulating novel transcripts. Hum. Mol. Genet. 2016, 25, 1771–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paracchini, S.; Diaz, R.; Stein, J. Advances in Dyslexia Genetics—New Insights into the Role of Brain Asymmetries. Adv. Genet. 2016, 96, 53–97. [Google Scholar] [CrossRef] [PubMed]
- Pearson, K.; Lee, A. On the inheritance of characters not capable if exact quantitative measurement. Philos. Trans. R. Soc. Lond. Ser. A Math. Phys. Sci. 1901, 195, 79–150. [Google Scholar]
- Wray, N.R.; Wijmenga, C.; Sullivan, P.F.; Yang, J.; Visscher, P.M. Common Disease Is More Complex than Implied by the Core Gene Omnigenic Model. Cell 2018, 173, 1573–1580. [Google Scholar] [CrossRef] [Green Version]
- Rietveld, C.A.; Medland, S.E.; Derringer, J.; Yang, J.; Esko, T.; Martin, N.W.; Westra, H.-J.; Shakhbazov, K.; Abdellaoui, A.; Agrawal, A.; et al. GWAS of 126,559 Individuals Identifies Genetic Variants Associated with Educational Attainment. Science 2013, 340, 1467–1471. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Wedow, R.; Okbay, A.; Kong, E.; Maghzian, O.; Zacher, M.; Nguyen-Viet, T.A.; Bowers, P.; Sidorenko, J.; Linnér, R.K.; et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat. Genet. 2018, 50, 1112–1121. [Google Scholar] [CrossRef] [Green Version]
- Selzam, S.; Dale, P.S.; Wagner, R.K.; DeFries, J.C.; Cederlöf, M.; O’Reilly, P.F.; Krapohl, E.; Plomin, R. Genome-Wide Polygenic Scores Predict Reading Performance throughout the School Years. Sci. Stud. Read. 2017, 21, 334–349. [Google Scholar] [CrossRef] [Green Version]
- Gialluisi, A.; Andlauer, T.F.M.; Mirza-Schreiber, N.; Moll, K.; Becker, J.; Hoffmann, P.; Ludwig, K.U.; Czamara, D.; Pourcain, B.S.; Brandler, W.; et al. Genome-wide association scan identifies new variants associated with a cognitive predictor of dyslexia. Transl. Psychiatry 2019, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Gialluisi, A.; Andlauer, T.F.M.; Mirza-Schreiber, N.; Moll, K.; Becker, J.; Hoffmann, P.; Ludwig, K.U.; Czamara, D.; Pourcain, B.S.; Honbolygó, F.; et al. Genome-wide association study reveals new insights into the heritability and genetic correlates of developmental dyslexia. Mol. Psychiatry 2020. [Google Scholar] [CrossRef]
- De Kovel, C.G.F.; Francks, C. The molecular genetics of hand preference revisited. Sci. Rep. 2019, 91, 5986. [Google Scholar] [CrossRef] [Green Version]
- Ocklenburg, S.; Metzen, D.; Schlüter, C.; Fraenz, C.; Arning, L.; Streit, F.; Güntürkün, O.; Kumsta, R.; Genç, E. Polygenic scores for handedness and their association with asymmetries in brain structure. Anat. Embryol. 2021, 1, 1–13. [Google Scholar] [CrossRef]
- Kong, X.-Z.; Postema, M.; Schijven, D.; Castillo, A.C.; Pepe, A.; Crivello, F.; Joliot, M.; Mazoyer, B.; Fisher, S.E.; Francks, C. Large-Scale Phenomic and Genomic Analysis of Brain Asymmetrical Skew. Cereb. Cortex 2021, 31, 4151–4168. [Google Scholar] [CrossRef] [PubMed]
- Sha, Z.; Pepe, A.; Schijven, D.; Carrion Castillo, A.; Roe, J.M.; Westerhausen, R.; Joliot, M.; Fisher, S.E.; Crivello, F.; Francks, C. Left-handedness and its genetic influences are associated with structural asymmetries mapped across the cerebral cortex in 31,864 individuals. bioRxiv 2021. [Google Scholar] [CrossRef]
- Eriksson, N.; MacPherson, J.M.; Tung, J.Y.; Hon, L.S.; Naughton, B.; Saxonov, S.; Avey, L.; Wojcicki, A.; Pe’Er, I.; Mountain, J. Web-Based, Participant-Driven Studies Yield Novel Genetic Associations for Common Traits. PLoS Genet. 2010, 6, e1000993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scerri, T.S.; Brandler, W.M.; Paracchini, S.; Morris, A.P.; Ring, S.M.; Richardson, A.J.; Talcott, J.B.; Stein, J.; Monaco, A.P. PCSK6 is associated with handedness in individuals with dyslexia. Hum. Mol. Genet. 2010, 20, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.J.; Woodhead, Z.V.J.; Thompson, P.A.; Bishop, D.V.M. Assessing the reliability of an online behavioural laterality battery: A pre-registered study. Laterality 2020, 26, 359–397. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paracchini, S. Recent Advances in Handedness Genetics. Symmetry 2021, 13, 1792. https://doi.org/10.3390/sym13101792
Paracchini S. Recent Advances in Handedness Genetics. Symmetry. 2021; 13(10):1792. https://doi.org/10.3390/sym13101792
Chicago/Turabian StyleParacchini, Silvia. 2021. "Recent Advances in Handedness Genetics" Symmetry 13, no. 10: 1792. https://doi.org/10.3390/sym13101792
APA StyleParacchini, S. (2021). Recent Advances in Handedness Genetics. Symmetry, 13(10), 1792. https://doi.org/10.3390/sym13101792