Symmetric and Asymmetric Components of Shape Variation in the Diatom Genus Frustulia (Bacillariophyta)


Abstract
1. Introduction
2. Materials and Methods
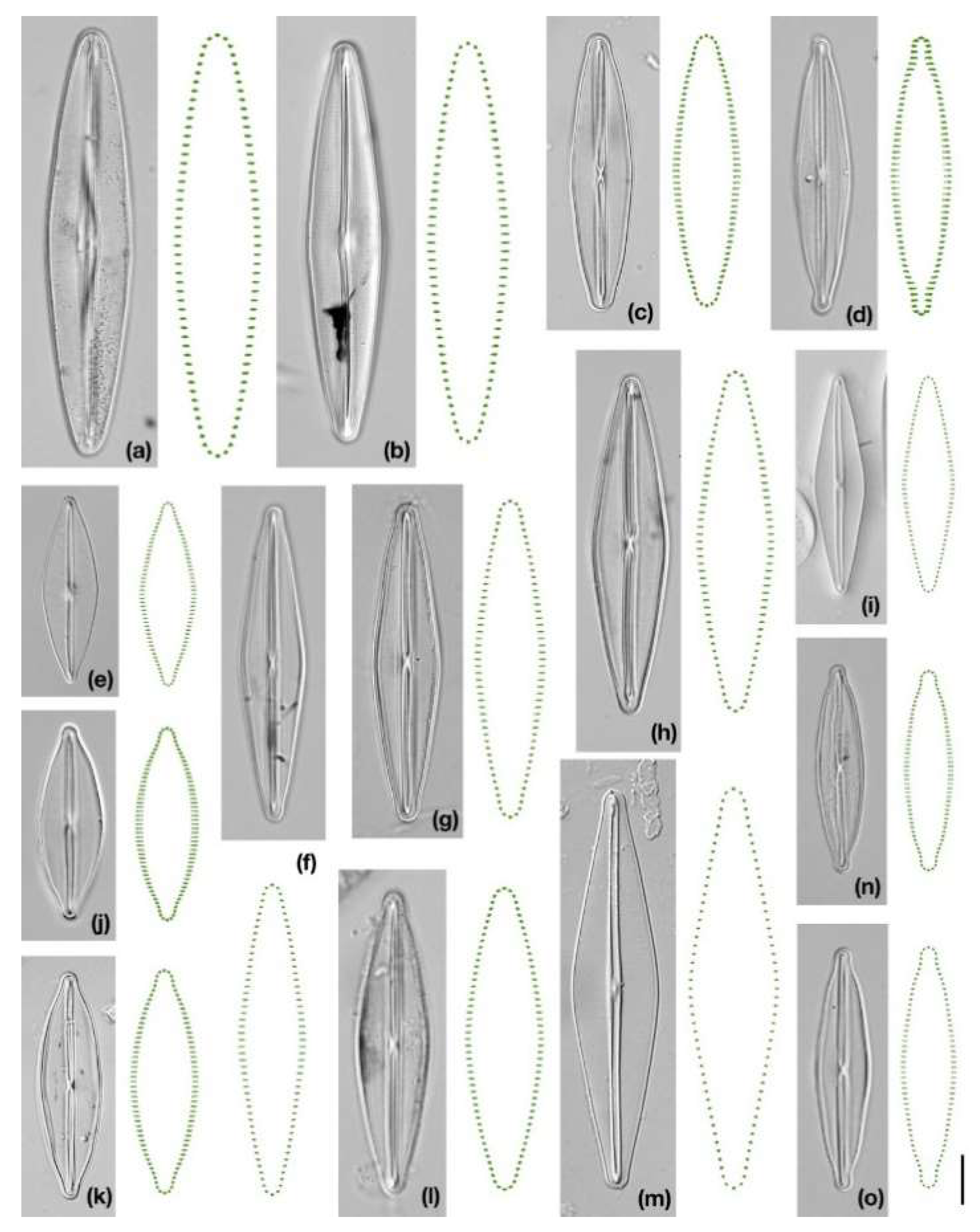
2.1. Characteristics of Strains and Data Acquisition
2.2. Geometric Morphometric Analyses
2.3. Reconstruction of Phylogeny and Trait Mapping
3. Results
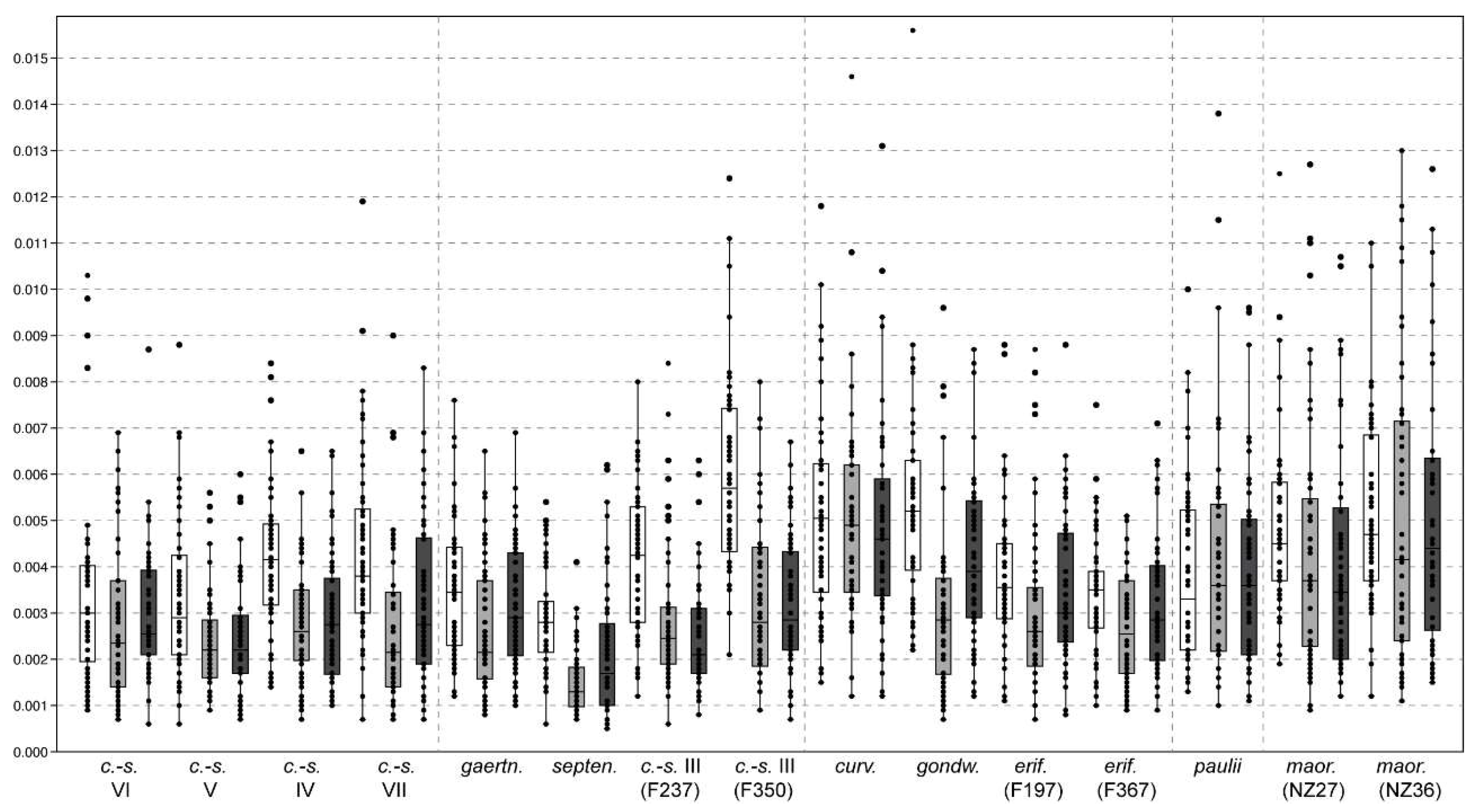
3.1. Quantification of Components of Shape Variation
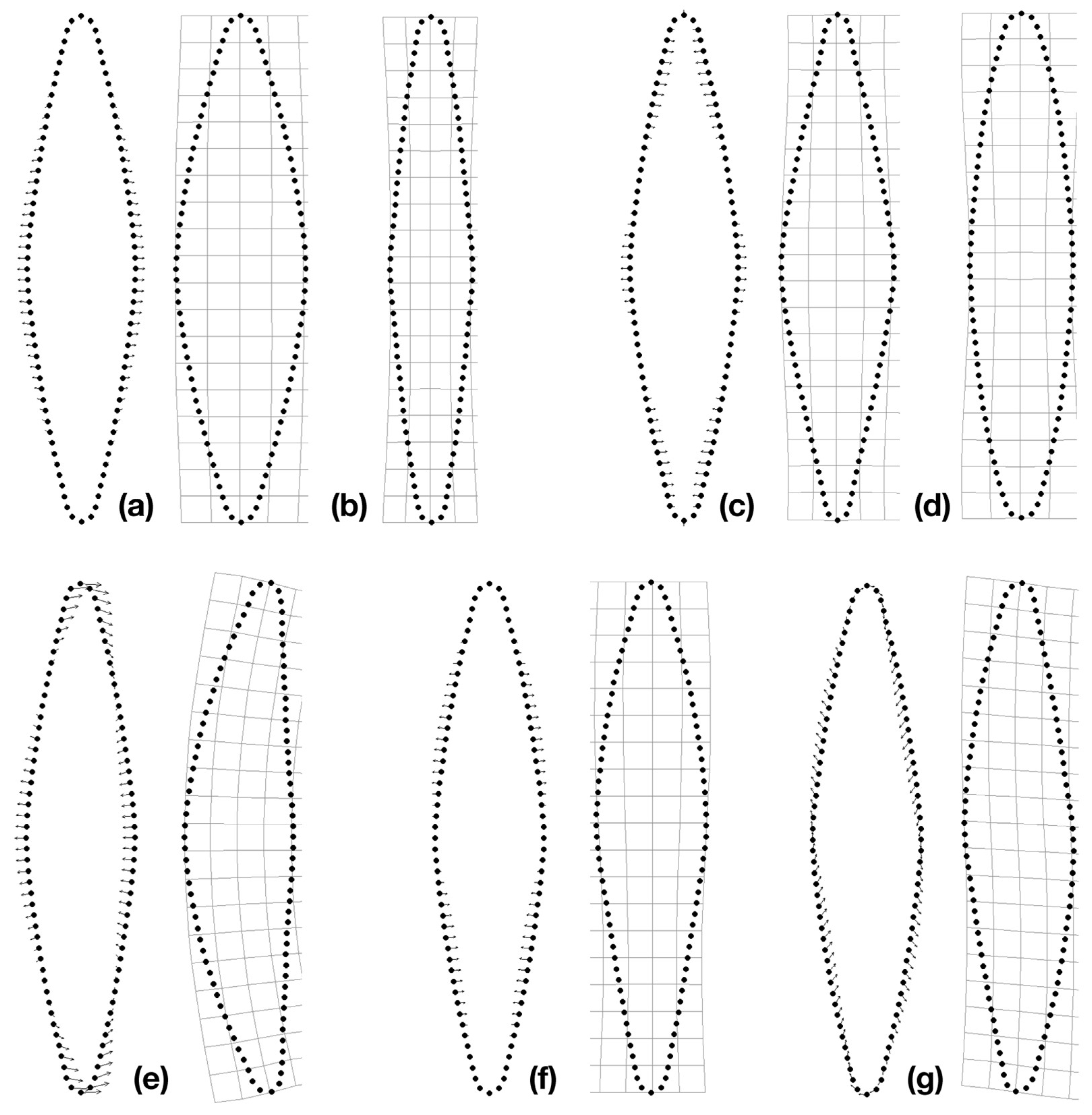
3.2. Reconstruction of Shape Changes
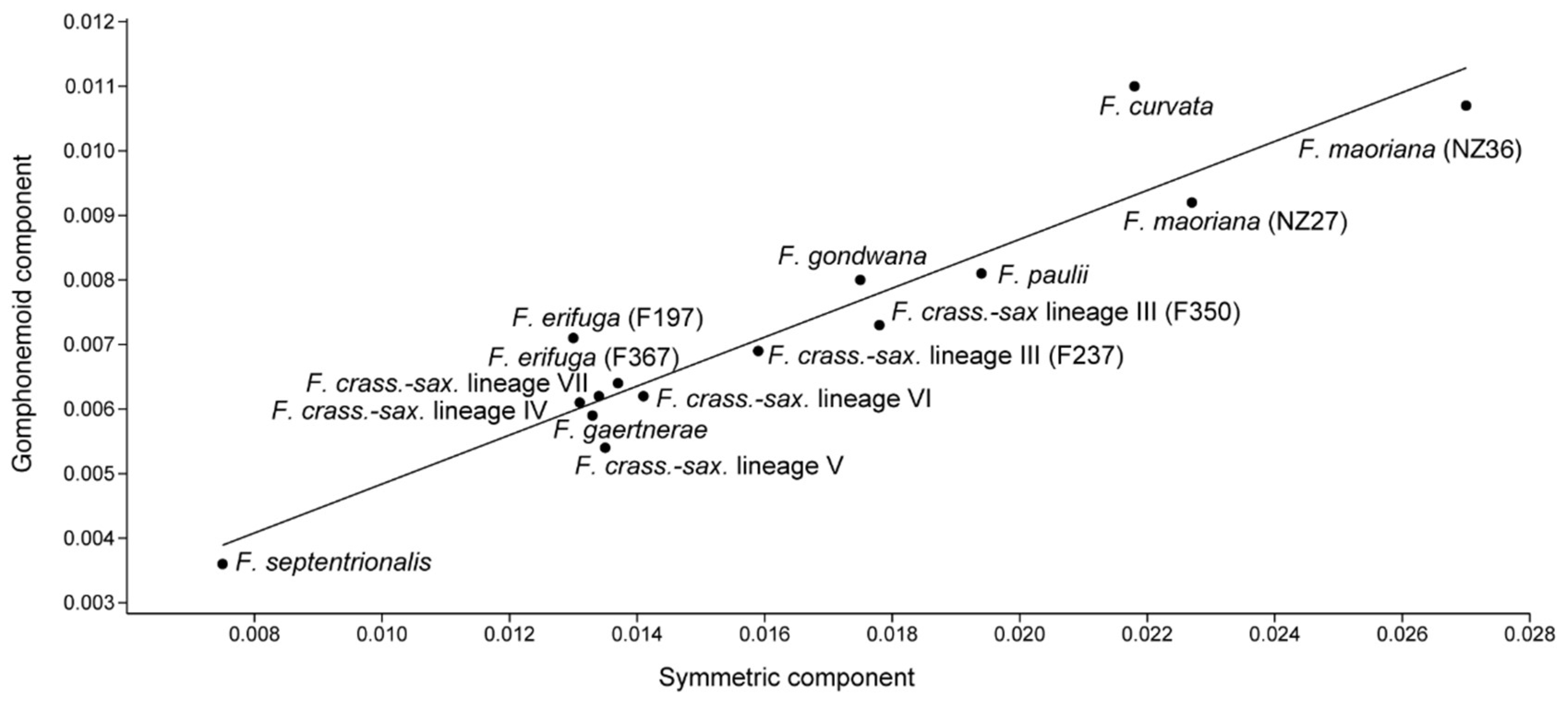
3.3. Phylogeny and Shape Components
4. Discussion
4.1. Shape Variation within Lineages
4.2. Phylogeny and Asymmetric Variation
4.3. Common Asymmetric Shape Variation across Lineages
4.4. Relationship between Asymmetric and Symmetric Variation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatoms Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 1990; p. 747. [Google Scholar]
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Mann, D.G.; Vanormelingen, P. An inordinate fondness? The number, distributions, and origins of diatom species. J. Eukaryot. Microbiol. 2013, 60, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Ruck, E.C.; Theriot, E.C. Origin and evolution of the canal raphe system in diatoms. Protist 2011, 162, 723–737. [Google Scholar] [CrossRef]
- Nakov, T.; Ruck, E.C.; Galachyants, Y.; Spaulding, S.A.; Theriot, E.C. Molecular phylogeny of the Cymbellales (Bacillariophyceae, Heterokontophyta) with a comparison of models for accommodating rate variation across sites. Phycologia 2014, 53, 359–373. [Google Scholar] [CrossRef]
- Ashworth, M.P.; Lobban, C.S.; Witkowski, A.; Theriot, E.C.; Sabir, M.J.; Baeshen, M.N.; Hajarah, N.H.; Baeshen, N.A.; Sabir, J.S.; Jansen, R.K. Molecular and morphological investigations of the stauros-bearing, raphid pennate diatoms (Bacillariophyceae): Craspedostauros EJ Cox, and Staurotropis TBB Paddock, and their relationship to the rest of the Mastogloiales. Protist 2017, 168, 48–70. [Google Scholar] [CrossRef] [PubMed]
- Mann, D. The origins of shape and form in diatoms: The interplay between morphometric studies and systemaics. In Shape and Form in Plants and Fungi; Academic Press: Cambridge, UK, 1994; pp. 17–38. [Google Scholar]
- Cox, E.J. Morphogenetic information and the selection of taxonomic characters for raphid diatom systematics. Plant Ecol. Evol. 2010, 143, 271–277. [Google Scholar] [CrossRef]
- Cox, E.J. Ontogeny, homology, and terminology—wall morphogenesis as an aid to character recognition and character state definition for pennate diatom systematics. J. Phycol. 2012, 48, 1–31. [Google Scholar] [CrossRef]
- MacDonald, J.D. On the structure of the Diatomaceous frustule, and its genetic cycle. J. Nat. Hist. 1869, 3, 1–8. [Google Scholar] [CrossRef]
- Pfitzer, E. Über den Bau und die Zellteilung der Diatomeen. Botanische Zeitung 1869, 27, 774–776. [Google Scholar]
- Geitler, L. Der Formwechsel der pennaten Diatomeen (Kieselalgen). Arch. Protistenkd. 1932, 78, 1–226. [Google Scholar]
- Edgar, R.K.; Kociolek, J.P.; Edgar, S.M. Life cycle-associated character variation in Aulacoseira krameri sp. nov., a new miocene species from Oregon, USA. Diatom Res. 2004, 19, 7–32. [Google Scholar] [CrossRef]
- Veselá, J.; Neustupa, J.; Pichrtová, M.; Poulíčková, A. Morphometric study of Navicula morphospecies (Bacillariophyta) with respect to diatom life cycle. Fottea 2009, 9, 307–316. [Google Scholar] [CrossRef]
- English, J.D.; Potapova, M.G. Ontogenetic and interspecific valve shape variation in the Pinnatae group of the genus Surirella and the description of S. Lacrimula sp. nov. Diatom Res. 2012, 27, 9–27. [Google Scholar] [CrossRef]
- Woodard, K.; Kulichová, J.; Poláčková, T.; Neustupa, J. Morphometric allometry of representatives of three naviculoid genera throughout their life cycle. Diatom Res. 2016, 31, 231–242. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Phenotypic plasticity, developmental instability, and robustness: The concepts and how they are connected. Front. Ecol. Evol. 2019, 7, 56. [Google Scholar] [CrossRef]
- Kulichová, J.; Neustupa, J.; Vrbová, K.; Levkov, Z.; Kopalová, K. Asymmetry in Luticola species. Diatom Res. 2019, 34, 67–74. [Google Scholar] [CrossRef]
- Kooistra, W.H.; De Stefano, M.; Mann, D.G.; Salma, N.; Medlin, L.K. Phylogenetic position of Toxarium, a pennate-like lineage within centric diatoms (Bacillariophyceae). J. Phycol. 2003, 39, 185–197. [Google Scholar] [CrossRef]
- Cantonati, M.; Angeli, N.; Virtanen, L.; Wojtal, A.Z.; Gabrieli, J.; Falasco, E.; Lavoie, I.; Morin, S.; Marchetto, A.; Fortin, C. Achnanthidium minutissimum (Bacillariophyta) valve deformities as indicators of metal enrichment in diverse widely-distributed freshwater habitats. Sci. Total Environ. 2014, 475, 201–215. [Google Scholar] [CrossRef]
- Olenici, A.; Blanco, S.; Borrego-Ramos, M.; Momeu, L.; Baciu, C. Exploring the effects of acid mine drainage on diatom teratology using geometric morphometry. Ecotoxicology 2017, 26, 1018–1030. [Google Scholar] [CrossRef]
- Cerisier, A.; Vedrenne, J.; Lavoie, I.; Morin, S. Assessing the severity of diatom deformities using geometric morphometry. Bot. Lett. 2019, 166, 32–40. [Google Scholar] [CrossRef]
- Lange-Bertalot, H. Navicula sensu stricto. 10 genera separated from Navicula sensu lato. Frustulia. In Diatoms of Europe—Diatoms of European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag, K. G.: Ruggell, Liechtenstein, 2001; Volume 2, p. 526. [Google Scholar]
- Siver, P.A.; Baskette, G. A morphological examination of Frustulia (Bacillariophyceae) from the Ocala National Forest, Florida, USA. Can. J. Bot. 2004, 82, 629–644. [Google Scholar] [CrossRef]
- Veselá, J.; Urbánková, P.; Černá, K.; Neustupa, J. Ecological variation within traditional diatom morphospecies: Diversity of Frustulia rhomboides sensu lato (Bacillariophyceae) in European freshwater habitats. Phycologia 2012, 51, 552–561. [Google Scholar] [CrossRef]
- Urbánková, P.; Scharfen, V.; Kulichová, J. Molecular and automated identification of the diatom genus Frustulia in northern Europe. Diatom Res. 2016, 31, 217–229. [Google Scholar] [CrossRef]
- Bouchard, A.J.; Hamilton, P.B.; Savoie, A.M.; Starr, J.R. Molecular and morphological data reveal hidden diversity in common North American Frustulia species (Amphipleuraceae). Diatom Res. 2019, 34, 205–223. [Google Scholar] [CrossRef]
- Urbánková, P.; Veselá, J. DNA-barcoding: A case study in the diatom genus Frustulia (Bacillariophyceae). Nova Hedwig. 2013, 142, 147–162. [Google Scholar]
- Nakov, T.; Beaulieu, J.M.; Alverson, A.J. Accelerated diversification is related to life history and locomotion in a hyperdiverse lineage of microbial eukaryotes (Diatoms, Bacillariophyta). N. Phytol. 2018, 219, 462–473. [Google Scholar] [CrossRef]
- Rohlf, F.J. The tps series of software. Hystrix 2015, 26, 9–12. [Google Scholar]
- Kloster, M.; Kauer, G.; Beszteri, B. SHERPA: An image segmentation and outline feature extraction tool for diatoms and other objects. BMC Bioinform. 2014, 15, 218. [Google Scholar] [CrossRef]
- Savriama, Y. A step-by-step guide for geometric morphometrics of floral symmetry. Front. Plant Sci. 2018, 9, 1433. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, D.H.; Fink, W.L. Geometric Morphometrics for Biologists: A Primer; Elsevier: Amsterdam, The Netherlands; Academic Press: London, UK, 2004; p. 455. [Google Scholar]
- Klingenberg, C.P. Evolution and development of shape: Integrating quantitative approaches. Nat. Rev. Genet. 2010, 11, 623–635. [Google Scholar] [CrossRef]
- MacLeod, N. Use of landmark and outline morphometrics to investigate thecal form variation in crushed gogiid echinoderms. Palaeoworld 2015, 24, 408–429. [Google Scholar] [CrossRef]
- MacLeod, N. Morphometrics: History, development methods and prospects. Zool. Syst. 2017, 42, 4–33. [Google Scholar]
- Savriama, Y.; Neustupa, J.; Klingenberg, C.P. Geometric morphometrics of symmetry and allometry in Micrasterias rotata (Zygnemophyceae, Viridiplantae). Nova Hedwig. Suppl. 2010, 136, 43–54. [Google Scholar] [CrossRef]
- Savriama, Y.; Klingenberg, C.P. Beyond bilateral symmetry: Geometric morphometric methods for any type of symmetry. BMC Evol. Biol. 2011, 11, 280. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.C.; Otárola-Castillo, E. Geomorph: An R package for the collection and analysis of geometric morphometric shape data. Methods Ecol. Evol. 2013, 4, 393–399. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 6 January 2018).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Yoon, H.S.; Hackett, J.D.; Bhattacharya, D. A single origin of the peridinin—and fucoxanthin-containing plastids in dinoflagellates through tertiary endosymbiosis. Proc. Natl. Acad. Sci. USA 2002, 99, 11724–11729. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. Ali View: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acid. Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Sánchez, R.; Serra, F.; Tárraga, J.; Medina, I.; Carbonell, J.; Pulido, L.; de María, A.; Capella-Gutíerrez, S.; Huerta-Cepas, J.; Gabaldón, T. Phylemon 2.0: A suite of web-tools for molecular evolution, phylogenetics, phylogenomics and hypotheses testing. Nucleic Acids Res. 2011, 39, W470–W474. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Chernomor, O.; Von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Boettiger, C.; Coop, G.; Ralph, P. Is your phylogeny informative? Measuring the power of comparative methods. Evolution 2012, 66, 2240–2251. [Google Scholar] [CrossRef]
- Mann, D.G. The species concept in diatoms. Phycologia 1999, 38, 437–495. [Google Scholar] [CrossRef]
- Woodard, K.; Neustupa, J. Morphometric asymmetry of frustule outlines in the pennate diatom Luticola poulickovae (Bacillariophyceae). Symmetry 2016, 8, 150. [Google Scholar] [CrossRef]
- Urbánková, P.; Kulichová, J.; Kilroy, C. Frustulia curvata and Frustulia paulii, two diatom species new to science. Diatom Res. 2015, 30, 65–73. [Google Scholar] [CrossRef]
- Poulickova, A. Morphology, cytology and sexual reproduction in aerophytic cave diatom Luticola dismutica (Bacillariophyceae). Preslia 2008, 80, 87–99. [Google Scholar]
- Waddington, C.H. Canalization of development and the inheritance of acquired characters. Nature 1942, 150, 563–565. [Google Scholar] [CrossRef]
- Mann, D.G. Symmetry and cell division in raphid diatoms. Ann. Bot. 1983, 52, 573–581. [Google Scholar] [CrossRef]
- Cox, E.J.; Williams, D.M. Systematics of naviculoid diatoms (Bacillariophyta): A preliminary analysis of protoplast and frustule characters for family and order level classification. Syst. Biodivers. 2006, 4, 385–399. [Google Scholar] [CrossRef]
- Pavlov, A.; Nakov, T.; Levkov, Z.; Furey, P.; Lowe, R.; Ector, L. Luticola grupcei (Bacillariophyceae)—a new freshwater diatom from Mountain Baba (Macedonia) and Great Smoky Mountains National Park (USA): Comparison with the type material of L. Goeppertiana (Bleisch) D.G. Mann. Nov. Hedwig. 2009, 89, 147–164. [Google Scholar] [CrossRef]
- Mann, D.G.; Stickle, A.J. Nuclear movements and frustule symmetry in raphid pennate diatoms. In Proceedings of the 9th International Diatom Symposium, Bristol, UK, 24–30 August 1986; Round, F.E., Ed.; Biopress: Bristol, UK; Koeltz Scientific Books Koenigstein: Oberreifenberg, Germany, 1988; pp. 281–289. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lineage | Strain | Symmetric | Cymbelloid | Gomphonemoid | Sigmoid | Digit. | |||
|---|---|---|---|---|---|---|---|---|---|
| abb. 1 | Code | Total | Total | Median | Total | Median | Total | Median | Error 2 |
| c.-s. III 3 | F237 | 0.0159 | 0.0090 | 0.0042 | 0.0069 | 0.0025 | 0.0063 | 0.0021 | 8% |
| c.-s. III | F350 | 0.0178 | 0.0115 | 0.0057 | 0.0073 | 0.0028 | 0.0073 | 0.0028 | 9% |
| c.-s. IV | F259 | 0.0131 | 0.0088 | 0.0041 | 0.0061 | 0.0026 | 0.0064 | 0.0028 | 9% |
| c.-s. V | F77 | 0.0135 | 0.0073 | 0.0029 | 0.0054 | 0.0022 | 0.0055 | 0.0022 | 7% |
| c.-s. VI | F288 | 0.0141 | 0.0078 | 0.0030 | 0.0062 | 0.0023 | 0.0065 | 0.0025 | 11% |
| c.-s. VII | NZ39 | 0.0134 | 0.0087 | 0.0038 | 0.0062 | 0.0022 | 0.0065 | 0.0027 | 8% |
| curv. 4 | F381 | 0.0218 | 0.0121 | 0.0050 | 0.0110 | 0.0049 | 0.0113 | 0.0046 | 5% |
| erif. 5 | F197 | 0.0130 | 0.0078 | 0.0035 | 0.0071 | 0.0026 | 0.0065 | 0.0030 | 8% |
| erif. | F367 | 0.0137 | 0.0074 | 0.0035 | 0.0064 | 0.0025 | 0.0060 | 0.0029 | 6% |
| gaertn. 6 | 28-9C | 0.0133 | 0.0076 | 0.0035 | 0.0059 | 0.0022 | 0.0065 | 0.0029 | 12% |
| gondw. 7 | NZ37 | 0.0175 | 0.0107 | 0.0052 | 0.0080 | 0.0029 | 0.0073 | 0.0039 | 4% |
| maor. 8 | NZ27 | 0.0227 | 0.0104 | 0.0045 | 0.0092 | 0.0037 | 0.0087 | 0.0034 | 5% |
| maor. | NZ36 | 0.0270 | 0.0112 | 0.0047 | 0.0107 | 0.0042 | 0.0104 | 0.0044 | 3% |
| paulii | NZ13 | 0.0194 | 0.0092 | 0.0033 | 0.0081 | 0.0036 | 0.0091 | 0.0036 | 8% |
| septen. 9 | 26-4B | 0.0075 | 0.0055 | 0.0028 | 0.0036 | 0.0013 | 0.0043 | 0.0017 | 21% |
| Symmetric | Cymbelloid | Gomphonemoid | Sigmoid | Mean CS 1 | |
|---|---|---|---|---|---|
| Symmetric | 0.80 ** | 0.85 *** | 0.78 ** | −0.74 * | |
| Cymbelloid | 0.74 * | 0.87 *** | 0.86 *** | n.s. | |
| Gomphonemoid | 0.76 * | 0.83 ** | 0.88 *** | n.s. | |
| Sigmoid | n.s. | n.s. | 0.88 ** | n.s. | |
| Mean CS | −0.72 * | n.s. | n.s. | n.s. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulichová, J.; Urbánková, P. Symmetric and Asymmetric Components of Shape Variation in the Diatom Genus Frustulia (Bacillariophyta). Symmetry 2020, 12, 1626. https://doi.org/10.3390/sym12101626
Kulichová J, Urbánková P. Symmetric and Asymmetric Components of Shape Variation in the Diatom Genus Frustulia (Bacillariophyta). Symmetry. 2020; 12(10):1626. https://doi.org/10.3390/sym12101626
Chicago/Turabian StyleKulichová, Jana, and Pavla Urbánková. 2020. "Symmetric and Asymmetric Components of Shape Variation in the Diatom Genus Frustulia (Bacillariophyta)" Symmetry 12, no. 10: 1626. https://doi.org/10.3390/sym12101626
APA StyleKulichová, J., & Urbánková, P. (2020). Symmetric and Asymmetric Components of Shape Variation in the Diatom Genus Frustulia (Bacillariophyta). Symmetry, 12(10), 1626. https://doi.org/10.3390/sym12101626