Chiral Neuronal Motility: The Missing Link between Molecular Chirality and Brain Asymmetry


Abstract
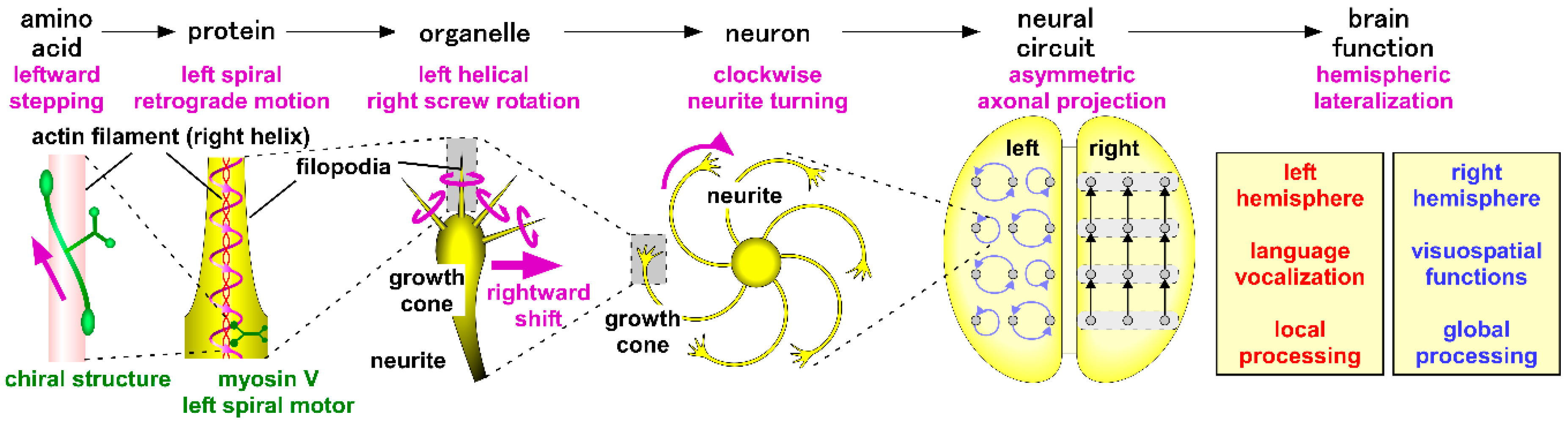
1. Introduction
2. Left–Right Brain Asymmetry in Humans
3. Left–Right Brain Asymmetry in Non-Human Animals
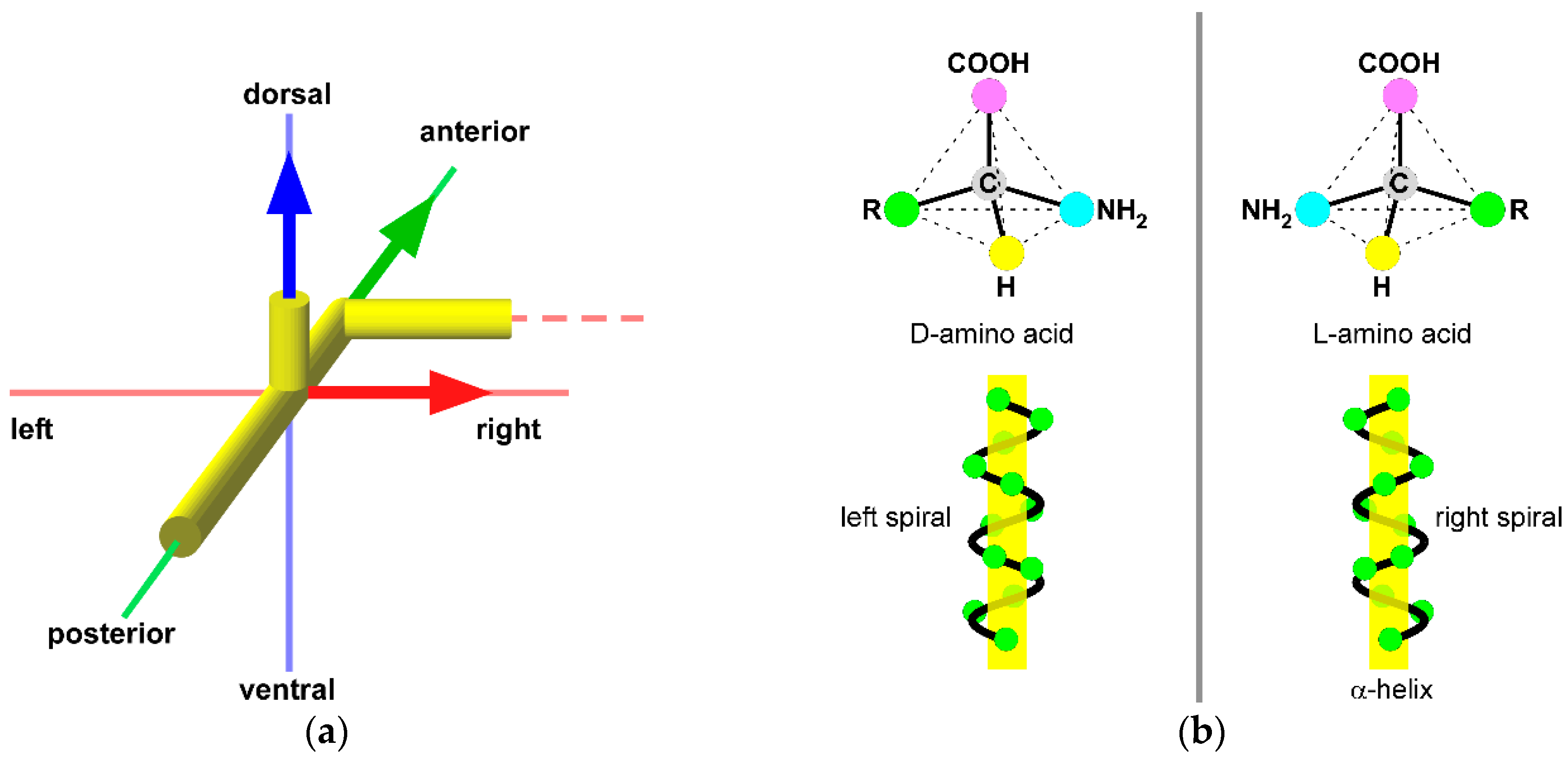
4. Development of Body Asymmetry
5. Brain Asymmetry versus Visceral Asymmetry
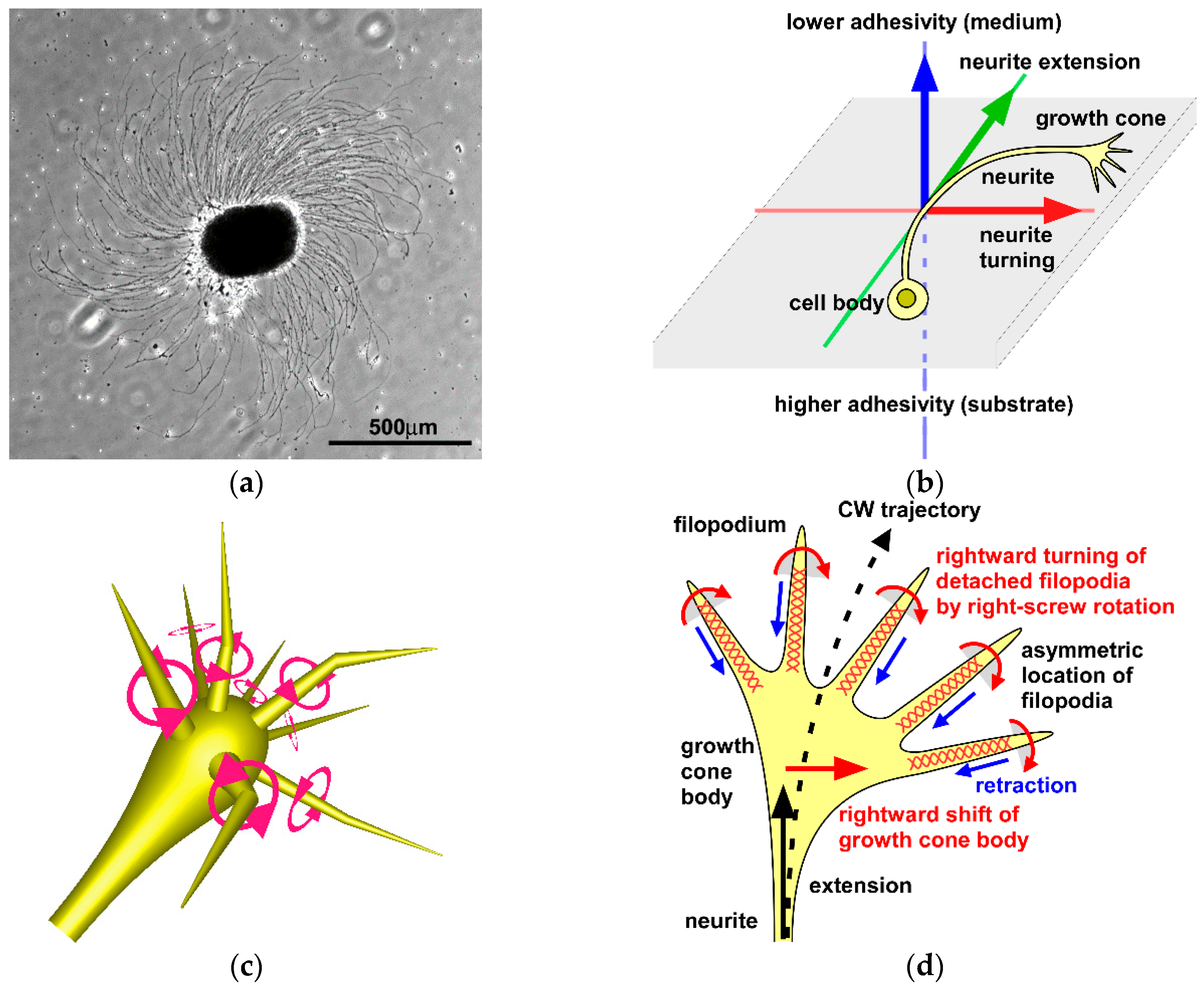
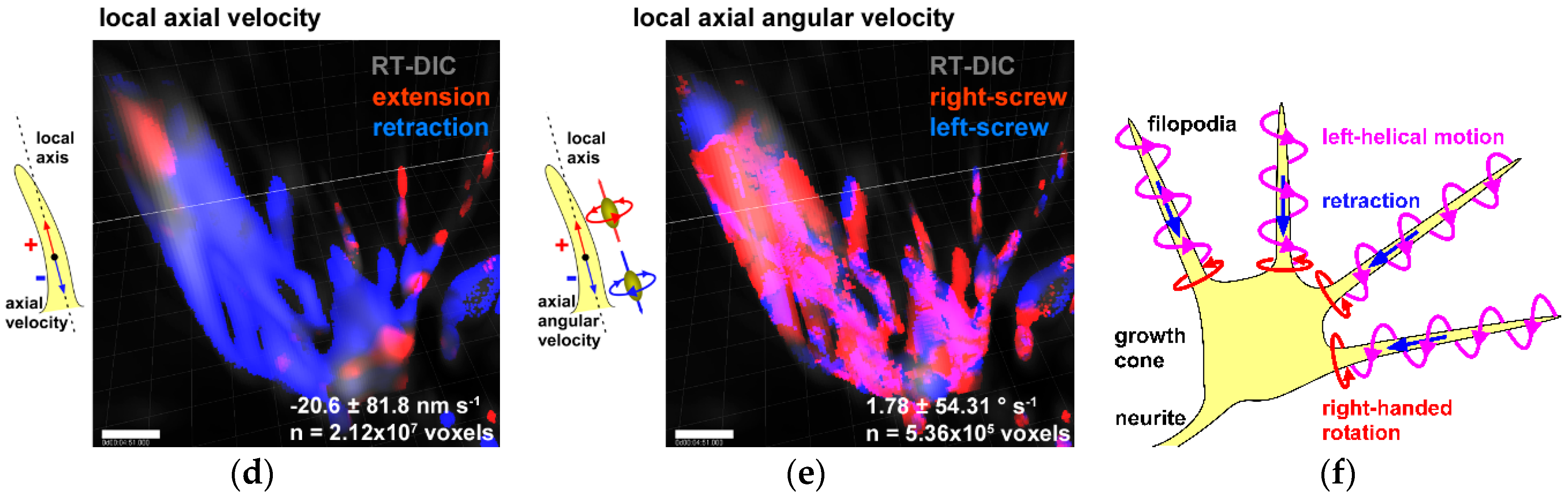
6. Chiral Neuronal Motility
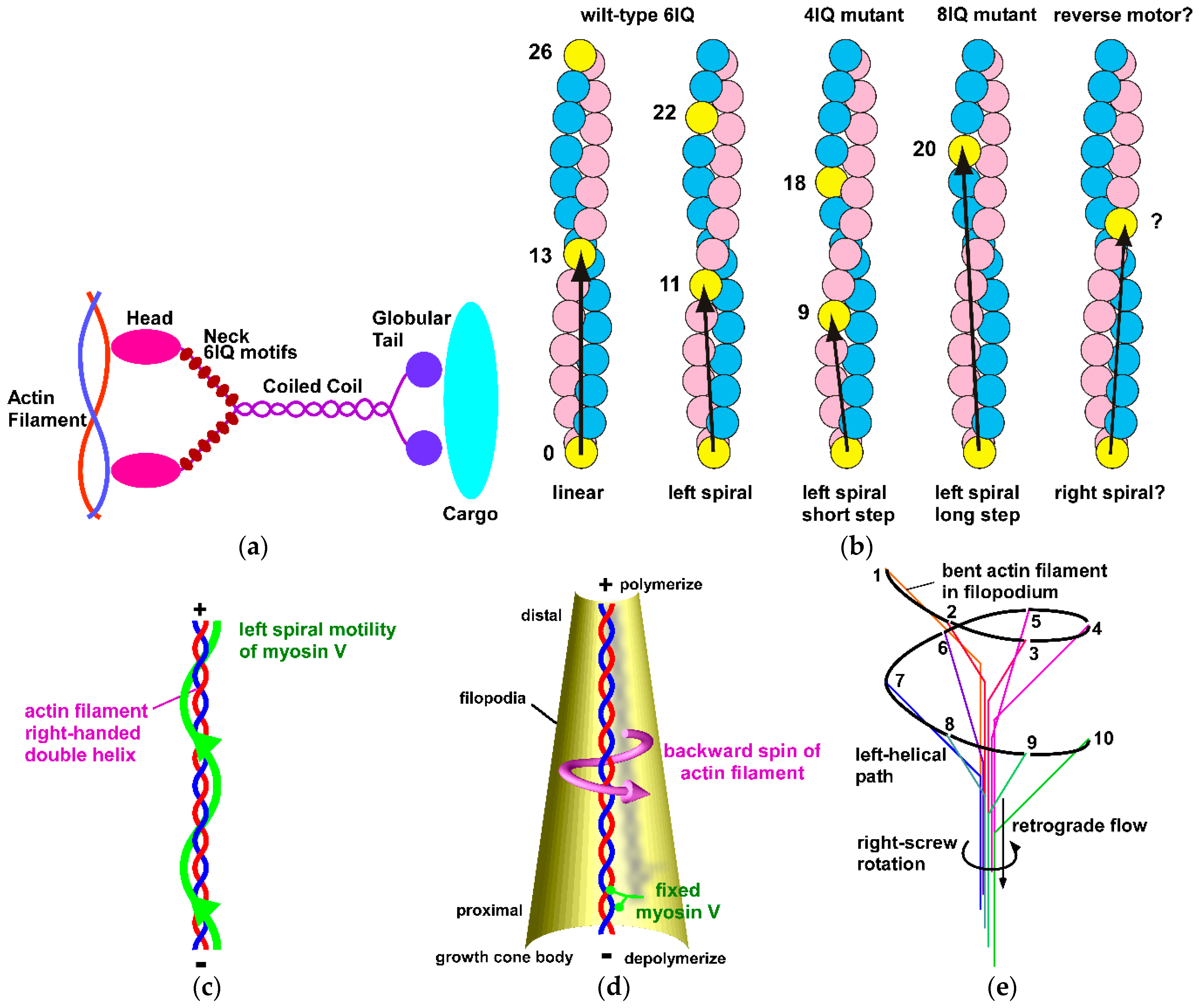
7. Molecular Mechanism Generating Chiral Neuronal Motility
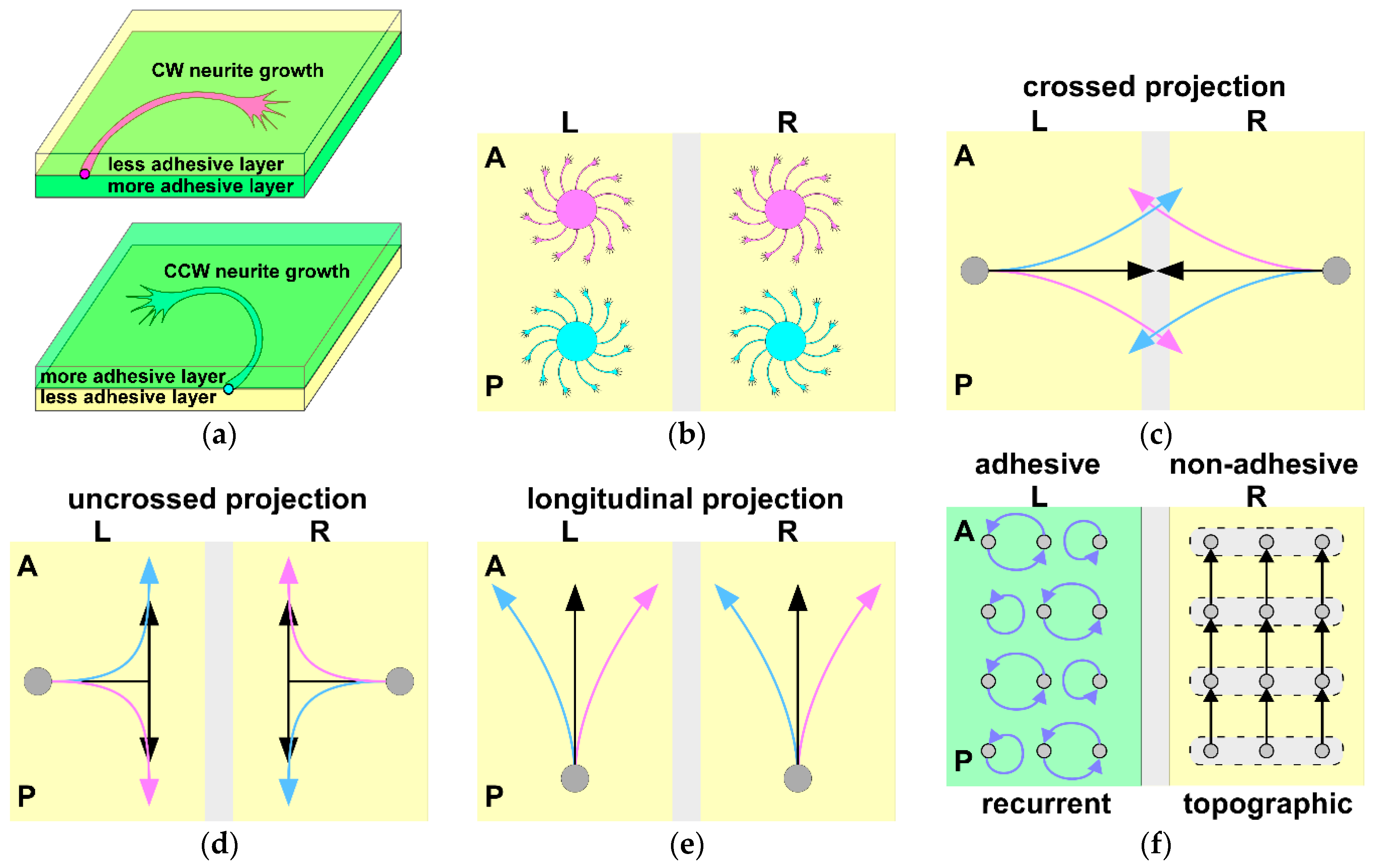
8. Role of Chiral Neuronal Motility in Formation of Brain Asymmetry
9. Methods for Analyzing Chirality and Left–Right Asymmetry
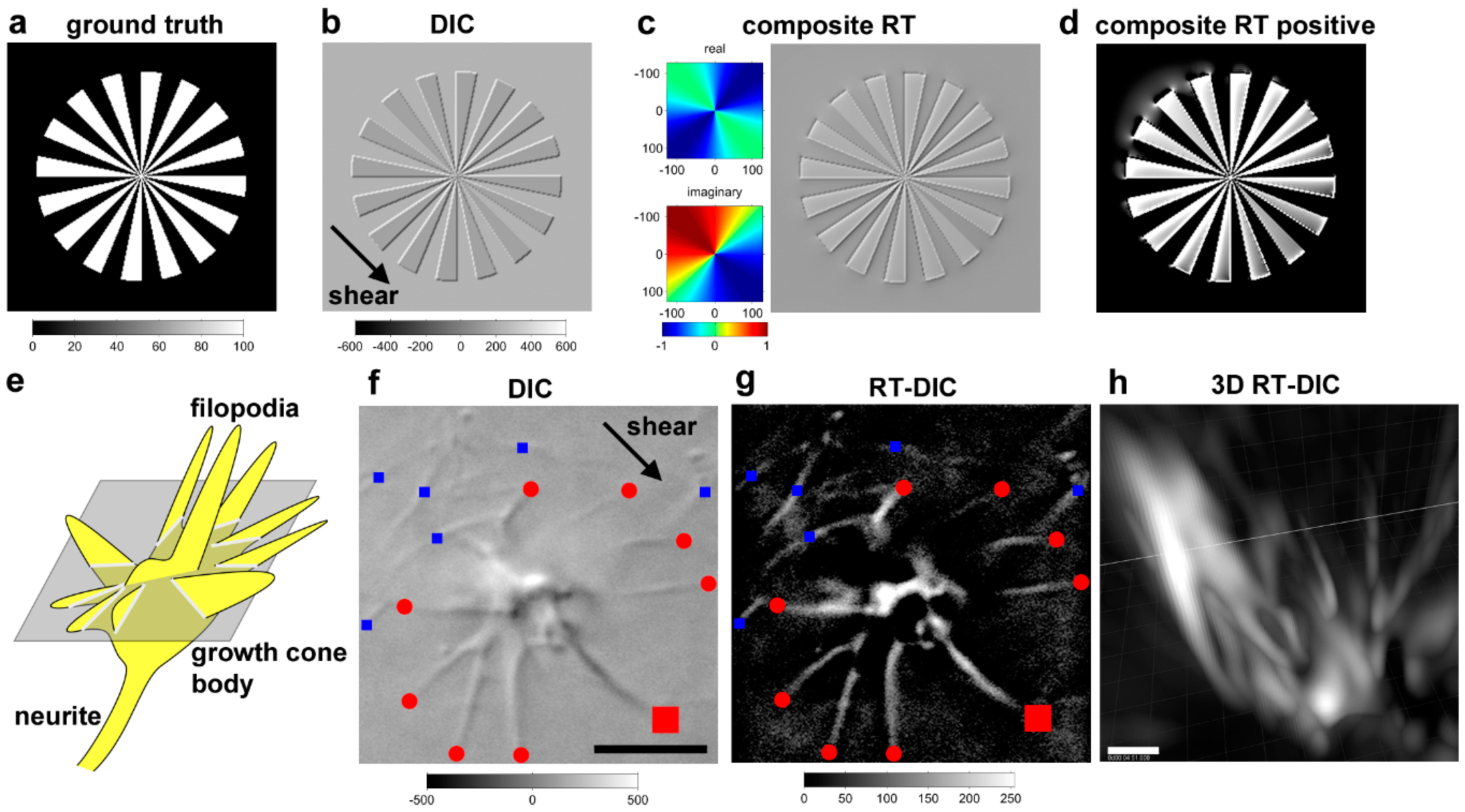
9.1. Riesz Transform-DIC Microscopy for Less-Toxic Label-Free 3D Time-Lapse Imaging
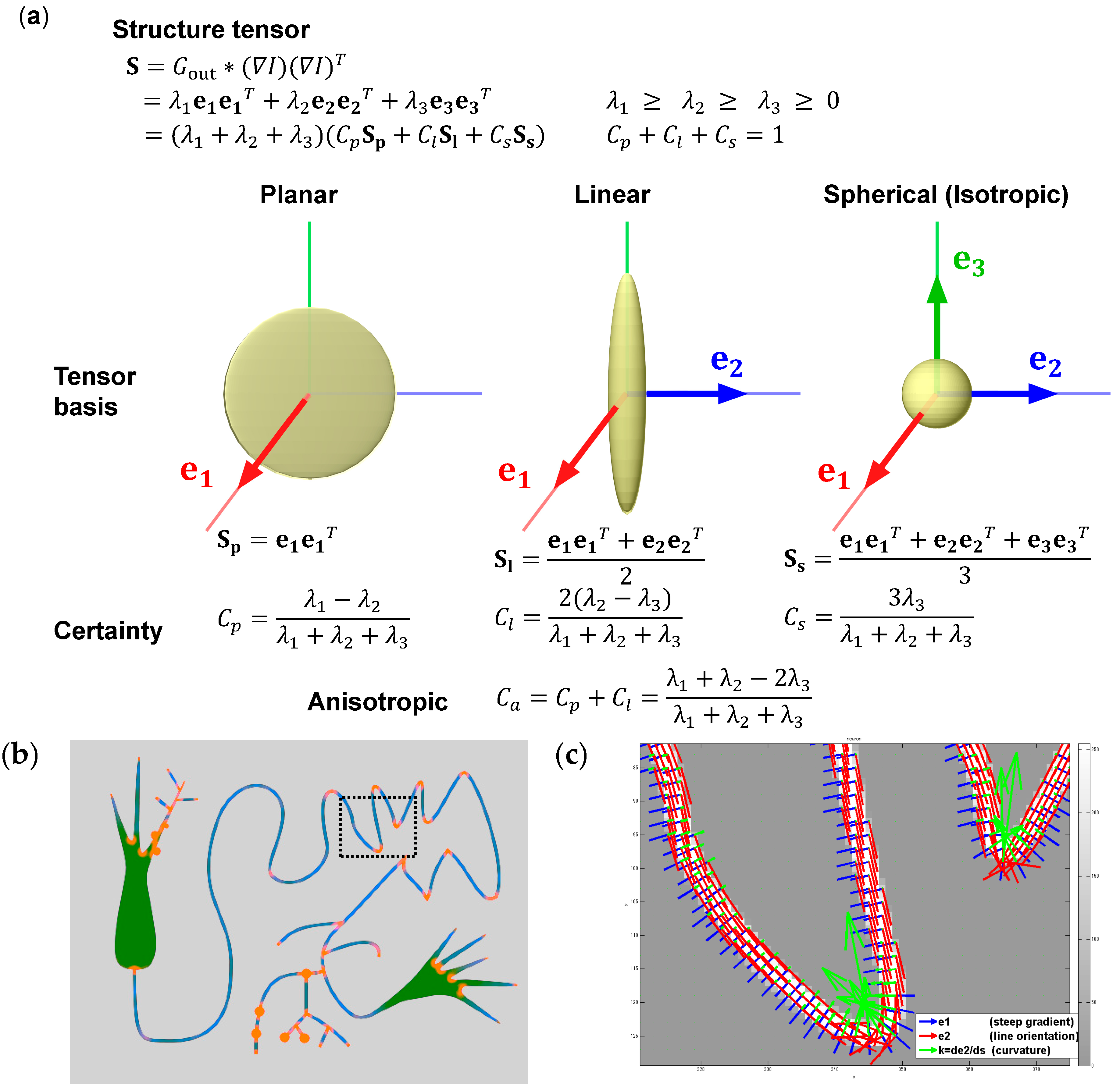
9.2. Structure Tensor for Analysis of 3D Morphology
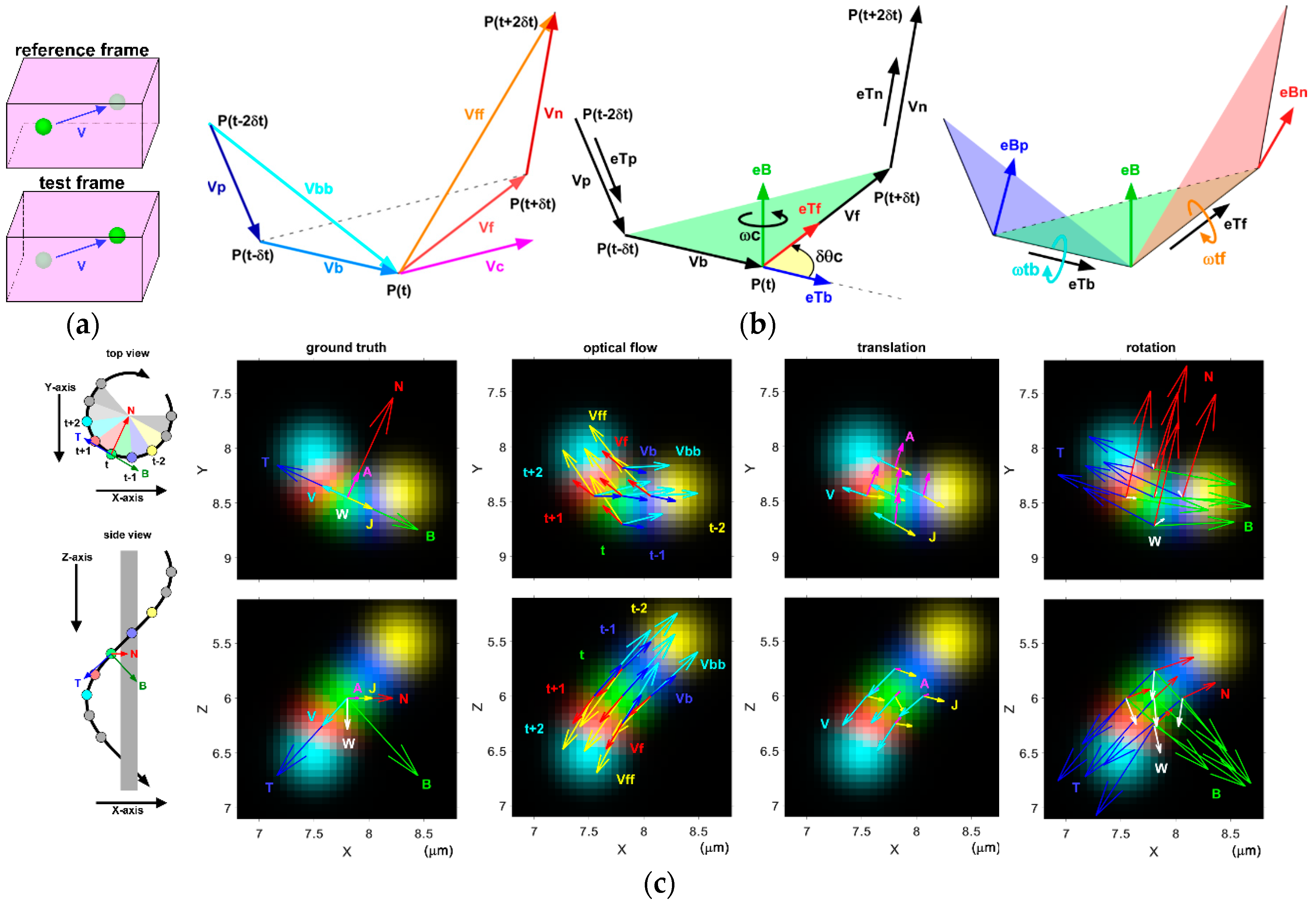
9.3. Optical Flow for Analysis of 3D Motility
10. Conclusions and Future Perspectives
Supplementary Materials
Funding
Conflicts of Interest
References
- Geschwind, N.; Galaburda, A.M. Cerebral Lateralization: Biological Mechanisms, Associations, and Pathology; MIT Press: Cambridge, MA, USA, 1987; 283p. [Google Scholar]
- Sun, T.; Walsh, C.A. Molecular approaches to brain asymmetry and handedness. Nat. Rev. Neurosci. 2006, 7, 655–662. [Google Scholar] [CrossRef]
- Toga, A.W.; Thompson, P.M. Mapping brain asymmetry. Nat. Rev. Neurosci. 2003, 4, 37–48. [Google Scholar] [CrossRef]
- Corballis, M.C. From mouth to hand: Gesture, speech, and the evolution of right-handedness. Behav. Brain Sci. 2003, 26, 199–208, discussion 208–260. [Google Scholar] [CrossRef] [PubMed]
- Hepper, P.G.; Shahidullah, S.; White, R. Handedness in the human fetus. Neuropsychologia 1991, 29, 1107–1111. [Google Scholar] [CrossRef]
- Hepper, P.G.; Wells, D.L.; Lynch, C. Prenatal thumb sucking is related to postnatal handedness. Neuropsychologia 2005, 43, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Broca, P. Sur le siège de la faculté du langage articulé (15 juin). Bulletins de la Société Anthropologque de Paris 1865, 6, 377–393. [Google Scholar] [CrossRef]
- Gazzaniga, M.S.; Bogen, J.E.; Sperry, R.W. Observations on visual perception after disconnexion of the cerebral hemispheres in man. Brain 1965, 88, 221–236. [Google Scholar] [CrossRef]
- Sperry, R. Some effects of disconnecting the cerebral hemispheres. Science 1982, 217, 1223–1226. [Google Scholar] [CrossRef]
- Gazzaniga, M.S. Forty-five years of split-brain research and still going strong. Nat. Rev. Neurosci. 2005, 6, 653–659. [Google Scholar] [CrossRef]
- Corbetta, M.; Shulman, G.L. Spatial neglect and attention networks. Annu. Rev. Neurosci. 2011, 34, 569–599. [Google Scholar] [CrossRef]
- Kleinman, J.T.; Newhart, M.; Davis, C.; Heidler-Gary, J.; Gottesman, R.F.; Hillis, A.E. Right hemispatial neglect: Frequency and characterization following acute left hemisphere stroke. Brain Cogn. 2007, 64, 50–59. [Google Scholar] [CrossRef]
- Navon, D. Forest before trees: The precedence of global features in visual perception. Cogn. Psychol. 1977, 9, 353–383. [Google Scholar] [CrossRef]
- Robertson, L.C.; Lamb, M.R.; Knight, R.T. Effects of lesions of temporal-parietal junction on perceptual and attentional processing in humans. J. Neurosci. Off. J. Soc. Neurosci. 1988, 8, 3757–3769. [Google Scholar] [CrossRef]
- Fink, G.R.; Halligan, P.W.; Marshall, J.C.; Frith, C.D.; Frackowiak, R.S.; Dolan, R.J. Where in the brain does visual attention select the forest and the trees? Nature 1996, 382, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Gainotti, G.; Marra, C. Differential contribution of right and left temporo-occipital and anterior temporal lesions to face recognition disorders. Front. Hum. Neurosci. 2011, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Galaburda, A.M.; LeMay, M.; Kemper, T.L.; Geschwind, N. Right-left asymmetrics in the brain. Science 1978, 199, 852–856. [Google Scholar] [CrossRef]
- Van Essen, D.C. A Population-Average, Landmark- and Surface-based (PALS) atlas of human cerebral cortex. Neuroimage 2005, 28, 635–662. [Google Scholar] [CrossRef] [PubMed]
- LeMay, M. Morphological cerebral asymmetries of modern man, fossil man, and nonhuman primate. Ann. N. Y. Acad. Sci. 1976, 280, 349–366. [Google Scholar] [CrossRef] [PubMed]
- Halpern, M.E.; Gunturkun, O.; Hopkins, W.D.; Rogers, L.J. Lateralization of the vertebrate brain: Taking the side of model systems. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 10351–10357. [Google Scholar] [CrossRef]
- Rogers, L.; Vallortigara, G. When and Why Did Brains Break Symmetry? Symmetry 2015, 7, 2181. [Google Scholar] [CrossRef]
- Rogers, L.J. A Matter of Degree: Strength of Brain Asymmetry and Behaviour. Symmetry 2017, 9, 57. [Google Scholar] [CrossRef]
- Gunturkun, O.; Ocklenburg, S. Ontogenesis of Lateralization. Neuron 2017, 94, 249–263. [Google Scholar] [CrossRef]
- Kawakami, R.; Shinohara, Y.; Kato, Y.; Sugiyama, H.; Shigemoto, R.; Ito, I. Asymmetrical allocation of NMDA receptor epsilon2 subunits in hippocampal circuitry. Science 2003, 300, 990–994. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, Y.; Hirase, H.; Watanabe, M.; Itakura, M.; Takahashi, M.; Shigemoto, R. Left-right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. Proc. Natl. Acad. Sci. USA 2008, 105, 19498–19503. [Google Scholar] [CrossRef] [PubMed]
- Andrew, R.J.; Rogers, L.J. The nature of lateralization in tetrapods. In Comparative Vertebrate Lateralization; Cambridge University Press: Cambridge, UK, 2002; pp. 94–125. [Google Scholar]
- Hopkins, W.D. Chimpanzee right-handedness: Internal and external validity in the assessment of hand use. Cortex 2006, 42, 90–93. [Google Scholar] [CrossRef]
- Rogers, L.J. Lateralization in its many forms, and its evolution and development. Evol. Hemisph. Spéc. Primates 2007, 5, 23. [Google Scholar]
- Bisazza, A.; Cantalupo, C.; Robins, A.; Rogers, L.J.; Vallortigara, G. Right-pawedness in toads. Nature 1996, 379, 408. [Google Scholar] [CrossRef]
- Robins, A.; Lippolis, G.; Bisazza, A.; Vallortigara, G.; Rogers, L.J. Lateralized agonistic responses and hindlimb use in toads. Anim. Behav. 1998, 56, 875–881. [Google Scholar] [CrossRef]
- Harris, L.J. Footedness in parrots: Three centuries of research, theory, and mere surmise. Can. J. Psychol. 1989, 43, 369–396. [Google Scholar] [CrossRef]
- Heffner, H.E.; Heffner, R.S. Temporal lobe lesions and perception of species-specific vocalizations by macaques. Science 1984, 226, 75–76. [Google Scholar] [CrossRef]
- Hauser, M.D.; Andersson, K. Left hemisphere dominance for processing vocalizations in adult, but not infant, rhesus monkeys: Field experiments. Proc. Natl. Acad. Sci. USA 1994, 91, 3946–3948. [Google Scholar] [CrossRef] [PubMed]
- Hook-Costigan, M.A.; Rogers, L.J. Lateralized use of the mouth in production of vocalizations by marmosets. Neuropsychologia 1998, 36, 1265–1273. [Google Scholar] [CrossRef]
- Nottebohm, F. Asymmetries in newral control of vocalization in the canary. In Lateralization in the Nervous System; Academic Press: Cambridge, MA, USA, 1977; pp. 23–44. [Google Scholar]
- Van Ruijssevelt, L.; Washington, S.D.; Hamaide, J.; Verhoye, M.; Keliris, G.A.; Van der Linden, A. Song Processing in the Zebra Finch Auditory Forebrain Reflects Asymmetric Sensitivity to Temporal and Spectral Structure. Front. Neurosci. 2017, 11, 549. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.; Koboroff, A.; Kaplan, G. Lateral Asymmetry of Brain and Behaviour in the Zebra Finch, Taeniopygia guttata. Symmetry 2018, 10, 679. [Google Scholar] [CrossRef]
- Ehret, G. Left hemisphere advantage in the mouse brain for recognizing ultrasonic communication calls. Nature 1987, 325, 249–251. [Google Scholar] [CrossRef] [PubMed]
- Fitch, R.H.; Brown, C.P.; O’Connor, K.; Tallal, P. Functional lateralization for auditory temporal processing in male and female rats. Behav. Neurosci. 1993, 107, 844–850. [Google Scholar] [CrossRef]
- Bauer, R.H. Lateralization of Neural Control for Vocalization by the Frog (Rana-Pipiens). Psychobiology 1993, 21, 243–248. [Google Scholar]
- Rogers, L.J. Development and function of lateralization in the avian brain. Brain Res. Bull. 2008, 76, 235–244. [Google Scholar] [CrossRef]
- Vallortigara, G.; Rogers, L.J.; Bisazza, A.; Lippolis, G.; Robins, A. Complementary right and left hemifield use for predatory and agonistic behaviour in toads. Neuroreport 1998, 9, 3341–3344. [Google Scholar] [CrossRef]
- Braitenberg, V.; Kemali, M. Exceptions to bilateral symmetry in the epithalamus of lower vertebrates. J. Comp. Neurol. 1970, 138, 137–146. [Google Scholar] [CrossRef]
- Concha, M.L.; Wilson, S.W. Asymmetry in the epithalamus of vertebrates. J. Anat. 2001, 199, 63–84. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, H.; Amo, R.; Okamoto, H. Phylogeny and ontogeny of the habenular structure. Front. Neurosci. 2011, 5, 138. [Google Scholar] [CrossRef] [PubMed]
- Concha, M.L.; Bianco, I.H.; Wilson, S.W. Encoding asymmetry within neural circuits. Nat. Rev. Neurosci. 2012, 13, 832–843. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, N.; Morimoto, K.; Tsubokawa, T.; Higashijima, S.; Okamoto, H.; Yoshihara, Y. From the olfactory bulb to higher brain centers: Genetic visualization of secondary olfactory pathways in zebrafish. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 4756–4767. [Google Scholar] [CrossRef] [PubMed]
- Frasnelli, E. Brain and behavioral lateralization in invertebrates. Front. Psychol. 2013, 4, 939. [Google Scholar] [CrossRef]
- Frasnelli, E.; Vallortigara, G.; Rogers, L.J. Left-right asymmetries of behaviour and nervous system in invertebrates. Neurosci. Biobehav. Rev. 2012, 36, 1273–1291. [Google Scholar] [CrossRef] [PubMed]
- Pascual, A.; Huang, K.L.; Neveu, J.; Preat, T. Neuroanatomy: Brain asymmetry and long-term memory. Nature 2004, 427, 605–606. [Google Scholar] [CrossRef]
- Hobert, O.; Johnston, R.J., Jr.; Chang, S. Left-right asymmetry in the nervous system: The Caenorhabditis elegans model. Nat. Rev. Neurosci. 2002, 3, 629–640. [Google Scholar] [CrossRef]
- Hobert, O. Development of left/right asymmetry in the Caenorhabditis elegans nervous system: From zygote to postmitotic neuron. Genesis 2014, 52, 528–543. [Google Scholar] [CrossRef]
- Afzelius, B.A. A human syndrome caused by immotile cilia. Science 1976, 193, 317–319. [Google Scholar] [CrossRef]
- Layton, W.M., Jr. Random determination of a developmental process: Reversal of normal visceral asymmetry in the mouse. J. Hered. 1976, 67, 336–338. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.A.; Wolpert, L. The development of handedness in left/right asymmetry. Development 1990, 109, 1–9. [Google Scholar] [PubMed]
- Levin, M.; Mercola, M. The compulsion of chirality: Toward an understanding of left-right asymmetry. Genes Dev. 1998, 12, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Tanaka, Y.; Okada, Y.; Takeda, S.; Harada, A.; Kanai, Y.; Kido, M.; Hirokawa, N. Randomization of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 1998, 95, 829–837. [Google Scholar] [CrossRef]
- Hirokawa, N.; Tanaka, Y.; Okada, Y.; Takeda, S. Nodal flow and the generation of left-right asymmetry. Cell 2006, 125, 33–45. [Google Scholar] [CrossRef]
- Blum, M.; Schweickert, A.; Vick, P.; Wright, C.V.; Danilchik, M.V. Symmetry breakage in the vertebrate embryo: When does it happen and how does it work? Dev. Biol. 2014, 393, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Yoshiba, S.; Watanabe, D.; Ikeuchi, S.; Goto, T.; Marshall, W.F.; Hamada, H. De novo formation of left-right asymmetry by posterior tilt of nodal cilia. PLoS Biol. 2005, 3, e268. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Shiratori, H.; Saijoh, Y.; Hamada, H. Determination of left-right patterning of the mouse embryo by artificial nodal flow. Nature 2002, 418, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Gray, R.S.; Roszko, I.; Solnica-Krezel, L. Planar cell polarity: Coordinating morphogenetic cell behaviors with embryonic polarity. Dev. Cell 2011, 21, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Tingler, M.; Kurz, S.; Maerker, M.; Ott, T.; Fuhl, F.; Schweickert, A.; LeBlanc-Straceski, J.M.; Noselli, S.; Blum, M. A Conserved Role of the Unconventional Myosin 1d in Laterality Determination. Curr. Biol. 2018, 28, 810–816. [Google Scholar] [CrossRef]
- Juan, T.; Géminard, C.; Coutelis, J.-B.; Cerezo, D.; Polès, S.; Noselli, S.; Fürthauer, M. Myosin1D is an evolutionarily conserved regulator of animal left–right asymmetry. Nat. Commun. 2018, 9, 1942. [Google Scholar] [CrossRef]
- Saydmohammed, M.; Yagi, H.; Calderon, M.; Clark, M.J.; Feinstein, T.; Sun, M.; Stolz, D.B.; Watkins, S.C.; Amack, J.D.; Lo, C.W.; et al. Vertebrate myosin 1d regulates left–right organizer morphogenesis and laterality. Nat. Commun. 2018, 9, 3381. [Google Scholar] [CrossRef]
- McDowell, G.; Rajadurai, S.; Levin, M. From cytoskeletal dynamics to organ asymmetry: A nonlinear, regulative pathway underlies left-right patterning. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed]
- Levin, M. Left-right asymmetry in embryonic development: A comprehensive review. Mech. Dev. 2005, 122, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Levin, M. A unified model for left-right asymmetry? Comparison and synthesis of molecular models of embryonic laterality. Dev. Biol. 2013, 379, 1–15. [Google Scholar] [CrossRef]
- Levin, M.; Johnson, R.L.; Stern, C.D.; Kuehn, M.; Tabin, C. A molecular pathway determining left-right asymmetry in chick embryogenesis. Cell 1995, 82, 803–814. [Google Scholar] [CrossRef]
- Namigai, E.K.; Kenny, N.J.; Shimeld, S.M. Right across the tree of life: The evolution of left-right asymmetry in the Bilateria. Genesis 2014, 52, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Hozumi, S.; Maeda, R.; Taniguchi, K.; Kanai, M.; Shirakabe, S.; Sasamura, T.; Speder, P.; Noselli, S.; Aigaki, T.; Murakami, R.; et al. An unconventional myosin in Drosophila reverses the default handedness in visceral organs. Nature 2006, 440, 798–802. [Google Scholar] [CrossRef]
- Speder, P.; Adam, G.; Noselli, S. Type ID unconventional myosin controls left-right asymmetry in Drosophila. Nature 2006, 440, 803–807. [Google Scholar] [CrossRef]
- Taniguchi, K.; Maeda, R.; Ando, T.; Okumura, T.; Nakazawa, N.; Hatori, R.; Nakamura, M.; Hozumi, S.; Fujiwara, H.; Matsuno, K. Chirality in planar cell shape contributes to left-right asymmetric epithelial morphogenesis. Science 2011, 333, 339–341. [Google Scholar] [CrossRef]
- Gonzalez-Morales, N.; Geminard, C.; Lebreton, G.; Cerezo, D.; Coutelis, J.B.; Noselli, S. The Atypical Cadherin Dachsous Controls Left-Right Asymmetry in Drosophila. Dev. Cell 2015, 33, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Lebreton, G.; Géminard, C.; Lapraz, F.; Pyrpassopoulos, S.; Cerezo, D.; Spéder, P.; Ostap, E.M.; Noselli, S. Molecular to organismal chirality is induced by the conserved myosin 1D. Science 2018, 362, 949–952. [Google Scholar] [CrossRef]
- Kennedy, D.N.; O’Craven, K.M.; Ticho, B.S.; Goldstein, A.M.; Makris, N.; Henson, J.W. Structural and functional brain asymmetries in human situs inversus totalis. Neurology 1999, 53, 1260–1265. [Google Scholar] [CrossRef]
- Ihara, A.; Hirata, M.; Fujimaki, N.; Goto, T.; Umekawa, Y.; Fujita, N.; Terazono, Y.; Matani, A.; Wei, Q.; Yoshimine, T.; et al. Neuroimaging study on brain asymmetries in situs inversus totalis. J. Neurol. Sci. 2010, 288, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Vingerhoets, G.; Li, X.; Hou, L.; Bogaert, S.; Verhelst, H.; Gerrits, R.; Siugzdaite, R.; Roberts, N. Brain structural and functional asymmetry in human situs inversus totalis. Brain Struct. Funct. 2018, 223, 1937–1952. [Google Scholar] [CrossRef] [PubMed]
- McManus, I.C.; Martin, N.; Stubbings, G.F.; Chung, E.M.; Mitchison, H.M. Handedness and situs inversus in primary ciliary dyskinesia. Proc. Biol. Sci. 2004, 271, 2579–2582. [Google Scholar] [CrossRef]
- Tanaka, S.; Kanzaki, R.; Yoshibayashi, M.; Kamiya, T.; Sugishita, M. Dichotic listening in patients with situs inversus: Brain asymmetry and situs asymmetry. Neuropsychologia 1999, 37, 869–874. [Google Scholar] [CrossRef]
- Vingerhoets, G.; Gerrits, R.; Bogaert, S. Atypical brain functional segregation is more frequent in situs inversus totalis. Cortex 2018, 106, 12–25. [Google Scholar] [CrossRef]
- McManus, C. Right Hand, Left Hand: The Origins of Asymmetry in Brains, Bodies, Atoms and Cultures; Harvard University Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Barth, K.A.; Miklosi, A.; Watkins, J.; Bianco, I.H.; Wilson, S.W.; Andrew, R.J. fsi zebrafish show concordant reversal of laterality of viscera, neuroanatomy, and a subset of behavioral responses. Curr. Boil. 2005, 15, 844–850. [Google Scholar] [CrossRef] [PubMed]
- McManus, C. Reversed bodies, reversed brains, and (some) reversed behaviors: Of zebrafish and men. Dev. Cell 2005, 8, 796–797. [Google Scholar] [CrossRef]
- Kawakami, R.; Dobi, A.; Shigemoto, R.; Ito, I. Right isomerism of the brain in inversus viscerum mutant mice. PLoS ONE 2008, 3, e1945. [Google Scholar] [CrossRef] [PubMed]
- Heacock, A.M.; Agranoff, B.W. Clockwise growth of neurites from retinal explants. Science 1977, 198, 64–66. [Google Scholar] [CrossRef]
- Xu, J.; Van Keymeulen, A.; Wakida, N.M.; Carlton, P.; Berns, M.W.; Bourne, H.R. Polarity reveals intrinsic cell chirality. Proc. Natl. Acad. Sci. USA 2007, 104, 9296–9300. [Google Scholar] [CrossRef]
- Tamada, A.; Kawase, S.; Murakami, F.; Kamiguchi, H. Autonomous right-screw rotation of growth cone filopodia drives neurite turning. J. Cell. Biol. 2010, 188, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Hsu, J.J.; Zhao, X.; Guo, C.; Wong, M.N.; Huang, Y.; Li, Z.; Garfinkel, A.; Ho, C.M.; Tintut, Y.; et al. Left-right symmetry breaking in tissue morphogenesis via cytoskeletal mechanics. Circ. Res. 2012, 110, 551–559. [Google Scholar] [CrossRef]
- Wan, L.Q.; Chin, A.S.; Worley, K.E.; Ray, P. Cell chirality: Emergence of asymmetry from cell culture. Philos. Trans. R. Soc.Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef]
- Wan, L.Q.; Ronaldson, K.; Park, M.; Taylor, G.; Zhang, Y.; Gimble, J.M.; Vunjak-Novakovic, G. Micropatterned mammalian cells exhibit phenotype-specific left-right asymmetry. Proc. Natl. Acad. Sci. USA 2011, 108, 12295–12300. [Google Scholar] [CrossRef]
- Naganathan, S.R.; Furthauer, S.; Nishikawa, M.; Julicher, F.; Grill, S.W. Active torque generation by the actomyosin cell cortex drives left-right symmetry breaking. eLife 2014, 3, e04165. [Google Scholar] [CrossRef]
- Sato, K.; Hiraiwa, T.; Maekawa, E.; Isomura, A.; Shibata, T.; Kuranaga, E. Left-right asymmetric cell intercalation drives directional collective cell movement in epithelial morphogenesis. Nat. Commun. 2015, 6, 10074. [Google Scholar] [CrossRef]
- Tee, Y.H.; Shemesh, T.; Thiagarajan, V.; Hariadi, R.F.; Anderson, K.L.; Page, C.; Volkmann, N.; Hanein, D.; Sivaramakrishnan, S.; Kozlov, M.M.; et al. Cellular chirality arising from the self-organization of the actin cytoskeleton. Nat. Cell. Biol. 2015, 17, 445–457. [Google Scholar] [CrossRef]
- Yamanaka, H.; Kondo, S. Rotating pigment cells exhibit an intrinsic chirality. Genes Cells 2015, 20, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Dimonte, A.; Adamatzky, A.; Erokhin, V.; Levin, M. On chirality of slime mould. Biosystems 2016, 140, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Inaki, M.; Liu, J.; Matsuno, K. Cell chirality: Its origin and roles in left-right asymmetric development. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed]
- Raymond, M.J., Jr.; Ray, P.; Kaur, G.; Singh, A.V.; Wan, L.Q. Cellular and Nuclear Alignment Analysis for Determining Epithelial Cell Chirality. Ann. Biomed. Eng. 2016, 44, 1475–1486. [Google Scholar] [CrossRef]
- Tamada, A.; Igarashi, M. Revealing chiral cell motility by 3D Riesz transform-differential interference contrast microscopy and computational kinematic analysis. Nat. Commun. 2017, 8, 2194. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.; Chin, A.S.; Worley, K.E.; Fan, J.; Kaur, G.; Wu, M.; Wan, L.Q. Intrinsic cellular chirality regulates left-right symmetry breaking during cardiac looping. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef]
- Chin, A.S.; Worley, K.E.; Ray, P.; Kaur, G.; Fan, J.; Wan, L.Q. Epithelial Cell Chirality Revealed by Three-Dimensional Spontaneous Rotation. Proc. Natl. Acad. Sci. USA 2018, 115, 12188–12193. [Google Scholar] [CrossRef]
- Inaki, M.; Sasamura, T.; Matsuno, K. Cell Chirality Drives Left-Right Asymmetric Morphogenesis. Front. Cell Dev. Biol. 2018, 6, 34. [Google Scholar] [CrossRef]
- Romijn, H.J.; Mud, M.T.; Wolters, P.S.; Corner, M.A. Neurite formation in dissociated cerebral cortex in vitro: Evidence for clockwise outgrowth and autotopic contacts. Brain Res. 1980, 192, 575–580. [Google Scholar] [CrossRef]
- Schwartz, M.; Agranoff, B.W. Outgrowth and maintenance of neurites from cultured goldfish retinal ganglion cells. Brain Res. 1981, 206, 331–343. [Google Scholar] [CrossRef]
- Shirasaki, R.; Tamada, A.; Katsumata, R.; Murakami, F. Guidance of cerebellofugal axons in the rat embryo: Directed growth toward the floor plate and subsequent elongation along the longitudinal axis. Neuron 1995, 14, 961–972. [Google Scholar] [CrossRef]
- Tamada, A.; Shirasaki, R.; Murakami, F. Floor plate chemoattracts crossed axons and chemorepels uncrossed axons in the vertebrate brain. Neuron 1995, 14, 1083–1093. [Google Scholar] [CrossRef][Green Version]
- Yamamoto, N.; Tamada, A.; Murakami, F. Wiring of the brain by a range of guidance cues. Prog. Neurobiol. 2002, 68, 393–407. [Google Scholar] [CrossRef]
- Landreth, G.E.; Agranoff, B.W. Explant culture of adult goldfish retina: Effect of prior optic nerve crush. Brain Res. 1976, 118, 299–303. [Google Scholar] [CrossRef][Green Version]
- Berg, J.S.; Powell, B.C.; Cheney, R.E. A millennial myosin census. Mol. Biol. Cell. 2001, 12, 780–794. [Google Scholar] [CrossRef]
- Cheney, R.E.; O’Shea, M.K.; Heuser, J.E.; Coelho, M.V.; Wolenski, J.S.; Espreafico, E.M.; Forscher, P.; Larson, R.E.; Mooseker, M.S. Brain myosin-V is a two-headed unconventional myosin with motor activity. Cell 1993, 75, 13–23. [Google Scholar] [CrossRef]
- Rodriguez, O.C.; Cheney, R.E. Human myosin-Vc is a novel class V myosin expressed in epithelial cells. J. Cell Sci. 2002, 115, 991–1004. [Google Scholar]
- Ali, M.Y.; Uemura, S.; Adachi, K.; Itoh, H.; Kinosita, K., Jr.; Ishiwata, S. Myosin V is a left-handed spiral motor on the right-handed actin helix. Nat. Struct. Biol. 2002, 9, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Wang, F.; Schmitz, S.; Xu, Y.; Xu, Q.; Molloy, J.E.; Veigel, C.; Sellers, J.R. Neck length and processivity of myosin V. J. Biol. Chem. 2003, 278, 29201–29207. [Google Scholar] [CrossRef]
- Sakamoto, T.; Yildez, A.; Selvin, P.R.; Sellers, J.R. Step-size is determined by neck length in myosin V. Biochemistry 2005, 44, 16203–16210. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.K.; Bridgman, P.C. Nerve growth cone lamellipodia contain two populations of actin filaments that differ in organization and polarity. J. Cell Biol. 1992, 119, 1219–1243. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.L.; Hammer, J.; Bridgman, P.C. Subcellular localization of myosin V in nerve growth cones and outgrowth from dilute-lethal neurons. J. Cell Sci. 1997, 110 Pt 4, 439–449. [Google Scholar]
- Mehta, A.D.; Rock, R.S.; Rief, M.; Spudich, J.A.; Mooseker, M.S.; Cheney, R.E. Myosin-V is a processive actin-based motor. Nature 1999, 400, 590–593. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Mabuchi, K.; Ikebe, R.; Ikebe, M. Mechanoenzymatic characterization of human myosin Vb. Biochemistry 2006, 45, 2729–2738. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Watanabe, T.M.; Sato, O.; Awata, J.; Homma, K.; Umeki, N.; Higuchi, H.; Ikebe, R.; Ikebe, M. Human myosin Vc is a low duty ratio nonprocessive motor. J. Boil. Chem. 2008, 283, 10581–10592. [Google Scholar] [CrossRef]
- Takagi, Y.; Yang, Y.; Fujiwara, I.; Jacobs, D.; Cheney, R.E.; Sellers, J.R.; Kovacs, M. Human myosin Vc is a low duty ratio, nonprocessive molecular motor. J. Boil. Chem. 2008, 283, 8527–8537. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.W.; Ross, L.S.; Parrett, T.; Easter, S.S., Jr. The development of a simple scaffold of axon tracts in the brain of the embryonic zebrafish, Brachydanio rerio. Development 1990, 108, 121–145. [Google Scholar] [PubMed]
- Easter, S.S., Jr.; Ross, L.S.; Frankfurter, A. Initial tract formation in the mouse brain. J. Neurosci. Off. J. Soc. Neurosci. 1993, 13, 285–299. [Google Scholar] [CrossRef]
- Mastick, G.S.; Easter, S.S., Jr. Initial organization of neurons and tracts in the embryonic mouse fore- and midbrain. Dev. Biol. 1996, 173, 79–94. [Google Scholar] [CrossRef]
- Murphy, D.B.; Davidson, M.W. Fundamentals of Light Microscopy and Electronic Imaging, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; 538p. [Google Scholar]
- Felsberg, M.; Sommer, G. The monogenic signal. IEEE Trans. Signal Process. 2001, 49, 3136–3144. [Google Scholar] [CrossRef]
- Larkin, K.G.; Bone, D.J.; Oldfield, M.A. Natural demodulation of two-dimensional fringe patterns. I. General background of the spiral phase quadrature transform. J. Opt. Soc. Am. A Opt. Image Sci. Vis. 2001, 18, 1862–1870. [Google Scholar] [CrossRef] [PubMed]
- Gabor, D. Theory of communication. Part 1: The analysis of information. J. Inst. Electr. Eng. Part III Radio Commun. Eng. 1946, 93, 429–441. [Google Scholar] [CrossRef]
- Bigün, J.; Granlund, G. Optimal orientation detection of linear symmetry. In Proceedings of the IEEE First International Conference on Computer Vision (ICCV), London, UK, 8–11 June 1987. [Google Scholar]
- Horn, B.K.; Schunck, B.G. Determining optical flow. Artif. Intell. 1981, 17, 185–203. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Left Hemisphere (Human) | Right Hemisphere (Human) |
| Right hand control (dominant in 90%) | Left hand control (dominant in 10%) |
| Language | Visuospatial functions |
| Aphasia | Left hemispatial neglect |
| Right allocentric neglect | Left egocentric neglect |
| Local feature perception | Global feature perception |
| Facial recognition | |
| Left Hemisphere (Non-Human Vertebrates) | Right Hemisphere (Non-Human Vertebrates) |
| Vocal communication | Social recognition |
| Routine behaviors | Emergency behaviors |
| Discrimination of categorized stimuli | Discrimination of geometric cues |
| Food searching | Detection and escape from predators |
| Prey capture | Aggressive behaviors |
| Local information processing | Global information processing |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamada, A. Chiral Neuronal Motility: The Missing Link between Molecular Chirality and Brain Asymmetry. Symmetry 2019, 11, 102. https://doi.org/10.3390/sym11010102
Tamada A. Chiral Neuronal Motility: The Missing Link between Molecular Chirality and Brain Asymmetry. Symmetry. 2019; 11(1):102. https://doi.org/10.3390/sym11010102
Chicago/Turabian StyleTamada, Atsushi. 2019. "Chiral Neuronal Motility: The Missing Link between Molecular Chirality and Brain Asymmetry" Symmetry 11, no. 1: 102. https://doi.org/10.3390/sym11010102
APA StyleTamada, A. (2019). Chiral Neuronal Motility: The Missing Link between Molecular Chirality and Brain Asymmetry. Symmetry, 11(1), 102. https://doi.org/10.3390/sym11010102