Fc Engineering Strategies to Advance IgA Antibodies as Therapeutic Agents


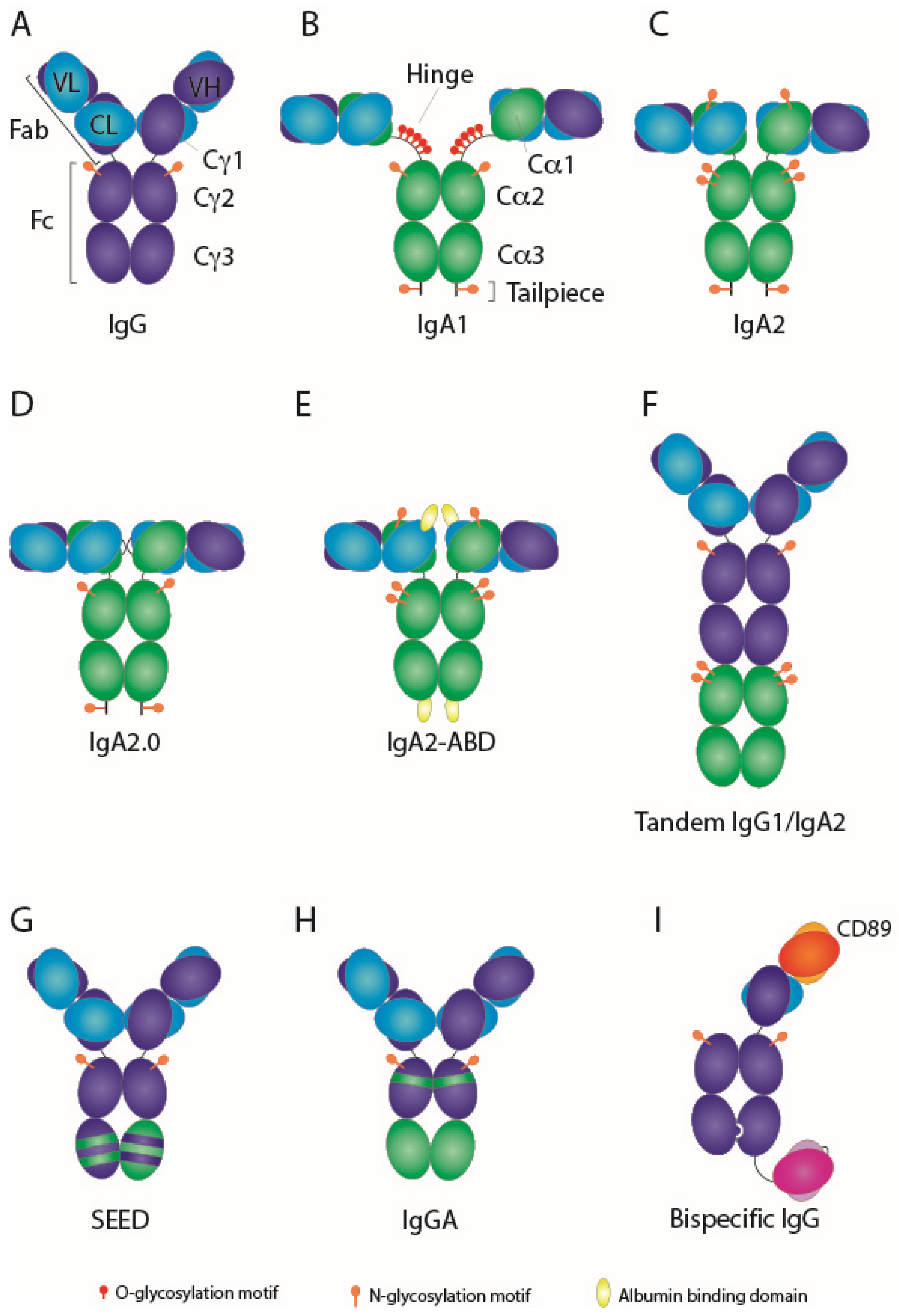{kind=link}
Abstract
1. Introduction: IgA and FcαRI
2. Enhancing IgA Developability
3. IgA Glyco-Engineering
4. FcRn-Based Half-Life Extension Strategies
5. IgA-IgG Hybrid Molecules
6. FcαRI Engagers as IgA Alternative
7. Perspective
Author Contributions
Funding
Conflicts of Interest
References
- Macpherson, A.J.; Gatto, D.; Sainsbury, E.; Harriman, G.R.; Hengartner, H.; Zinkernagel, R.M. A primitive T cell-independent mechanism of intestinal mucosal IgA responses to commensal bacteria. Science 2000, 288, 2222–2226. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Geuking, M.B.; McCoy, K.D. Homeland Security: IgA immunity at the frontiers of the body. Trends Immunol. 2012, 33, 160–167. [Google Scholar] [CrossRef]
- Chintalacharuvu, K.R.; Raines, M.; Morrison, S.L. Divergence of human alpha-chain constant region gene sequences. A novel recombinant alpha 2 gene. J. Immunol. 1994, 152, 5299–5304. [Google Scholar]
- Steffen, U.; Koeleman, C.A.; Sokolova, M.V.; Bang, H.; Kleyer, A.; Rech, J.; Unterweger, H.; Schicht, M.; Garreis, F.; Hahn, J.; et al. IgA subclasses have different effector functions associated with distinct glycosylation profiles. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, A.J.; Yilmaz, B.; Limenitakis, J.P.; Ganal-Vonarburg, S.C. IgA Function in Relation to the Intestinal Microbiota. Annu. Rev. Immunol. 2018, 36, 359–381. [Google Scholar] [CrossRef] [PubMed]
- Johansen, F.E.; Kaetzel, C.S. Regulation of the polymeric immunoglobulin receptor and IgA transport: New advances in environmental factors that stimulate pIgR expression and its role in mucosal immunity. Mucosal Immunol. 2011, 4, 598–602. [Google Scholar] [CrossRef]
- Brandtzaeg, P.; Prydz, H. Direct evidence for an integrated function of J chain and secretory component in epithelial transport of immunoglobulins. Nature 1984, 311, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Leong, K.W.; Ding, J.L. The unexplored roles of human serum IgA. DNA Cell Biol. 2014, 33, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Van Egmond, M.; Van Garderen, E.; Van Spriel, A.B.; Damen, C.A.; Van Amersfoort, E.S.; Van Zandbergen, G.; Van Hattum, J.; Kuiper, J.; Van De Winkel, J.G.J. FcαRI-positive liver Kupffer cells: Reappraisal of the function of immunoglobulin A in immunity. Nat. Med. 2000, 6, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, B.; Launay, P.; Kanamaru, Y.; Moura, I.C.; Pfirsch, S.; Ruffié, C.; Hénin, D.; Benhamou, M.; Pretolani, M.; Blank, U.; et al. Identification of FcαRI as an inhibitory receptor that controls inflammation: Dual role of FcRγ ITAM. Immunity 2005, 22, 31–42. [Google Scholar] [CrossRef]
- van Spriel, A.B.; Leusen, J.H.W.; Vilé, H.; van de Winkel, J.G.J. Mac-1 (CD11b/CD18) as Accessory Molecule for FcαR (CD89) Binding of IgA. J. Immunol. 2002, 169, 3831–3836. [Google Scholar] [CrossRef] [PubMed]
- Carayannopoulos, L.; Hexham, J.M.; Capra, J.D. Localization of the binding site for the monocyte immunoglobulin (Ig) A-Fc receptor (CD89) to the domain boundary between Cα2 and Cα3 in human IgA1. J. Exp. Med. 1996, 183, 1579–1586. [Google Scholar] [CrossRef]
- Geissmann, F.; Launay, P.; Pasquier, B.; Lepelletier, Y.; Leborgne, M.; Lehuen, A.; Brousse, N.; Monteiro, R.C. A Subset of Human Dendritic Cells Expresses IgA Fc Receptor (CD89), Which Mediates Internalization and Activation Upon Cross-Linking by IgA Complexes. J. Immunol. 2001, 166, 346–352. [Google Scholar] [CrossRef]
- Wines, B.D.; Sardjono, C.T.; Trist, H.M.; Lay, C.-S.; Hogarth, P.M. The Interaction of FcαRI with IgA and Its Implications for Ligand Binding by Immunoreceptors of the Leukocyte Receptor Cluster. J. Immunol. 2001, 166, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Ten Broeke, T.; Honing, H.; Brandsma, A.M.; Jacobino, S.; Bakema, J.E.; Kanters, D.; Van Der Linden, J.A.M.; Bracke, M.; Koenderman, L.; Leusen, J.H.W. FcαRI dynamics are regulated by GSK-3 and PKCζ during cytokine mediated inside-out signaling. Front. Immunol. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rossato, E.; Ben Mkaddem, S.; Kanamaru, Y.; Hurtado-Nedelec, M.; Hayem, G.; Descatoire, V.; Vonarburg, C.; Miescher, S.; Zuercher, A.W.; Monteiro, R.C. Reversal of arthritis by human monomeric IgA through the receptor-mediated SH2 domain-containing phosphatase 1 inhibitory pathway. Arthritis Rheumatol. 2015, 67, 1766–1777. [Google Scholar] [CrossRef] [PubMed]
- Valerius, T.; Stockmeyer, B.; van Spriel, A.B.; Graziano, R.F.; van den Herik-Oudijk, I.E.; Repp, R.; Deo, Y.M.; Lund, J.; Kalden, J.R.; Gramatzki, M.; et al. FcalphaRI (CD89) as a novel trigger molecule for bispecific antibody therapy. Blood 1997, 90, 4485–4492. [Google Scholar] [CrossRef]
- Dechant, M.; Beyer, T.; Schneider-Merck, T.; Weisner, W.; Peipp, M.; van de Winkel, J.G.J.; Valerius, T. Effector Mechanisms of Recombinant IgA Antibodies against Epidermal Growth Factor Receptor. J. Immunol. 2007, 179, 2936–2943. [Google Scholar] [CrossRef]
- Dechant, M.; Vidarsson, G.; Stockmeyer, B.; Repp, R.; Glennie, M.J.; Gramatzki, M.; Van de Winkel, J.G.J.; Valerius, T. Chimeric IgA antibodies against HLA class II effectively trigger lymphoma cell killing. Blood 2002, 100, 4574–4580. [Google Scholar] [CrossRef]
- Huls, G.; Heijnen, I.A.F.M.; Cuomo, E.; Van Der Linden, J.; Boel, E.; Van De Winkel, J.G.J.; Logtenberg, T. Antitumor immune effector mechanisms recruited by phage display-derived fully human IgG1 and IgA1 monoclonal antibodies. Cancer Res. 1999, 59, 5778–5784. [Google Scholar]
- Van Egmond, M.; Van Vuuren, A.J.H.; Morion, H.C.; Van Spriel, A.B.; Shen, L.; Hofhuis, F.M.A.; Saito, T.; Mayadas, T.N.; Verbeek, J.S.; Van De Winkel, J.G.J. Human immunoglobulin A receptor (FcαRI, CD89) function in transgenic mice requires both FCR γ chain and CR3 (CD11b/CD18). Blood 1999, 93, 4387–4394. [Google Scholar] [CrossRef] [PubMed]
- Launay, P.; Grossetête, B.; Arcos-Fajardo, M.; Gaudin, E.; Torres, S.P.; Beaudoin, L.; De Serre, N.P.M.; Lehuen, A.; Monteiro, R.C. Fcα receptor (CD89) mediates the development of immunoglobulin a (IgA) nephropathy (Berger’s disease): Evidence for pathogenic soluble receptor-IgA complexes in patients and CD89 transgenic mice. J. Exp. Med. 2000, 191, 1999–2009. [Google Scholar] [CrossRef] [PubMed]
- Boross, P.; Lohse, S.; Nederend, M.; Jansen, J.H.M.; van Tetering, G.; Dechant, M.; Peipp, M.; Royle, L.; Liew, L.P.; Boon, L.; et al. IgA EGFR antibodies mediate tumour killing in vivo. EMBO Mol. Med. 2013, 5, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Leusen, J.H.W. IgA as therapeutic antibody. Mol. Immunol. 2015, 68, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Logtenberg, M.E.W.; Jansen, J.H.M.; Raaben, M.; Toebes, M.; Franke, K.; Brandsma, A.M.; Matlung, H.L.; Fauster, A.; Gomez-Eerland, R.; Bakker, N.A.M.; et al. Glutaminyl cyclase is an enzymatic modifier of the CD47- SIRPα axis and a target for cancer immunotherapy. Nat. Med. 2019, 25, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Lohse, S.; Meyer, S.; Meulenbroek, L.A.P.M.; Jansen, J.H.M.; Nederend, M.; Kretschmer, A.; Klausz, K.; Möginger, U.; Derer, S.; Rösner, T.; et al. An anti-EGFR IgA that displays improved pharmacokinetics and myeloid effector cell engagement in vivo. Cancer Res. 2016, 76, 403–417. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Nederend, M.; Marco Jansen, J.H.; Reiding, K.R.; Jacobino, S.R.; Meeldijk, J.; Bovenschen, N.; Wuhrer, M.; Valerius, T.; Ubink, R.; et al. Improved in vivo anti-tumor effects of IgA-Her2 antibodies through half-life extension and serum exposure enhancement by FcRn targeting. MAbs 2016, 8, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Rouwendal, G.J.A.; van der Lee, M.M.; Meyer, S.; Reiding, K.R.; Schouten, J.; de Roo, G.; Egging, D.F.; Leusen, J.H.W.; Boross, P.; Wuhrer, M.; et al. A comparison of anti-HER2 IgA and IgG1 in vivo efficacy is facilitated by high N-glycan sialylation of the IgA. MAbs 2016, 8, 74–86. [Google Scholar] [CrossRef]
- Deo, Y.M.; Sundarapandiyan, K.; Keler, T.; Wallace, P.K.; Graziano, R.F. Bispecific molecules directed to the Fc receptor for IgA (Fc alpha RI, CD89) and tumor antigens efficiently promote cell-mediated cytotoxicity of tumor targets in whole blood. J. Immunol. 1998, 160, 1677–1686. [Google Scholar]
- Keler, T.; Wallace, P.K.; Vitale, L.A.; Russoniello, C.; Sundarapandiyan, K.; Graziano, R.F.; Deo, Y.M. Differential Effect of Cytokine Treatment on Fcα Receptor I- and Fcγ Receptor I-Mediated Tumor Cytotoxicity by Monocyte-Derived Macrophages. J. Immunol. 2000, 164, 5746–5752. [Google Scholar] [CrossRef]
- Brandsma, A.M.; Broeke, T.T.; Nederend, M.; Meulenbroek, L.A.P.M.; Van Tetering, G.; Meyer, S.; Jansen, J.H.M.; Buitrago, M.A.B.; Nagelkerke, S.Q.; Németh, I.; et al. Simultaneous targeting of FcgRs and FcaRI enhances tumor cell killing. Cancer Immunol. Res. 2015, 3, 1316–1324. [Google Scholar] [CrossRef] [PubMed]
- Brandsma, A.M.; Bondza, S.; Evers, M.; Koutstaal, R.; Nederend, M.; Marco Jansen, J.H.; Rösner, T.; Valerius, T.; Leusen, J.H.W.; Ten Broeke, T. Potent Fc receptor signaling by IgA leads to superior killing of cancer cells by neutrophils compared to IgG. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Roberts, J.T.; Barb, A.W. Multiple Variables at the Leukocyte Cell Surface Impact Fc γ Receptor-Dependent Mechanisms. Front. Immunol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Rosales, C. Fc γ Receptor Heterogeneity in Leukocyte Functional Responses. Front. Immunol. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Treffers, L.W.; Van Houdt, M.; Bruggeman, C.W.; Heineke, M.H.; Zhao, X.W.; Van Der Heijden, J.; Nagelkerke, S.Q.; Verkuijlen, P.J.J.H.; Geissler, J.; Lissenberg-Thunnissen, S.; et al. FcγRIIIb restricts antibody-dependent destruction of cancer cells by human neutrophils. Front. Immunol. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Peipp, M.; Van Bueren, J.J.L.; Schneider-Merck, T.; Bleeker, W.W.K.; Dechant, M.; Beyer, T.; Repp, R.; Van Berkel, P.H.C.; Vink, T.; Van De Winkel, J.G.J.; et al. Antibody fucosylation differentially impacts cytotoxicity mediated by NK and PMN effector cells. Blood 2008, 112, 2390–2399. [Google Scholar] [CrossRef]
- Hassler, J.R. IgA nephropathy: A brief review. Semin. Diagn. Pathol. 2020, 37, 143–147. [Google Scholar] [CrossRef]
- Woof, J.M.; Ken, M.A. The function of immunoglobulin A in immunity. J. Pathol. 2006, 208, 270–282. [Google Scholar] [CrossRef]
- Lohse, S.; Brunke, C.; Derer, S.; Peipp, M.; Boross, P.; Kellner, C.; Beyer, T.; Dechant, M.; Van Der Winkel, J.G.J.; Leusen, J.H.W.; et al. Characterization of a mutated IgA2 antibody of the m(1) allotype against the epidermal growth factor receptor for the recruitment of monocytes and macrophages. J. Biol. Chem. 2012, 287, 25139–25150. [Google Scholar] [CrossRef]
- Lua, W.H.; Ling, W.L.; Yeo, J.Y.; Poh, J.J.; Lane, D.P.; Gan, S.K.E. The effects of Antibody Engineering CH and CL in Trastuzumab and Pertuzumab recombinant models: Impact on antibody production and antigen-binding. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Evers, M.; Ten Broeke, T.; Jansen, J.H.M.; Nederend, M.; Hamdan, F.; Reiding, K.R.; Meyer, S.; Moerer, P.; Brinkman, I.; Rösner, T.; et al. Novel chimerized IgA CD20 antibodies: Improving neutrophil activation against CD20-positive malignancies. MAbs 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Pritsch, O.; Magnac, C.; Dumas, G.; Bouvet, J.P.; Alzari, P.; Dighiero, G. Can isotype switch modulate antigen-binding affinity and influence clonal selection? Eur. J. Immunol. 2000. [Google Scholar] [CrossRef]
- Tudor, D.; Yu, H.; Maupetit, J.; Drillet, A.S.; Bouceba, T.; Schwartz-Cornil, I.; Lopalco, L.; Tuffery, P.; Bomsel, M. Isotype modulates epitope specificity, affinity, and antiviral activities of anti-HIV-1 human broadly neutralizing 2F5 antibody. Proc. Natl. Acad. Sci. USA 2012. [Google Scholar] [CrossRef] [PubMed]
- Correa, A.; Trajtenberg, F.; Obal, G.; Pritsch, O.; Dighiero, G.; Oppezzo, P.; Buschiazzo, A. Structure of a human IgA1 Fab fragment at 1.55 Å resolution: Potential effect of the constant domains on antigen-affinity modulation. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013. [Google Scholar] [CrossRef] [PubMed]
- Grevys, A.; Bern, M.; Foss, S.; Bratlie, D.B.; Moen, A.; Gunnarsen, K.S.; Aase, A.; Michaelsen, T.E.; Sandlie, I.; Andersen, J.T. Fc Engineering of Human IgG1 for Altered Binding to the Neonatal Fc Receptor Affects Fc Effector Functions. J. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Posgai, M.T.; Tonddast-Navaei, S.; Jayasinghe, M.; Ibrahim, G.M.; Stan, G.; Herr, A.B. FcαRI binding at the IgA1 CH2–CH3 interface induces long-range conformational changes that are transmitted to the hinge region. Proc. Natl. Acad. Sci. USA 2018. [Google Scholar] [CrossRef] [PubMed]
- Gregory, R.L.; Rundegren, J.; Arnold, R.R. Separation of human IgA1 and IgA2 using jacalin-agarose chromatography. J. Immunol. Methods 1987, 99, 101–106. [Google Scholar] [CrossRef]
- Nilson, B.H.K.; Solomon, A.; Bjorck, L.; Akerstrom, B. Protein L from Peptostreptococcus magnus binds to the κ light chain variable domain. J. Biol. Chem. 1992, 267, 2234–2239. [Google Scholar]
- Boes, A.; Spiegel, H.; Delbrück, H.; Fischer, R.; Schillberg, S.; Sack, M. Affinity purification of a framework 1 engineered mouse/human chimeric IgA2 antibody from tobacco. Biotechnol. Bioeng. 2011, 108, 2804–2814. [Google Scholar] [CrossRef]
- Puligedda, R.D.; Vigdorovich, V.; Kouiavskaia, D.; Kattala, C.D.; Zhao, J.; Al-Saleem, F.H.; Chumakov, K.; Sather, D.N.; Dessain, S.K. Human IgA Monoclonal Antibodies That Neutralize Poliovirus, Produced by Hybridomas and Recombinant Expression. Antibodies 2020, 9, 5. [Google Scholar] [CrossRef]
- Beyer, T.; Lohse, S.; Berger, S.; Peipp, M.; Valerius, T.; Dechant, M. Serum-free production and purification of chimeric IgA antibodies. J. Immunol. Methods 2009, 346, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Jack Borrok, M.; Luheshi, N.M.; Beyaz, N.; Davies, G.C.; Legg, J.W.; Wu, H.; Dall’Acqua, W.F.; Tsui, P. Enhancement of antibody-dependent cell-mediated cytotoxicity by endowing igG with FcαRI (CD89) binding. MAbs 2015, 7, 743–751. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.H.; Aperlo, C.; Li, Y.; Kurosawa, E.; Lan, Y.; Lo, K.M.; Huston, J.S. SEEDbodies: Fusion proteins based on strand-exchange engineered domain (SEED) CH3 heterodimers in an Fc analogue platform for asymmetric binders or immunofusions and bispecific antibodies. Protein Eng. Des. Sel. 2010, 23, 195–202. [Google Scholar] [CrossRef]
- Kelton, W.; Mehta, N.; Charab, W.; Lee, J.; Lee, C.H.; Kojima, T.; Kang, T.H.; Georgiou, G. IgGA: A “cross-isotype” engineered human Fc antibody domain that displays both IgG-like and IgA-like effector functions. Chem. Biol. 2014, 21, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Xu, L.; Tao, F.; Xie, K.; Wu, Z.; Li, Y.; Li, J.; Chen, K.; Pi, C.; Mendelsohn, A.; et al. Simultaneous exposure to FcγR and FcaR on monocytes and macrophages enhances antitumor activity in vivo. Oncotarget 2017, 8, 39356–39366. [Google Scholar] [CrossRef] [PubMed]
- Brunke, C.; Lohse, S.; Derer, S.; Peipp, M.; Boross, P.; Kellner, C.; Beyer, T.; Dechant, M.; Royle, L.; Liew, L.P.; et al. Effect of a tail piece cysteine deletion on biochemical and functional properties of an epidermal growth factor receptor-directed IgA2 min(1) antibody. MAbs 2013, 5, 936–945. [Google Scholar] [CrossRef]
- Plomp, R.; De Haan, N.; Bondt, A.; Murli, J.; Dotz, V.; Wuhrer, M. Comparative glycomics of immunoglobulin A and G from saliva and plasma reveals biomarker potential. Front. Immunol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Atkin, J.D.; Pleass, R.J.; Owens, R.J.; Woof, J.M. Mutagenesis of the human IgA1 heavy chain tailpiece that prevents dimer assembly. J. Immunol. 1996, 157, 156–159. [Google Scholar]
- Blaese, R.M.; Strober, W.; Levy, A.L.; Waldmann, T.A. Hypercatabolism of IgG, IgA, IgM, and albumin in the Wiskott-Aldrich syndrome. A unique disorder of serum protein metabolism. J. Clin. Investig. 1971, 50, 2331–2338. [Google Scholar] [CrossRef]
- Challacombe, S.J.; Russell, M.W. Estimation of the intravascular half-lives of normal rhesus monkey IgG, IgA and IgM. Immunology 1979, 36, 331–338. [Google Scholar]
- Delacroix, D.L.; Elkon, K.B.; Geubel, A.P. Changes in size, subclass, and metabolic properties of serum immunoglobulin A in liver diseases and in other diseases with high serum immunoglobulin A. J. Clin. Investig. 1983, 71, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.T.; Foss, S.; Kenanova, V.E.; Olafsen, T.; Leikfoss, I.S.; Roopenian, D.C.; Wu, A.M.; Sandlie, I. Anti-carcinoembryonic antigen single-chain variable fragment antibody variants bind mouse and human neonatal Fc receptor with different affinities that reveal distinct cross-species differences in serum half-life. J. Biol. Chem. 2012, 287, 22927–22937. [Google Scholar] [CrossRef] [PubMed]
- Snoeck, V.; Peter, I.R.; Cox, E. The IgA system: A comparison of structure and function in different species. Vet. Res. 2006, 37, 455–467. [Google Scholar] [CrossRef]
- Li, F.; Ulrich, M.L.; Shih, V.F.S.; Cochran, J.H.; Hunter, J.H.; Westendorf, L.; Neale, J.; Benjamin, D.R. Mouse strains influence clearance and efficacy of antibody and antibody–drug conjugate via Fc–FcGR interaction. Mol. Cancer Ther. 2019, 18, 780–787. [Google Scholar] [CrossRef]
- Sharma, S.K.; Chow, A.; Monette, S.; Vivier, D.; Pourat, J.; Edwards, K.J.; Dilling, T.R.; Abdel-Atti, D.; Zeglis, B.M.; Poirier, J.T.; et al. Fc-mediated anomalous biodistribution of therapeutic antibodies in immunodeficient mouse models. Cancer Res. 2018, 78, 1820–1832. [Google Scholar] [CrossRef] [PubMed]
- Mattu, T.S.; Pleass, R.J.; Willis, A.C.; Kilian, M.; Wormald, M.R.; Lellouch, A.C.; Rudd, P.M.; Woof, J.M.; Dwek, R.A. The glycosylation and structure of human serum IgA1, Fab, and Fc regions and the role of N-glycosylation on Fcα receptor interactions. J. Biol. Chem. 1998, 273, 2260–2272. [Google Scholar] [CrossRef] [PubMed]
- Rifai, A.; Fadden, K.; Morrison, S.L.; Chintalacharuvu, K.R. The N-glycans determine the differential blood clearance and hepatic uptake of human immunoglobulin (Ig)A1 and IgA2 isotypes. J. Exp. Med. 2000, 191, 2171–2181. [Google Scholar] [CrossRef]
- Lee, S.J.; Evers, S.; Roeder, D.; Parlow, A.F.; Risteli, J.; Risteli, L.; Lee, Y.C.; Feizi, T.; Langen, H.; Nussenzweig, M.C. Mannose receptor-mediated regulation of serum glycoprotein homeostasis. Science 2002, 295, 1898–1901. [Google Scholar] [CrossRef]
- Heystek, H.C.; Moulon, C.; Woltman, A.M.; Garonne, P.; van Kooten, C. Human Immature Dendritic Cells Efficiently Bind and Take up Secretory IgA Without the Induction of Maturation. J. Immunol. 2002, 168, 102–107. [Google Scholar] [CrossRef]
- Zhong, X.; Ma, W.; Meade, C.L.; Tam, A.S.; Llewellyn, E.; Cornell, R.; Cote, K.; Scarcelli, J.J.; Marshall, J.K.; Tzvetkova, B.; et al. Transient CHO expression platform for robust antibody production and its enhanced N-glycan sialylation on therapeutic glycoproteins. Biotechnol. Prog. 2019, 35, 1–12. [Google Scholar] [CrossRef]
- Su, C.T.-T.; Lua, W.-H.; Ling, W.-L.; Gan, S.K.-E. Allosteric Effects between the Antibody Constant and Variable Regions: A Study of IgA Fc Mutations on Antigen Binding. Antibodies 2018, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Nussinov, R.; Ma, B. Antigen binding allosterically promotes Fc receptor recognition. MAbs 2019. [Google Scholar] [CrossRef]
- Lombana, T.N.; Rajan, S.; Zorn, J.A.; Mandikian, D.; Chen, E.C.; Estevez, A.; Yip, V.; Bravo, D.D.; Phung, W.; Farahi, F.; et al. Production, characterization, and in vivo half-life extension of polymeric IgA molecules in mice. MAbs 2019, 11, 1122–1138. [Google Scholar] [CrossRef]
- Dicker, M.; Tschofen, M.; Maresch, D.; König, J.; Juarez, P.; Orzaez, D.; Altmann, F.; Steinkellner, H.; Strasser, R. Transient glyco-engineering to produce recombinant IgA1 with defined N-and O-glycans in plants. Front. Plant Sci. 2016, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Göritzer, K.; Maresch, D.; Altmann, F.; Obinger, C.; Strasser, R. Exploring Site-Specific N-Glycosylation of HEK293 and Plant-Produced Human IgA Isotypes. J. Proteome Res. 2017, 16, 2560–2570. [Google Scholar] [CrossRef] [PubMed]
- Göritzer, K.; Turupcu, A.; Maresch, D.; Novak, J.; Altmann, F.; Oostenbrink, C.; Obinger, C.; Strasser, R. Distinct Fcα receptor N-glycans modulate the binding affinity to immunoglobulin A (IgA) antibodies. J. Biol. Chem. 2019, 294, 13995–14008. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.M.; Wall, S.B.; Takahashi, K.; Novak, J.; Renfrow, M.B.; Herr, A.B. Analysis of IgA1 N-glycosylation and its contribution to FcαRI binding. Biochemistry 2008, 47, 11285–11299. [Google Scholar] [CrossRef] [PubMed]
- Story, C.M.; Mikulska, J.E.; Simister, N.E. A major histocompatibility complex class I-like Fc receptor cloned from human placenta: Possible role in transfer of immunoglobulin G from mother to fetus. J. Exp. Med. 1994, 180, 2377–2381. [Google Scholar] [CrossRef]
- Firan, M.; Bawdon, R.; Radu, C.; Ober, R.J.; Eaken, D.; Antohe, F.; Ghetie, V.; Ward, E.S. The MHC class I-related receptor, FcRn, plays an essential role in the maternofetal transfer of γ-globulin in humans. Int. Immunol. 2001, 13, 993–1002. [Google Scholar] [CrossRef]
- Dickinson, B.L.; Badizadegan, K.; Wu, Z.; Ahouse, J.C.; Zhu, X.; Simister, N.E.; Blumberg, R.S.; Lencer, W.I. Bidirectional FcRn-dependent IgG transport in a polarized human intestinal epithelial cell line. J. Clin. Investig. 1999, 104, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Goebl, N.A.; Babbey, C.M.; Datta-Mannan, A.; Witcher, D.R.; Wroblewski, V.J.; Dunn, K.W. Neonatal Fc receptor mediates internalization of Fc in transfected human endothelial cells. Mol. Biol. Cell 2008, 19, 5490–5505. [Google Scholar] [CrossRef] [PubMed]
- Ward, E.S.; Ober, R.J. Targeting FcRn to Generate Antibody-Based Therapeutics. Trends Pharmacol. Sci. 2018, 39, 892–904. [Google Scholar] [CrossRef]
- Zhu, X.; Meng, G.; Dickinson, B.L.; Li, X.; Mizoguchi, E.; Miao, L.; Wang, Y.; Robert, C.; Wu, B.; Smith, P.D.; et al. MHC Class I-Related Neonatal Fc Receptor for IgG Is Functionally Expressed in Monocytes, Intestinal Macrophages, and Dendritic Cells. J. Immunol. 2001, 166, 3266–3276. [Google Scholar] [CrossRef] [PubMed]
- Junghans, R.P.; Anderson, C.L. The protection receptor for IgG catabolism is the β2-microglobulin-containing neonatal intestinal transport receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 5512–5516. [Google Scholar] [CrossRef] [PubMed]
- Israel, E.J.; Wilsker, D.F.; Hayes, K.C.; Schoenfeld, D.; Simister, N.E. Increased clearance of IgG in mice that lack β2-microglobulin: Possible protective role of FcRn. Immunology 1996, 89, 573–578. [Google Scholar] [CrossRef]
- Ghetie, V.; Hubbard, J.G.; Kim, J.K.; Tsen, M.F.; Lee, Y.; Ward, E.S. Abnormally short serum half-lives of IgG in β2-microglobulin-deficient mice. Eur. J. Immunol. 1996, 26, 690–696. [Google Scholar] [CrossRef] [PubMed]
- Muda, M.; Gross, A.W.; Dawson, J.P.; He, C.; Kurosawa, E.; Schweickhardt, R.; Dugas, M.; Soloviev, M.; Bernhardt, A.; Fischer, D.; et al. Therapeutic assessment of SEED: A new engineered antibody platform designed to generate mono- and bispecific antibodies. Protein Eng. Des. Sel. 2011, 24, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Kelton, C.; Wesolowski, J.S.; Soloviev, M.; Schweickhardt, R.; Fischer, D.; Kurosawa, E.; McKenna, S.D.; Gross, A.W. Anti-EGFR biparatopic-SEED antibody has enhanced combination-activity in a single molecule. Arch. Biochem. Biophys. 2012, 526, 219–225. [Google Scholar] [CrossRef]
- Chintalacharuvu, K.R.; Vuong, L.U.C.; Loi, L.A.; Larrick, J.W.; Morrison, S.L. Hybrid IgA2/IgG1 antibodies with tailor-made effector functions. Clin. Immunol. 2001, 101, 21–31. [Google Scholar] [CrossRef]
- Stockmeyer, B.; Dechant, M.; van Egmond, M.; Tutt, A.L.; Sundarapandiyan, K.; Graziano, R.F.; Repp, R.; Kalden, J.R.; Gramatzki, M.; Glennie, M.J.; et al. Triggering FCα-Receptor I (CD89) Recruits Neutrophils as Effector Cells for CD20-Directed Antibody Therapy. J. Immunol. 2000, 165, 5954–5961. [Google Scholar] [CrossRef]
- Li, B.; Xu, L.; Pi, C.; Yin, Y.; Xie, K.; Tao, F.; Li, R.; Gu, H.; Fang, J. CD89-mediated recruitment of macrophages via a bispecific antibody enhances anti-tumor efficacy. Oncoimmunology 2018, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Leusen, J.H.W.; Boross, P. Regulation of complement and modulation of its activity in mAb therapy of cancer. MAbs 2014, 6, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Geller, A.; Yan, J. The role of membrane bound complement regulatory proteins in tumor development and cancer immunotherapy. Front. Immunol. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, Q.; Xiao, W.; Zhao, Y.; Pi, J.; Xu, H.; Zhao, H.; Xu, J.; Evans, C.E.; Jin, H. Advances in Anti-Tumor Treatments Targeting the CD47/SIRPα Axis. Front. Immunol. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Hidalgo, A.; Chilvers, E.R.; Summers, C.; Koenderman, L. The Neutrophil Life Cycle. Trends Immunol. 2019, 40, 584–597. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Tetering, G.; Evers, M.; Chan, C.; Stip, M.; Leusen, J. Fc Engineering Strategies to Advance IgA Antibodies as Therapeutic Agents. Antibodies 2020, 9, 70. https://doi.org/10.3390/antib9040070
van Tetering G, Evers M, Chan C, Stip M, Leusen J. Fc Engineering Strategies to Advance IgA Antibodies as Therapeutic Agents. Antibodies. 2020; 9(4):70. https://doi.org/10.3390/antib9040070
Chicago/Turabian Stylevan Tetering, Geert, Mitchell Evers, Chilam Chan, Marjolein Stip, and Jeanette Leusen. 2020. "Fc Engineering Strategies to Advance IgA Antibodies as Therapeutic Agents" Antibodies 9, no. 4: 70. https://doi.org/10.3390/antib9040070
APA Stylevan Tetering, G., Evers, M., Chan, C., Stip, M., & Leusen, J. (2020). Fc Engineering Strategies to Advance IgA Antibodies as Therapeutic Agents. Antibodies, 9(4), 70. https://doi.org/10.3390/antib9040070