Engineering a Novel Antibody-Peptide Bispecific Fusion Protein Against MERS-CoV


 ,
,  ,
, 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Gene Construction
2.2. Protein Expression and Purification
2.3. MERS-CoV S Protein Binding
2.4. MERS-CoV Neutralization Assay
2.5. Cell-Cell Fusion Assay
3. Results
3.1. Generation of Anti-MERS-CoV Fusion Proteins
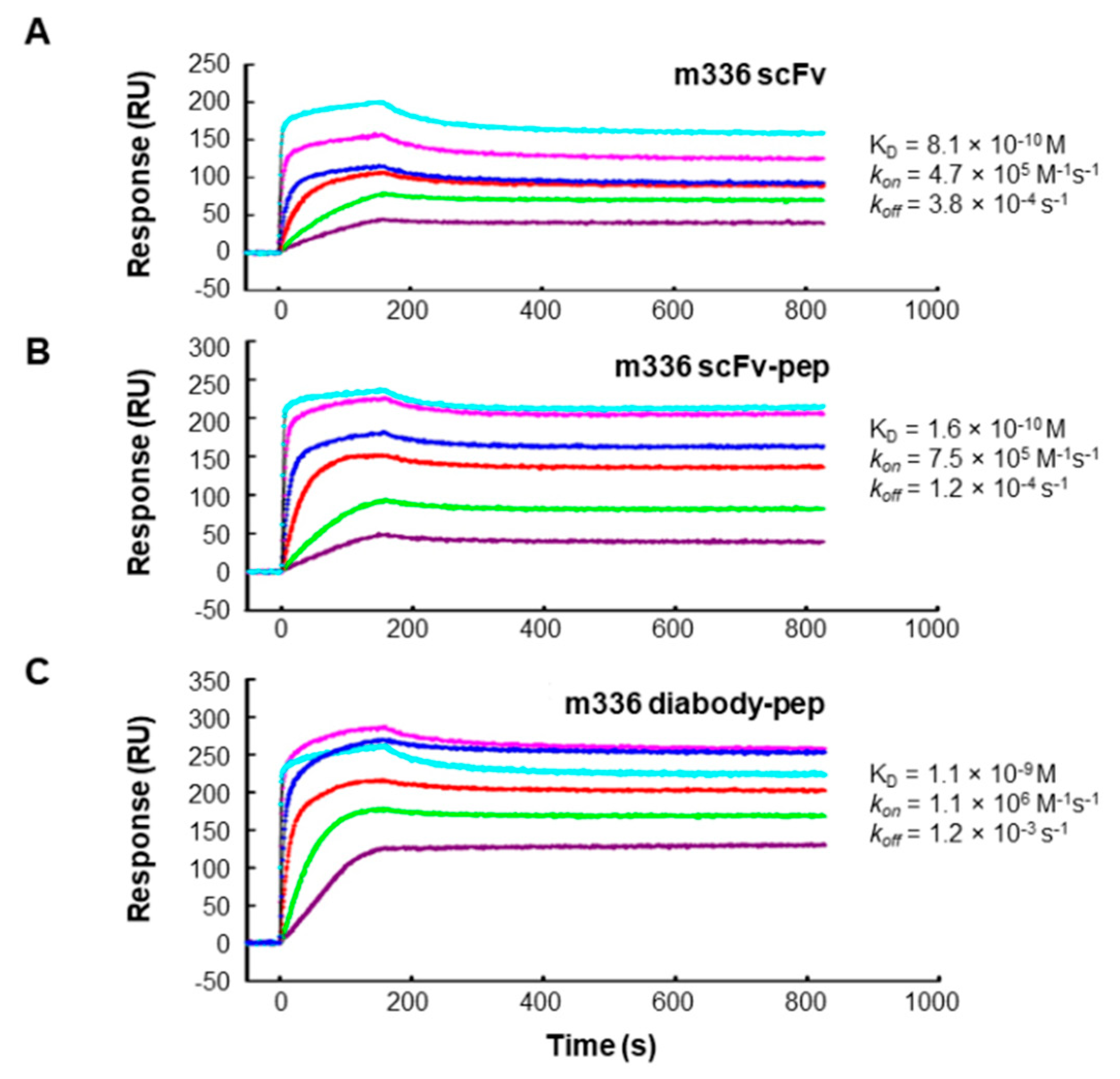
3.2. Interactions between Fusion Proteins and MERS-CoV S Protein
3.3. Neutralization of MERS-CoV Pseudovirus Infection
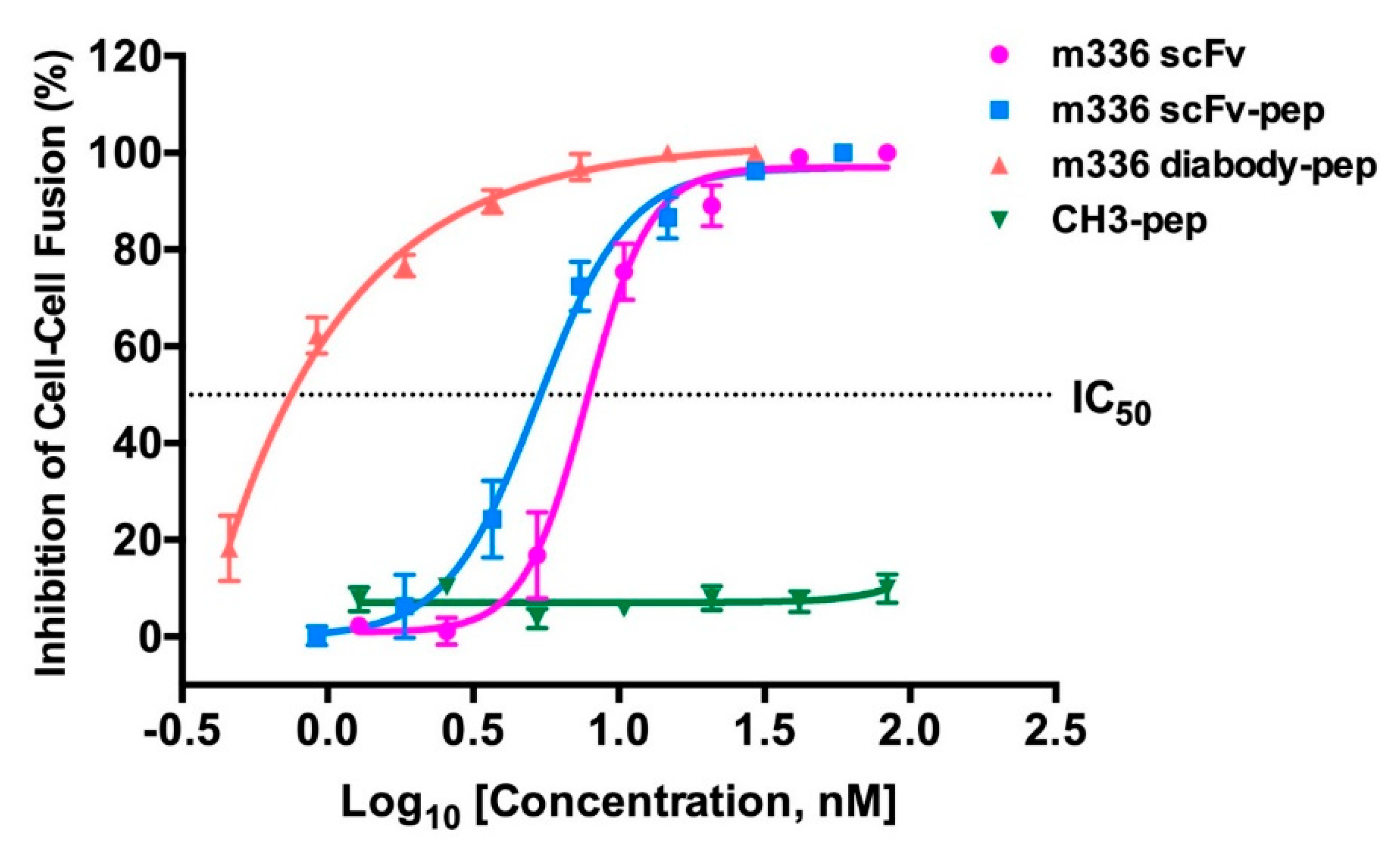
3.4. Inhibition of MERS S Protein-Mediated Cell-Cell Fusion
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhu, Z.; Dimitrov, A.S.; Bossart, K.N.; Crameri, G.; Bishop, K.A.; Choudhry, V.; Mungall, B.A.; Feng, Y.; Choudhary, A.; Zhang, M.; et al. Potent Neutralization of Hendra and Nipah Viruses by Human Monoclonal Antibodies. J. Virol. 2006, 80, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Bossart, K.N.; Geisbert, T.W.; Feldmann, H.; Zhu, Z.; Feldmann, F.; Geisbert, J.B.; Yan, L.; Feng, Y.; Brining, D.; Scott, D.P.; et al. A Neutralizing Human Monoclonal Antibody Protects African Green Monkeys from Hendra Virus Challenge. Sci. Transl. Med. 2011, 3, 105ra103. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Wong, G.; Audet, J.; Bello, A.; Fernando, L.; Alimonti, J.B.; Faustherbovendo, H.; Wei, H.; Aviles, J.; Hiatt, E.; et al. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 2014, 514, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhu, Z.; Li, S.; Deng, Y.; Wu, Y.; Zhang, N.; Puri, V.; Wang, C.; Zou, P.; Lei, C.; et al. A broadly neutralizing germline-like human monoclonal antibody against dengue virus envelope domain III. PLoS Pathog. 2019, 15, e1007836. [Google Scholar] [CrossRef]
- Brinkmann, U.; Kontermann, R.E. The making of bispecific antibodies. mAbs 2017, 9, 182–212. [Google Scholar] [CrossRef]
- Xu, T.; Ying, T.; Wang, L.; Zhang, X.D.; Wang, Y.; Kang, L.; Huang, T.; Cheng, L.; Wang, L.; Zhao, Q. A native-like bispecific antibody suppresses the inflammatory cytokine response by simultaneously neutralizing tumor necrosis factor-alpha and interleukin-17A. Oncotarget 2017, 8, 81860–81872. [Google Scholar] [CrossRef]
- Jin, Y.; Lei, C.; Hu, D.; Dimitrov, D.S.; Ying, T. Human monoclonal antibodies as candidate therapeutics against emerging viruses. Front. Med. 2017, 11, 462–470. [Google Scholar] [CrossRef]
- Bardhi, A.; Wu, Y.; Chen, W.; Li, W.; Zhu, Z.; Zheng, J.H.; Wong, H.; Jeng, E.; Jones, J.; Ochsenbauer, C.; et al. Potent In Vivo NK Cell-Mediated Elimination of HIV-1-Infected Cells Mobilized by a gp120-Bispecific and Hexavalent Broadly Neutralizing Fusion Protein. J. Virol. 2017, 91, e00937-17. [Google Scholar] [CrossRef]
- Kong, D.; Wang, Y.; Ji, P.; Li, W.; Ying, T.; Huang, J.; Wang, C.; Wu, Y.; Wang, Y.; Chen, W.; et al. A defucosylated bispecific multivalent molecule exhibits broad HIV-1-neutralizing activity and enhanced antibody-dependent cellular cytotoxicity against reactivated HIV-1 latently infected cells. AIDS 2018, 32, 1749–1761. [Google Scholar] [CrossRef]
- Wu, Y.; Xue, J.; Wang, C.; Li, W.; Wang, L.; Chen, W.; Prabakaran, P.; Kong, D.; Jin, Y.; Hu, D.; et al. Rapid elimination of broadly neutralizing antibodies correlates with treatment failure in the acute phase of SHIV infection. J. Virol. 2019, 93, e01077-19. [Google Scholar] [CrossRef]
- Pu, J.; Wang, Q.; Xu, W.; Lu, L.; Jiang, S. Development of Protein- and Peptide-Based HIV Entry Inhibitors Targeting gp120 or gp41. Viruses 2019, 11, 705. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.; Fouchier, R.A. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Chen, X.; Chughtai, A.A.; Dyda, A.; MacIntyre, C.R. Comparative epidemiology of Middle East respiratory syndrome coronavirus (MERS-CoV) in Saudi Arabia and South Korea. Emerg. Microbes Infect. 2017, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Lau, S.K.; To, K.K.; Cheng, V.C.; Woo, P.C.; Yuen, K.Y. Middle East respiratory syndrome coronavirus: Another zoonotic betacoronavirus causing SARS-like disease. Clin. Microbiol. Rev. 2015, 28, 465–522. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.W.; Li, K.S.M.; To, K.K.W.; Cheng, V.C.C.; Chen, H.; Yuen, K. Is the discovery of the novel human betacoronavirus 2c EMC/2012 (HCoV-EMC) the beginning of another SARS-like pandemic? J. Infect. 2012, 65, 477–489. [Google Scholar] [CrossRef]
- Alqahtani, A.S.; Wiley, K.E.; Tashani, M.; Heywood, A.E.; Willaby, H.W.; BinDhim, N.F.; Booy, R.; Rashid, H. Camel exposure and knowledge about MERS-CoV among Australian Hajj pilgrims in 2014. Virol. Sin. 2016, 31, 89–93. [Google Scholar] [CrossRef]
- Woo, P.C.Y.; Lau, S.K.P.; Huang, Y.; Yuen, K. Coronavirus Diversity, Phylogeny and Interspecies Jumping. Exp. Biol. Med. 2009, 234, 1117–1127. [Google Scholar] [CrossRef]
- Weiss, S.R.; Navas-Martin, S. Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef]
- Azhar, E.I.; El-Kafrawy, S.A.; Farraj, S.A.; Hassan, A.M.; Al-Saeed, M.S.; Hashem, A.M.; Madani, T.A. Evidence for camel-to-human transmission of MERS coronavirus. N. Engl. J. Med. 2014, 370, 2499–2505. [Google Scholar] [CrossRef]
- Vijaykrishna, D.; Smith, G.J.; Zhang, J.X.; Peiris, J.S.; Chen, H.; Guan, Y. Evolutionary insights into the ecology of coronaviruses. J. Virol. 2007, 81, 4012–4020. [Google Scholar] [CrossRef]
- Annan, A.; Baldwin, H.J.; Corman, V.M.; Klose, S.M.; Owusu, M.; Nkrumah, E.E.; Badu, E.K.; Anti, P.; Agbenyega, O.; Meyer, B.; et al. Human betacoronavirus 2c EMC/2012-related viruses in bats, Ghana and Europe. Emerg. Infect. Dis. 2013, 19, 456–459. [Google Scholar] [CrossRef] [PubMed]
- Reusken, C.; Haagmans, B.L.; Muller, M.A.; Gutierrez, C.; Godeke, G.; Meyer, B.; Muth, D.; Raj, V.S.; De Vries, L.S.; Corman, V.M.; et al. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. Lancet Infect. Dis. 2013, 13, 859–866. [Google Scholar] [CrossRef]
- Lau, S.K.; Wernery, R.; Wong, E.Y.; Joseph, S.; Tsang, A.K.; Patteril, N.A.; Elizabeth, S.K.; Chan, K.H.; Muhammed, R.; Kinne, J.; et al. Polyphyletic origin of MERS coronaviruses and isolation of a novel clade A strain from dromedary camels in the United Arab Emirates. Emerg. Microbes Infect. 2016, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Milne-Price, S.; Miazgowicz, K.L.; Munster, V.J. The emergence of the Middle East respiratory syndrome coronavirus. Pathog. Dis. 2014, 71, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Assiri, A.; Altawfiq, J.A.; Alrabeeah, A.A.; Alrabiah, F.; Alhajjar, S.; Albarrak, A.; Flemban, H.; Alnassir, W.N.; Balkhy, H.H.; Alhakeem, R.F.; et al. Epidemiological, demographic, and clinical characteristics of 47 cases of Middle East respiratory syndrome coronavirus disease from Saudi Arabia: A descriptive study. Lancet Infect. Dis. 2013, 13, 752–761. [Google Scholar] [CrossRef]
- Memish, Z.A.; Mishra, N.; Olival, K.J.; Fagbo, S.F.; Kapoor, V.; Epstein, J.H.; Alhakeem, R.; Durosinloun, A.; Al Asmari, M.; Islam, A.; et al. Middle East respiratory syndrome coronavirus in bats, Saudi Arabia. Emerg. Infect. Dis. 2013, 19, 1819–1823. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Xia, S.; Ying, T.; Jiang, S. Urgent development of effective therapeutic and prophylactic agents to control the emerging threat of Middle East respiratory syndrome (MERS). Emerg. Microbes Infect. 2015, 4, e37. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Eden, J.; Holmes, E.C.; Wang, L. Adaptive evolution of bat dipeptidyl peptidase 4 (dpp4): Implications for the origin and emergence of Middle East respiratory syndrome coronavirus. Virol. J. 2013, 10, 304. [Google Scholar] [CrossRef]
- Raj, V.S.; Mou, H.; Smits, S.L.; Dekkers, D.H.W.; Muller, M.A.; Dijkman, R.; Muth, D.; Demmers, J.; Zaki, A.M.; Fouchier, R.A.M.; et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 2013, 495, 251–254. [Google Scholar] [CrossRef]
- Ying, T.; Chen, W.; Feng, Y.; Wang, Y.; Gong, R.; Dimitrov, D.S. Engineered Soluble Monomeric IgG1 CH3 Domain: Generation, mechanisms of function, and implications for design of biological therapeutics*. J. Biol. Chem. 2013, 288, 25154–25164. [Google Scholar] [CrossRef]
- Boonacker, E.; Van Noorden, C.J.F. The multifunctional or moonlighting protein CD26/DPPIV. Eur. J. Cell Biol. 2003, 82, 53–73. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Lu, L.; Du, L.; Debnath, A.K. A predicted receptor-binding and critical neutralizing domain in S protein of the novel human coronavirus HCoV-EMC. J. Infect. 2013, 66, 464–466. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Lu, L.; Liu, Q.; Xu, W.; Du, L. Receptor-binding domains of spike proteins of emerging or re-emerging viruses as targets for development of antiviral vaccines. Emerg. Microbes Infect. 2012, 1, e13. [Google Scholar] [CrossRef] [PubMed]
- Van Doremalen, N.; Falzarano, D.; Ying, T.; de Wit, E.; Bushmaker, T.; Feldmann, F.; Okumura, A.; Wang, Y.; Scott, D.P.; Hanley, P.W.; et al. Efficacy of antibody-based therapies against Middle East respiratory syndrome coronavirus (MERS-CoV) in common marmosets. Antivir. Res. 2017, 143, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Ying, T.; Du, L.; Ju, T.W.; Prabakaran, P.; Lau, C.C.; Lu, L.; Liu, Q.; Wang, L.; Feng, Y.; Wang, Y.; et al. Exceptionally potent neutralization of Middle East respiratory syndrome coronavirus by human monoclonal antibodies. J. Virol. 2014, 88, 7796–7805. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.S.; Ying, T.; Tao, X.; Garron, T.; Algaissi, A.; Wang, Y.; Wang, L.; Peng, B.H.; Jiang, S.; Dimitrov, D.S.; et al. Passive transfer of a germline-like neutralizing human monoclonal antibody protects transgenic mice against lethal Middle East respiratory syndrome coronavirus infection. Sci. Rep. 2016, 6, 31629. [Google Scholar] [CrossRef]
- Houser, K.V.; Gretebeck, L.; Ying, T.; Wang, Y.; Vogel, L.; Lamirande, E.W.; Bock, K.W.; Moore, I.N.; Dimitrov, D.S.; Subbarao, K. Prophylaxis with a Middle East respiratory syndrome coronavirus (MERS-CoV)-specific human monoclonal antibody protects rabbits from MERS-CoV infection. J. Infect. Dis. 2016, 213, 1557–1561. [Google Scholar] [CrossRef]
- Ying, T.; Prabakaran, P.; Du, L.; Shi, W.; Feng, Y.; Wang, Y.; Wang, L.; Li, W.; Jiang, S.; Dimitrov, D.S.; et al. Junctional and allele-specific residues are critical for MERS-CoV neutralization by an exceptionally potent germline-like antibody. Nat. Commun. 2015, 6, 8223. [Google Scholar] [CrossRef]
- Channappanavar, R.; Lu, L.; Xia, S.; Du, L.; Meyerholz, D.K.; Perlman, S.; Jiang, S. Protective Effect of Intranasal Regimens Containing Peptidic Middle East Respiratory Syndrome Coronavirus Fusion Inhibitor Against MERS-CoV Infection. J. Infect. Dis. 2015, 212, 1894–1903. [Google Scholar] [CrossRef]
- Lu, L.; Liu, Q.; Zhu, Y.; Chan, K.H.; Qin, L.; Li, Y.; Wang, Q.; Chan, J.F.; Du, L.; Yu, F.; et al. Structure-based discovery of Middle East respiratory syndrome coronavirus fusion inhibitor. Nat. Commun. 2014, 5, 3067. [Google Scholar] [CrossRef]
- Wang, C.; Hua, C.; Xia, S.; Li, W.; Lu, L.; Jiang, S. Combining a Fusion Inhibitory Peptide Targeting the MERS-CoV S2 Protein HR1 Domain and a Neutralizing Antibody Specific for the S1 Protein Receptor-Binding Domain (RBD) Showed Potent Synergism against Pseudotyped MERS-CoV with or without Mutations in RBD. Viruses 2019, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Screening, Ranking, and Epitope Mapping of Anti-Human IL-9 Supernatants. Available online: http://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_5540.pdf (accessed on 28 October 2019).
- Du, L.; Zhao, G.; Yang, Y.; Qiu, H.; Wang, L.; Kou, Z.; Tao, X.; Yu, H.; Sun, S.; Tseng, C.T.K.; et al. A Conformation-Dependent Neutralizing Monoclonal Antibody Specifically Targeting Receptor-Binding Domain in Middle East Respiratory Syndrome Coronavirus Spike Protein. J. Virol. 2014, 88, 7045–7053. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Kou, Z.; Ma, C.; Tao, X.; Wang, L.; Zhao, G.; Chen, Y.; Yu, F.; Tseng, C.T.K.; Zhou, Y.; et al. A Truncated Receptor-Binding Domain of MERS-CoV Spike Protein Potently Inhibits MERS-CoV Infection and Induces Strong Neutralizing Antibody Responses: Implication for Developing Therapeutics and Vaccines. PLoS ONE 2013, 8, e81587. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Xu, W.; Wang, Q.; Wang, C.; Hua, C.; Li, W.; Lu, L.; Jiang, S. Peptide-Based Membrane Fusion Inhibitors Targeting HCoV-229E Spike Protein HR1 and HR2 Domains. Int. J. Mol. Sci. 2018, 19, 487. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Liu, Q.; Wang, Q.; Sun, Z.; Su, S.; Du, L.; Ying, T.; Lu, L.; Jiang, S. Middle East respiratory syndrome coronavirus (MERS-CoV) entry inhibitors targeting spike protein. Virus Res. 2014, 194, 200–210. [Google Scholar] [CrossRef]
- Ying, T.; Wang, Y.; Feng, Y.; Prabakaran, P.; Gong, R.; Wang, L.; Crowder, K.; Dimitrov, D.S. Engineered antibody domains with significantly increased transcytosis and half-life in macaques mediated by FcRn. mAbs 2015, 7, 922–930. [Google Scholar] [CrossRef]
- Ying, T.; Gong, R.; Ju, T.W.; Prabakaran, P.; Dimitrov, D.S. Engineered Fc based antibody domains and fragments as novel scaffolds. Biochim. Biophys. Acta 2014, 1844, 1977–1982. [Google Scholar] [CrossRef]
- Ying, T.; Feng, Y.; Wang, Y.; Chen, W.; Dimitrov, D.S. Monomeric IgG1 Fc molecules displaying unique Fc receptor interactions that are exploitable to treat inflammation-mediated diseases. mAbs 2014, 6, 1201–1210. [Google Scholar] [CrossRef]
- Yu, F.; Song, H.; Wu, Y.; Chang, S.Y.; Wang, L.; Li, W.; Hong, B.; Xia, S.; Wang, C.; Khurana, S.; et al. A potent germline-like human monoclonal antibody targets a pH-sensitive epitope on H7N9 influenza Hemagglutinin. Cell Host Microbe 2017, 22, 471–483. [Google Scholar] [CrossRef]
- Hong, B.; Wen, Y.; Ying, T. Recent Progress on Neutralizing Antibodies against Hepatitis B Virus and its Implications. Infect. Disord. Drug Targets 2019, 19, 213–223. [Google Scholar] [CrossRef]
- Wu, Y.; Jiang, S.; Ying, T. Single-Domain Antibodies as Therapeutics against Human Viral Diseases. Front. Immuno. 2017, 8, 1802. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, N.; Zuo, T.; Shi, X.; Poon, K.V.; Wu, Y.; Gao, F.; Li, D.; Wang, R.; Guo, J.; et al. Potent Neutralization of MERS-CoV by Human Neutralizing Monoclonal Antibodies to the Viral Spike Glycoprotein. Sci. Transl. Med. 2014, 6, 234ra59. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Agnihothram, S.; Jiao, Y.; Stanhope, J.; Graham, R.L.; Peterson, E.C.; Avnir, Y.; Tallarico, A.S.C.; Sheehan, J.; Zhu, Q.; et al. Identification of human neutralizing antibodies against MERS-CoV and their role in virus adaptive evolution. Proc. Natl. Acad. Sci. USA 2014, 111, E2018–E2026. [Google Scholar] [CrossRef]
- Cockrell, A.S.; Peck, K.M.; Yount, B.; Agnihothram, S.; Scobey, T.; Curnes, N.R.; Baric, R.S.; Heise, M.T. Mouse Dipeptidyl Peptidase 4 Is Not a Functional Receptor for Middle East Respiratory Syndrome Coronavirus Infection. J. Virol. 2014, 88, 5195–5199. [Google Scholar] [CrossRef] [PubMed]
- Van Doremalen, N.; Munster, V.J. Animal models of Middle East respiratory syndrome coronavirus infection. Antivir. Res. 2015, 122, 28–38. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fusion Proteins | kon (M−1s−1) | koff (s−1) | KD (M) |
|---|---|---|---|
| CH3-pep | 5.99 × 104 | 6.56 × 104 | 1.09 |
| m336 scFv | 4.66 × 105 | 3.80 × 10−4 | 8.14 × 10−10 |
| m336 scFv-pep | 7.47 × 105 | 1.20 × 10−4 | 1.60 × 10−10 |
| m336 diabody-pep | 1.14 × 106 | 1.23 × 10−3 | 1.08 × 10−9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Xu, J.; Kong, Y.; Liang, R.; Li, W.; Li, J.; Lu, J.; Dimitrov, D.S.; Yu, F.; Wu, Y.; et al. Engineering a Novel Antibody-Peptide Bispecific Fusion Protein Against MERS-CoV. Antibodies 2019, 8, 53. https://doi.org/10.3390/antib8040053
Wang L, Xu J, Kong Y, Liang R, Li W, Li J, Lu J, Dimitrov DS, Yu F, Wu Y, et al. Engineering a Novel Antibody-Peptide Bispecific Fusion Protein Against MERS-CoV. Antibodies. 2019; 8(4):53. https://doi.org/10.3390/antib8040053
Chicago/Turabian StyleWang, Lili, Jiyan Xu, Yu Kong, Ruiying Liang, Wei Li, Jinyao Li, Jun Lu, Dimiter S. Dimitrov, Fei Yu, Yanling Wu, and et al. 2019. "Engineering a Novel Antibody-Peptide Bispecific Fusion Protein Against MERS-CoV" Antibodies 8, no. 4: 53. https://doi.org/10.3390/antib8040053
APA StyleWang, L., Xu, J., Kong, Y., Liang, R., Li, W., Li, J., Lu, J., Dimitrov, D. S., Yu, F., Wu, Y., & Ying, T. (2019). Engineering a Novel Antibody-Peptide Bispecific Fusion Protein Against MERS-CoV. Antibodies, 8(4), 53. https://doi.org/10.3390/antib8040053