Antibodies and Derivatives Targeting DR4 and DR5 for Cancer Therapy



Abstract

{kind=link}
{kind=link}
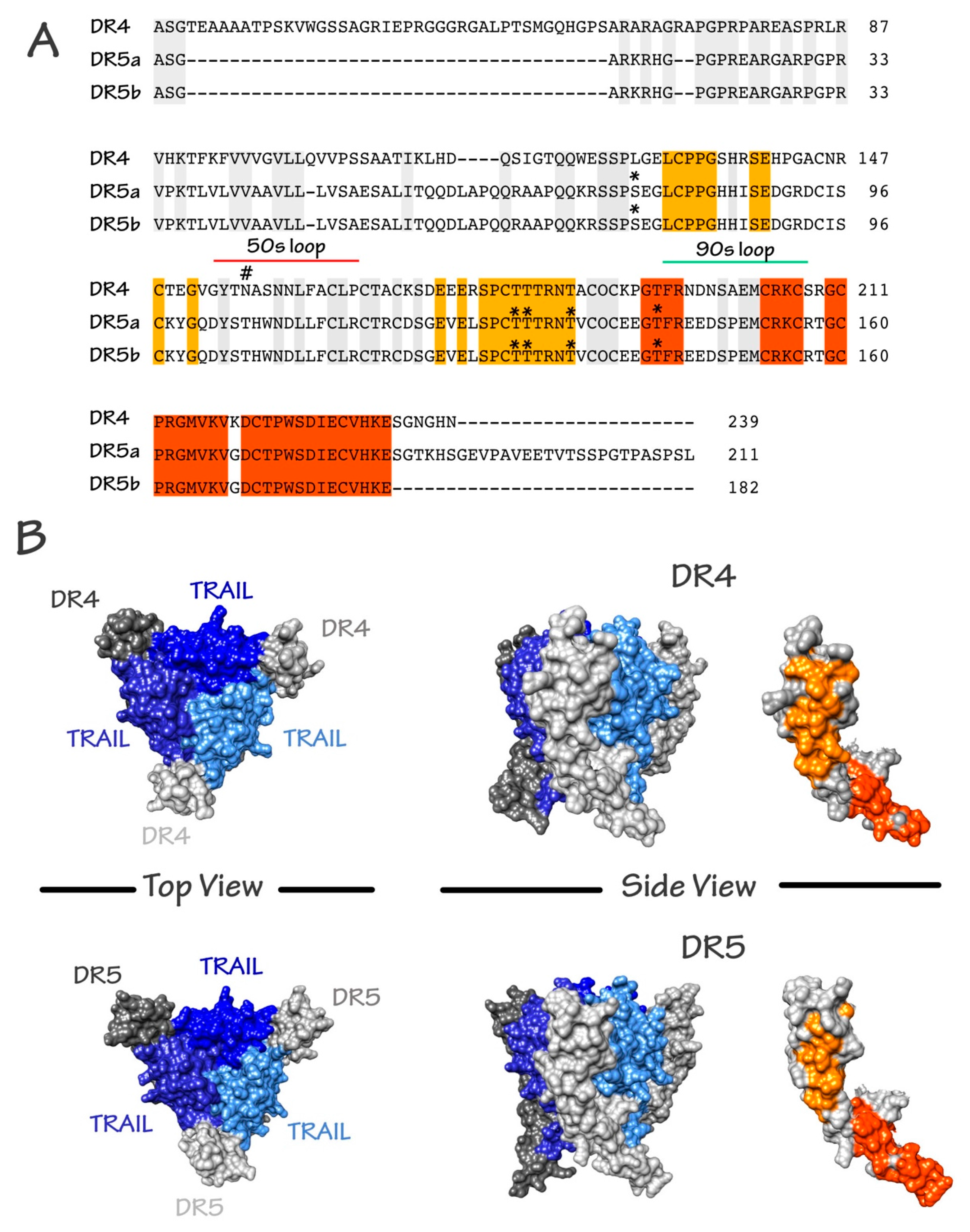{kind=link}
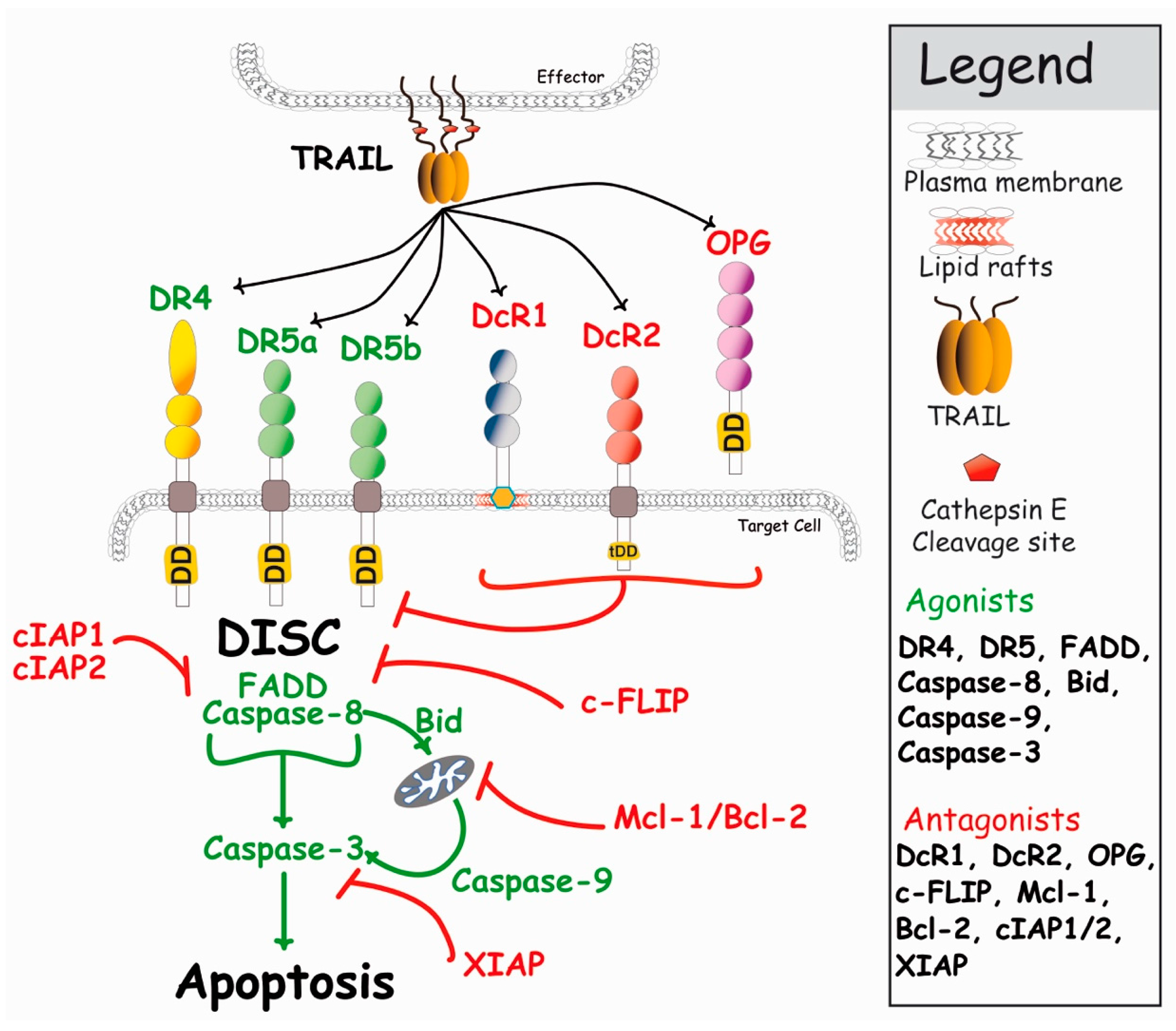{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
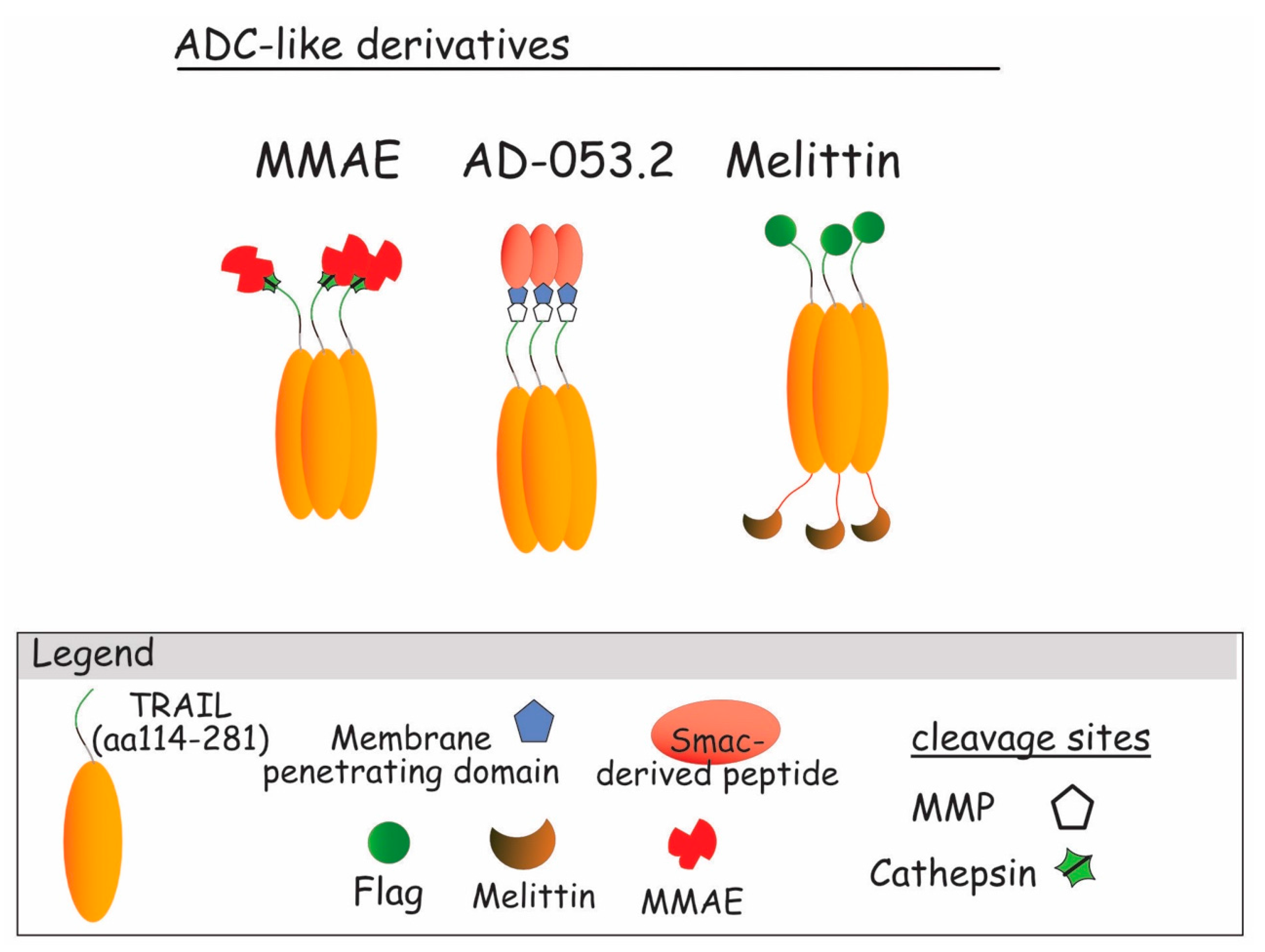{kind=link}
1. Introduction
2. Apoptosis Induced by TRAIL
3. TRAIL in Clinical Trials
3.1. First Attempts to Use TRAIL in Clinical Trials
3.2. Novel TRAIL Versions
4. First Antibodies in Clinical Trials
5. Novel TRAIL-Related Derivatives
5.1. Multivalent-Based Antibodies or Peptides
5.2. Bi- and Tri-Specific Derivatives
5.2.1. scFv-TRAIL
5.2.2. Unconventional and Bispecific Antibodies
5.3. Chimeric Antigen Receptors (CARs)
5.4. Antibody Drug Conjugate-Like TRAIL Derivatives (ADC-Like)
6. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| Aa | amino acids |
| ADC | antibody-drug conjugate |
| ADCC | antibody-dependant cell-mediated cytotoxicity |
| APO | apoptosis antigen |
| Bcl-2 | B-cell lymphoma 2, apoptosis inhibitor |
| BsAbs | bispecific antibodies |
| CARs | chimeric antibody receptors |
| CD | cluster of differentiation |
| CDC | complement-dependent cellular cytotoxicity |
| c-FLIP | cellular FLICE (FADD-like IL-1β-converting enzyme)-inhibitory protein |
| CPT-TRAIL | circularly permuted TRAIL |
| DC | dendritic cell |
| DcR | decoy receptor |
| DcR1 | decoy receptor 1, TRAIL-R3 |
| DcR2 | decoy receptor 2, TRAIL-R4 |
| DD | death domain |
| DISC | death-inducing signaling complex |
| DR | death receptor |
| DR4 | death receptor 4, TRAIL-R1 |
| DR5 | death receptor 5, TRAIL-R2 |
| Fab | fragment antigen-binding |
| Fas | fibroblast associated surface antigen |
| Fas-L | as ligand |
| Fc | crystalized fragment |
| FcR | crystalized fragment receptor |
| GPI | glycosylphosphatidylinisotol |
| KD | Kringle domain |
| Mcl-1 | myeloid cell leukemia-1 |
| moAbs | monoclonal antibodies |
| NK cell | natural killer cell |
| OPG | steoprotegerin, TRAIL-R5 |
| PBLs | eripheral blood cells |
| ROS | reactive oxygen species |
| rhTRAIL | human recombinant TRAIL |
| scFv | single chain fragment variable |
| sc-TRAIL | single chain TRAIL |
| sTRAIL | soluble TRAIL |
| TNC-TRAIL | tenascin-C-TRAIL |
| TNF | tumor necrosis factor |
| TNF-α | tumor necrosis factor alpha |
| TRAIL | TNF-related apoptosis-inducing ligand |
References
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed]
- DeVita, V.T., Jr.; Chu, E. A history of cancer chemotherapy. Cancer Res. 2008, 68, 8643–8653. [Google Scholar] [CrossRef] [PubMed]
- Carswell, E.A.; Old, L.J.; Kassel, R.L.; Green, S.; Fiore, N.; Williamson, B. An endotoxin-induced serum factor that causes necrosis of tumors. Proc. Natl. Acad. Sci. USA 1975, 72, 3666–3670. [Google Scholar] [CrossRef] [PubMed]
- Lejeune, F.J.; Ruegg, C.; Lienard, D. Clinical applications of TNF-alpha in cancer. Curr. Opin. Immunol. 1998, 10, 573–580. [Google Scholar] [CrossRef]
- Trauth, B.C.; Klas, C.; Peters, A.M.; Matzku, S.; Möller, P.; Falk, W.; Debatin, K.M.; Krammer, P.H. Monoclonal antibody-mediated tumor regression by induction of apoptosis. Science 1989, 245, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Yonehara, S.; Ishii, A.; Yonehara, M. A cell-killing monoclonal antibody (anti-Fas) to a cell surface antigen co-downregulated with the receptor of tumor necrosis factor. J. Exp. Med. 1989, 169, 1747–1756. [Google Scholar] [CrossRef] [PubMed]
- Wiley, S.R.; Schooley, K.; Smolak, P.J.; Din, W.S.; Huang, C.; Nicholl, J.K.; Sutherland, G.R.; Smith, T.D.; Rauch, C.; Smith, C.A.; et al. Identification and characterization of a new member of the TNF family that induces apoptosis. Immunity 1995, 3, 673–682. [Google Scholar] [CrossRef]
- Smith, C.A.; Farrah, T.; Goodwin, R.G. The TNF receptor superfamily of cellular and viral proteins: Activation, costimulation, and death. Cell 1994, 76, 959–962. [Google Scholar] [CrossRef]
- Pitti, R.M.; Marsters, S.A.; Ruppert, S.; Donahue, C.J.; Moore, A.; Ashkenazi, A. Induction of apoptosis by Apo-2 ligand, a new member of the TNF cytokine family. J. Biol. Chem. 1996, 271, 12687–12690. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.; Bodmer, J.-L.; Thome, M.; Hofmann, K.; Holler, N.; Tschopp, J. Characterization of two receptors for TRAIL. FEBS Lett. 1997, 416, 329–334. [Google Scholar] [CrossRef]
- MacFarlane, M.; Ahmad, M.; Srinivasula, S.M.; Fernandes-Alnemri, T.; Cohen, G.M.; Alnemri, E.S. Identification and molecular cloning of two novel receptors for the cytotoxic ligand TRAIL. J. Biol. Chem. 1997, 272, 25417–25420. [Google Scholar] [CrossRef] [PubMed]
- Marsters, S.A.; Sheridan, J.P.; Pitti, R.M.; Huang, A.; Skubatch, M.; Baldwin, D.; Yuan, J.; Gurney, A.; Goddard, A.D.; Godowski, P.; et al. A novel receptor for Apo2L/TRAIL contains a truncated death domain. Curr. Biol. 1997, 7, 1003–1006. [Google Scholar] [CrossRef]
- Walczak, H.; Degli-Esposti, M.A.; Johnson, R.S.; Smolak, P.J.; Waugh, J.Y.; Boiani, N.; Timour, M.S.; Gerhart, M.J.; Schooley, K.A.; Smith, C.A.; et al. TRAIL-R2: A novel apoptosis-mediating receptor for TRAIL. EMBO J. 1997, 16, 5386–5397. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, J.; Watanabe-Fukunaga, R.; Adachi, M.; Matsuzawa, A.; Kasugai, T.; Kitamura, Y.; Itoh, N.; Suda, T.; Nagata, S. Lethal effect of the anti-Fas antibody in mice. Nature 1993, 364, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Walczak, H.; Miller, R.E.; Ariail, K.; Gliniak, B.; Griffith, T.S.; Kubin, M.; Chin, W.; Jones, J.; Woodward, A.; Le, T.; et al. Tumoricidal activity of tumor necrosis factor-related apoptosis-inducing ligand in vivo. Nat. Med. 1999, 5, 157–163. [Google Scholar] [PubMed]
- Micheau, O.; Shirley, S.; Dufour, F. Death receptors as targets in cancer. Br. J. Pharmacol. 2013, 169, 1723–1744. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Pai, R.C.; Fong, S.; Leung, S.; Lawrence, D.A.; Marsters, S.A.; Blackie, C.; Chang, L.; McMurtrey, A.E.; Hebert, A.; et al. Safety and antitumor activity of recombinant soluble Apo2 ligand. J. Clin. Investig. 1999, 104, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; El-Deiry, W.S. TRAIL and apoptosis induction by TNF-family death receptors. Oncogene 2003, 22, 8628–8633. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.O.; Herndon, J.M.; Barreiro, R.; Griffith, T.S.; Ferguson, T.A. TRAIL: A mechanism of tumor surveillance in an immune privileged site. J. Immunol. 2002, 169, 4739–4744. [Google Scholar] [CrossRef] [PubMed]
- Smyth, M.J.; Takeda, K.; Hayakawa, Y.; Peschon, J.J.; van den Brink, M.R.M.; Yagita, H. Nature’s TRAIL—On a path to cancer immunotherapy. Immunity 2003, 18, 1–6. [Google Scholar] [CrossRef]
- Bodmer, J.L.; Schneider, P.; Tschopp, J. The molecular architecture of the TNF superfamily. Trends Biochem. Sci. 2002, 27, 19–26. [Google Scholar] [CrossRef]
- Dufour, F.; Rattier, T.; Shirley, S.; Picarda, G.; Constantinescu, A.A.; Morlé, A.; Zakaria, A.B.; Marcion, G.; Causse, S.; Szegezdi, E.; et al. N-glycosylation of mouse TRAIL-R and human TRAIL-R1 enhances TRAIL-induced death. Cell Death Differ. 2017, 24, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Shiraishi, T.; Horinaka, M.; Wakada, M.; Sakai, T. Glycosylation modulates TRAIL-R1/death receptor 4 protein: Different regulations of two pro-apoptotic receptors for TRAIL by tunicamycin. Oncol. Rep. 2007, 18, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.W.; Punnoose, E.A.; Januario, T.; Lawrence, D.A.; Pitti, R.M.; Lancaster, K.; Lee, D.; von Goetz, M.; Yee, S.F.; Totpa, K.; et al. Death-receptor O-glycosylation controls tumor-cell sensitivity to the proapoptotic ligand Apo2L/TRAIL. Nat. Med. 2007, 13, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Mirandola, P.; Ponti, C.; Gobbi, G.; Sponzilli, L.; Vaccarezza, M.; Cocco, L.; Zauli, G.; Secchiero, P.; Manzoli, F.M.; Vitale, M. Activated human NK and CD8+ T cells express both TNF-related apoptosis-inducing ligand (TRAIL) and TRAIL receptors but are resistant to TRAIL-mediated cytotoxicity. Blood 2004, 104, 2418–2424. [Google Scholar] [CrossRef] [PubMed]
- Emery, J.G.; McDonnell, P.; Burke, M.B.; Deen, K.C.; Lyn, S.; Silverman, C.; Dul, E.; Appelbaum, E.R.; Eichman, C.; DiPrinzio, R.; et al. Osteoprotegerin is a receptor for the cytotoxic ligand TRAIL. J. Biol. Chem. 1998, 273, 14363–14367. [Google Scholar] [CrossRef] [PubMed]
- Truneh, A.; Sharma, S.; Silverman, C.; Khandekar, S.; Reddy, M.P.; Deen, K.C.; Mclaughlin, M.M.; Srinivasula, S.M.; Livi, G.P.; Marshall, L.A.; et al. Temperature-sensitive differential affinity of TRAIL for its receptors. DR5 is the highest affinity receptor. J. Biol. Chem. 2000, 275, 23319–23325. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, H.N.; Ashkenazi, A. Apo2L/TRAIL and its death and decoy receptors. Cell Death Differ. 2003, 10, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Udagawa, N.; Takahashi, N. Action of RANKL and OPG for osteoclastogenesis. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Omran, O.M.; Ata, H.S. Expression of tumor necrosis factor-related apoptosis-inducing ligand death receptors DR4 and DR5 in human nonmelanoma skin cancer. Am. J. Dermatopathol. 2014, 36, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Sanlioglu, A.D.; Korcum, A.F.; Pestereli, E.; Erdogan, G.; Karaveli, S.; Savas, B.; Griffith, T.S.; Sanlioglu, S. TRAIL death receptor-4 expression positively correlates with the tumor grade in breast cancer patients with invasive ductal carcinoma. Int. J. Radiat. Oncol. Biol. Phys. 2007, 69, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Macher-Goeppinger, S.; Aulmann, S.; Tagscherer, K.E.; Wagener, N.; Haferkamp, A.; Penzel, R.; Brauckhoff, A.; Hohenfellner, M.; Sykora, J.; Walczak, H.; et al. Prognostic value of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) and TRAIL receptors in renal cell cancer. Clin. Cancer Res. 2009, 15, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Ganten, T.M.; Sykora, J.; Koschny, R.; Batke, E.; Aulmann, S.; Mansmann, U.; Stremmel, W.; Sinn, H.-P.; Walczak, H. Prognostic significance of tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) receptor expression in patients with breast cancer. J. Mol. Med. 2009, 87, 995–1007. [Google Scholar] [CrossRef] [PubMed]
- Van Noesel, M.M.; van Bezouw, S.; Salomons, G.S.; Voûte, P.A.; Pieters, R.; Baylin, S.B.; Herman, J.G.; Versteeg, R. Tumor-specific down-regulation of the tumor necrosis factor-related apoptosis-inducing ligand decoy receptors DcR1 and DcR2 is associated with dense promoter hypermethylation. Cancer Res. 2002, 62, 2157–2161. [Google Scholar] [PubMed]
- Gottwald, L.; Piekarski, J.; Kubiak, R.; Szwalski, J.; Pasz-Walczak, G.; Sęk, P.; Spych, M.; Suzin, J.; Tyliński, W.; Jeziorski, A. Membrane expression of TRAIL receptors DR4, DR5, DcR1 and DcR2 in the normal endometrium, atypical endometrial hyperplasia and endometrioid adenocarcinoma: A tissue microarray study. Arch. Gynecol. Obstet. 2013, 288, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Anees, M.; Hoak, P.; Schiefer, A.-I.; Vanhara, P.; EI-Gazzar, A.; Perco, P.; Kiesewette, B.; Mullauer, L.; Streubel, B.; Raderer, M.; et al. The potential evasion of immune surveillance in mucosa associated lymphoid tissue lymphoma by DcR2-mediated up-regulation of nuclear factor-kappaB. Leuk. Lymphoma 2015, 56, 1440–1449. [Google Scholar] [CrossRef] [PubMed]
- Micheau, O. Posttranslational modifications and death receptor Signalling. In TRAIL, Fas Ligand, TNF and TLR3 in Cancer; Micheau, O., Ed.; Springer: Cham, Switzerland, 2017; pp. 247–290. [Google Scholar]
- Bodmer, J.L.; Meier, P.; Tschopp, J.; Schneider, P. Cysteine 230 is essential for the structure and activity of the cytotoxic ligand TRAIL. J. Biol. Chem. 2000, 275, 20632–20637. [Google Scholar] [CrossRef] [PubMed]
- Belkahla, H.; Herlem, G.; Picaud, F.; Gharbi, T.; Hémadi, M.; Ammar, S.; Micheau, O. TRAIL-NP hybrids for cancer therapy: A review. Nanoscale 2017, 9, 5755–5768. [Google Scholar] [CrossRef] [PubMed]
- Gieffers, C.; Kluge, M.; Merz, C.; Sykora, J.; Thiemann, M.; Schaal, R.; Fischer, C.; Marcus, B.; Abhari, B.A.; Hohenberger, P.; Fulda, S.; et al. APG350 induces superior clustering of TRAIL receptors and shows therapeutic antitumor efficacy independent of cross-linking via Fcgamma receptors. Mol. Cancer Ther. 2013, 12, 2735–2747. [Google Scholar] [CrossRef] [PubMed]
- Shirley, S.; Morizot, A.; Micheau, O. Regulating TRAIL Receptor-Induced Cell Death at the Membrane: A Deadly Discussion. Recent Pat. Anti-Cancer Drug Discov. 2011, 6, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Mongkolsapaya, J.; Grimes, J.M.; Chen, N.; Xu, X.-N.; Stuart, D.I.; Jones, E.Y.; Screaton, G.R. Structure of the TRAIL-DR5 complex reveals mechanisms conferring specificity in apoptotic initiation. Nat. Struct. Biol. 1999, 6, 1048–1053. [Google Scholar] [PubMed]
- Ramamurthy, V.; Yamniuk, A.P.; Lawrence, E.J.; Yong, W.; Schneeweis, L.A.; Cheng, L.; Murdock, M.; Corbett, M.J.; Doyle, M.L.; Sheriff, S. The structure of the death receptor 4-TNF-related apoptosis-inducing ligand (DR4-TRAIL) complex. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Scaffidi, C.; Schmitz, I.; Zha, J.; Korsmeyer, S.J.; Krammer, P.H.; Peter, M.E. Differential modulation of apoptosis sensitivity in CD95 type I and type II cells. J. Biol. Chem. 1999, 274, 22532–22538. [Google Scholar] [CrossRef] [PubMed]
- Chinnaiyan, A.M. The apoptosome: Heart and soul of the cell death machine. Neoplasia 1999, 1, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Jacquemin, G.; Shirley, S.; Micheau, O. Combining naturally occurring polyphenols with TNF-related apoptosis-inducing ligand: A promising approach to kill resistant cancer cells? Cell. Mol. Life Sci. 2010, 67, 3115–3130. [Google Scholar] [CrossRef] [PubMed]
- Merino, D.; Lalaoui, N.; Morizot, A.; Schneider, P.; Solary, E.; Micheau, O. Differential inhibition of TRAIL-mediated DR5-DISC formation by decoy receptors 1 and 2. Mol. Cell. Biol. 2006, 26, 7046–7055. [Google Scholar] [CrossRef] [PubMed]
- Shirley, S.; Micheau, O. Targeting c-FLIP in cancer. Cancer Lett. 2013, 332, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Zamai, L.; Ahmad, M.; Bennett, I.M.; Azzoni, L.; Alnemri, E.S.; Perussia, B. Natural killer (NK) cell-mediated cytotoxicity: Differential use of TRAIL and Fas ligand by immature and mature primary human NK cells. J. Exp. Med. 1998, 188, 2375–2380. [Google Scholar] [CrossRef] [PubMed]
- Brincks, E.L.; Katewa, A.; Kucaba, T.A.; Griffith, S.T.; Legge, K.L. CD8 T cells utilize TRAIL to control influenza virus infection. J. Immunol. 2008, 181, 4918–4925. [Google Scholar] [CrossRef] [PubMed]
- Griffith, T.S.; Wiley, S.R.; Kubin, M.Z.; Sedger, L.M.; Maliszewski, C.R.; Fanger, N.A. Monocyte-mediated tumoricidal activity via the tumor necrosis factor-related cytokine, TRAIL. J. Exp. Med. 1999, 189, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Halaas, O.; Vik, R.; Ashkenazi, A.; Espevik, T. Lipopolysaccharide induces expression of APO2 ligand/TRAIL in human monocytes and macrophages. Scand. J. Immunol. 2000, 51, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.; Thome, M.; Burns, K.; Bodmer, J.-L.; Hofmann, K.; Kataoka, T.; Holler, N.; Tschopp, J. TRAIL receptors 1 (DR4) and 2 (DR5) signal FADD-dependent apoptosis and activate NF-kappaB. Immunity 1997, 7, 831–836. [Google Scholar] [CrossRef]
- Holler, N.; Tardivel, A.; Kovacsovics-Bankowski, M.; Hertig, S.; Gaide, O.; Martinon, F.; Tinel, A.; Deperthes, D.; Calderara, S.; Schulthess, T.; et al. Two adjacent trimeric Fas ligands are required for Fas signaling and formation of a death-inducing signaling complex. Mol. Cell. Biol. 2003, 23, 1428–1440. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, K.; Kwagh, J.G.; Dicker, D.T.; Herlyn, M.; Rustgi, A.K.; Chen, Y.; El-Deiry, W.S. Death induction by recombinant native TRAIL and its prevention by a caspase 9 inhibitor in primary human esophageal epithelial cells. J. Biol. Chem. 2004, 279, 40044–40052. [Google Scholar] [CrossRef] [PubMed]
- Dufour, F.; Rattier, T.; Constantinescu, A.A.; Zischler, L.; Morlé, A.; Mabrouk, H.B.; Humblin, E.; Jacquemin, G.; Szegezdi, E.; Delacote, F.; et al. TRAIL receptor gene editing unveils TRAIL-R1 as a master player of apoptosis induced by TRAIL and ER stress. Oncotarget 2017, 8, 9974–9985. [Google Scholar] [PubMed]
- Sun, A.Y.; Shen, Y.L.; Yin, J.C.; Zhang, H.; Tang, Y.N.; Wei, D.Z. Improvement of expression level and bioactivity of soluble tumor necrosis factor-related apoptosis-inducing ligand (Apo2L/TRAIL) by a novel zinc ion feeding strategy. Biotechnol. Lett. 2006, 28, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Krippner-Heidenreich, A.; Grunwald, I.; Zimmermann, G.; Kühnle, M.; Gerspach, J.; Sterns, T.; Shnyder, D.; Gill, J.H.; Männel, D.N.; Pfizenmaier, K.; et al. Single-chain TNF, a TNF derivative with enhanced stability and antitumoral activity. J. Immunol. 2008, 180, 8176–8183. [Google Scholar] [CrossRef] [PubMed]
- Schneider, B.; Münkel, S.; Krippner-Heidenreich, A.; Grunwald, I.; Wels, W.S.; Wajant, H.; Pfizenmaier, K.; Gerspach, J. Potent antitumoral activity of TRAIL through generation of tumor-targeted single-chain fusion proteins. Cell Death Dis. 2010, 1, e68. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Qiu, L.; Hou, J.; Zhao, Y.; Pan, L.; Yang, S.; Leng, Y.; Xi, H.; Zhang, X.; Cui, J.; Wei, N. Recombinant Circularly Permuted TRAIL (CPT) for the Treatment of Relapsed or Refractory Multiple Myeloma: An Open-Label, Multicenter Phase II Clinical Trial. Blood 2012, 120, 78. [Google Scholar]
- Berg, D.; Lehne, M.; Muller, N.; Siegmund, D.; Munkel, S.; Sebald, W.; Pfizenmaier, K.; Wajant, H. Enforced covalent trimerization increases the activity of the TNF ligand family members TRAIL and CD95L. Cell Death Differ. 2007, 14, 2021–2034. [Google Scholar] [CrossRef] [PubMed]
- Gieffers, C.; Kluge, M.; Hill, O.; Meinolf, T.; Merz, C.; Branschädel, M.; Harald, F. APG350, a dimerized single chain TRAIL receptor agonist with enhanced functional properties. Cancer Res. 2011, 71. [Google Scholar] [CrossRef]
- Wang, H.Z.; Davis, J.S.; Wu, X.W. Immunoglobulin Fc Domain Fusion to TRAIL Significantly Prolongs Its Plasma Half-Life and Enhances Its Antitumor Activity. Mol. Cancer Ther. 2014, 13, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, A.B.; Picaud, F.; Rattier, T.; Pudlo, M.; Saviot, L.; Chassagnon, R.; Lherminier, J.; Gharbi, T.; Micheau, O.; Herlem, G. Nanovectorization of TRAIL with Single Wall Carbon Nanotubes Enhances Tumor Cell Killing. Nano Lett. 2015, 15, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Holland, P.M.; Miller, R.; Jones, J.; Douangpanya, H.; Piasecki, J.; Roudier, M.; Dougall, W.C. Combined therapy with the RANKL inhibitor RANK-Fc and rhApo2L/TRAIL/dulanermin reduces bone lesions and skeletal tumor burden in a model of breast cancer skeletal metastasis. Cancer Biol. Ther. 2010, 9, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Soria, J.C.; Smit, E.; Khayat, D.; Besse, B.; Yang, X.; Hsu, C.-P.; Reese, D.; Wiezorek, J.; Blackhall, F. Phase 1b study of dulanermin (recombinant human Apo2L/TRAIL) in combination with paclitaxel, carboplatin, and bevacizumab in patients with advanced non-squamous non-small-cell lung cancer. J. Clin. Oncol. 2010, 28, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Wang, A.; Yang, S.F. Antitumor activity of a novel recombinant mutant human tumor necrosis factor-related apoptosis-inducing ligand. Acta Pharmacol. Sin. 2005, 26, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P. Production of recombinant TRAIL and TRAIL receptor: Fc chimeric proteins. Methods Enzymol. 2000, 322, 325–345. [Google Scholar] [PubMed]
- Muller, N.; Schneider, B.; Pfizenmaier, K.; Wajant, H. Superior serum half life of albumin tagged TNF ligands. Biochem. Biophys. Res. Commun. 2010, 396, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Su, D.; Zhang, J.; Ge, S.; Li, Y.; Wang, F.; Gravel, M.; Roulston, A.; Song, Q.; Xu, W.; et al. Improvement of Pharmacokinetic Profile of TRAIL via Trimer-Tag Enhances its Antitumor Activity in vivo. Sci. Rep. 2017, 7, 8953. [Google Scholar] [CrossRef] [PubMed]
- Hutt, M.; Marquardt, L.; Seifert, O.; Siegemund, M.; Müller, I.; Kulms, D.; Pfizenmaier, K.; Kontermann, R.E. Superior properties of Fc-comprising scTRAIL fusion proteins. Mol. Cancer Ther. 2017. [Google Scholar] [CrossRef]
- Quintavalle, C.; Condorelli, G. Dulanermin in cancer therapy: Still much to do. Transl. Lung Cancer Res. 2012, 1, 158–159. [Google Scholar] [PubMed]
- Belada, D.; Mayer, J.; Czuczman, M.S.; Flinn, I.W.; Durbin-Johnson, B.; Bray, G.L. Phase II study of dulanermin plus rituximab in patients with relapsed follicular non-Hodgkin’s lymphoma (NHL). J. Clin. Oncol. 2010, 28, 8104. [Google Scholar] [CrossRef]
- Blackhall, F.H.; Márk, Z.; Zatloukal, P.; Szima, B.; Albert, I.; Juhász, E.; Yang, X.; Smethurst, D.; Hei, Y.; Soria, J. A randomized phase II study of paclitaxel (P) and carboplatin (C) ± bevacizumab (B) ± dulanermin (D) in non-small cell lung cancer (NSCLC). J. Clin. Oncol. 2010, 28, 7534. [Google Scholar] [CrossRef]
- Kasubhai, S.M.; Bendell, J.C.; Kozloff, M.; Kapp, A.V.; Ashkenazi, A.; Royer-Joo, S.; Portera, C.C. Phase Ib study of dulanermin combined with FOLFIRI (with or without bevacizumab [BV]) in previously treated patients (Pts) with metastatic colorectal cancer (mCRC). J. Clin. Oncol. 2012, 30, 3543. [Google Scholar]
- Kozloff, M.; Messersmith, W.A.; Kapp, A.V.; Ashkenazi, A.; Royer-Joo, S.; Portera, C.C.; Wainberg, Z.A. Phase Ib study of dulanermin combined with first-line FOLFOX plus bevacizumab (BV) in patients (Pts) with metastatic colorectal cancer (mCRC). J. Clin. Oncol. 2012, 30, 3552. [Google Scholar]
- Pan, Y.; Xu, R.; Peach, M.; Huang, C.-P.; Branstetter, D.; Novotny, W.; Herbst, R.S.; Eckhardt, S.G.; Holland, P.M. Evaluation of pharmacodynamic biomarkers in a Phase 1a trial of dulanermin (rhApo2L/TRAIL) in patients with advanced tumours. Br. J. Cancer 2011, 105, 1830–1838. [Google Scholar] [CrossRef] [PubMed]
- Soria, J.C.; Márk, Z.; Zatloukal, P.; Szima, B.; Albert, I.; Juhász, E.; Pujol, J.L.; Kozielski, J.; Baker, N.; Smethurst, D.; et al. Randomized phase II study of dulanermin in combination with paclitaxel, carboplatin, and bevacizumab in advanced non-small-cell lung cancer. J. Clin. Oncol. 2011, 29, 4442–4451. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Lin, J.; Xu, R. The molecular mechanisms of TRAIL resistance in cancer cells: Help in designing new drugs. Curr. Pharm. Des. 2014, 20, 6714–6722. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, L.; van der Sloot, A.M.; Reis, C.R.; Deegan, S.; Ryan, A.E.; Dhami, S.P.S.; Murillo, L.S.; Cool, R.H.; de Sampaio, P.C.; Thompson, K.; et al. Decoy receptors block TRAIL sensitivity at a supracellular level: The role of stromal cells in controlling tumour TRAIL sensitivity. Oncogene 2015, 35, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Selvarajoo, K. A systems biology approach to overcome TRAIL resistance in cancer treatment. Prog. Biophys. Mol. Biol. 2017, 128, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.C.; Kim, K.M.; Lee, K.S.; Namkoong, S.; Lee, S.J.; Han, J.A.; Jeoung, D.; Ha, K.S.; Kwon, Y.G.; Kim, Y.M. Serum bioactive lysophospholipids prevent TRAIL-induced apoptosis via PI3K/Akt-dependent cFLIP expression and Bad phosphorylation. Cell Death Differ. 2004, 11, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Rathore, R.; McCallum, J.E.; Varghese, E.; Florea, A.-M.; Büsselberg, D. Overcoming chemotherapy drug resistance by targeting inhibitors of apoptosis proteins (IAPs). Apoptosis 2017, 22, 898–919. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Feng, L.; Hu, B.; Lu, Y.; Wang, W.; Guo, W.; Wang, S.; Jiao, B.; Pang, J.; Fu, W.; et al. miR-133a Promotes TRAIL Resistance in Glioblastoma via Suppressing Death Receptor 5 and Activating NF-kappaB Signaling. Mol. Ther. Nucleic Acids 2017, 8, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Breunig, C.; Pahl, J.; Küblbeck, M.; Miller, M.; Antonelli, D.; Erdem, N.; Wirth, C.; Will, R.; Bott, A.; Cerwenka, A.; et al. MicroRNA-519a-3p mediates apoptosis resistance in breast cancer cells and their escape from recognition by natural killer cells. Cell Death Dis. 2017, 8, e2973. [Google Scholar] [CrossRef] [PubMed]
- Morizot, A.; Mérino, D.; Lalaoui, N.; Jacquemin, G.; Granci, V.; Iessi, E.; Lanneau, D.; Bouyer, F.; Solary, E.; Chauffert, B.; et al. Chemotherapy overcomes TRAIL-R4-mediated TRAIL resistance at the DISC level. Cell Death Differ. 2011, 18, 700–711. [Google Scholar] [CrossRef] [PubMed]
- A database of privately and publicly funded clinical studies conducted around the world. Available online: https://clinicaltrials.gov (accessed on 16 October 2017).
- De Miguel, D.; Lemke, J.; Anel, A.; Walczak, H.; Martinez-Lostao, L. Onto better TRAILs for cancer treatment. Cell Death Differ. 2016, 23, 733–747. [Google Scholar] [CrossRef] [PubMed]
- Lemke, J.; von Karstedt, S.; Zinngrebe, J.; Walczak, H. Getting TRAIL back on track for cancer therapy. Cell Death Differ. 2014, 21, 1350–1364. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.; Allen, J.E.; Prabhu, V.V.; Talekar, M.K.; Finnberg, N.K.; El-Deiry, W.S. Targeting TRAIL in the treatment of cancer: New developments. Expert Opin. Ther. Targets 2015, 19, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Natoni, A.; Farlane, M.M.; Lnoue, S.; Walewska, R.; Majid, A.; Knee, D.; Stover, D.R.; Dyer, M.J.S.; Cohen, G.M. TRAIL signals to apoptosis in chronic lymphocytic leukaemia cells primarily through TRAIL-R1 whereas cross-linked agonistic TRAIL-R2 antibodies facilitate signalling via TRAIL-R2. Br. J. Haematol. 2007, 139, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Wajant, H.; Moosmayer, D.; Wuest, T.; Bartke, T.; Gerlach, E.; Schonherr, U.; Peters, N.; Scheurich, P.; Pfizenmaier, K. Differential activation of TRAIL-R1 and -2 by soluble and membrane TRAIL allows selective surface antigen-directed activation of TRAIL-R2 by a soluble TRAIL derivative. Oncogene 2001, 20, 4101–4106. [Google Scholar] [CrossRef] [PubMed]
- Leng, Y.; Hou, J.; Jin, J.; Zhang, M.; Ke, X.; Jiang, B.; Pan, L.; Yang, L.; Zhou, F.; Wang, J.; et al. Circularly permuted TRAIL plus thalidomide and dexamethasone versus thalidomide and dexamethasone for relapsed/refractory multiple myeloma: A phase 2 study. Cancer Chemother. Pharmacol. 2017, 79, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Hou, J.; Zhao, Y.; Ke, X.; Wang, Z.; Qiu, L.; Xi, H.; Wang, F.; Wei, N.; Liu, Y.; et al. A multicenter, open-label phase II study of recombinant CPT (Circularly Permuted TRAIL) plus thalidomide in patients with relapsed and refractory multiple myeloma. Am. J. Hematol. 2014, 89, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Somasekharan, S.P.; Koc, M.; Morizot, A.; Micheau, O.; Sorensen, P.H.B.; Gaide, O.; Andera, L.; Martinou, J.-C. TRAIL promotes membrane blebbing, detachment and migration of cells displaying a dysfunctional intrinsic pathway of apoptosis. Apoptosis 2013, 18, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Secchiero, P.; Zerbinati, C.; Rimondi, E.; Corallini, F.; Milani, D.; Grill, V.; Forti, G.; Capitani, S.; Zauli, G. TRAIL promotes the survival, migration and proliferation of vascular smooth muscle cells. Cell. Mol. Life Sci. 2004, 61, 1965–1974. [Google Scholar] [CrossRef] [PubMed]
- Ishimura, N.; Isomoto, H.; Bronk, S.F.; Gores, G.J. Trail induces cell migration and invasion in apoptosis-resistant cholangiocarcinoma cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G129–G136. [Google Scholar] [CrossRef] [PubMed]
- Pukac, L.; Kanakaraj, P.; Humphreys, R.; Alderson, R.; Bloom, M.; Sung, C.; Riccobene, T.; Johnson, R.; Fiscella, M.; Mahoney, M.; et al. HGS-ETR1, a fully human TRAIL-receptor 1 monoclonal antibody, induces cell death in multiple tumour types in vitro and in vivo. Br. J. Cancer 2005, 92, 1430–1441. [Google Scholar] [CrossRef] [PubMed]
- Tolcher, A.W.; Mita, M.; Meropol, N.J.; von Mehren, M.; Patnaik, A.; Padavic, K.; Hill, M.; Mays, T.; McCoy, T.; Fox, N.L.; et al. Phase I pharmacokinetic and biologic correlative study of mapatumumab, a fully human monoclonal antibody with agonist activity to tumor necrosis factor-related apoptosis-inducing ligand receptor-1. J. Clin. Oncol. 2007, 25, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Ciuleanu, T.; Bazin, I.; Lungulescu, D.; Miron, L.; Bondarenko, I.; Deptala, A.; Rodriguez-Torres, M.; Giantonio, B.; Fox, N.L.; Wissel, P.; et al. A randomized, double-blind, placebo-controlled phase II study to assess the efficacy and safety of mapatumumab with sorafenib in patients with advanced hepatocellular carcinoma. Ann. Oncol. 2016, 27, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Greco, F.A.; Bonomi, P.; Crawford, J.; Kelly, K.; Oh, Y.; Halpern, W.; Lo, L.; Gallant, G.; Klein, J. Phase 2 study of mapatumumab, a fully human agonistic monoclonal antibody which targets and activates the TRAIL receptor-1, in patients with advanced non-small cell lung cancer. Lung Cancer 2008, 61, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Hotte, S.J.; Hirte, H.W.; Chen, E.X.; Siu, L.L.; Le, L.H.; Corey, A.; Iacobucci, A.; MacLean, M.; Lo, L.; Fox, N.L.; et al. A phase 1 study of mapatumumab (fully human monoclonal antibody to TRAIL-R1) in patients with advanced solid malignancies. Clin. Cancer Res. 2008, 14, 3450–3455. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Morton, C.L.; Kolb, E.A.; Gorlick, R.; Keir, S.T.; Carol, H.; Lock, R.; Kang, M.H.; Reynolds, C.P.; Maris, J.M.; et al. Initial testing (stage 1) of mapatumumab (HGS-ETR1) by the pediatric preclinical testing program. Pediatr. Blood Cancer 2010, 54, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Nelson, D.; Alberts, S.R.; Poordad, F.; Leong, S.; Teitelbaum, U.R.; Woods, L.; Fox, N.; O’Neil, B.H. Phase Ib study of mapatumumab in combination with sorafenib in patients with advanced hepatocellular carcinoma (HCC) and chronic viral hepatitis. J. Clin. Oncol. 2011, 29, 261. [Google Scholar] [CrossRef]
- Trarbach, T.; Moehler, M.; Heinemann, V.; Köhne, C.-H.; Przyborek, M.; Schulz, C.; Sneller, V.; Gallant, G.; Kanzler, S. Phase II trial of mapatumumab, a fully human agonistic monoclonal antibody that targets and activates the tumour necrosis factor apoptosis-inducing ligand receptor-1 (TRAIL-R1), in patients with refractory colorectal cancer. Br. J. Cancer 2010, 102, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Von Pawel, J.; Harvey, J.H.; Spigel, D.R.; Dediu, M.; Reck, M.; Cebotaru, C.L.; Humphreys, R.C.; Gribbin, M.J.; Fox, N.L.; Camidge, D.R. Phase II trial of mapatumumab, a fully human agonist monoclonal antibody to tumor necrosis factor-related apoptosis-inducing ligand receptor 1 (TRAIL-R1), in combination with paclitaxel and carboplatin in patients with advanced non-small-cell lung cancer. Clin. Lung Cancer 2014, 15, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Von Pawel, J.; Harvey, J.H.; Spigel, D.R.; Dediu, M.; Reck, M.; Cebotaru, C.L.; Kumm, E.; Gallant, G.; Fox, N.; Camidge, D.R. A randomized phase II trial of mapatumumab, a TRAIL-R1 agonist monoclonal antibody, in combination with carboplatin and paclitaxel in patients with advanced NSCLC. J. Clin. Oncol. 2010, 28. [Google Scholar] [CrossRef]
- Younes, A.; Harvey, J.H.; Spigel, D.R.; Dediu, M.; Reck, M.; Cebotaru, C.L.; Kumm, E.; Gallant, G.; Fox, N.; Camidge, D.R. A Phase 1b/2 trial of mapatumumab in patients with relapsed/refractory non-Hodgkin’s lymphoma. Br. J. Cancer 2010, 103, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xiao, X.; Zhu, Z.; Dimitrov, D.S. Identification and characterization of a novel agonistic anti-DR4 human monoclonal antibody. MAbs 2010, 2, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Chuntharapai, A.; Dodge, K.; Grimmer, K.; Schroeder, K.; Marsters, S.A.; Koeppen, H.; Ashkenazi, A.; Kim, K.J. Isotype-dependent inhibition of tumor growth in vivo by monoclonal antibodies to death receptor 4. J. Immunol. 2001, 166, 4891–4898. [Google Scholar] [CrossRef] [PubMed]
- Sung, E.-S.; Park, K.-J.; Lee, S.-H.; Jang, Y.-S.; Park, S.-K.; Park, Y.-H.; Kwag, W.-J.; Kwon, M.-H.; Kim, Y.-S. A novel agonistic antibody to human death receptor 4 induces apoptotic cell death in various tumor cells without cytotoxicity in hepatocytes. Mol. Cancer Ther. 2009, 8, 2276–2285. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Jin, A.; Ozawa, T.; Tajiri, K.; Obata, T.; Ishida, I.; Jin, F.; Kishi, H.; Muraguchi, A. Post-translational modification of TRAIL receptor type 1 on various tumor cells and the susceptibility of tumors to TRAIL-induced apoptosis. Biochem. Biophys. Res. Commun. 2010, 395, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Burvenich, I.J.; Lee, F.-T.; Guo, N.; Gan, H.K.; Rigopoulos, A.; Parslow, A.C.; O’Keefe, G.J.; Gong, S.J.; Tochon-Danguy, H.; Rudd, S.E.; et al. In Vitro and In Vivo Evaluation of 89Zr-DS-8273a as a Theranostic for Anti-Death Receptor 5 Therapy. Theranostics 2016, 6, 2225–2234. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, G.A.; Condamine, T.; Mony, S.; Hashimoto, A.; Wang, F.; Liu, Q.; Forero, A.; Bendell, J.; Witt, R.; Hockstein, N.; et al. Selective Targeting of Myeloid-Derived Suppressor Cells in Cancer Patients Using DS-8273a, an Agonistic TRAIL-R2 Antibody. Clin. Cancer Res. 2017, 23, 2942–2950. [Google Scholar] [CrossRef] [PubMed]
- Forero, A.; Bendell, J.C.; Kumar, P.; Janisch, L.; Rosen, M.; Wang, Q.; Copigneaux, C.; Desai, M.; Senaldi, G.; Maitland, M.L. First-in-human study of the antibody DR5 agonist DS-8273a in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Herbst, R.S.; Kurzrock, R.; Hong, D.S.; Valdivieso, M.; Hsu, C.-P.; Goyal, L.; Juan, G.; Hwang, Y.; Wong, S.; Hill, J.S.; et al. A first-in-human study of conatumumab in adult patients with advanced solid tumors. Clin. Cancer Res. 2010, 16, 5883–5891. [Google Scholar] [CrossRef] [PubMed]
- LoRusso, P.M.; Hong, D.; Heath, E.; Kurzrock, R.; Wang, D.; Hsu, M.; Goyal, L.; Wiezorek, J.; Storgard, C.; Herbst, R. First-in-human study of AMG 655, a pro-apoptotic TRAIL receptor-2 agonist, in adult patients with advanced solid tumors. J. Clin. Oncol. 2007, 25, 3534. [Google Scholar]
- Camidge, D.R.; Herbst, R.S.; Gordon, M.S.; Eckhardt, S.G.; Kurzrock, R.; Durbin, B.; Ing, J.; Tohnya, T.M.; Sager, J.; Ashkenazi, A.; et al. A phase I safety and pharmacokinetic study of the death receptor 5 agonistic antibody PRO95780 in patients with advanced malignancies. Clin. Cancer Res. 2010, 16, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.; Chen, J.-J.; Yu, Y.; Li, B.; Sun, S.-Y.; Zhang, B.; Cao, L. Drozitumab, a human antibody to death receptor 5, has potent antitumor activity against rhabdomyosarcoma with the expression of caspase-8 predictive of response. Clin. Cancer Res. 2011, 17, 3181–3192. [Google Scholar] [CrossRef] [PubMed]
- Forero-Torres, A.; Infante, J.R.; Waterhouse, D.; Wong, L.; Vickers, S.; Arrowsmith, E.; He, A.R.; Har, L.; Trent, D.; Wade, J.; et al. Phase 2, multicenter, open-label study of tigatuzumab (CS-1008), a humanized monoclonal antibody targeting death receptor 5, in combination with gemcitabine in chemotherapy-naive patients with unresectable or metastatic pancreatic cancer. Cancer Med. 2013, 2, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Forero-Torres, A.; Shah, J.; Wood, T.; Posey, J.; Carlisle, R.; Copigneaux, C.; Luo, F.; Wojtowicz-Praga, S.; Percent, I.; Saleh, M. Phase I trial of weekly tigatuzumab, an agonistic humanized monoclonal antibody targeting death receptor 5 (DR5). Cancer Biother. Radiopharm. 2010, 25, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, K.; Liu, W.; Zhao, L.; Wang, Z.; Liu, D.; Ohtsuka, T. Tumoricidal activity of a novel anti-human DR5 monoclonal antibody without hepatocyte cytotoxicity. Nat. Med. 2001, 7, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Belyanskaya, L.L.; Marti, T.M.; Hopkins-Donaldson, S.; Kurtz, S.; Felley-Bosco, E.; Stahel, R.A. Human agonistic TRAIL receptor antibodies Mapatumumab and Lexatumumab induce apoptosis in malignant mesothelioma and act synergistically with cisplatin. Mol. Cancer 2007, 6, 66. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Qiu, Y.; Hao, Z.; Cai, J.; Zhang, S.; Liu, Y.; Zheng, D. A novel humanized anti-tumor necrosis factor-related apoptosis-inducing ligand-R2 monoclonal antibody induces apoptotic and autophagic cell death. IUBMB Life 2017. [Google Scholar] [CrossRef] [PubMed]
- Rocha Lima, C.M.; Bayraktar, S.; Flores, A.M.; MacIntyre, J.; Montero, A.; Baranda, J.C.; Wallmark, J.; Portera, C.; Raja, R.; et al. Phase Ib study of drozitumab combined with first-line mFOLFOX6 plus bevacizumab in patients with metastatic colorectal cancer. Cancer Investig. 2012, 30, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Shimada, O.; Wu, X.; Jin, X.; Abdel-Muneem Nouh, M.A.; Fiscella, M.; Albert, V.; Matsuda, T.; Kakehi, Y. Human agonistic antibody to tumor necrosis factor-related apoptosis-inducing ligand receptor 2 induces cytotoxicity and apoptosis in prostate cancer and bladder cancer cells. Urology 2007, 69, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; de Vries, E.G.; Infante, J.R.; Oldenhuis, C.N.; Gietema, J.A.; Yang, L.; Bilic, S.; Parker, K.; Goldbrunner, M.; Scott, J.W.; et al. Safety, pharmacokinetics, and pharmacodynamics of the DR5 antibody LBY135 alone and in combination with capecitabine in patients with advanced solid tumors. Investig. New Drugs 2014, 32, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. Safety and tolerability of TRAIL receptor agonists in cancer treatment. Eur. J. Clin. Pharmacol. 2015, 71, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Holland, P.M. Death receptor agonist therapies for cancer, which is the right TRAIL? Cytokine Growth Factor Rev. 2014, 25, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A. Targeting the extrinsic apoptotic pathway in cancer: Lessons learned and future directions. J. Clin. Investig. 2015, 125, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Kasibhatla, S.; Tseng, B. Why target apoptosis in cancer treatment? Mol. Cancer Ther. 2003, 2, 573–580. [Google Scholar] [PubMed]
- Baetu, T.M.; Hiscott, J. On the TRAIL to apoptosis. Cytokine Growth Factor Rev. 2002, 13, 199–207. [Google Scholar] [CrossRef]
- Lemke, J.; von Karstedt, S.; El Hay, M.A.; Conti, A.; Arce, F.; Montinaro, A.; Papenfuss, K.; El-Bahrawy, M.A.; Walczak, H. Selective CDK9 inhibition overcomes TRAIL resistance by concomitant suppression of cFlip and Mcl-1. Cell Death Differ. 2014, 21, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.A.; Powley, I.R.; Jukes-Jones, R.; Horn, S.; Feoktistova, M.; Feoktistova, L.; Schwabe, J.W.R.; Leverkus, M.; Cain, K.; MacFarlane, M. Co-operative and Hierarchical Binding of c-FLIP and Caspase-8: A Unified Model Defines How c-FLIP Isoforms Differentially Control Cell Fate. Mol. Cell. 2016, 61, 834–849. [Google Scholar] [CrossRef] [PubMed]
- Ndozangue-Touriguine, O.; Sebbagh, M.; Merino, D.; Micheau, O.; Bertoglio, J.; Breard, J. A mitochondrial block and expression of XIAP lead to resistance to TRAIL-induced apoptosis during progression to metastasis of a colon carcinoma. Oncogene 2008, 27, 6012–6022. [Google Scholar] [CrossRef] [PubMed]
- Finlay, D.; Vamos, M.; González-López, M.; Ardecky, R.J.; Ganji, S.R.; Yuan, H.; Su, Y.; Cooley, T.R.; Hauser, C.T.; Welsh, K.; et al. Small-molecule IAP antagonists sensitize cancer cells to TRAIL-induced apoptosis: Roles of XIAP and cIAPs. Mol. Cancer Ther. 2014, 13, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Perimenis, P.; Galaris, A.; Voulgari, A.; Prassa, M.; Pintzas, A. IAP antagonists Birinapant and AT-406 efficiently synergise with either TRAIL, BRAF, or BCL-2 inhibitors to sensitise BRAFV600E colorectal tumour cells to apoptosis. BMC Cancer 2016, 16, 624. [Google Scholar] [CrossRef] [PubMed]
- Wang, S. The promise of cancer therapeutics targeting the TNF-related apoptosis-inducing ligand and TRAIL receptor pathway. Oncogene 2008, 27, 6207–6215. [Google Scholar] [CrossRef] [PubMed]
- Griffith, T.S.; Rauch, C.T.; Smolak, P.J.; Waugh, J.Y.; Boiani, N.; Lynch, D.H.; Smith, C.A.; Goodwin, R.G.; Kubin, M.Z. Functional analysis of TRAIL receptors using monoclonal antibodies. J. Immunol. 1999, 162, 2597–2605. [Google Scholar] [PubMed]
- Takeda, K.; Yamaguchi, N.; Akiba, H.; Kojima, Y.; Hayakawa, Y.; Tanner, J.E.; Sayers, T.J.; Seki, N.; Okumura, K.; Yagita, H.; et al. Induction of Tumor-specific T Cell Immunity by Anti-DR5 Antibody Therapy. J. Exp. Med. 2004, 199, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Nomura, T.; Takai, T.; Chiba, T.; Honjo, T.; Tsubata, T. Antigen receptor cross-linking by anti-immunoglobulin antibodies coupled to cell surface membrane induces rapid apoptosis of normal spleen B cells. Scand. J. Immunol. 1998, 47, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Szegezdi, E.; Leverkus, M. Guiding the Killer and Bringing in Accomplices: Bispecific Antibody Treatment for Malignant Melanoma. J. Investig. Dermatol. 2016, 136, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Tamada, T.; Shinmi, D.; Ikeda, M.; Yonezawa, Y.; Kataoka, S.; Kuroki, R.; Mori, E.; Motoki, K. TRAIL-R2 Superoligomerization Induced by Human Monoclonal Agonistic Antibody KMTR2. Sci. Rep. 2015, 5, 17936. [Google Scholar] [CrossRef] [PubMed]
- Motoki, K.; Mori, E.; Matsumoto, A.; Thomas, M.; Tomura, T.; Humphreys, R.; Albert, V.; Muto, M.; Yoshida, H.; Aoki, M.; et al. Enhanced apoptosis and tumor regression induced by a direct agonist antibody to tumor necrosis factor-related apoptosis-inducing ligand receptor 2. Clin. Cancer Res. 2005, 11, 3126–3135. [Google Scholar] [CrossRef] [PubMed]
- Nagane, M.; Shimizu, S.; Mori, E.; Kataoka, S.; Shiokawa, Y. Predominant antitumor effects by fully human anti-TRAIL-receptor 2 (DR5) monoclonal antibodies in human glioma cells in vitro and in vivo. Neuro Oncol. 2010, 12, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.D.; Kordich, J.J.; Huang, T.H.; Piasecki, J.; Bush, T.L.; Sullivan, T.; Foltz, I.N.; Chang, W.; Douangpanya, H.; Dang, T.; et al. Apo2L/TRAIL and the Death Receptor 5 Agonist Antibody AMG 655 Cooperate to Promote Receptor Clustering and Antitumor Activity. Cancer Cell 2014, 26, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Tuthill, M.H.; Montinaro, A.; Zinngrebe, J.; Prieske, K.; Draber, P.; Prieske, S.; Newsom-Davis, T.; von Karstedt, S.; Graves, J.; Walczak, H. TRAIL-R2-specific antibodies and recombinant TRAIL can synergise to kill cancer cells. Oncogene 2015, 34, 2138–2144. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.C.; Han, X.; Sun, X.; Shen, M.; Huang, J.; Li, Y.; Ozawa, T.; Pang, D.; Jin, S.; Kishi, H.; et al. Fully human monoclonal antibodies to TRAIL-R1 enhance TRAIL-induced apoptosis via activation of caspase-8 pathway. Biochem. Biophys. Res. Commun. 2016, 475, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.L. Cancer Therapeutic Targets; Springer: New York, NY, USA, 2017; p. 1105. [Google Scholar]
- Hendriks, D.; Choi, G.; de Bruyn, M.; Wiersma, V.R.; Bremer, E. Antibody-Based Cancer Therapy: Successful Agents and Novel Approaches. Int. Rev. Cell Mol. Biol. 2017, 331, 289–383. [Google Scholar] [PubMed]
- Wajant, H. Principles of antibody-mediated TNF receptor activation. Cell Death Differ. 2015, 22, 1727–1741. [Google Scholar] [CrossRef] [PubMed]
- Pavet, V.; Beyrath, J.; Pardin, C.; Morizot, A.; Lechner, M.-C.; Briand, J.-P.; Wendland, M.; Maison, W.; Fournel, S.; Micheau, O.; et al. Multivalent DR5 peptides activate the TRAIL death pathway and exert tumoricidal activity. Cancer Res. 2010, 70, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Si, Y.; Liu, G.; Li, S.; Zhang, J.; Ma, Y. The tetravalent anti-DR5 antibody without cross-linking direct induces apoptosis of cancer cells. Biomed. Pharmacother. 2015, 70, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, K.P.; Isaacs, R.; Bilic, S.; Kentsch, K.; Huet, H.A.; Hofmann, A.; Rasco, D.; Kundamal, N.; Tang, Z.; Cooksey, J.; et al. Unexpected hepatotoxicity in a phase I study of TAS266, a novel tetravalent agonistic Nanobody(R) targeting the DR5 receptor. Cancer Chemother. Pharmacol. 2015, 75, 887–895. [Google Scholar]
- Siegel, M.R.; Sisler, H.D. Inhibition of Protein Synthesis in Vitro by Cycloheximide. Nature 1963, 200, 675–676. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Meyer, E.; Debatin, K.M. Metabolic inhibitors sensitize for CD95 (APO-1/Fas)-induced apoptosis by down-regulating Fas-associated death domain-like interleukin 1-converting enzyme inhibitory protein expression. Cancer Res. 2000, 60, 3947–3956. [Google Scholar] [PubMed]
- Bruhns, P.; Iannascoli, B.; England, P.; Mancardi, D.A.; Fernandez, N.; Jorieux, S.; Daëron, M. Specificity and affinity of human Fc gamma receptors and their polymorphic variants for human IgG subclasses. Blood 2009, 113, 3716–3725. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ravetch, J.V. Antitumor activities of agonistic anti-TNFR antibodies require differential FcgammaRIIB coengagement in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 19501–19506. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Meng, Q.; Gao, C.; Zhuang, G.; Huang, X.; Zhang, J.; Liu, B.; Fan, X.; Zhang, M. Analysis of aDR5scFv with Specific Identification and Function. Monoclon. Antib. Immunodiagn. Immunother. 2016, 35, 25–31. [Google Scholar]
- Piao, X.H.; Ozawa, T.; Hamana, H.; Shitaoka, K.; Jin, A.; Kishi, H.; Muraguchi, A. TRAIL-receptor 1 IgM antibodies strongly induce apoptosis in human cancer cells in vitro and in vivo. Oncoimmunology 2016, 5, e1131380. [Google Scholar] [CrossRef] [PubMed]
- Kaplan-Lefko, P.J.; Graves, J.D.; Zoog, S.J.; Pan, Y.; Wall, J.; Branstetter, D.G.; Moriguchi, J.; Coxon, A.; Huard, J.N.; et al. Conatumumab, a fully human agonist antibody to death receptor 5, induces apoptosis via caspase activation in multiple tumor types. Cancer Biol. Ther. 2010, 9, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, Z.A.; Yeap, S.K.; Ali, A.M.; Ho, W.Y.; Alitheen, N.B.M.; Hamid, M. scFv Antibody: Principles and Clinical Application. Clin. Dev. Immunol. 2012, 980250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Yang, Y.; Fan, D.; Xiong, D. The development of bispecific antibodies and their applications in tumor immune escape. Exp. Hematol. Oncol. 2017, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Naoum, G.E.; Buchsbaum, D.J.; Tawadros, F.; Farooqi, A.; Arafat, W.O. Journey of TRAIL from Bench to Bedside and its Potential Role in Immuno-Oncology. Oncol. Rev. 2017, 11, 332. [Google Scholar] [CrossRef] [PubMed]
- Arabpour, M.; Poelstra, K.; Helfrich, W.; Bremer, E.; Haisma, H.J. Targeted elimination of activated hepatic stellate cells by an anti-epidermal growth factor-receptor single chain fragment variable antibody-tumor necrosis factor-related apoptosis-inducing ligand (scFv425-sTRAIL). J. Gene Med. 2014, 16, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; Samplonius, D.F.; van Genne, L.; Dijkstra, M.H.; Kroesen, B.J.; de Leij, L.F.M.H.; Helfrich, W. Simultaneous inhibition of epidermal growth factor receptor (EGFR) signaling and enhanced activation of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) receptor-mediated apoptosis induction by an scFv:sTRAIL fusion protein with specificity for human EGFR. J. Biol. Chem. 2005, 280, 10025–10033. [Google Scholar] [PubMed]
- Bremer, E.; Samplonius, D.; Kroesen, B.-J.; van Genne, L.; de Leij, L.; Helfrich, W. Exceptionally potent anti-tumor bystander activity of an scFv:sTRAIL fusion protein with specificity for EGP2 toward target antigen-negative tumor cells. Neoplasia 2004, 6, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Hartung, F.; Pardo, L.A. Guiding TRAIL to cancer cells through Kv10.1 potassium channel overcomes resistance to doxorubicin. Eur. Biophys. J. 2016, 45, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Tatzel, K.; Kuroki, L.; Dmitriev, I.; Kashentseva, E.; Curiel, D.T.; Goedegebuure, S.P.; Powell, M.A.; Mutch, D.G.; Hawkins, W.G.; Spitzer, D. Membrane-proximal TRAIL species are incapable of inducing short circuit apoptosis signaling: Implications for drug development and basic cytokine biology. Sci. Rep. 2016, 6, 22661. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, M.; Rybczynska, A.A.; Wei, Y.; Schwenkert, M.; Fey, G.H.; Dierckx, R.A.; van Waarde, A.; Helfrich, W.; Bremer, E. Melanoma-associated Chondroitin Sulfate Proteoglycan (MCSP)-targeted delivery of soluble TRAIL potently inhibits melanoma outgrowth in vitro and in vivo. Mol. Cancer 2010, 9, 301. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Ni, C.-W.; Lin, Y.-Z.; Yin, L.; Jiang, C.-B.; Lv, C.-T.; Le, Y.; Lang, Y.; Zhao, C.-Y.; Yang, K.; et al. Targeted induction of apoptosis in glioblastoma multiforme cells by an MRP3-specific TRAIL fusion protein in vitro. Tumour Biol. 2014, 35, 1157–1168. [Google Scholar] [CrossRef] [PubMed]
- Hartung, F.; Stuhmer, W.; Pardo, L.A. Tumor cell-selective apoptosis induction through targeting of K(V)10.1 via bifunctional TRAIL antibody. Mol. Cancer 2011, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, D.S. Clinical implications of EGFR expression in the development and progression of solid tumors: Focus on non-small cell lung cancer. Oncologist 2006, 11, 358–373. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.N.; Bassoff, N.; Reinshagen, C.; Bhere, D.; Nowicki, M.O.; Lawler, S.E.; Roux, J.; Shah, K. Bi-specific molecule against EGFR and death receptors simultaneously targets proliferation and death pathways in tumors. Sci. Rep. 2017, 7, 2602. [Google Scholar] [CrossRef] [PubMed]
- Siegemund, M.; Pollak, N.; Seifert, O.; Wahl, K.; Hanak, K.; Vogel, A.; Nussler, A.K.; Göttsch, D.; Münkel, S.; Bantel, H.; et al. Superior antitumoral activity of dimerized targeted single-chain TRAIL fusion proteins under retention of tumor selectivity. Cell Death Dis. 2012, 3, e295. [Google Scholar] [CrossRef] [PubMed]
- Siegemund, M.; Seifert, O.; Zarani, M.; Džinić, T.; De Leo, V.; Göttsch, D.; Münkel, S.; Hutt, M.; Pfizenmaier, K.; Kontermann, R.E. An optimized antibody-single-chain TRAIL fusion protein for cancer therapy. MAbs 2016, 8, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Holbro, T.; Hynes, N.E. ErbB receptors: Directing key signaling networks throughout life. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Kouno, M.; Lin, C.; Schechter, N.M.; Siegel, D.; Yang, X.; Seykora, J.T.; Stanley, J.R. Targeted delivery of tumor necrosis factor-related apoptosis-inducing ligand to keratinocytes with a pemphigus mAb. J. Investig. Dermatol. 2013, 133, 2212–2220. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yang, H.; Jia, D.; Nie, Q.; Cai, H.; Fan, Q.; Wan, L.; Li, L.; Lu, X. Fusion to an albumin-binding domain with a high affinity for albumin extends the circulatory half-life and enhances the in vivo antitumor effects of human TRAIL. J. Control Release 2016, 228, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Hu, M.; Tang, B.; Liu, X.; Zhuang, H.; Yang, J.; Hua, Z.-C. Annexin V-TRAIL fusion protein is a more sensitive and potent apoptotic inducer for cancer therapy. Sci. Rep. 2013, 3, 3565. [Google Scholar] [CrossRef] [PubMed]
- Sang, M.; Zhang, J.; Li, B.; Chen, Y. TRAIL-CM4 fusion protein shows in vitro antibacterial activity and a stronger antitumor activity than solo TRAIL protein. Protein Expr. Purif. 2016, 122, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Prigozhina, T.B.; Szafer, F.; Aronin, A.; Tzdaka, K.; Amsili, S.; Makdasi, E.; Shani, N.; Elhalel, M.D. Fn14.TRAIL fusion protein is oligomerized by TWEAK into a superefficient TRAIL analog. Cancer Lett. 2017, 400, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Du, P.; Jiang, S.-H.; Jin, G.-H.; Huang, Q.-L.; Hua, Z.-C. Enhancement of antitumor properties of TRAIL by targeted delivery to the tumor neovasculature. Mol. Cancer Ther. 2008, 7, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Duan, N.; Zou, W.; Zhang, C.; Lai, Y.; Shen, P.; Hua, Z. Fused hydrophobic elastin-like-peptides (ELP) enhance biological activity of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). Protein Pept. Lett. 2015, 22, 1000–1006. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Sui, A.; Wang, Z.; Liu, S.; Zhou, Q.; Liu, X.; Zhang, H. Induction of non-small cell lung carcinoma apoptosis using soluble RGD-TRAIL by targeting the integrin receptor of tumor cells. Mol. Med. Rep. 2012, 6, 1355–1360. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, X.; Sha, H.; Zhang, L.; Bian, X.; Han, X.; Liu, B. sTRAIL-iRGD is a promising therapeutic agent for gastric cancer treatment. Sci. Rep. 2017, 7, 579. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Ma, X.; Wang, H.; Xi, Y.; Qian, M.; Yang, W.; Luo, D.; Fan, L.; Xia, X.; Zhou, J.; et al. The novel fusion protein sTRAIL-TMTP1 exhibits a targeted inhibition of primary tumors and metastases. J. Mol. Med. 2014, 92, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Wang, Z.; Huang, L.; Shen, Y. Efficient refolding of the bifunctional therapeutic fusion protein VAS-TRAIL by a triple agent solution. Protein Expr. Purif. 2016, 125, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qiao, X.; Shang, Y.; Zhang, S.; Li, Y.; He, H.; Chen, S.-Z. RGD and NGR modified TRAIL protein exhibited potent anti-metastasis effects on TRAIL-insensitive cancer cells in vitro and in vivo. Amino Acids 2017, 49, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Yang, H.; Shi, Q.; Fan, Q.; Wan, L.; Lu, X. Targeted Delivery to Tumor-associated Pericytes via an Affibody with High Affinity for PDGFRbeta Enhances the in vivo Antitumor Effects of Human TRAIL. Theranostics 2017, 7, 2261–2276. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Tatzel, K.; Wang, X.; Belt, B.; Binder, P.; Kuroki, L.; Powell, M.A.; Mutch, D.G.; Hawkins, W.G.; Spitzer, D. Mesothelin’s minimal MUC16 binding moiety converts TR3 into a potent cancer therapeutic via hierarchical binding events at the plasma membrane. Oncotarget 2016, 7, 31534–31549. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, M.; Wei, Y.; Wiersma, V.R.; Samplonius, D.F.; Klip, H.G.; van der Zee, A.G.J.; Yang, B.; Helfrich, W.; Bremer, E. Cell surface delivery of TRAIL strongly augments the tumoricidal activity of T cells. Clin. Cancer Res. 2011, 17, 5626–5637. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; Samplonius, D.G.; Peipp, M.; van Genne, L.; Kroesen, B.J.; Fey, G.H.; Gramatzki, M.; de Leij, L.F.M.H.; Helfrich, W. Target cell-restricted apoptosis induction of acute leukemic T cells by a recombinant tumor necrosis factor-related apoptosis-inducing ligand fusion protein with specificity for human CD7. Cancer Res. 2005, 65, 3380–3388. [Google Scholar] [CrossRef] [PubMed]
- Stieglmaier, J.; Bremer, E.; Kellner, C.; Liebig, T.M.; ten Cate, B.; Peipp, M.; Schulze-Koops, H.; Pfeiffer, M.; Bühring, H.-J.; Greil, J.; et al. Selective induction of apoptosis in leukemic B-lymphoid cells by a CD19-specific TRAIL fusion protein. Cancer Immunol. Immunother. 2008, 57, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Uckun, F.M.; Myerd, D.E.; Qazi, S.; Ozer, Z.; Rose, R.; D’Cruz, O.J.; Ma, H. Recombinant human CD19L-sTRAIL effectively targets B cell precursor acute lymphoblastic leukemia. J. Clin. Investig. 2015, 125, 1006–1018. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Li, S.; Li, Z.; Peng, H.; Yuan, X.; Jiang, L.; Zhang, Y.; Fan, D.; Hu, X.; Yang, M.; et al. Human umbilical cord mesenchymal stem cells as vehicles of CD20-specific TRAIL fusion protein delivery: A double-target therapy against non-Hodgkin’s lymphoma. Mol. Pharm. 2013, 10, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Madhumathi, J.; Sridevi, S.; Verma, R.S. Novel TNF-related Apoptotic-inducing Ligand-based Immunotoxin for Therapeutic Targeting of CD25 Positive Leukemia. Target Oncol. 2016, 11, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Ten Cate, B.; Bremer, E.; de Bruyn, M.; Bijma, T.; Samplonius, D.F.; Schwemmlein, M.; Huls, G.; Fey, G.H.; Helfrich, W. A novel AML-selective TRAIL fusion protein that is superior to Gemtuzumab Ozogamicin in terms of in vitro selectivity, activity and stability. Leukemia 2009, 23, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- El-Mesery, M.; Trebing, J.; Schäfer, V.; Weisenberger, D.; Siegmund, D.; Wajant, H. CD40-directed scFv-TRAIL fusion proteins induce CD40-restricted tumor cell death and activate dendritic cells. Cell Death Dis. 2013, 4, e916. [Google Scholar] [CrossRef] [PubMed]
- Wiersma, V.R.; He, Y.; Samplonius, D.F.; van Ginkel, R.J.; Gerssen, J.; Eggleton, P.; Zhou, J.; Bremer, E.; Helfrich, W. A CD47-blocking TRAIL fusion protein with dual pro-phagocytic and pro-apoptotic anticancer activity. Br. J. Haematol. 2014, 164, 304–307. [Google Scholar] [CrossRef] [PubMed]
- Trebing, J.; El-Mesery, M.; Schäfer, V.; Weisenberger, D.; Siegmund, D.; Silence, K.; Wajant, H. CD70-restricted specific activation of TRAILR1 or TRAILR2 using scFv-targeted TRAIL mutants. Cell Death Dis. 2014, 5, e1035. [Google Scholar] [CrossRef] [PubMed]
- Wiersma, V.R.; de Bruyn, M.; Shi, C.; Gooden, M.J.M.; Wouters, M.C.A.; Samplonius, D.F.; Hendriks, D.; Nijman, H.W.; Wei, Y.; Zhou, J.; et al. C-type lectin-like molecule-1 (CLL1)-targeted TRAIL augments the tumoricidal activity of granulocytes and potentiates therapeutic antibody-dependent cell-mediated cytotoxicity. MAbs 2015, 7, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, D.; He, Y.; Koopmans, I.; Wiersma, V.R.; van Ginkel, R.J.; Samplonius, D.F.; Helfrich, W.; Bremer, E. Programmed Death Ligand 1 (PD-L1)-targeted TRAIL combines PD-L1-mediated checkpoint inhibition with TRAIL-mediated apoptosis induction. Oncoimmunology 2016, 5, e1202390. [Google Scholar] [CrossRef] [PubMed]
- Dobson, C.L.; Main, S.; Newton, P.; Chodorge, M.; Cadwallader, K.; Humphreys, R.; Albert, V.; Vaughan, T.J.; Minter, R.R.; Edwards, B.M. Human monomeric antibody fragments to TRAIL-R1 and TRAIL-R2 that display potent in vitro agonism. MAbs 2009, 1, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Swers, J.S.; Grinberg, L.; Wang, L.; Feng, H.; Lekstrom, K.; Carrasco, R.; Xiao, Z.; Inigo, I.; Leow, C.C.; Wu, H.; Tice, D.A.; Baca, M. Multivalent scaffold proteins as superagonists of TRAIL receptor 2-induced apoptosis. Mol. Cancer Ther. 2013, 12, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Park, K.J.; Kim, S.J.; Kwon, O.; Jeong, K.J.; Kim, A.; Kim, Y.S. Generation of bivalent and bispecific kringle single domains by loop grafting as potent agonists against death receptors 4 and 5. J. Mol. Biol. 2011, 411, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Park, K.J.; Sung, E.S.; Kim, A.; Choi, J.D.; Kim, J.D.; Kim, S.H.; Kwon, M.K.; Kim, Y.S. Engineering of a human kringle domain into agonistic and antagonistic binding proteins functioning in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 9567–9571. [Google Scholar] [CrossRef] [PubMed]
- Park, K.J.; Lee, C.H.; Kim, A.; Jeong, K.J.; Kim, C.H.; Kim, Y.S. Death receptors 4 and 5 activate Nox1 NADPH oxidase through riboflavin kinase to induce reactive oxygen species-mediated apoptotic cell death. J. Biol. Chem. 2012, 287, 3313–3325. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.M.; Lee, Y.J.; Kim, Y.S.; Jeong, K.J. High-level production of Fc-fused kringle domain in Pichia pastoris. J. Ind. Microbiol. Biotechnol. 2014, 41, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Milutinovic, S.; Kashyap, A.K.; Yanagi, T.; Wimer, C.; Zhou, S.; O’Neil, R.; Kurtzman, A.L.; Faynboym, A.; Xu, L.; Hannum, C.H.; et al. Dual Agonist Surrobody Simultaneously Activates Death Receptors DR4 and DR5 to Induce Cancer Cell Death. Mol. Cancer Ther. 2016, 15, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Michaelson, J.S.; Demarest, S.J.; Miller, B.; Amatucci, A.; Snyder, W.B.; Wu, X.; Huang, F.; Phan, S.; Gao, S.; Doern, A.; et al. Anti-tumor activity of stability-engineered IgG-like bispecific antibodies targeting TRAIL-R2 and LTbetaR. MAbs 2009, 1, 128–141. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Hendriks, D.; van Ginkel, R.; Samplonius, D.; Bremer, E.; Helfrich, W. Melanoma-Directed Activation of Apoptosis Using a Bispecific Antibody Directed at MCSP and TRAIL Receptor-2/Death Receptor-5. J. Investig. Dermatol. 2016, 136, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Brunker, P.; Wartha, K.; Friess, T.; Grau-Richards, S.; Waldhauer, I.; Koller, C.F.; Weiser, B.; Majety, M.; Runza, V.; Niu, H.; et al. RG7386, a Novel Tetravalent FAP-DR5 Antibody, Effectively Triggers FAP-Dependent, Avidity-Driven DR5 Hyperclustering and Tumor Cell Apoptosis. Mol. Cancer Ther. 2016, 15, 946–957. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Hong, B.-J.; Lee, J.H.; Yeo, S.; Jung, H.Y.; Chung, J.; Ahn, G.-O.; Hahn, S.K. Hyaluronate-Death Receptor 5 Antibody Conjugates for Targeted Treatment of Liver Metastasis. Biomacromolecules 2016, 17, 3085–3093. [Google Scholar] [CrossRef] [PubMed]
- Gross, G.; Gorochov, G.; Waks, T.; Eshhar, Z. Generation of effector T cells expressing chimeric T cell receptor with antibody type-specificity. Transplant. Proc. 1989, 21, 127–130. [Google Scholar] [PubMed]
- Gross, G.; Waks, T.; Eshhar, Z. Expression of immunoglobulin-T-cell receptor chimeric molecules as functional receptors with antibody-type specificity. Proc. Natl. Acad. Sci. USA 1989, 86, 10024–10028. [Google Scholar] [CrossRef] [PubMed]
- Sadelain, M.; Brentjens, R.; Riviere, I. The promise and potential pitfalls of chimeric antigen receptors. Curr. Opin. Immunol. 2009, 21, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, E.; Kishi, H.; Ozawa, T.; Hamana, H.; Nakagawa, H.; Jin, A.; Lin, Z.; Muraguchi, A. A chimeric antigen receptor for TRAIL-receptor 1 induces apoptosis in various types of tumor cells. Biochem. Biophys. Res. Commun. 2014, 453, 798–803. [Google Scholar] [CrossRef] [PubMed]
- Roybal, K.T.; Rupp, L.J.; Morsut, L.; Walker, W.J.; McNally, K.A.; Park, J.S.; Lim, W.A. Precision Tumor Recognition by T Cells With Combinatorial Antigen-Sensing Circuits. Cell 2016, 164, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.; Goetsch, L.; Dumontet, C.; Corvaïa, N. Strategies and challenges for the next generation of antibody drug conjugates. Nat. Rev. Drug Discov. 2017, 16, 315–337. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.M.; Morris, C.Q. Antibody-Drug Conjugates (ADCs) for Personalized Treatment of Solid Tumors: A Review. Adv. Ther. 2017, 34, 1015–1035. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.Q.; Wang, H.B.; Xie, Z.M.; Li, Z.H.; Tang, X.J.; Xu, Y.C.; Zhang, C.; Naranmandura, H.; Chen, S.Q. Novel conjugation of tumor-necrosis-factor-related apoptosis-inducing ligand (TRAIL) with monomethyl auristatin E for efficient antitumor drug delivery. Adv. Mater. 2013, 25, 4718–4722. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.Q.; Zhao, W.B.; Lai, J.; Ding, D.; Wei, X.; Li, Y.; Liu, W.; Yang, X.; Xu, Y.; Chen, S. Hetero-modification of TRAIL trimer for improved drug delivery and in vivo antitumor activities. Sci. Rep. 2015, 5, 14872. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Yang, X.; Zhao, W.; Xu, Y.; Pan, L.; Chen, S. Optimizing Multistep Delivery of PEGylated Tumor-Necrosis-Factor-Related Apoptosis-Inducing Ligand-Toxin Conjugates for Improved Antitumor Activities. Bioconjug. Chem. 2017, 28, 2180–2189. [Google Scholar] [CrossRef] [PubMed]
- Pieczykolan, J.S.; Kubiński, K.; Masłyk, M.; Pawlak, S.D.; Pieczykolan, A.; Rózga, P.K.; Szymanik, M.; Gałązka, M.; Teska-Kamińska, M.; Żerek, B.; et al. AD-O53.2—A novel recombinant fusion protein combining the activities of TRAIL/Apo2L and Smac/Diablo, overcomes resistance of human cancer cells to TRAIL/Apo2L. Investig. New Drugs 2014, 32, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, T.; Zhang, N.; Yang, M.; Li, B.; Lü, X.; Cao, X.; Ling, C. Melittin, a major component of bee venom, sensitizes human hepatocellular carcinoma cells to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis by activating CaMKII-TAK1-JNK/p38 and inhibiting IkappaBalpha kinase-NFkappaB. J. Biol. Chem. 2009, 284, 3804–3813. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Han, Y.; Fu, H.; Liu, M.; Wu, J.; Chen, X.; Zhang, S.; Chen, Y. Construction and expression of sTRAIL-melittin combining enhanced anticancer activity with antibacterial activity in Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 2877–2884. [Google Scholar] [CrossRef] [PubMed]










© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubuisson, A.; Micheau, O. Antibodies and Derivatives Targeting DR4 and DR5 for Cancer Therapy. Antibodies 2017, 6, 16. https://doi.org/10.3390/antib6040016
Dubuisson A, Micheau O. Antibodies and Derivatives Targeting DR4 and DR5 for Cancer Therapy. Antibodies. 2017; 6(4):16. https://doi.org/10.3390/antib6040016
Chicago/Turabian StyleDubuisson, Agathe, and Olivier Micheau. 2017. "Antibodies and Derivatives Targeting DR4 and DR5 for Cancer Therapy" Antibodies 6, no. 4: 16. https://doi.org/10.3390/antib6040016
APA StyleDubuisson, A., & Micheau, O. (2017). Antibodies and Derivatives Targeting DR4 and DR5 for Cancer Therapy. Antibodies, 6(4), 16. https://doi.org/10.3390/antib6040016