
Neutralizing Activity of SARS-CoV-2 Antibodies in Patients with COVID-19 and Vaccinated Individuals

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Methods
2.3. Statistical Analysis
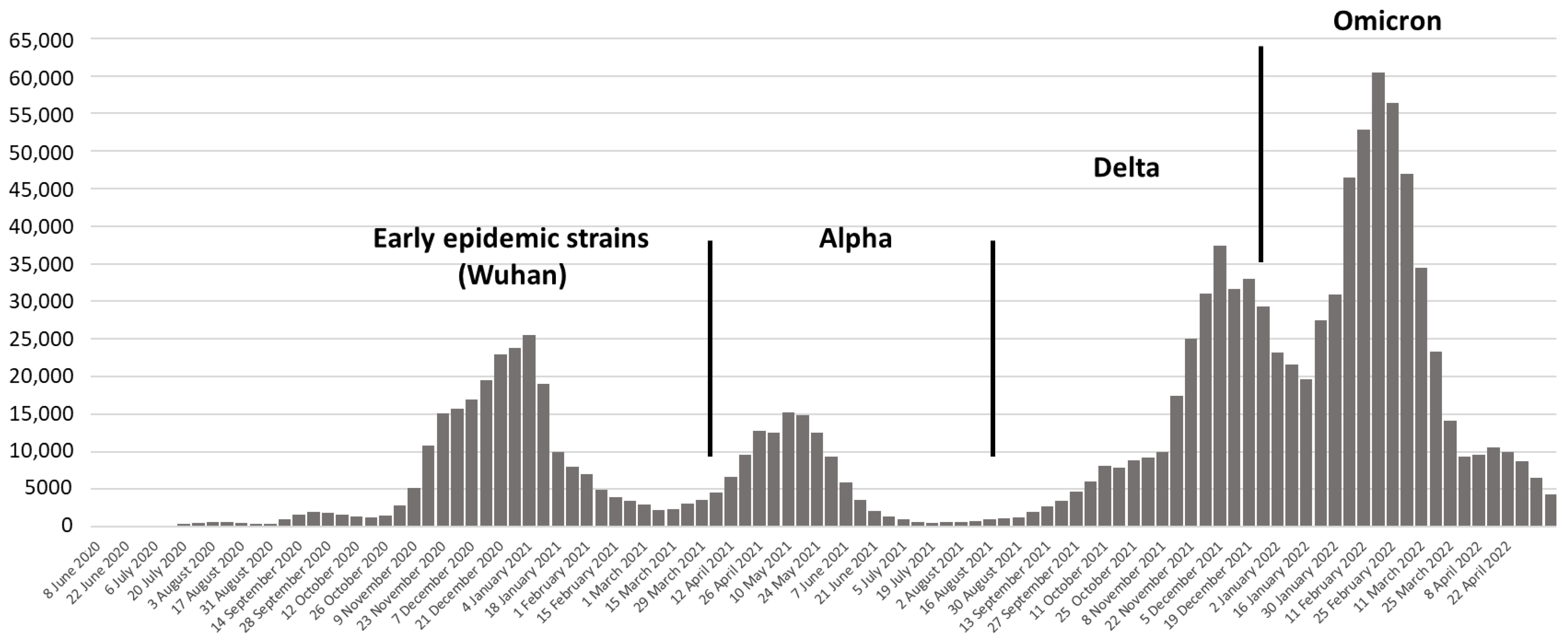
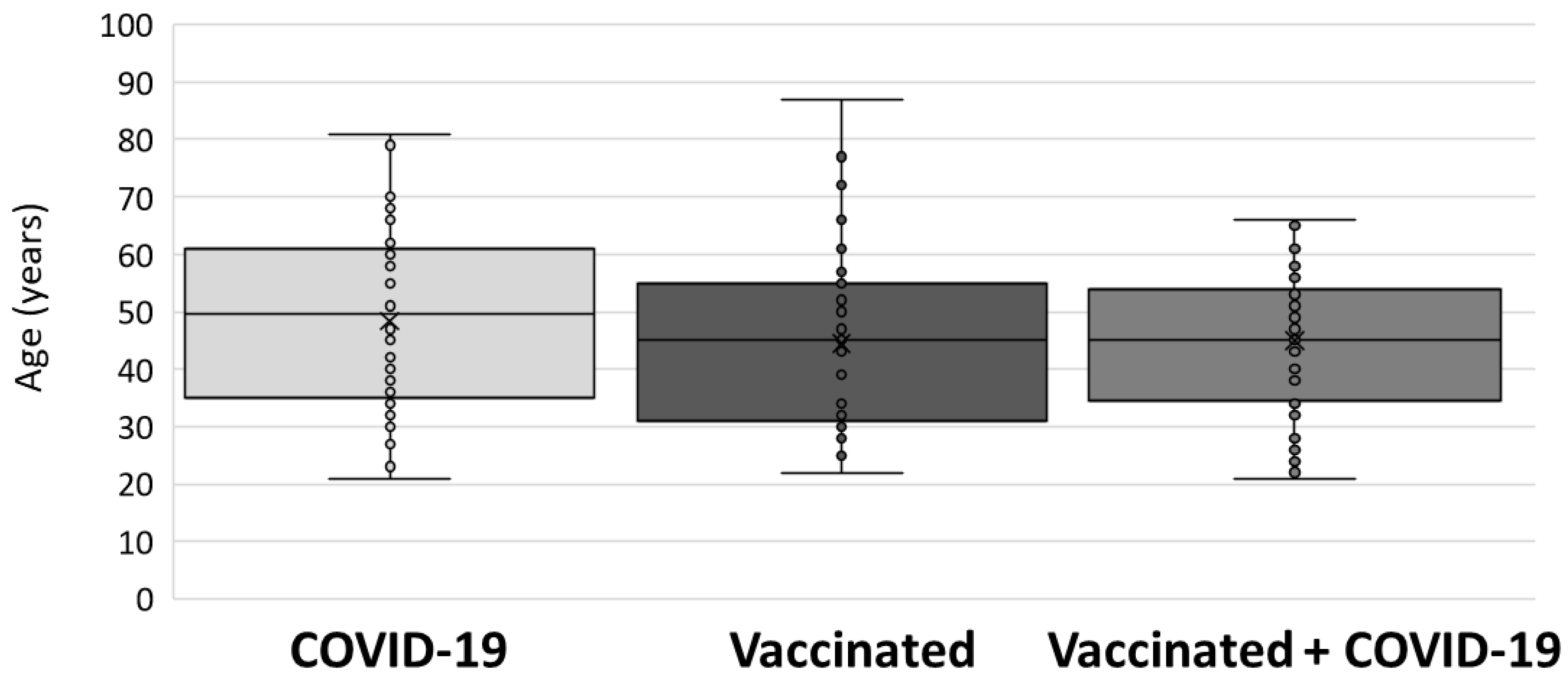
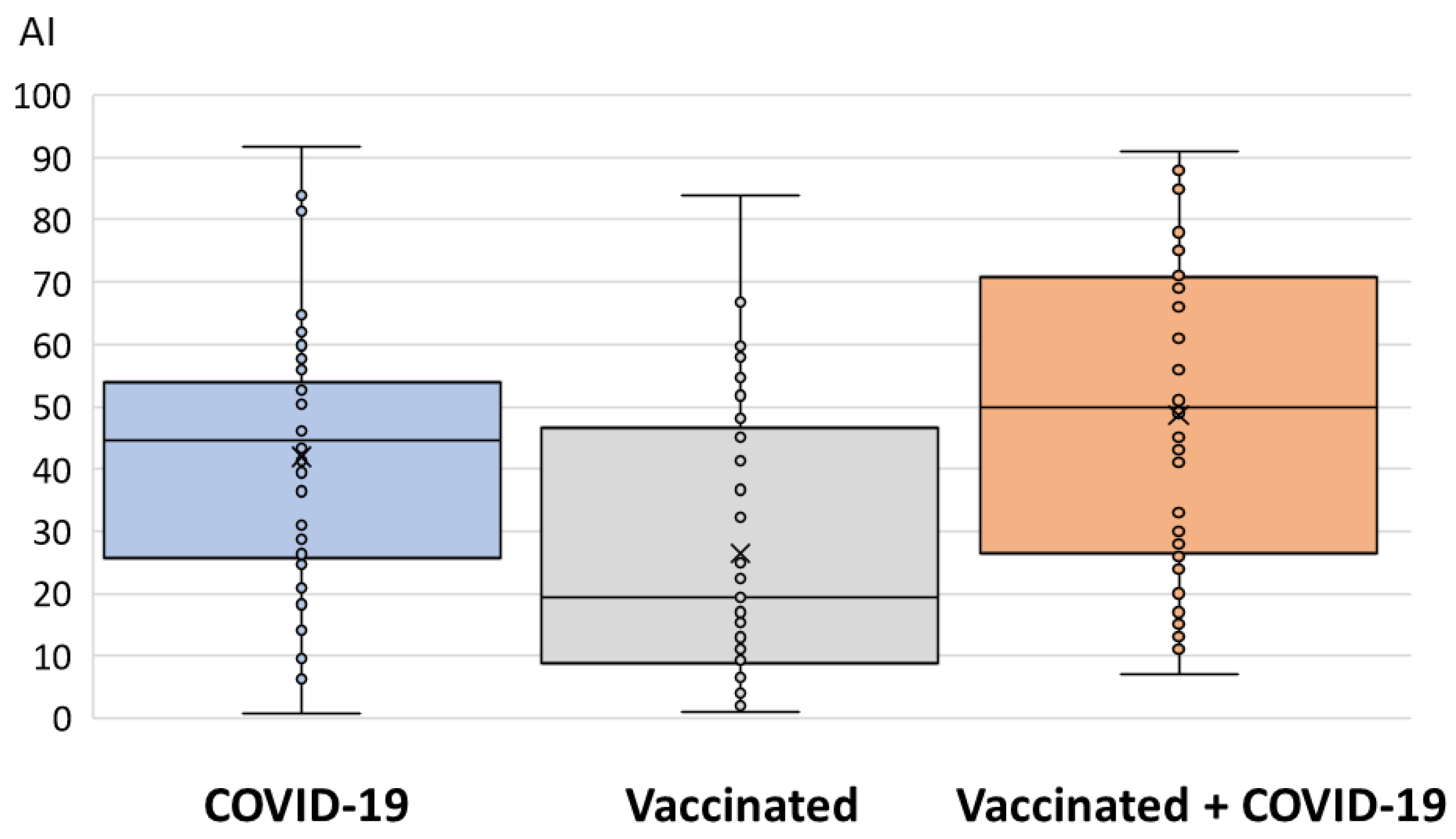
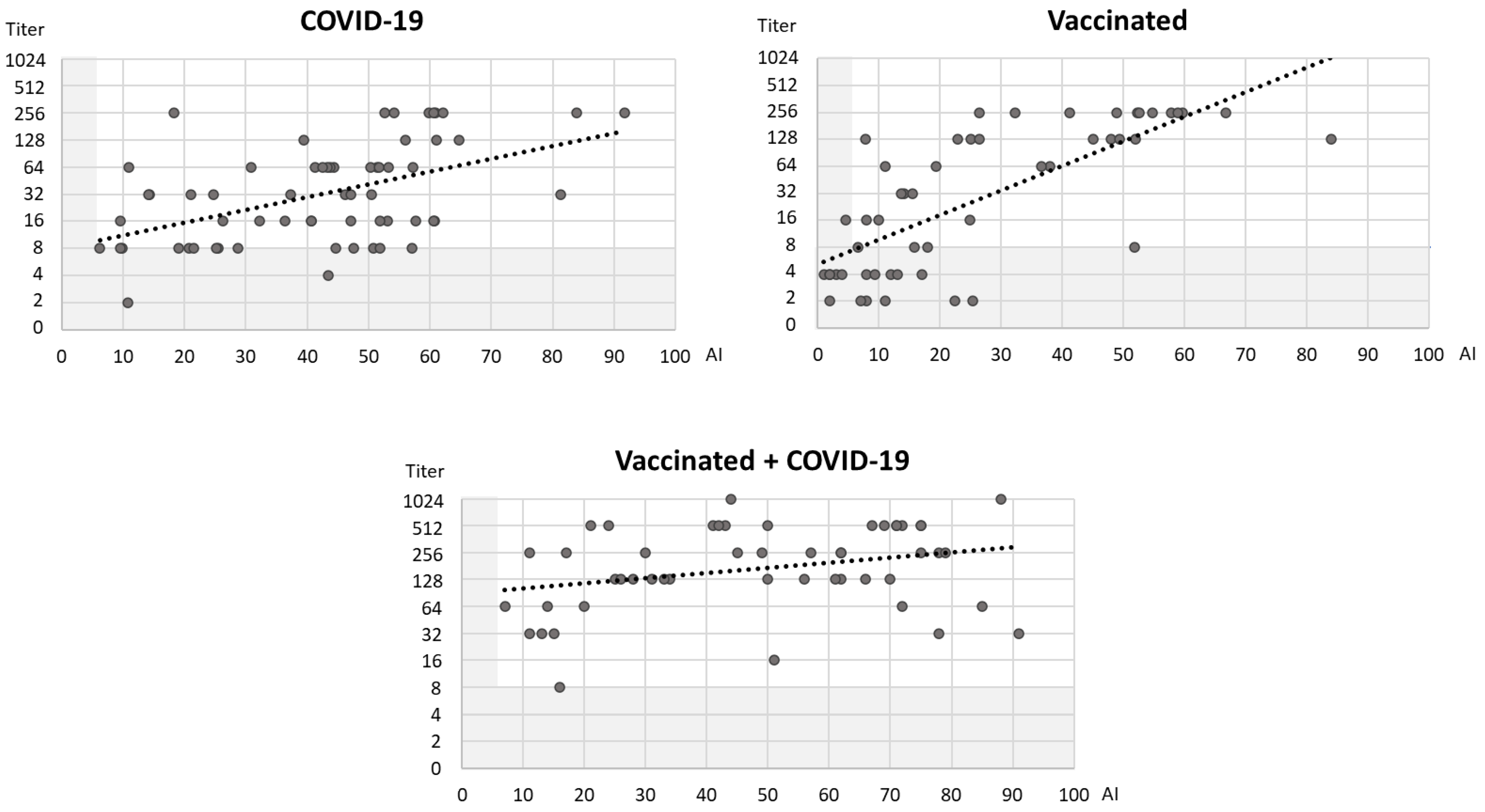
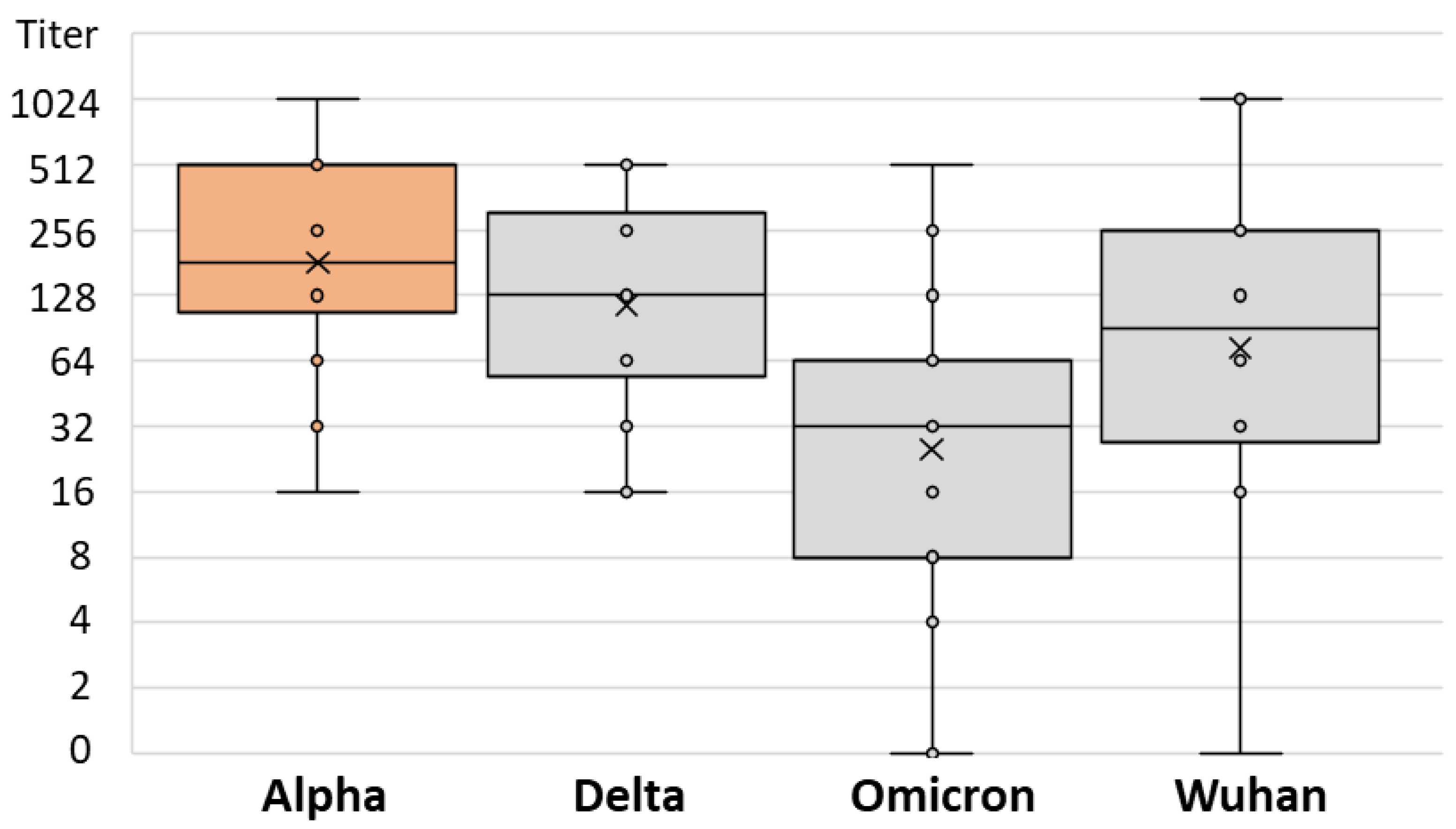
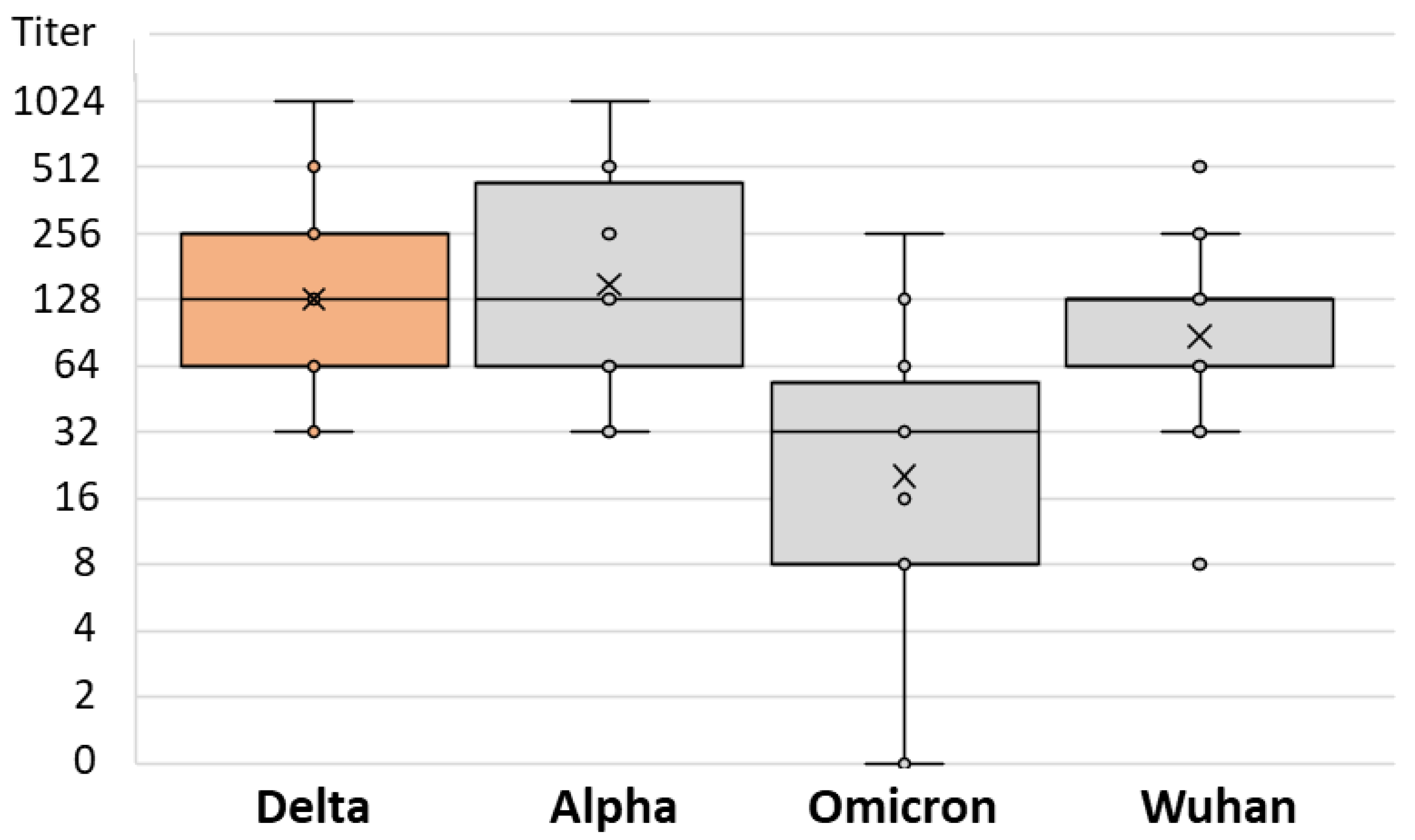
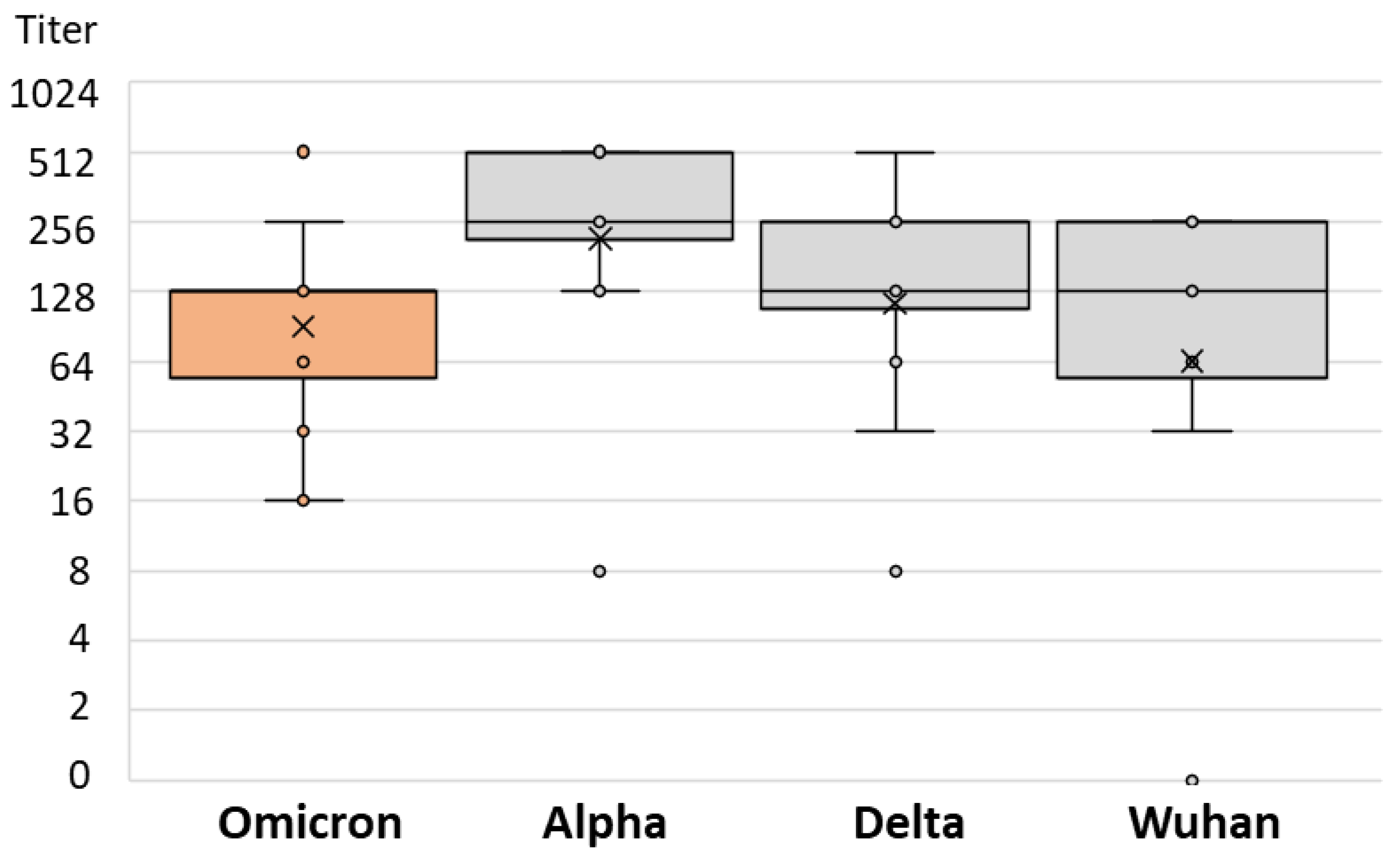
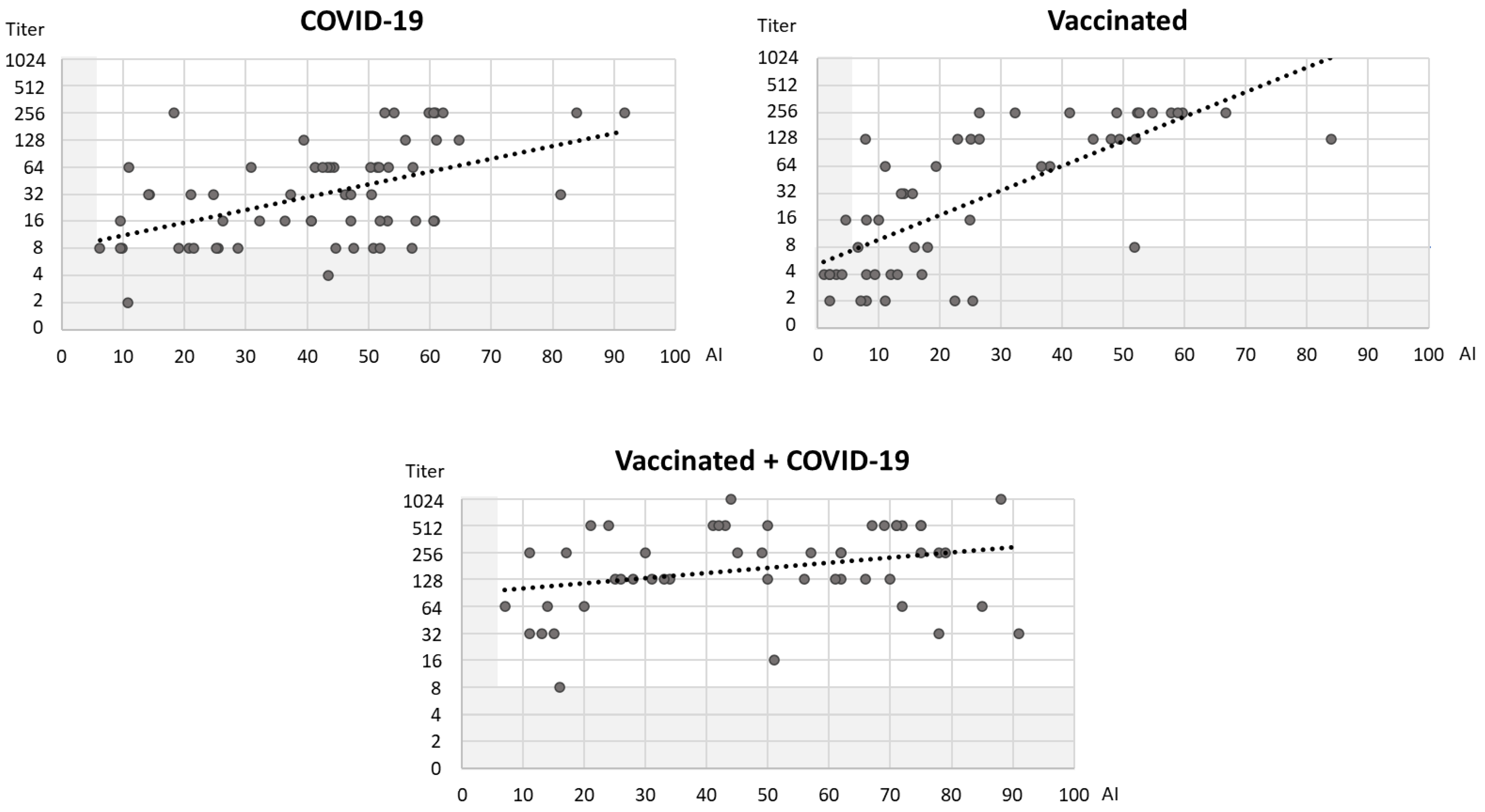
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 26 August 2023).
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S.W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Wang, M.Y.; Zhao, R.; Gao, L.J.; Gao, X.F.; Wang, D.P.; Cao, J.M. SARS-CoV-2: Structure, Biology, and Structure-Based Therapeutics Development. Front. Cell Infect. Microbiol. 2020, 10, 587269. [Google Scholar] [CrossRef] [PubMed]
- Gomari, M.M.; Tarighi, P.; Choupani, E.; Abkhiz, S.; Mohamadzadeh, M.; Rostami, N.; Sadroddiny, E.; Baammi, S.; Uversky, V.N.; Dokholyan, N.V. Structural evolution of Delta lineage of SARS-CoV-2. Int. J. Biol. Macromol. 2023, 226, 1116–1140. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lai, S.; Gao, G.F.; Shi, W. The emergence, genomic diversity and global spread of SARS-CoV-2. Nature 2021, 600, 408–418. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; Wenseleers, T.; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef] [PubMed]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.d.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.A.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Mlcochova, P.; Kemp, S.A.; Dhar, M.S.; Papa, G.; Meng, B.; Ferreira, I.A.T.M.; Datir, R.; Collier, D.A.; Albecka, A.; Singh, S.; et al. SARS-CoV-2 B.1.617.2 Delta variant replication and immune evasion. Nature 2021, 599, 114–119. [Google Scholar] [CrossRef]
- Wang, L.; Møhlenberg, M.; Wang, P.; Zhou, H. Immune Evasion of Neutralizing Antibodies by SARS-CoV-2 Omicron. Cytokine Growth Factor Rev. 2023, 70, 13–25. [Google Scholar] [CrossRef]
- Zhang, G.F.; Meng, W.; Chen, L.; Ding, L.; Feng, J.; Perez, J.; Ali, A.; Sun, S.; Liu, Z.; Huang, Y.; et al. Neutralizing Antibodies to SARS-CoV-2 Variants of Concern Including Delta and Omicron in Subjects Receiving MRNA-1273, BNT162b2, and Ad26.COV2.S Vaccines. J. Med. Virol. 2022, 94, 5678–5690. [Google Scholar] [CrossRef]
- Pango Lineages: Latest Epidemiological Lineages of SARS-CoV-2. Available online: https://cov-lineages.org/ (accessed on 18 September 2023).
- Volz, E.; Mishra, S.; Chand, M.; Barrett, J.C.; Johnson, R.; Geidelberg, L.; Hinsley, W.R.; Laydon, D.J.; Dabrera, G.; O’Toole, Á.; et al. COVID-19 Genomics UK (COG-UK) consortium. Assessing transmissibility of SARS-CoV-2 lineage B.1.1.7 in England. Nature 2021, 593, 266–269. [Google Scholar] [CrossRef]
- He, X.; Hong, W.; Pan, X.; Lu, G.; Wei, X. SARS-CoV-2 Omicron variant: Characteristics and prevention. Med. Commun. 2021, 2, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Salehi-Vaziri, M.; Fazlalipour, M.; Seyed Khorrami, S.M.; Azadmanesh, K.; Pouriayevali, M.H.; Jalali, T.; Shoja, Z.; Maleki, A. The ins and outs of SARS-CoV-2 variants of concern (VOCs). Arch. Virol. 2022, 167, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Duong, D. Alpha, Beta, Delta, Gamma: What’s important to know about SARS-CoV-2 variants of concern? CMAJ 2021, 193, E1059–E1060. [Google Scholar] [CrossRef]
- Alhamid, G.; Tombuloglu, H.; Rabaan, A.A.; Al-Suhaimi, E. SARS-CoV-2 detection methods: A comprehensive review. Saudi J. Biol. Sci. 2022, 29, 103465. [Google Scholar] [CrossRef]
- Perera, R.A.; Mok, C.K.; Tsang, O.T.; Lv, H.; Ko, R.L.; Wu, N.C.; Yuan, M.; Leung, W.S.; Chan, J.M.; Chik, T.S.; et al. Serological assays for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), March 2020. Euro Surveill. 2020, 25, 2000421. [Google Scholar] [CrossRef]
- Pieri, M.; Infantino, M.; Manfredi, M.; Nuccetelli, M.; Grossi, V.; Lari, B.; Tomassetti, F.; Sarubbi, S.; Russo, E.; Amedei, A.; et al. Performance evaluation of four surrogate Virus Neutralization Tests (sVNTs) in comparison to the in vivo gold standard test. Front. Biosci. 2022, 27, 74. [Google Scholar] [CrossRef]
- Pušnik, J.; Monzon-Posadas, W.O.; Zorn, J.; Peters, K.; Baum, M.; Proksch, H.; Schlüter, C.B.; Alter, G.; Menting, T.; Streeck, H.; et al. SARS-CoV-2 humoral and cellular immunity following different combinations of vaccination and breakthrough infection. Nat. Commun. 2023, 14, 572. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 183, 281–292. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced Sensitivity of SARS-CoV-2 Variant Delta to Antibody Neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef]
- Jørgensen, S.B.; Nygård, K.; Kacelnik, O.; Telle, K. Secondary Attack Rates for Omicron and Delta Variants of SARS-CoV-2 in Norwegian Households. JAMA 2022, 327, 1610–1611. [Google Scholar] [CrossRef]
- Hofmann, N.; Grossegesse, M.; Neumann, M.; Schaade, L.; Nitsche, A. Evaluation of a commercial ELISA as alternative to plaque reduction neutralization test to detect neutralizing antibodies against SARS-CoV-2. Sci. Rep. 2022, 12, 3549. [Google Scholar] [CrossRef]
- Chi, X.; Yan, R.; Zhang, J.; Zhang, G.; Zhang, Y.; Hao, M.; Zhang, Z.; Fan, P.; Dong, Y.; Yang, Y.; et al. A neutralizing human antibody binds to the N-terminal domain of the Spike protein of SARS-CoV-2. Science 2020, 369, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Yan, X.; Guo, X.; Cao, W.; Han, W.; Qi, C.; Feng, J.; Yang, D.; Gao, G.; Jin, G. A human SARS-CoV neutralizing antibody against epitope on S2 protein. Biochem. Biophys. Res. Commun. 2005, 333, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Stevanovic, V.; Brlek-Gorski, D.; Ferencak, I.; Ferenc, T.; Ujevic-Bosnjak, M.; Tabain, I.; Janev-Holcer, N.; Perkovic, I.; Anticevic, M. Emerging Trends in the Epidemiology of COVID-19: The Croatian ‘One Health’ Perspective. Viruses 2021, 13, 2354. [Google Scholar] [CrossRef] [PubMed]
- Ferenčak, I.; Kuzle, J.; Mišić, A.; Stevanović, V.; Bekavac, B.; Hruškar, Ž.; Vilibić-Čavlek, T.; Tabain, I. SARS-CoV-2 in Croatia-Genomic analysis of the third and fourth epidemic wave. In Proceedings of the 9th International Congress “Veterinary Science and Profession”, Zagreb, Croatia, 9 October 2021. [Google Scholar]
- Ferenčak, I.; Obrovac, M.; Žmak, L.; Kuzle, J.; Petrović, G.; Vilibić-Čavlek, T.; Jurić, D.; Jurić, A.; Hruškar, Ž.; Capak, K.; et al. SARS-CoV-2 Omicron Variant in Croatia-Rapid Detection of the First Case and Cross-Border Spread. Pathogens 2022, 11, 511. [Google Scholar] [CrossRef]
- Martínez-Flores, D.; Zepeda-Cervantes, J.; Cruz-Reséndiz, A.; Aguirre-Sampieri, S.; Sampieri, A.; Vaca, L. SARS-CoV-2 Vaccines Based on the Spike Glycoprotein and Implications of New Viral Variants. Front. Immunol. 2021, 12, 701501. [Google Scholar] [CrossRef]
- WHO. COMIRNATY® (Tozinameran), COVID-19 mRNA Vaccine (Nucleoside Modified)—Pfizer-BioNTech COVID-19 Vaccine. Available online: https://www.who.int/publications/m/item/comirnaty-covid-19-mrna-vaccine (accessed on 30 May 2023).
- Livingston, E.H.; Malani, P.N.; Creech, C.B. The Johnson & Johnson Vaccine for COVID-19. JAMA 2021, 325, 1575. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Bogdanic, M.; Borko, E.; Hruskar, Z.; Zilic, D.; Ferenc, T.; Tabain, I.; Barbic, L.; Vujica Ferenc, M.; Ferencak, I.; et al. Detection of SARS-CoV-2 Antibodies: Comparison of Enzyme Immunoassay, Surrogate Neutralization and Virus Neutralization Test. Antibodies 2023, 12, 35. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Laurie, M.T.; Liu, J.; Sunshine, S.; Peng, J.; Black, D.; Mitchell, A.M.; Mann, S.A.; Pilarowski, G.; Zorn, K.C.; Rubio, L.; et al. SARS-CoV-2 Variant Exposures Elicit Antibody Responses with Differential Cross-Neutralization of Established and Emerging Strains Including Delta and Omicron. J. Infect. Dis. 2022, 225, 1909–1914. [Google Scholar] [CrossRef]
- Changrob, S.; Fu, Y.; Guthmiller, J.J.; Halfmann, P.J.; Li, L.; Stamper, C.T.; Dugan, H.L.; Accola, M.; Rehrauer, W.; Zheng, N.Y.; et al. Cross-Neutralization of Emerging SARS-CoV-2 Variants of Concern by Antibodies Targeting Distinct Epitopes on Spike. mBio 2021, 12, e0297521. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.A.; McBride, S.K.; Leier, H.C.; Guzman, G.; Lyski, Z.L.; Schoen, D.; Winders, B.; Lee, J.Y.; Lee, D.X.; Messer, W.B.; et al. Vaccination before or after SARS-CoV-2 infection leads to robust humoral response and antibodies that effectively neutralize variants. Sci. Immunol. 2022, 7, eabn8014. [Google Scholar] [CrossRef] [PubMed]
- Bates, T.A.; McBride, S.K.; Winders, B.; Schoen, D.; Trautmann, L.; Curlin, M.E.; Tafesse, F.G. Antibody Response and Variant Cross-Neutralization after SARS-CoV-2 Breakthrough Infection. JAMA 2022, 327, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Suryawanshi, R.K.; Chen, I.P.; Ma, T.; Syed, A.M.; Brazer, N.; Saldhi, P.; Simoneau, C.R.; Ciling, A.; Khalid, M.M.; Sreekumar, B.; et al. Limited Cross-Variant Immunity after Infection with the SARS-CoV-2 Omicron Variant Without Vaccination. Nature 2022, 607, 351–355. [Google Scholar] [CrossRef]
- Marcotte, H.; Hammarström, L.; Pan-Hammarström, Q. Limited cross-variant neutralization after primary Omicron infection: Consideration for a variant-containing booster. Signal. Transduct. Target. Ther. 2022, 7, 294. [Google Scholar] [CrossRef]
- Servellita, V.; Syed, A.M.; Morris, M.K.; Brazer, N.; Saldhi, P.; Garcia-Knight, M.; Sreekumar, B.; Khalid, M.M.; Ciling, A.; Chen, P.-Y.; et al. Neutralizing Immunity in Vaccine Breakthrough Infections from the SARS-CoV-2 Omicron and Delta Variants. Cell 2022, 185, 1539–1548.e5. [Google Scholar] [CrossRef]
- Chakraborty, C.; Bhattacharya, M.; Sharma, A.R.; Mallik, B. Omicron (B.1.1.529)—A new heavily mutated variant: Mapped location and probable properties of its mutations with an emphasis on S-glycoprotein. Int. J. Biol. Macromol. 2022, 219, 980–997. [Google Scholar] [CrossRef]
- Yu, W.; Guo, Y.; Zhang, S.; Kong, Y.; Shen, Z.; Zhang, J. Proportion of asymptomatic infection and nonsevere disease caused by SARS-CoV-2 Omicron variant: A systematic review and analysis. J. Med. Virol. 2022, 94, 5790–5801. [Google Scholar] [CrossRef]
- Tober-Lau, P.; Gruell, H.; Vanshylla, K.; Koch, W.M.; Hillus, D.; Schommers, P.; Suárez, I.; Suttorp, N.; Sander, L.E.; Klein, F.; et al. Cross-Variant Neutralizing Serum Activity after SARS-CoV-2 Breakthrough Infections. Emerg. Infect. Dis. 2022, 28, 1050–1052. [Google Scholar] [CrossRef]
- Lavezzo, E.; Pacenti, M.; Manuto, L.; Boldrin, C.; Cattai, M.; Grazioli, M.; Bianca, F.; Sartori, M.; Caldart, F.; Castelli, G.; et al. Neutralising Reactivity against SARS-CoV-2 Delta and Omicron Variants by Vaccination and Infection History. Genome Med. 2022, 14, 61. [Google Scholar] [CrossRef]
- Villas-Boas, L.S.; Paula, A.V.D.; Silva, A.R.D.; Paiao, H.G.O.; Tozetto-Mendoza, T.R.; Manuli, E.R.; Leal, F.E.; Ferraz, A.D.B.C.; Sabino, E.C.; Bierrenbach, A.L.; et al. Absence of Neutralizing Antibodies against the Omicron SARS-CoV-2 Variant in Convalescent Sera from Individuals Infected with the Ancestral SARS-CoV-2 Virus or Its Gamma Variant. Clinics 2022, 77, 100068. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.Y.; Yang, J.S.; Hwang, S.Y.; Hyun, H.; Seong, H.; Gu Yoon, J.; Yoon, S.Y.; Cheong, H.J.; Kim, W.J.; Park, W.J.; et al. Duration of Humoral Immunity and Cross-Neutralizing Activity Against the Alpha, Beta, and Delta Variants after Wild-Type Severe Acute Respiratory Syndrome Coronavirus 2 Infection: A Prospective Cohort Study. J. Infect. Dis. 2022, 226, 975–978. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Lew, J.; Kroeker, A.; Baid, K.; Aftanas, P.; Nirmalarajah, K.; Maguire, F.; Kozak, R.; McDonald, R.; Lang, A.; et al. Immunogenicity of Convalescent and Vaccinated Sera against Clinical Isolates of Ancestral SARS-CoV-2, Beta, Delta, and Omicron Variants. Med 2022, 3, 422–432.e3. [Google Scholar] [CrossRef]
- Barnes, E.; Goodyear, C.S.; Willicombe, M.; Gaskell, C.; Siebert, S.; I de Silva, T.; Murray, S.M.; Rea, D.; Snowden, J.A.; Carroll, M.; et al. SARS-CoV-2-specific immune responses and clinical outcomes after COVID-19 vaccination in patients with immune-suppressive disease. Nat. Med. 2023, 29, 1760–1774. [Google Scholar] [CrossRef] [PubMed]
- Premuzic, V.; Stevanovic, R.; Vilibic-Cavlek, T.; Sirovica, M.; Stalman, S.; Bogdanic, M.; Zilic, D.; Nakic, D.; Santini Dusevic, D.; Vojkovic, M.; et al. Serological Response to SARS-CoV-2 Vaccine in Hemodialyzed Patients and the Association with Later COVID-19 Positivity. Antibodies 2023, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Kolarić, B.; Ambriović-Ristov, A.; Tabain, I.; Vilibić-Čavlek, T. Waning immunity six months after BioNTech/Pfizer COVID-19 vaccination among nursing home residents in Zagreb, Croatia. Croat. Med. J. 2021, 62, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Liew, M.Y.; Mathews, J.I.; Li, A.; Singh, R.; Jaramillo, S.A.; Weiss, Z.F.; Bowman, K.; Ankomah, P.O.; Ghantous, F.; Lewis, G.D.; et al. Delayed and Attenuated Antibody Responses to Coronavirus Disease 2019 Vaccination with Poor Cross-Variant Neutralization in Solid-Organ Transplant Recipients—A Prospective Longitudinal Study. Open Forum Infect. Dis. 2023, 10, ofad369. [Google Scholar] [CrossRef]
- Mathur, P.; Kottilil, S. Immunomodulatory therapies for COVID-19. Front. Med. 2022, 9, 921452. [Google Scholar] [CrossRef]
- Money, K.M.; Baber, U.; Saart, E.; Samaan, S.; Sloane, J.A. Blunted Post-COVID-19 Humoral Immunity in Patients with CNS Demyelinating Disorders on Anti-CD20 Treatments. Front. Neurol. 2022, 13, 843081. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Postvaccinal Infection | SARS-CoV-2 Strain | NT Antibodies N (%) | 95%CI | p |
|---|---|---|---|---|
| Alpha (n = 20) | Delta | 20 (100) | 83.1–100 * | |
| Omicron | 17 (85.0) | 69.3–100 | 0.217 | |
| Wuhan | 18 (90.0) | 76.8–100 | ||
| Delta (n = 18) | Alpha | 18 (100) | 81.5–100 * | |
| Omicron | 15 (83.3) | 58.6–96.4 | 0.041 | |
| Wuhan | 18 (100) | 81.5–100 * | ||
| Omicron (n = 12) | Alpha | 12 (100) | 73.5–100 * | |
| Delta | 12 (100) | 73.5–100 * | 0.357 | |
| Wuhan | 11 (91.7) | 61.5–99.8 |
| Postvaccinal Infection | Infection NT Titer Range | Infection Median NT Titer (IQR) | SARS-CoV-2 Strain | Strain NT Titer Range | Strain Median NT Titer (IQR) | p |
|---|---|---|---|---|---|---|
| Alpha | 16–1024 | 256 (128–512) | Delta | 16–512 | 128 (64–256) | 0.013 |
| Omicron | 0–512 | 32 (8–64) | ||||
| Wuhan | 0–1024 | 64 (16–128) | ||||
| Delta | 32–1024 | 128 (64–256) | Alpha | 32–1024 | 128 (64–512) | 0.001 |
| Omicron | 0–256 | 32 (8–64) | ||||
| Wuhan | 8–1024 | 64 (32–128) | ||||
| Omicron | 16–512 | 128 (64–128) | Alpha | 8–512 | 256 (192–512) | 0.029 |
| Delta | 32–256 | 128 (128–128) | ||||
| Wuhan | 0–256 | 128 (96–256) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilibic-Cavlek, T.; Stevanovic, V.; Kovac, S.; Borko, E.; Bogdanic, M.; Miletic, G.; Hruskar, Z.; Ferenc, T.; Coric, I.; Vujica Ferenc, M.; et al. Neutralizing Activity of SARS-CoV-2 Antibodies in Patients with COVID-19 and Vaccinated Individuals. Antibodies 2023, 12, 61. https://doi.org/10.3390/antib12040061
Vilibic-Cavlek T, Stevanovic V, Kovac S, Borko E, Bogdanic M, Miletic G, Hruskar Z, Ferenc T, Coric I, Vujica Ferenc M, et al. Neutralizing Activity of SARS-CoV-2 Antibodies in Patients with COVID-19 and Vaccinated Individuals. Antibodies. 2023; 12(4):61. https://doi.org/10.3390/antib12040061
Chicago/Turabian StyleVilibic-Cavlek, Tatjana, Vladimir Stevanovic, Snjezana Kovac, Ema Borko, Maja Bogdanic, Gorana Miletic, Zeljka Hruskar, Thomas Ferenc, Ivona Coric, Mateja Vujica Ferenc, and et al. 2023. "Neutralizing Activity of SARS-CoV-2 Antibodies in Patients with COVID-19 and Vaccinated Individuals" Antibodies 12, no. 4: 61. https://doi.org/10.3390/antib12040061
APA StyleVilibic-Cavlek, T., Stevanovic, V., Kovac, S., Borko, E., Bogdanic, M., Miletic, G., Hruskar, Z., Ferenc, T., Coric, I., Vujica Ferenc, M., Milasincic, L., Antolasic, L., & Barbic, L. (2023). Neutralizing Activity of SARS-CoV-2 Antibodies in Patients with COVID-19 and Vaccinated Individuals. Antibodies, 12(4), 61. https://doi.org/10.3390/antib12040061