

Determination of the Binding Epitope of an Anti-Mouse CCR9 Monoclonal Antibody (C9Mab-24) Using the 1× Alanine and 2× Alanine-Substitution Method

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Development of C9Mab-24
2.2. ELISA
3. Results
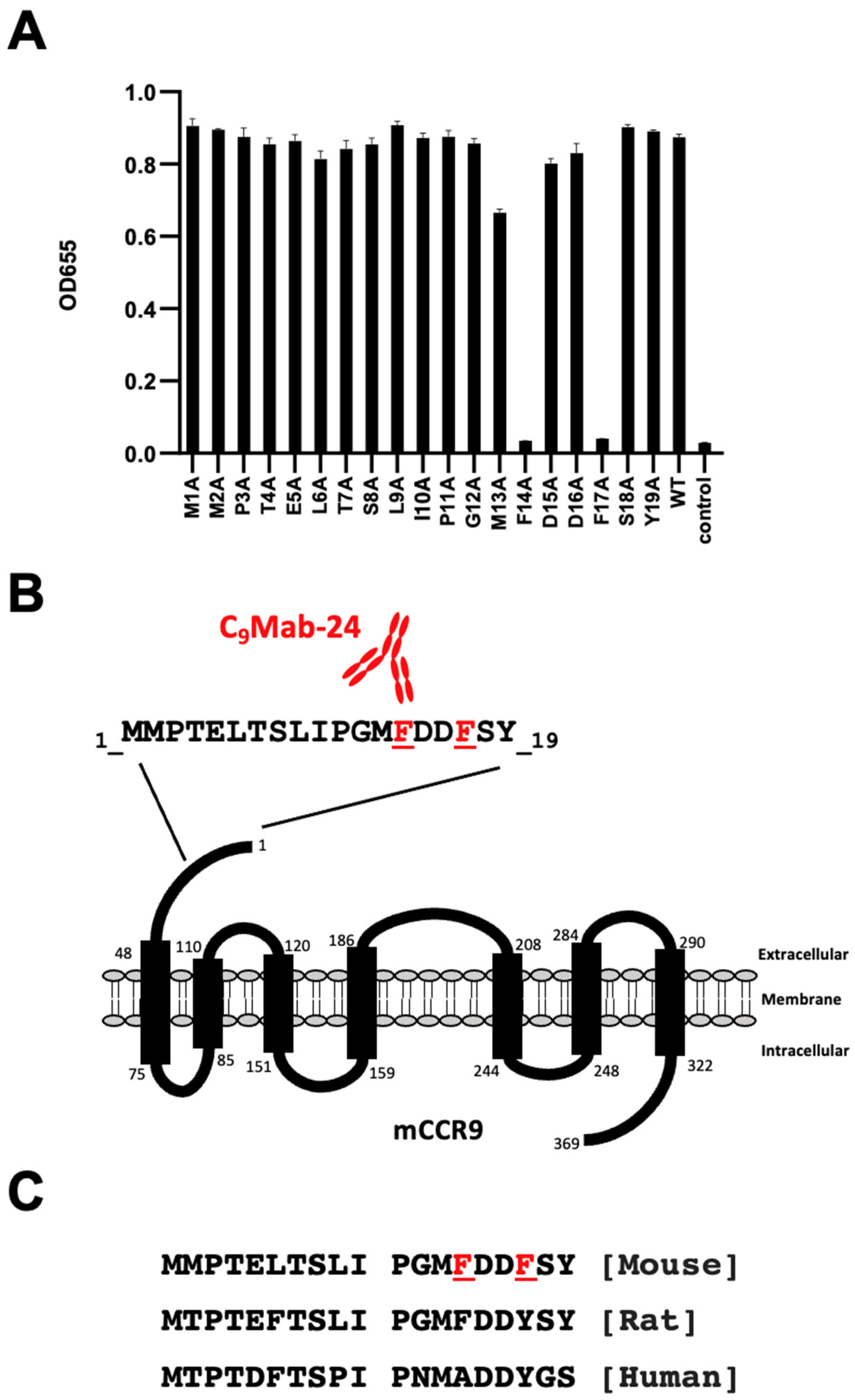
3.1. Epitope Determination Using 1× Ala-substituted mCCR9 Peptides
3.2. Epitope Determination Using 2× Ala-substituted mCCR9 Peptides
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wermers, J.D.; McNamee, E.N.; Wurbel, M.A.; Jedlicka, P.; Rivera-Nieves, J. The chemokine receptor CCR9 is required for the T-cell-mediated regulation of chronic ileitis in mice. Gastroenterology 2011, 140, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Doan, N.; Said, J.; Karunasiri, D.; Pullarkat, S.T. Strong expression of chemokine receptor CCR9 in diffuse large B-cell lymphoma and follicular lymphoma strongly correlates with gastrointestinal involvement. Hum. Pathol. 2014, 45, 1451–1458. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.A.; Agace, W.W.; Campbell, J.J.; Heath, H.M.; Parent, D.; Roberts, A.I.; Ebert, E.C.; Kassam, N.; Qin, S.; Zovko, M.; et al. Human G protein-coupled receptor GPR-9-6/CC chemokine receptor 9 is selectively expressed on intestinal homing T lymphocytes, mucosal lymphocytes, and thymocytes and is required for thymus-expressed chemokine-mediated chemotaxis. J. Exp. Med. 1999, 190, 1241–1256. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.J.; Crown, S.E.; Handel, T.M. Chemokine: Receptor structure, interactions, and antagonism. Annu. Rev. Immunol. 2007, 25, 787–820. [Google Scholar] [CrossRef] [PubMed]
- Tu, Z.; Xiao, R.; Xiong, J.; Tembo, K.M.; Deng, X.; Xiong, M.; Liu, P.; Wang, M.; Zhang, Q. CCR9 in cancer: Oncogenic role and therapeutic targeting. J. Hematol. Oncol. 2016, 9, 10. [Google Scholar] [CrossRef]
- Kunkel, E.J.; Campbell, J.J.; Haraldsen, G.; Pan, J.; Boisvert, J.; Roberts, A.I.; Ebert, E.C.; Vierra, M.A.; Goodman, S.B.; Genovese, M.C.; et al. Lymphocyte CC chemokine receptor 9 and epithelial thymus-expressed chemokine (TECK) expression distinguish the small intestinal immune compartment: Epithelial expression of tissue-specific chemokines as an organizing principle in regional immunity. J. Exp. Med. 2000, 192, 761–768. [Google Scholar] [CrossRef]
- Papadakis, K.A.; Prehn, J.; Nelson, V.; Cheng, L.; Binder, S.W.; Ponath, P.D.; Andrew, D.P.; Targan, S.R. The role of thymus-expressed chemokine and its receptor CCR9 on lymphocytes in the regional specialization of the mucosal immune system. J. Immunol. 2000, 165, 5069–5076. [Google Scholar] [CrossRef]
- Igaki, K.; Komoike, Y.; Nakamura, Y.; Watanabe, T.; Yamasaki, M.; Fleming, P.; Yang, L.; Soler, D.; Fedyk, E.; Tsuchimori, N. MLN3126, an antagonist of the chemokine receptor CCR9, ameliorates inflammation in a T cell mediated mouse colitis model. Int. Immunopharmacol. 2018, 60, 160–169. [Google Scholar] [CrossRef]
- López-Pacheco, C.; Soldevila, G.; Du Pont, G.; Hernández-Pando, R.; García-Zepeda, E.A. CCR9 Is a Key Regulator of Early Phases of Allergic Airway Inflammation. Mediat. Inflamm. 2016, 2016, 3635809. [Google Scholar] [CrossRef]
- Wurbel, M.A.; McIntire, M.G.; Dwyer, P.; Fiebiger, E. CCL25/CCR9 interactions regulate large intestinal inflammation in a murine model of acute colitis. PLoS ONE 2011, 6, e16442. [Google Scholar] [CrossRef]
- Wurbel, M.A.; Le Bras, S.; Ibourk, M.; Pardo, M.; McIntire, M.G.; Coco, D.; Geha, R.S.; Fiebiger, E.; Snapper, S.B. CCL25/CCR9 interactions are not essential for colitis development but are required for innate immune cell protection from chronic experimental murine colitis. Inflamm. Bowel. Dis. 2014, 20, 1165–1176. [Google Scholar] [CrossRef]
- Wendt, E.; Keshav, S. CCR9 antagonism: Potential in the treatment of Inflammatory Bowel Disease. Clin. Exp. Gastroenterol. 2015, 8, 119–130. [Google Scholar] [CrossRef]
- Amiya, T.; Nakamoto, N.; Chu, P.S.; Teratani, T.; Nakajima, H.; Fukuchi, Y.; Taniki, N.; Yamaguchi, A.; Shiba, S.; Miyake, R.; et al. Bone marrow-derived macrophages distinct from tissue-resident macrophages play a pivotal role in Concanavalin A-induced murine liver injury via CCR9 axis. Sci. Rep. 2016, 6, 35146. [Google Scholar] [CrossRef]
- Nakamoto, N. [Role of inflammatory macrophages and CCR9/CCL25 chemokine axis in the pathogenesis of liver injury as a therapeutic target]. Nihon Rinsho Meneki Gakkai Kaishi 2016, 39, 460–467. [Google Scholar] [CrossRef]
- Lu, L.; Du, H.; Huang, H.; Wang, C.; Wang, P.; Zha, Z.; Wu, Y.; Liu, X.; Weng, C.; Fang, X.; et al. CCR9 Promotes Migration and Invasion of Lung Adenocarcinoma Cancer Stem Cells. Int. J. Med. Sci. 2020, 17, 912–920. [Google Scholar] [CrossRef]
- Johnson-Holiday, C.; Singh, R.; Johnson, E.; Singh, S.; Stockard, C.R.; Grizzle, W.E.; Lillard, J.W., Jr. CCL25 mediates migration, invasion and matrix metalloproteinase expression by breast cancer cells in a CCR9-dependent fashion. Int. J. Oncol. 2011, 38, 1279–1285. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, T.; Chen, Y.; Gong, S.; Sun, X.; Zou, F.; Peng, R. CCL25/CCR9 Signal Promotes Migration and Invasion in Hepatocellular and Breast Cancer Cell Lines. DNA Cell Biol. 2016, 35, 348–357. [Google Scholar] [CrossRef]
- Singh, R.; Stockard, C.R.; Grizzle, W.E.; Lillard, J.W., Jr.; Singh, S. Expression and histopathological correlation of CCR9 and CCL25 in ovarian cancer. Int. J. Oncol. 2011, 39, 373–381. [Google Scholar] [CrossRef]
- Fusi, A.; Liu, Z.; Kümmerlen, V.; Nonnemacher, A.; Jeske, J.; Keilholz, U. Expression of chemokine receptors on circulating tumor cells in patients with solid tumors. J. Transl. Med. 2012, 10, 52. [Google Scholar] [CrossRef]
- Kühnelt-Leddihn, L.; Müller, H.; Eisendle, K.; Zelger, B.; Weinlich, G. Overexpression of the chemokine receptors CXCR4, CCR7, CCR9, and CCR10 in human primary cutaneous melanoma: A potential prognostic value for CCR7 and CCR10? Arch. Dermatol. Res. 2012, 304, 185–193. [Google Scholar] [CrossRef]
- Maciocia, P.M.; Wawrzyniecka, P.A.; Maciocia, N.C.; Burley, A.; Karpanasamy, T.; Devereaux, S.; Hoekx, M.; O’Connor, D.; Leon, T.; Rapoz-D’Silva, T.; et al. Anti-CCR9 chimeric antigen receptor T cells for T-cell acute lymphoblastic leukemia. Blood 2022, 140, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, Z.; Zhong, Y.; Lan, J.; Li, X.; Lin, H. CCR9-CCL25 interaction suppresses apoptosis of lung cancer cells by activating the PI3K/Akt pathway. Med. Oncol. 2015, 32, 66. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Mailey, B.; Ellenhorn, J.D.; Chu, P.G.; Lowy, A.M.; Kim, J. CC chemokine receptor 9 enhances proliferation in pancreatic intraepithelial neoplasia and pancreatic cancer cells. J. Gastrointest. Surg. 2009, 13, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Somovilla-Crespo, B.; Martín Monzón, M.T.; Vela, M.; Corraliza-Gorjón, I.; Santamaria, S.; Garcia-Sanz, J.A.; Kremer, L. 92R Monoclonal Antibody Inhibits Human CCR9(+) Leukemia Cells Growth in NSG Mice Xenografts. Front. Immunol. 2018, 9, 77. [Google Scholar] [CrossRef]
- Chamorro, S.; Vela, M.; Franco-Villanueva, A.; Carramolino, L.; Gutiérrez, J.; Gómez, L.; Lozano, M.; Salvador, B.; García-Gallo, M.; Martínez, A.C.; et al. Antitumor effects of a monoclonal antibody to human CCR9 in leukemia cell xenografts. MAbs 2014, 6, 1000–1012. [Google Scholar] [CrossRef]
- Wu, X.; Sun, M.; Yang, Z.; Lu, C.; Wang, Q.; Wang, H.; Deng, C.; Liu, Y.; Yang, Y. The Roles of CCR9/CCL25 in Inflammation and Inflammation-Associated Diseases. Front. Cell Dev. Biol. 2021, 9, 686548. [Google Scholar] [CrossRef]
- Tanaka, T.; Li, G.; Asano, T.; Saito, M.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Development of a Novel Anti-Mouse CCR2 Monoclonal Antibody (C(2)Mab-6) by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 80–86. [Google Scholar] [CrossRef]
- Asano, T.; Suzuki, H.; Goto, N.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Establishment of Novel Anti-Mouse CCR3 Monoclonal Antibodies (C(3)Mab-6 and C(3)Mab-7) by N-terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 94–100. [Google Scholar] [CrossRef]
- Takei, J.; Suzuki, H.; Asano, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-Mouse CCR4 Monoclonal Antibody (C(4)Mab-1) by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 87–93. [Google Scholar] [CrossRef]
- Asano, T.; Tanaka, T.; Suzuki, H.; Li, G.; Nanamiya, R.; Tateyama, N.; Isoda, Y.; Okada, Y.; Kobayashi, H.; Yoshikawa, T.; et al. Development of a Novel Anti-Mouse CCR6 Monoclonal Antibody (C(6)Mab-13) by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 343–349. [Google Scholar] [CrossRef]
- Kitamura, K.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Cx(6)Mab-1: A Novel Anti-Mouse CXCR6 Monoclonal Antibody Established by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 133–141. [Google Scholar] [CrossRef]
- Kobayashi, H.; Asano, T.; Suzuki, H.; Tanaka, T.; Yoshikawa, T.; Kaneko, M.K.; Kato, Y. Establishment of a Sensitive Monoclonal Antibody Against Mouse CCR9 (C9Mab-24) for Flow Cytometry. Monoclon. Antibodies Immunodiagn. Immunother. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Isoda, Y.; Tanaka, T.; Suzuki, H.; Asano, T.; Nakamura, T.; Yanaka, M.; Handa, S.; Komatsu, Y.; Okuno, S.; Takahashi, N.; et al. Epitope Mapping of an Anti-Mouse CXCR6 Monoclonal Antibody (Cx(6)Mab-1) Using the 2 × Alanine Scanning Method. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 275–278. [Google Scholar] [CrossRef]
- Jin, L.; Fendly, B.M.; Wells, J.A. High resolution functional analysis of antibody-antigen interactions. J. Mol. Biol. 1992, 226, 851–865. [Google Scholar] [CrossRef]
- Asano, T.; Suzuki, H.; Tanaka, T.; Saito, M.; Li, G.; Goto, N.; Nanamiya, R.; Kaneko, M.K.; Kato, Y. C(3)Mab-3: A Monoclonal Antibody for Mouse CC Chemokine Receptor 3 for Flow Cytometry. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 74–79. [Google Scholar] [CrossRef]
- Asano, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Tanaka, T.; Sano, M.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 3 Monoclonal Antibodies for Flow Cytometry. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 107–112. [Google Scholar] [CrossRef]
- Suzuki, H.; Saito, M.; Asano, T.; Tanaka, T.; Kitamura, K.; Kudo, Y.; Kaneko, M.K.; Kato, Y. C(8)Mab-3: An Anti-Mouse CCR8 Monoclonal Antibody for Immunocytochemistry. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 110–114. [Google Scholar] [CrossRef]
- Saito, M.; Tanaka, T.; Asano, T.; Nakamura, T.; Yanaka, M.; Handa, S.; Komatsu, Y.; Harigae, Y.; Tateyama, N.; Nanamiya, R.; et al. C(8)Mab-2: An Anti-Mouse C-C Motif Chemokine Receptor 8 Monoclonal Antibody for Immunocytochemistry. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 115–119. [Google Scholar] [CrossRef]
- Saito, M.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of an Anti-Mouse CCR8 Monoclonal Antibody (C(8)Mab-1) for Flow Cytometry and Immunocytochemistry. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 333–338. [Google Scholar] [CrossRef]
- Nanamiya, R.; Takei, J.; Asano, T.; Tanaka, T.; Sano, M.; Nakamura, T.; Yanaka, M.; Hosono, H.; Kaneko, M.K.; Kato, Y. Development of Anti-Human CC Chemokine Receptor 9 Monoclonal Antibodies for Flow Cytometry. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 101–106. [Google Scholar] [CrossRef]
- Takei, J.; Asano, T.; Li, G.; Saito, M.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of an Anti-Human CCR9 Monoclonal Antibody (C(9)Mab-1) Using Enzyme-Linked Immunosorbent Assay. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Suzuki, H.; Isoda, Y.; Asano, T.; Nakamura, T.; Yanaka, M.; Handa, S.; Takahashi, N.; Okuno, S.; Yoshikawa, T.; et al. Development of a Sensitive Anti-Human CCR9 Monoclonal Antibody (C(9)Mab-11) by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Maude, S.L.; Laetsch, T.W.; Buechner, J.; Rives, S.; Boyer, M.; Bittencourt, H.; Bader, P.; Verneris, M.R.; Stefanski, H.E.; Myers, G.D.; et al. Tisagenlecleucel in Children and Young Adults with B-Cell Lymphoblastic Leukemia. N. Engl. J. Med. 2018, 378, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Gower, M.; Tikhonova, A.N. Avoiding fratricide: A T-ALL order. Blood 2022, 140, 3–4. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Peptides | Sequences | C9Mab-24 |
|---|---|---|
| WT | MMPTELTSLIPGMFDDFSY | ++ |
| M1A | AMPTELTSLIPGMFDDFSY | ++ |
| M2A | MAPTELTSLIPGMFDDFSY | ++ |
| P3A | MMATELTSLIPGMFDDFSY | ++ |
| T4A | MMPAELTSLIPGMFDDFSY | ++ |
| E5A | MMPTALTSLIPGMFDDFSY | ++ |
| L6A | MMPTEATSLIPGMFDDFSY | ++ |
| T7A | MMPTELASLIPGMFDDFSY | ++ |
| S8A | MMPTELTALIPGMFDDFSY | ++ |
| L9A | MMPTELTSAIPGMFDDFSY | ++ |
| I10A | MMPTELTSLAPGMFDDFSY | ++ |
| P11A | MMPTELTSLIAGMFDDFSY | ++ |
| G12A | MMPTELTSLIPAMFDDFSY | ++ |
| M13A | MMPTELTSLIPGAFDDFSY | ++ |
| F14A | MMPTELTSLIPGMADDFSY | – |
| D15A | MMPTELTSLIPGMFADFSY | ++ |
| D16A | MMPTELTSLIPGMFDAFSY | ++ |
| F17A | MMPTELTSLIPGMFDDASY | – |
| S18A | MMPTELTSLIPGMFDDFAY | ++ |
| Y19A | MMPTELTSLIPGMFDDFSA | ++ |
| Peptides | Sequences | C9Mab-24 |
|---|---|---|
| WT | MMPTELTSLIPGMFDDFSY | ++ |
| M1A–M2A | AAPTELTSLIPGMFDDFSY | ++ |
| M2A–P3A | MAATELTSLIPGMFDDFSY | ++ |
| P3A–T4A | MMAAELTSLIPGMFDDFSY | ++ |
| T4A–E5A | MMPAALTSLIPGMFDDFSY | ++ |
| E5A–L6A | MMPTAATSLIPGMFDDFSY | ++ |
| L6A–T7A | MMPTEAASLIPGMFDDFSY | ++ |
| T7A–S8A | MMPTELAALIPGMFDDFSY | ++ |
| S8A–L9A | MMPTELTAAIPGMFDDFSY | ++ |
| L9A–I10A | MMPTELTSAAPGMFDDFSY | ++ |
| I10A–P11A | MMPTELTSLAAGMFDDFSY | ++ |
| P11A–G12A | MMPTELTSLIAAMFDDFSY | ++ |
| G12A–M13A | MMPTELTSLIPAAFDDFSY | ++ |
| M13A–F14A | MMPTELTSLIPGAADDFSY | – |
| F14A–D15A | MMPTELTSLIPGMAADFSY | – |
| D15A–D16A | MMPTELTSLIPGMFAAFSY | ++ |
| D16A–F17A | MMPTELTSLIPGMFDAASY | – |
| F17A–S18A | MMPTELTSLIPGMFDDAAY | – |
| S18A–Y19A | MMPTELTSLIPGMFDDFAA | ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, H.; Asano, T.; Tanaka, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Determination of the Binding Epitope of an Anti-Mouse CCR9 Monoclonal Antibody (C9Mab-24) Using the 1× Alanine and 2× Alanine-Substitution Method. Antibodies 2023, 12, 11. https://doi.org/10.3390/antib12010011
Kobayashi H, Asano T, Tanaka T, Suzuki H, Kaneko MK, Kato Y. Determination of the Binding Epitope of an Anti-Mouse CCR9 Monoclonal Antibody (C9Mab-24) Using the 1× Alanine and 2× Alanine-Substitution Method. Antibodies. 2023; 12(1):11. https://doi.org/10.3390/antib12010011
Chicago/Turabian StyleKobayashi, Hiyori, Teizo Asano, Tomohiro Tanaka, Hiroyuki Suzuki, Mika K. Kaneko, and Yukinari Kato. 2023. "Determination of the Binding Epitope of an Anti-Mouse CCR9 Monoclonal Antibody (C9Mab-24) Using the 1× Alanine and 2× Alanine-Substitution Method" Antibodies 12, no. 1: 11. https://doi.org/10.3390/antib12010011
APA StyleKobayashi, H., Asano, T., Tanaka, T., Suzuki, H., Kaneko, M. K., & Kato, Y. (2023). Determination of the Binding Epitope of an Anti-Mouse CCR9 Monoclonal Antibody (C9Mab-24) Using the 1× Alanine and 2× Alanine-Substitution Method. Antibodies, 12(1), 11. https://doi.org/10.3390/antib12010011