
Epitope Mapping of Anti-Mouse CCR3 Monoclonal Antibodies Using Flow Cytometry

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Plasmid Construction
2.3. Antibodies
2.4. Flow Cytometry
3. Results
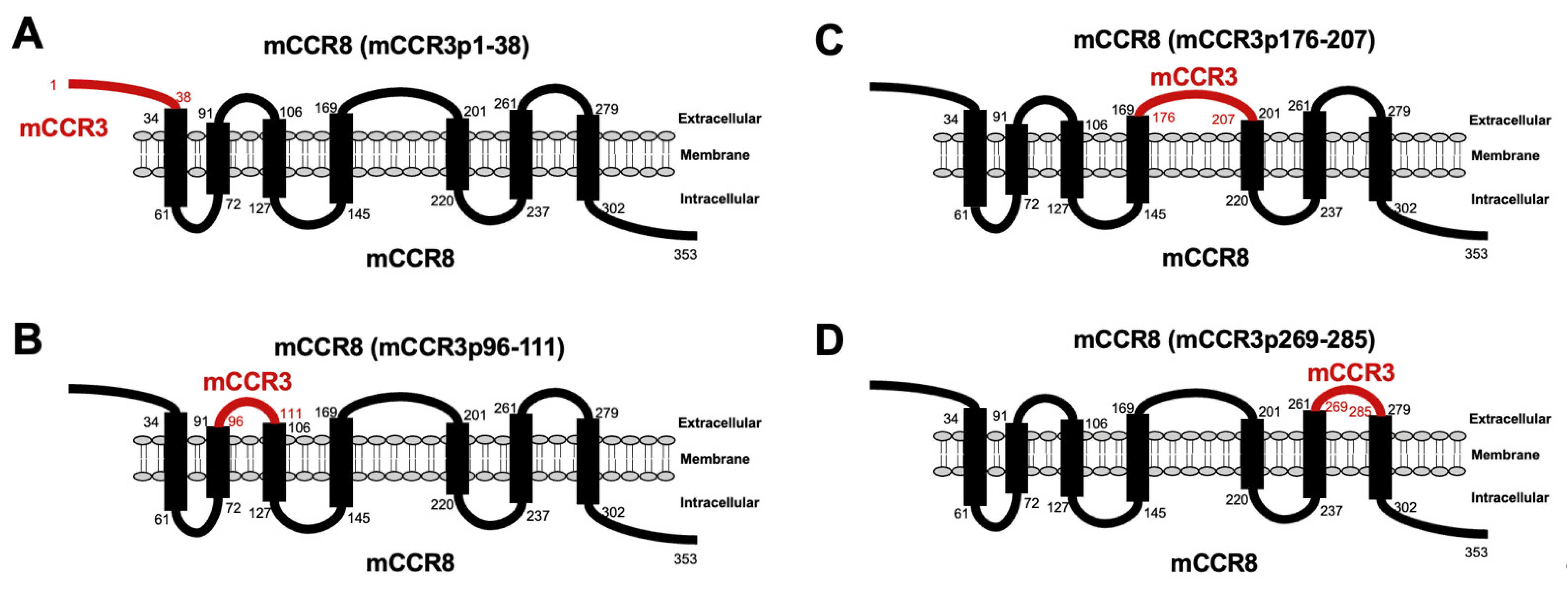
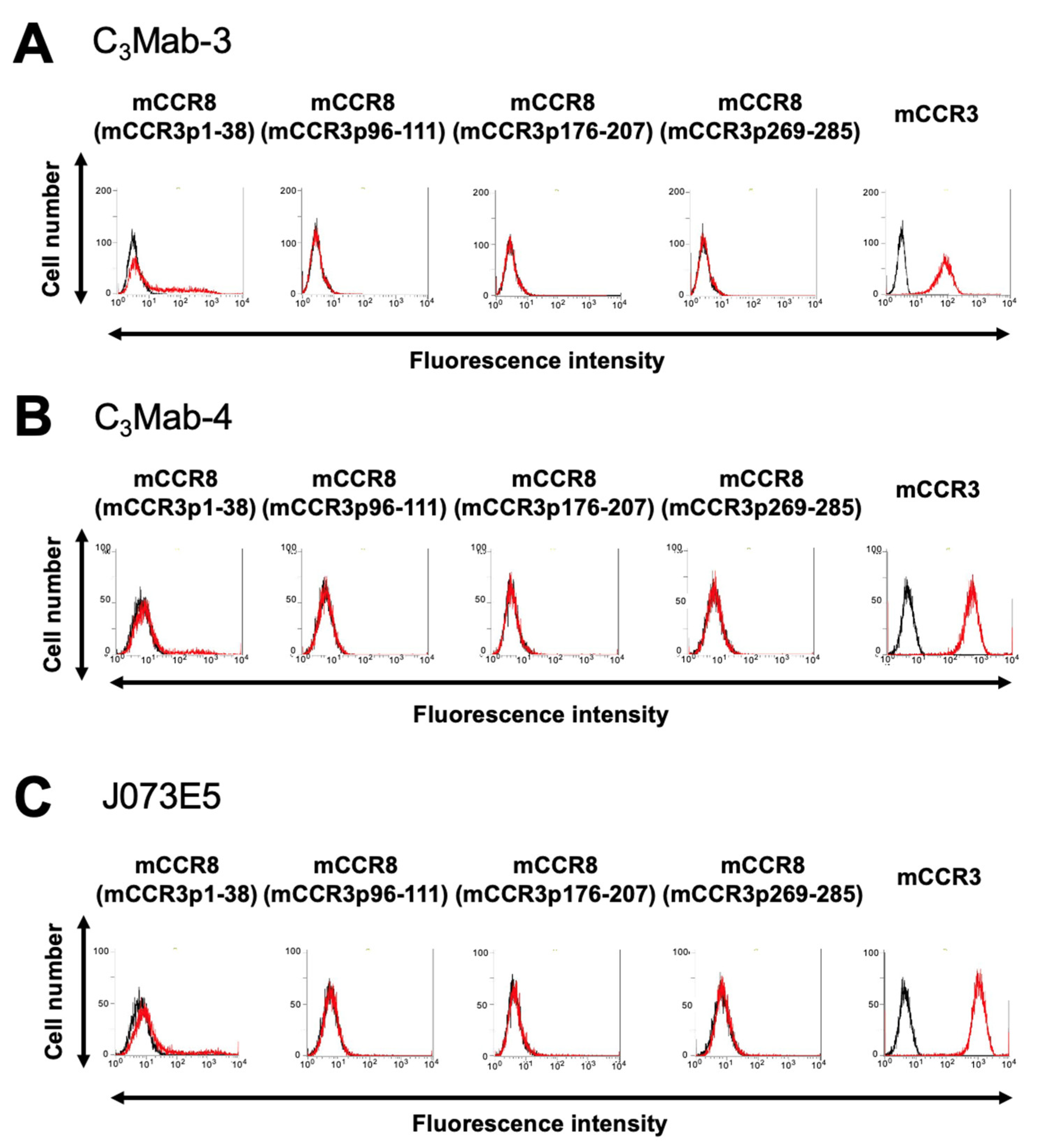
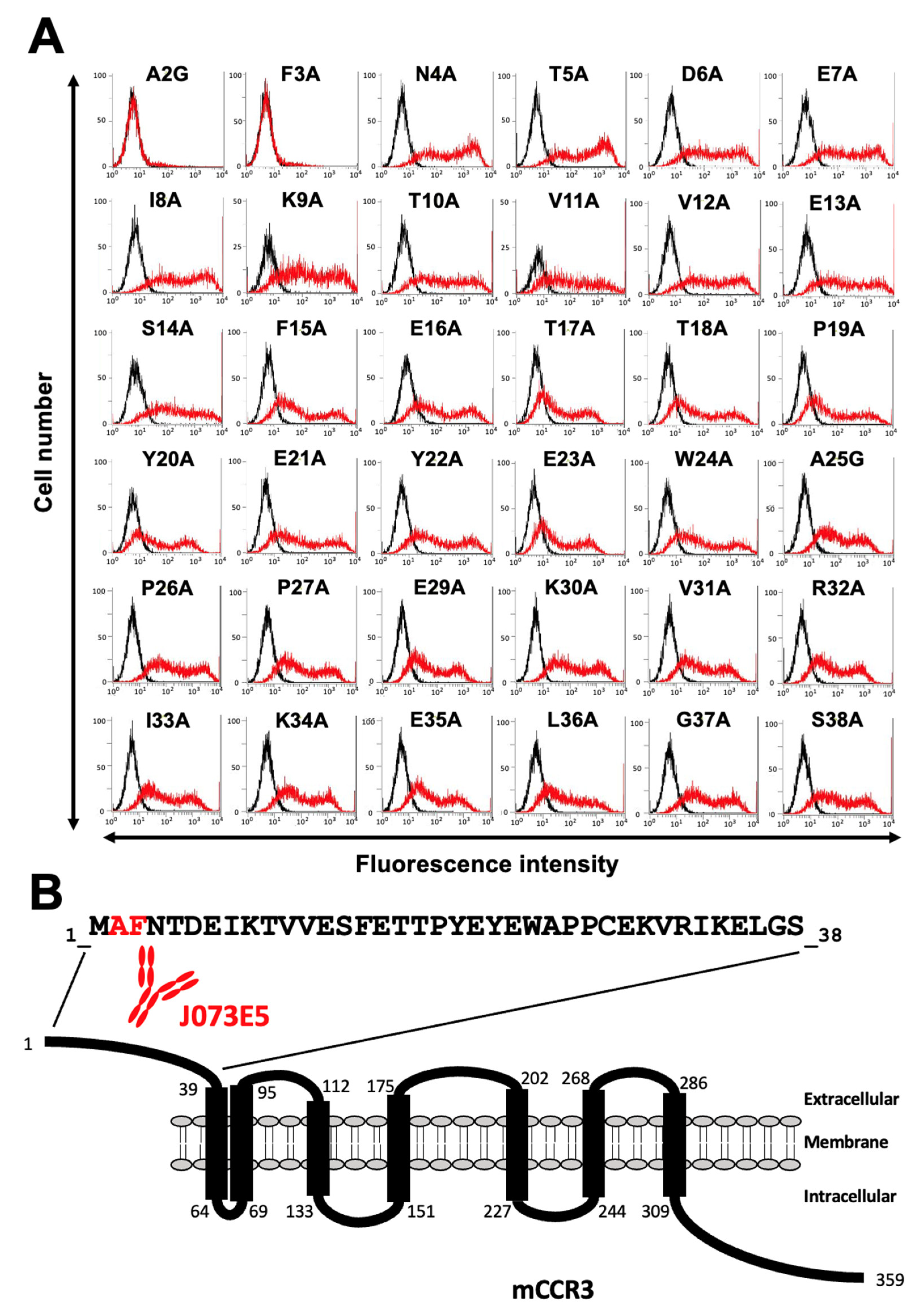
3.1. Determination of the Epitope of Anti-mCCR3 mAbs Using Flow Cytometry and Chimeric Proteins
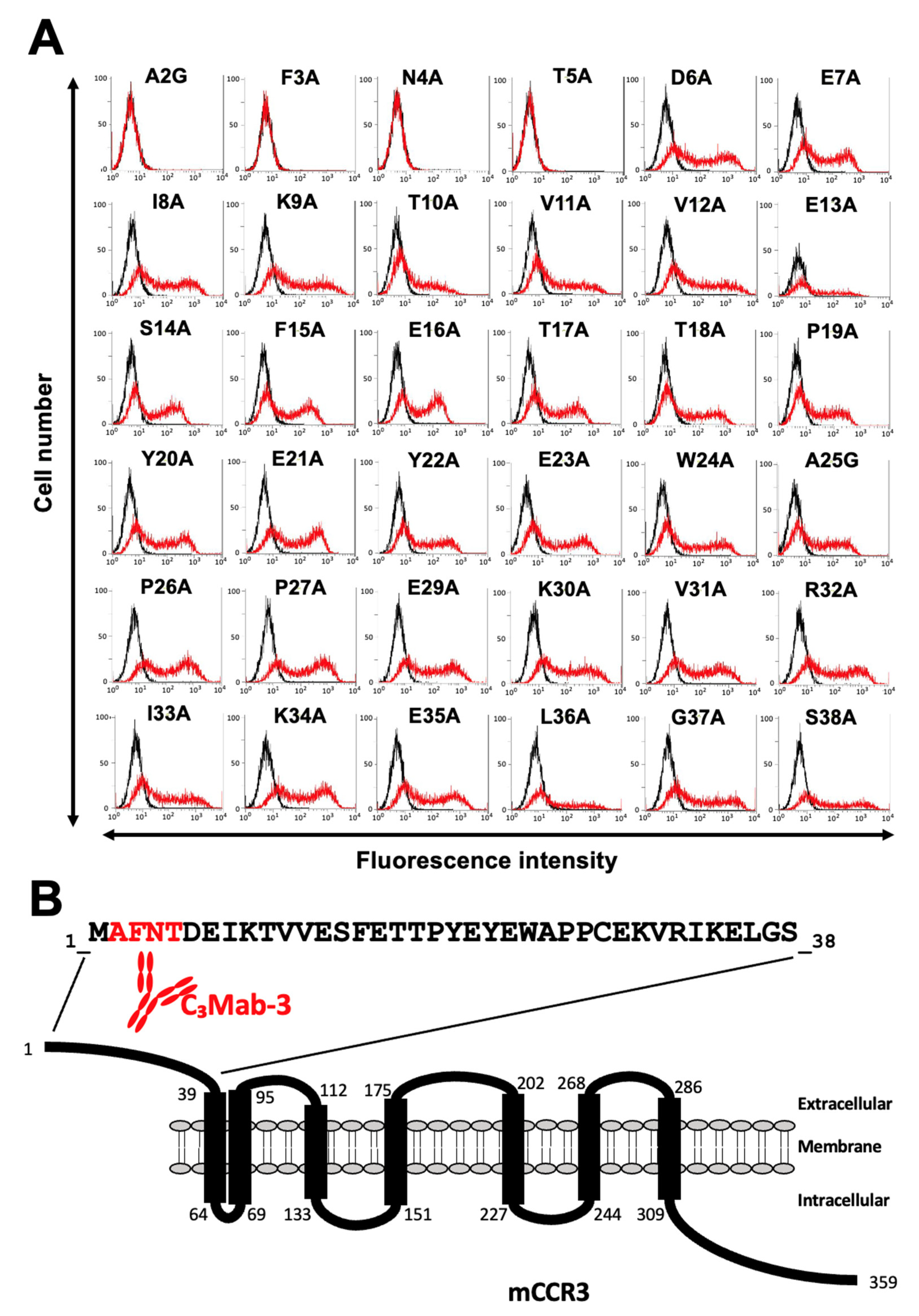
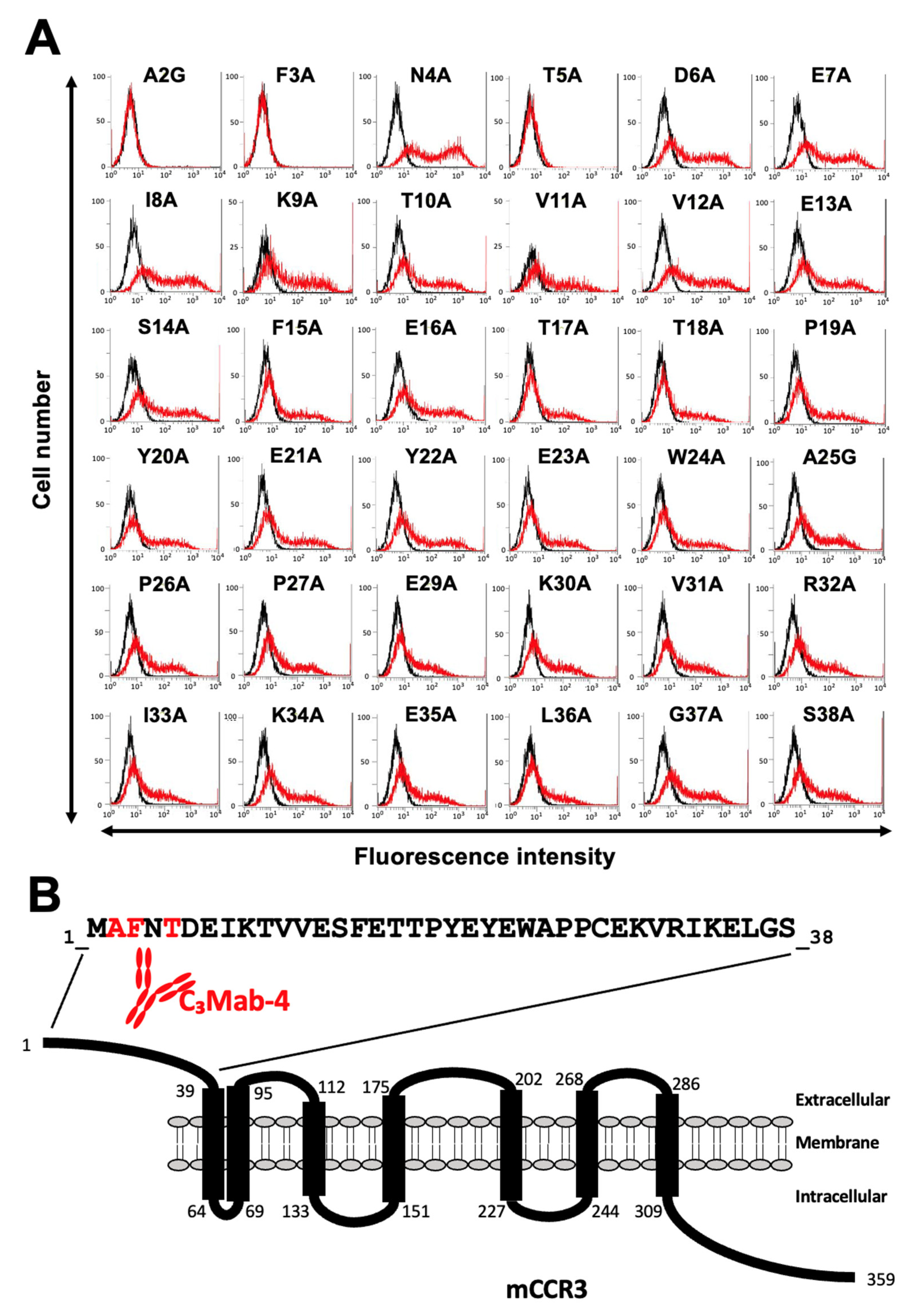
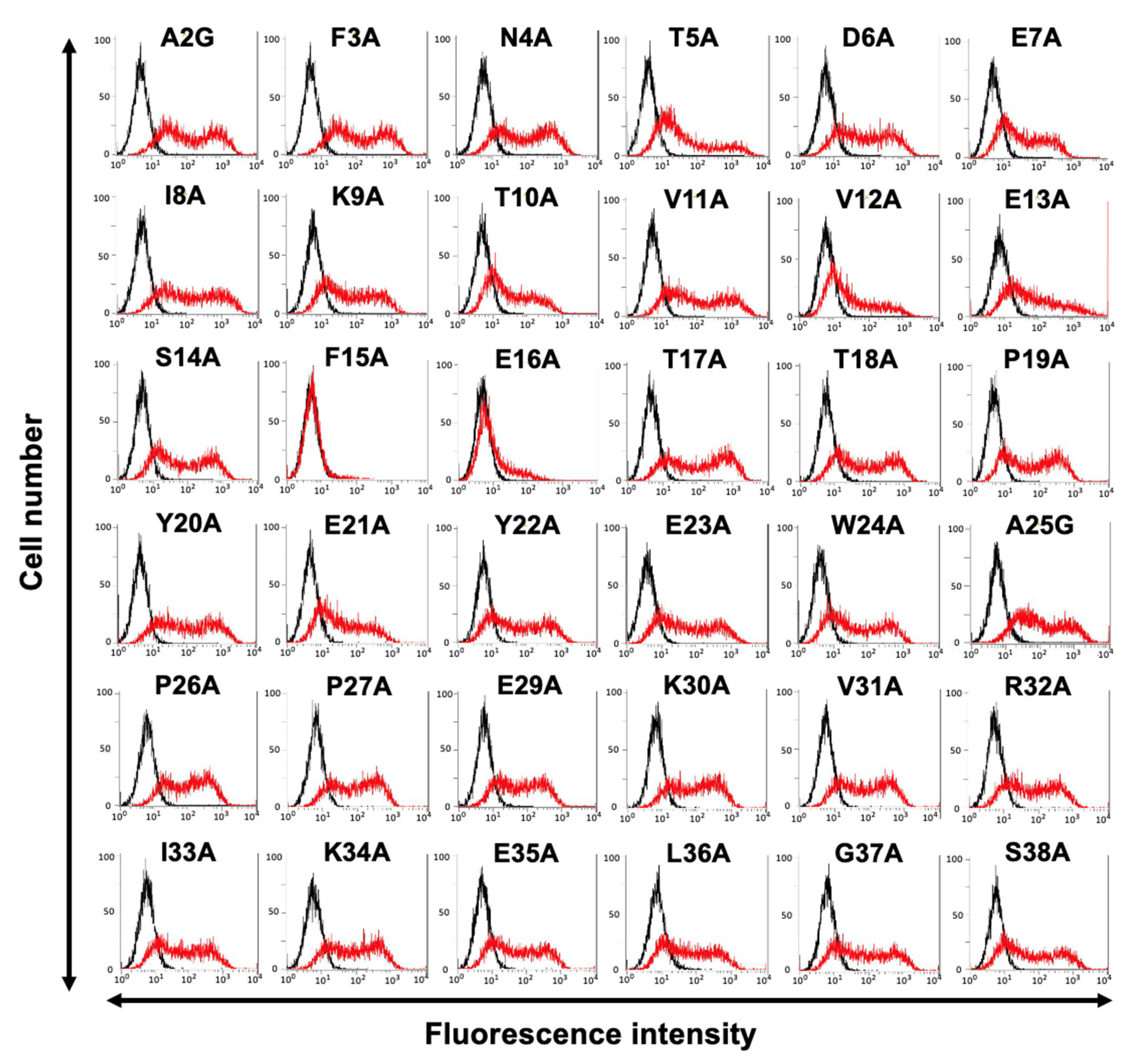
3.2. Determination of the C3Mab-3 Epitope Using Flow Cytometry and Alanine Scanning
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kufareva, I.; Salanga, C.L.; Handel, T.M. Chemokine and chemokine receptor structure and interactions: Implications for therapeutic strategies. Immunol. Cell Biol. 2015, 93, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Yoshie, O. Chemokines: A New Classification System and Their Role in Immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Charo, I.F.; Ransohoff, R.M. The Many Roles of Chemokines and Chemokine Receptors in Inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.M.; Baggiolini, M.; Charo, I.F.; Hébert, C.A.; Horuk, R.; Matsushima, K.; Miller, L.H.; Oppenheim, J.J.; Power, C.A. International union of pharmacology. XXII. Nomenclature for chemokine receptors. Pharmacol. Rev. 2000, 52, 145–176. [Google Scholar] [PubMed]
- Wise, E.L.; Duchesnes, C.; da Fonseca, P.; Allen, R.A.; Williams, T.J.; Pease, J.E. Small Molecule Receptor Agonists and Antagonists of CCR3 Provide Insight into Mechanisms of Chemokine Receptor Activation. J. Biol. Chem. 2007, 282, 27935–27943. [Google Scholar] [CrossRef] [PubMed]
- Bisset, L.R.; Schmid-Grendelmeier, P. Chemokines and their receptors in the pathogenesis of allergic asthma: Progress and perspective. Curr. Opin. Pulm. Med. 2005, 11, 35–42. [Google Scholar] [CrossRef]
- Mackay, C.R. Moving targets: Cell migration inhibitors as new anti-inflammatory therapies. Nat. Immunol. 2008, 9, 988–998. [Google Scholar] [CrossRef]
- Willems, L.I.; Ijzerman, A.P. Small molecule antagonists for chemokine CCR3 receptors. Med. Res. Rev. 2010, 30, 778–817. [Google Scholar] [CrossRef]
- Stellato, C.; Brummet, M.E.; Plitt, J.R.; Shahabuddin, S.; Baroody, F.M.; Liu, M.C.; Ponath, P.D.; Beck, L.A. Cutting Edge: Expression of the C-C Chemokine Receptor CCR3 in Human Airway Epithelial Cells. J. Immunol. 2001, 166, 1457–1461. [Google Scholar] [CrossRef]
- He, J.; Chen, Y.; Farzan, M.; Choe, H.; Ohagen, A.; Gartner, S.; Busciglio, J.; Yang, X.; Hofmann, W.; Newman, W.; et al. CCR3 and CCR5 are co-receptors for HIV-1 infection of microglia. Nature 1997, 385, 645–649. [Google Scholar] [CrossRef]
- Heath, H.; Qin, S.; Rao, P.; Wu, L.; LaRosa, G.; Kassam, N.; Ponath, P.D.; Mackay, C. Chemokine receptor usage by human eosinophils. The importance of CCR3 demonstrated using an antagonistic monoclonal antibody. J. Clin. Investig. 1997, 99, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Mackay, C.R.; Lanzavecchia, A. Selective Expression of the Eotaxin Receptor CCR3 by Human T Helper 2 Cells. Science 1997, 277, 2005–2007. [Google Scholar] [CrossRef] [PubMed]
- New, D.C.; Wong, Y.H. CC chemokine receptor-coupled signalling pathways. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao 2003, 35, 779–788. [Google Scholar] [PubMed]
- Kroeze, W.K.; Sheffler, D.J.; Roth, B.L. G-protein-coupled receptors at a glance. J. Cell Sci. 2003, 116, 4867–4869. [Google Scholar] [CrossRef] [PubMed]
- Kampen, G.T.; Stafford, S.; Adachi, T.; Jinquan, T.; Quan, S.; Grant, J.A.; Skov, P.S.; Poulsen, L.K.; Alam, R. Eotaxin induces degranulation and chemotaxis of eosinophils through the activation of ERK2 and p38 mitogen-activated protein kinases. Blood 2000, 95, 1911–1917. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Bertics, P.J. Chemoattractant-Induced Signaling via the Ras–ERK and PI3K–Akt Networks, along with Leukotriene C4 Release, Is Dependent on the Tyrosine Kinase Lyn in IL-5– and IL-3–Primed Human Blood Eosinophils. J. Immunol. 2011, 186, 516–526. [Google Scholar] [CrossRef]
- Fulkerson, P.C.; Fischetti, C.A.; McBride, M.L.; Hassman, L.M.; Hogan, S.P.; Rothenberg, M.E. A central regulatory role for eosinophils and the eotaxin/CCR3 axis in chronic experimental allergic airway inflammation. Proc. Natl. Acad. Sci. USA 2006, 103, 16418–16423. [Google Scholar] [CrossRef]
- Fukuda, K.; Kuo, C.-H.; Morohoshi, K.; Liu, F.-T.; Ono, S.J. The murine CCR3 receptor regulates both eosinophilia and hyperresponsiveness in IgE-mediated allergic conjunctivitis. Br. J. Ophthalmol. 2012, 96, 1132–1136. [Google Scholar] [CrossRef]
- Chantry, D.; Burgess, L.E. Chemokines in Allergy. Curr. Drug Targets-Inflamm. Allergy 2002, 1, 109–116. [Google Scholar] [CrossRef]
- Pease, J.E. Asthma, Allergy and Chemokines. Curr. Drug Targets 2006, 7, 3–12. [Google Scholar] [CrossRef]
- Miyazaki, D.; Nakamura, T.; Komatsu, N.; Nawata, N.; Ikeda, Y.; Inoue, Y.; Higashi, H.; Ono, S.J. Roles of Chemokines in Ocular Allergy and Possible Therapeutic Strategies. Cornea 2004, 23, S48–S54. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Lim, S.-J.; Won, K.Y.; Bae, G.; Kim, G.Y.; Min, J.W.; Noh, B.-J. Eosinophils in Colorectal Neoplasms Associated with Expression of CCL11 and CCL24. J. Pathol. Transl. Med. 2016, 50, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.K.; Mir, H.; Kapur, N.; Bae, S.; Singh, S. CC chemokines are differentially expressed in Breast Cancer and are associated with disparity in overall survival. Sci. Rep. 2019, 9, 4014. [Google Scholar] [CrossRef] [PubMed]
- Lorena, S.; Oliveira, D.; Dorta, R.; Landman, G.; Kowalski, L. Eotaxin expression in oral squamous cell carcinomas with and without tumour associated tissue eosinophilia. Oral Dis. 2003, 9, 279–283. [Google Scholar] [CrossRef]
- Gerard, C.; Rollins, B.J. Chemokines and disease. Nat. Immunol. 2001, 2, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Li, G.; Asano, T.; Saito, M.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Development of a Novel Anti-Mouse CCR2 Monoclonal Antibody (C2Mab-6) by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 80–86. [Google Scholar] [CrossRef]
- Tanaka, T.; Li, G.; Saito, M.; Suzuki, H.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of an Anti-human CCR2 Monoclonal Antibody (C2Mab-9) by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 188–193. [Google Scholar] [CrossRef]
- Asano, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Tanaka, T.; Sano, M.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 3 Monoclonal Antibodies for Flow Cytometry. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 107–112. [Google Scholar] [CrossRef]
- Asano, T.; Suzuki, H.; Goto, N.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Establishment of Novel Anti-Mouse CCR3 Monoclonal Antibodies (C3Mab-6 and C3Mab-7) by N-terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 94–100. [Google Scholar] [CrossRef]
- Asano, T.; Suzuki, H.; Tanaka, T.; Saito, M.; Li, G.; Goto, N.; Nanamiya, R.; Kaneko, M.K.; Kato, Y. C3Mab-3: A Monoclonal Antibody for Mouse CC Chemokine Receptor 3 for Flow Cytometry. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 74–79. [Google Scholar] [CrossRef]
- Takei, J.; Suzuki, H.; Asano, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-Mouse CCR4 Monoclonal Antibody (C4Mab-1) by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of an Anti-Mouse CCR8 Monoclonal Antibody (C8Mab-1) for Flow Cytometry and Immunocytochemistry. Monoclon. Antibodies Immunodiagn. Immunother. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Saito, M.; Asano, T.; Tanaka, T.; Kitamura, K.; Kudo, Y.; Kaneko, M.K.; Kato, Y. C8Mab-3: An Anti-Mouse CCR8 Monoclonal Antibody for Immunocytochemistry. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Nanamiya, R.; Takei, J.; Nakamura, T.; Yanaka, M.; Hosono, H.; Sano, M.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of Anti-Mouse CC Chemokine Receptor 8 Monoclonal Antibodies for Flow Cytometry. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Nanamiya, R.; Takei, J.; Asano, T.; Tanaka, T.; Sano, M.; Nakamura, T.; Yanaka, M.; Hosono, H.; Kaneko, M.K.; Kato, Y. Development of Anti-Human CC Chemokine Receptor 9 Monoclonal Antibodies for Flow Cytometry. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, K.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Cx6Mab-1: A Novel Anti-Mouse CXCR6 Monoclonal Antibody Established by N-Terminal Peptide Immunization. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Li, G.; Asano, T.; Kaneko, M.K.; Suzuki, H.; Kato, Y. Epitope Mapping of the Anti-Human CCR2 Monoclonal Antibody C2Mab-9. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Identification of the Binding Epitope of an Anti-mouse CCR4 Monoclonal Antibody, C4Mab-1. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 214–220. [Google Scholar] [CrossRef]
- Takei, J.; Asano, T.; Li, G.; Saito, M.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of an Anti-Human CCR9 Monoclonal Antibody (C9Mab-1) Using Enzyme-Linked Immunosorbent Assay. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 239–242. [Google Scholar] [CrossRef]
- Li, G.; Suzuki, H.; Asano, T.; Tanaka, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-EpCAM Monoclonal Antibody for Various Applications. Antibodies 2022, 11, 41. [Google Scholar] [CrossRef]
- Itai, S.; Fujii, Y.; Nakamura, T.; Chang, Y.-W.; Yanaka, M.; Saidoh, N.; Handa, S.; Suzuki, H.; Harada, H.; Yamada, S.; et al. Establishment of CMab-43, a Sensitive and Specific Anti-CD133 Monoclonal Antibody, for Immunohistochemistry. Monoclon. Antibodies Immunodiagn. Immunother. 2017, 36, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Kaneko, M.K.; Ogasawara, S.; Yamada, S.; Yanaka, M.; Nakamura, T.; Saidoh, N.; Yoshida, K.; Honma, R.; Kato, Y. Development of RAP Tag, a Novel Tagging System for Protein Detection and Purification. Monoclon. Antibodies Immunodiagn. Immunother. 2017, 36, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Yoshida, H.; Nosaki, S.; Kaneko, M.K.; Kato, Y. RAP Tag and PMab-2 Antibody: A Tagging System for Detecting and Purifying Proteins in Plant Cells. Front. Plant Sci. 2020, 11, 510444. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Kaneko, M.K.; Kato, Y. MAP Tag: A Novel Tagging System for Protein Purification and Detection. Monoclon. Antibodies Immunodiagn. Immunother. 2016, 35, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Wakasa, A.; Kaneko, M.K.; Kato, Y.; Takagi, J.; Arimori, T. Site-specific epitope insertion into recombinant proteins using the MAP tag system. J. Biochem. 2020, 168, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Kohli, L.L.; Stone, M.J. Characterization of binding between the chemokine eotaxin and peptides derived from the chemokine receptor CCR3. J. Biol. Chem. 2000, 275, 27250–27257. [Google Scholar] [CrossRef] [PubMed]
- Millard, C.J.; Ludeman, J.P.; Canals, M.; Bridgford, J.L.; Hinds, M.G.; Clayton, D.J.; Christopoulos, A.; Payne, R.J.; Stone, M.J. Structural Basis of Receptor Sulfotyrosine Recognition by a CC Chemokine: The N-Terminal Region of CCR3 Bound to CCL11/Eotaxin-1. Structure 2014, 22, 1571–1581. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.-H.; Xu, F.; Zhang, G.-S.; Wang, S.-B.; Xu, W.-H. CCR3 monoclonal antibody inhibits airway eosinophilic inflammation and mucus overproduction in a mouse model of asthma. Acta Pharmacol. Sin. 2006, 27, 1594–1599. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tateyama, N.; Asano, T.; Suzuki, H.; Li, G.; Yoshikawa, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Epitope Mapping of Anti-Mouse CCR3 Monoclonal Antibodies Using Flow Cytometry. Antibodies 2022, 11, 75. https://doi.org/10.3390/antib11040075
Tateyama N, Asano T, Suzuki H, Li G, Yoshikawa T, Tanaka T, Kaneko MK, Kato Y. Epitope Mapping of Anti-Mouse CCR3 Monoclonal Antibodies Using Flow Cytometry. Antibodies. 2022; 11(4):75. https://doi.org/10.3390/antib11040075
Chicago/Turabian StyleTateyama, Nami, Teizo Asano, Hiroyuki Suzuki, Guanjie Li, Takeo Yoshikawa, Tomohiro Tanaka, Mika K. Kaneko, and Yukinari Kato. 2022. "Epitope Mapping of Anti-Mouse CCR3 Monoclonal Antibodies Using Flow Cytometry" Antibodies 11, no. 4: 75. https://doi.org/10.3390/antib11040075
APA StyleTateyama, N., Asano, T., Suzuki, H., Li, G., Yoshikawa, T., Tanaka, T., Kaneko, M. K., & Kato, Y. (2022). Epitope Mapping of Anti-Mouse CCR3 Monoclonal Antibodies Using Flow Cytometry. Antibodies, 11(4), 75. https://doi.org/10.3390/antib11040075