Traditional Pollarding Practices for Dimorphic Ash Tree (Fraxinus dimorpha) Support Soil Fertility in the Moroccan High Atlas

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
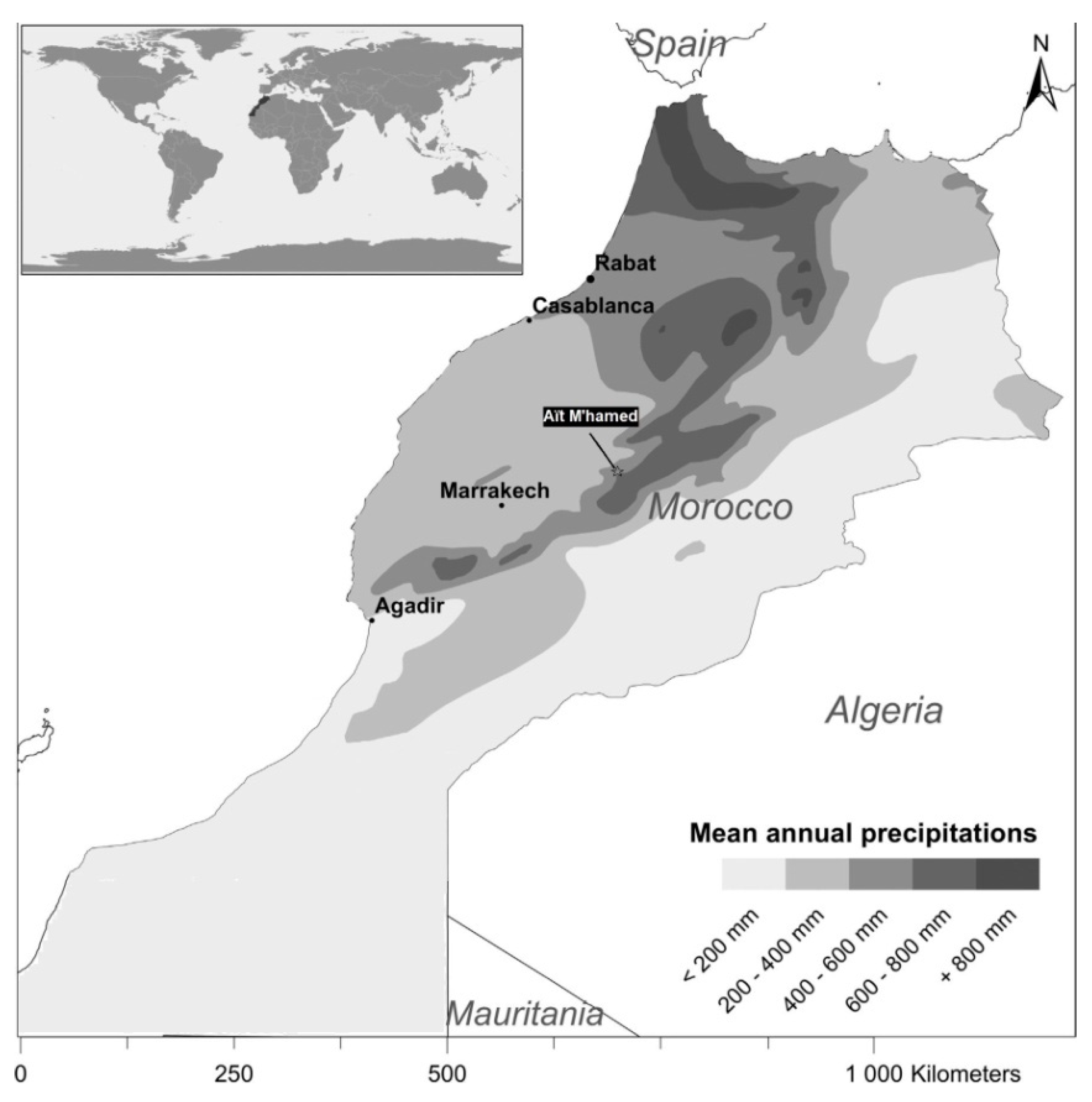
2.1. Study Site and Sampling
2.2. Soil Chemical and Physical Analysis
2.3. Mycorrhizal Attributes
2.3.1. Root Mycorrhization
2.3.2. Spore Enumeration
2.4. Statistical Analysis
3. Results
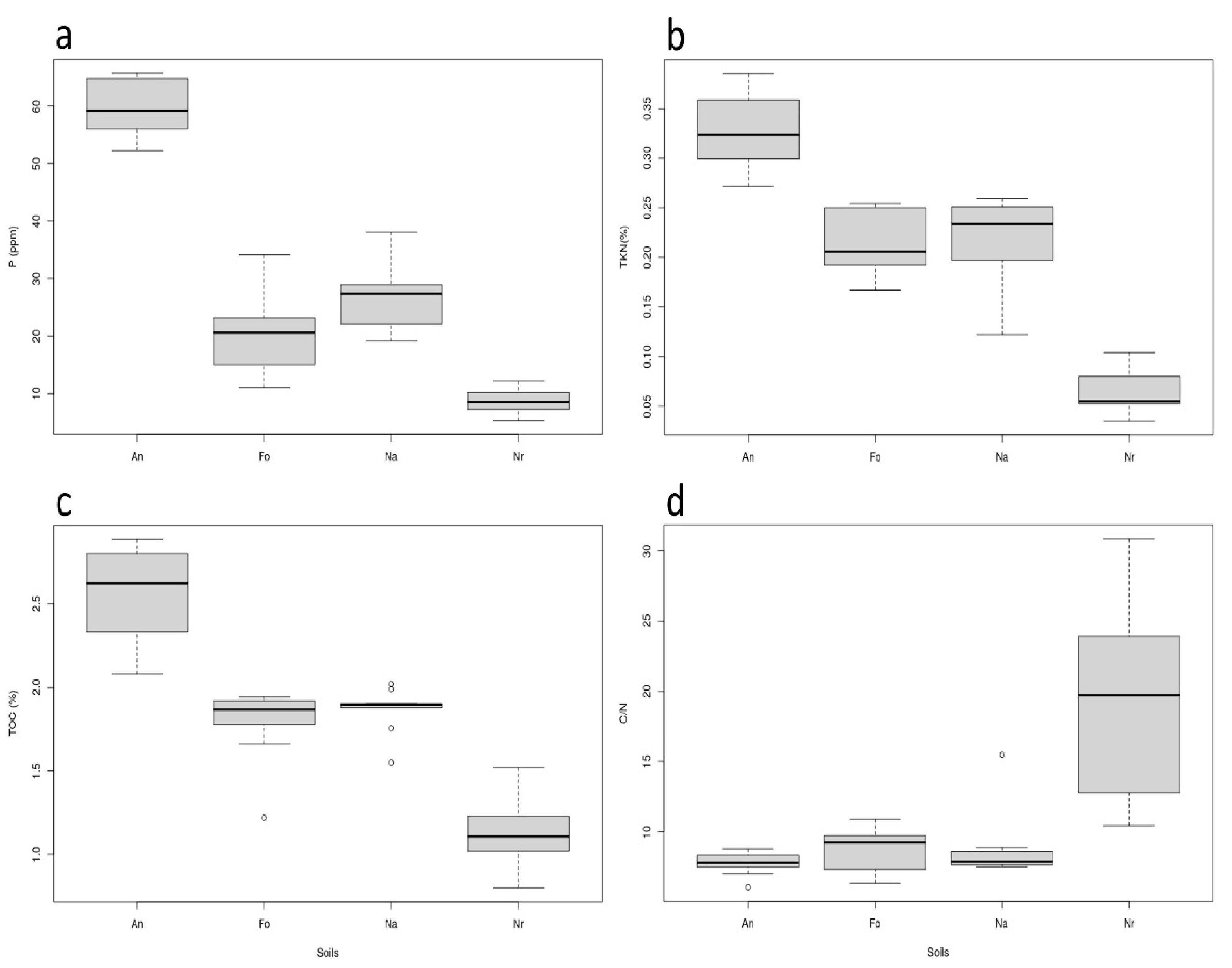
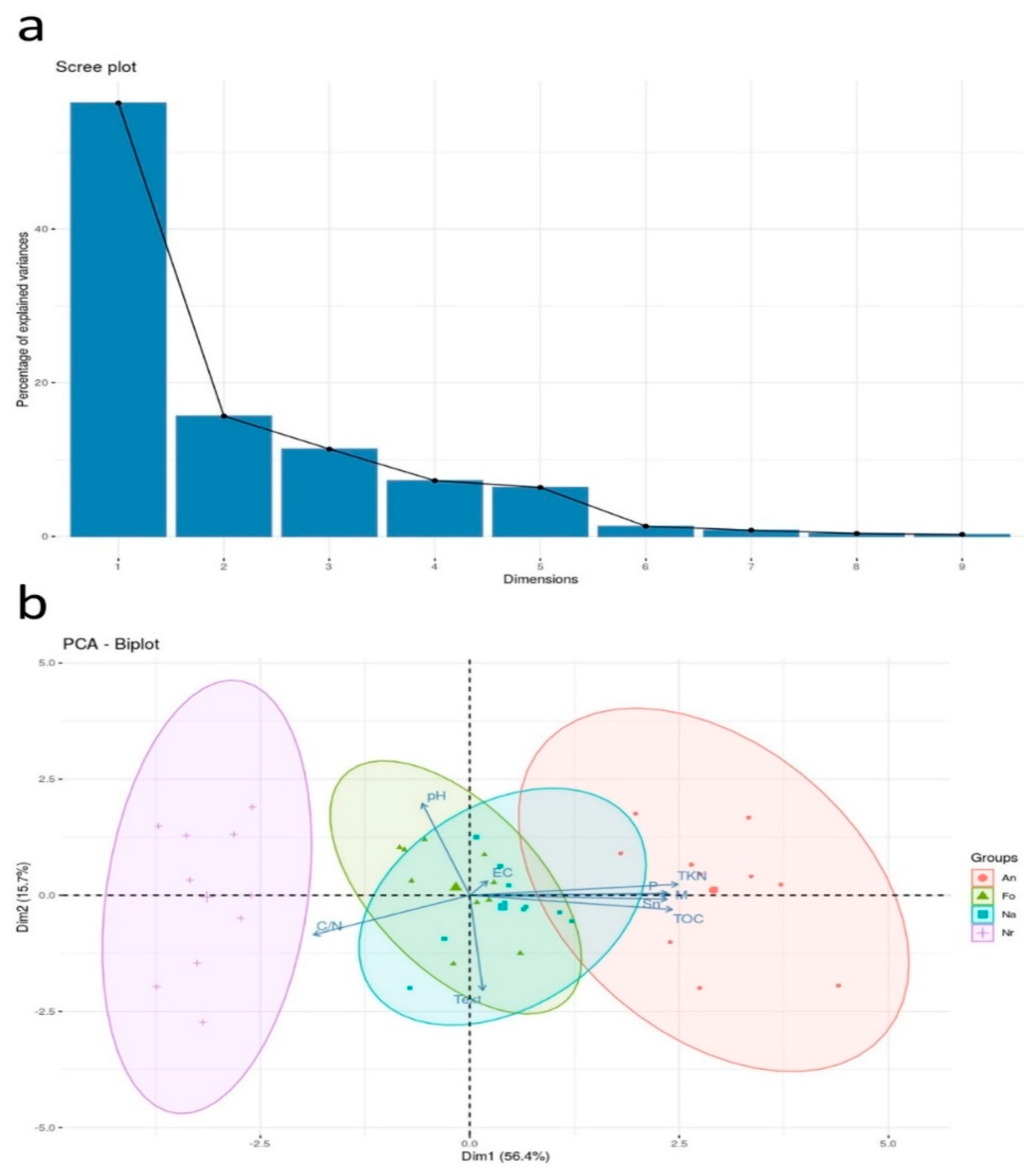
3.1. Soil Chemical and Physical Analyses
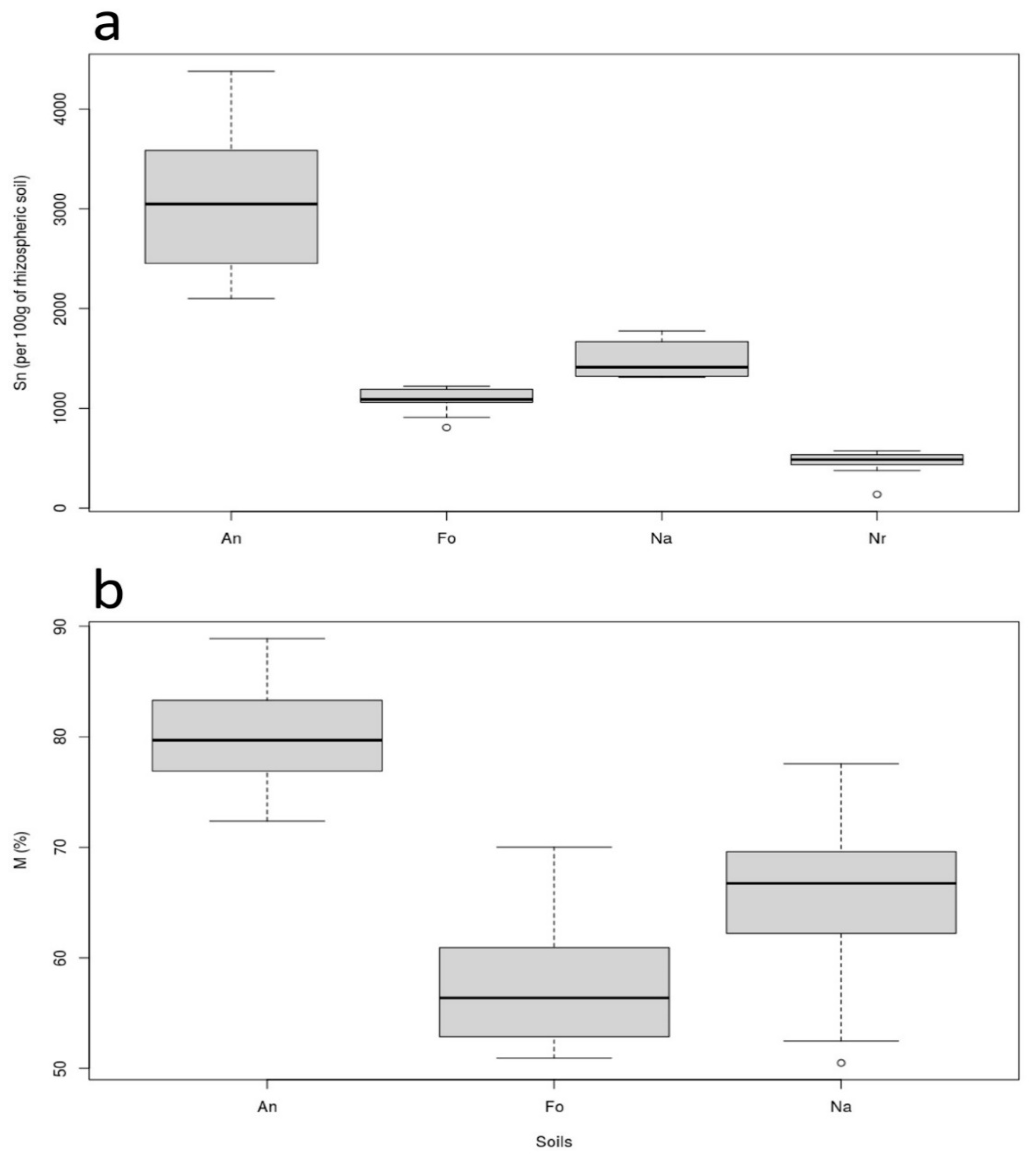
3.2. Soil Mycorrhizal Attributes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Parrotta, J.; Agnoletti, M. Traditional forest knowledge: Challenges and opportunities. Forest Ecol. Manag. 2007, 249, 1–4. [Google Scholar] [CrossRef]
- Maffi, L.; Woodley, E. Biocultural Diversity Conservation: A Global Sourcebook; Routledge: London, UK, 2012. [Google Scholar]
- De Freitas, J.G.; Bastos, M.R.; Dias, J.A. Traditional Ecological Knowledge as a Contribution to Climate Change Mitigation and Adaptation: The Case of the Portuguese Coastal Populations. In Handbook of Climate Change Communication: Case Studies in Climate Change Communication; Filho, W.L., Manolas, E., Azul, A.M., Azeiteiro, U.M., McGhie, H., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 257–269. [Google Scholar] [CrossRef]
- Wyllie de Echeverria, V.R.; Thornton, T.F. Using traditional ecological knowledge to understand and adapt to climate and biodiversity change on the Pacific coast of North America. Ambio 2019, 48, 1447–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrotta, J.; Yeo-Chang, Y.; Camacho, L.D. Traditional knowledge for sustainable forest management and provision of ecosystem services. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2016, 12, 1–4. [Google Scholar] [CrossRef]
- Genin, D.; M’Sou, S.; Ferradous, A.; Alifriqui, M. Another vision of sound tree and forest management: Insights from traditional ash shaping in the Moroccan Berber mountains. For. Ecol. Manag. 2018, 429, 180–188. [Google Scholar] [CrossRef]
- Genin, D.; Crochot, C.; MSou, S.; Araba, A.; Alifriqui, M. Meadow up a tree: Feeding flocks with a native ash tree in the Moroccan mountains. Pastoralism 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Genin, D.; Alifriqui, M. The agroforestry parklands with dimorphic ash trees of Aït M’Hamed. A well-guarded local specificity. In The Emergence of Local Specificities in the Mediterranean Hinterlands; Aderghal, M., Genin, D., Hanafi, A., Landel, P.A., Michon, G., Eds.; Laboratoire Population-Environnement-Développement: Marseille, France, 2019; pp. 30–51. ISBN 979-10-96763-08-5. [Google Scholar]
- Schweingruber, F.H. Wood Structure and Environment; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Howard, K.S.C.; Eldridge, D.J.; Soliveres, S. Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland. Basic Appl. Ecol. 2012, 13, 159–168. [Google Scholar] [CrossRef]
- Birhane, E.; Ahmed, S.; Hailemariam, M.; Negash, M.; Rannestad, M.M.; Norgrove, L. Carbon stock and woody species diversity in homegarden agroforestry along an elevation gradient in southern Ethiopia. Agrofor. Syst. 2020, 94, 1099–1110. [Google Scholar] [CrossRef]
- Rillig, M.C.; Mummey, D.L. Mycorrhizas and soil structure. New Phytol. 2006, 171, 41–53. [Google Scholar] [CrossRef]
- Schinner, F.; Öhlinger, R.; Kandeler, E.; Margesin, R. Methods in Soil Biology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Olsen, S.R. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. USDA Circular 1954, 24. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Measurement of the mycorrhization rate VA of a root system. In search for an estimation method with functional significance Presented at the Physiological and genetical aspects of mycorrhizae. In Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France, 1–5 July 1985; pp. 217–221. [Google Scholar]
- Gerdemann, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Scheffe, H. The Analysis of Variance; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1959; pp. 351–358. [Google Scholar]
- Jolliffe, I.T. Principal Component Analysis. In Series in Statistics; Springer: New York, NY, USA, 2012. [Google Scholar] [CrossRef]
- Singh, R.K.; Singh, A.; Garnett, S.T.; Zander, D.K.; Lobsang, D.K.; Tsering, D. Paisang (Quercus griffithii); a keystone tree species in sustainable agroecosystem management and livelihoods in Arunachal Pradesh, India. Environ. Manag. 2015, 55, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.; Mize, C.W.; McNabb, H.S. Populations of endogonaceous fungi at two locations in central Iowa. Can. J. Bot. 1982, 60, 2518–2529. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Q.; Li, Y.; Di, H.; Xu, J.; Li, J.; Pan, H. Effects of pasture management on soil fertility and microbial communities in the semi-arid grasslands of Inner Mongolia. J. Soils Sediments 2016, 16, 235–242. [Google Scholar] [CrossRef]
- Guangyu, Z.; Zhuangsheng, T.; Lei, C.; Zhouping, S.; Lei, D. Overgrazing depresses soil carbon stock through changing plant diversity in temperate grassland of the Loess Plateau. Plant Soil Environ. 2018, 64, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Yan, Y.; Xu, D.; Xu, X.; Wang, C.; Wang, X.; Eldridge, D.J. The fertile island effect collapses under extreme overgrazing: Evidence from a shrub-encroached grassland. Plant Soil 2020, 448, 201–212. [Google Scholar] [CrossRef]
- Faghihinia, M.; Zou, Y.; Chen, Z.; Bai, Y.; Li, W.; Marrs, R.; Staddon, P.L. The response of grassland mycorrhizal fungal abundance to a range of long-term grazing intensities. Rhizosphere 2020, 13, 100178. [Google Scholar] [CrossRef]
- Yang, X.; Chen, J.; Shen, Y.; Dong, F.; Chen, J. Global negative effects of livestock grazing on arbuscular mycorrhizas: A meta-analysis. Sci. Total Environ. 2020, 708, 134553. [Google Scholar] [CrossRef]
- Li, Y.; Wu, J.; Shen, J.; Liu, S.; Wang, C.; Chen, D.; Zhang, J. Soil microbial C:N ratio is a robust indicator of soil productivity for paddy fields. Sci. Rep. 2016, 6, 35266. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Dong, S.; Li, S.; Xiao, J.; Han, Y.; Yang, M.; Yeomans, J.C. Grazing enhances plant photosynthetic capacity by altering soil nitrogen in alpine grasslands on the Qinghai-Tibetan plateau. Agric. Ecosyst. Environ. 2019, 280, 161–168. [Google Scholar] [CrossRef]
- Wan, X.; Huang, Z.; He, Z.; Yu, Z.; Wang, M.; Davis, M.R.; Yang, Y. Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations. Plant Soil 2015, 387, 103–116. [Google Scholar] [CrossRef]
- Eom, A.-H.; Wilson, G.W.T.; Hartnett, D.C. Effects of ungulate grazers on arbuscular mycorrhizal symbiosis and fungal community structure in tallgrass prairie. Mycologia 2001, 93, 233–242. [Google Scholar] [CrossRef]
- Li, W.; Cao, W.; Wang, J.; Li, X.; Xu, C.; Shi, S. Effects of grazing regime on vegetation structure, productivity, soil quality, carbon and nitrogen storage of alpine meadow on the Qinghai-Tibetan Plateau. Ecol. Eng. 2017, 98, 123–133. [Google Scholar] [CrossRef]
- Brust, G.E. Chapter 9-Management Strategies for Organic Vegetable Fertility. In Safety and Practice for Organic Food; Biswas, D., Micallef, S.A., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 193–212. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, Q.; Noll, L.; Hu, Y.; Wanek, W. Drivers of microbial nitrogen use efficiency and soil inorganic N processes at the continental scale. Geophys. Res. Abstr. 2019, 21, 1–4. [Google Scholar]
- Khakpour, O.; Khara, J. Spore density and root colonization by arbuscular mycorrhizal fungi in some species in the northwest of Iran. Int. Res. J. Appl. Basic Sci. 2012, 3, 977–982. [Google Scholar]
- Tamil Pradha, K.; Sivakumar, K. Effects of am fungi on spores density and root colonization of chilli (Capsicum annum) at different levels of rock phosphate. Int. J. Curr. Res. Life Sci. 2018, 7, 1415–1419. [Google Scholar]
- Fakhech, A.; Ouahmane, L.; Hafidi, M. Seasonality of mycorrhizal attributes, soil phosphorus and nitrogen of Juniperus phoenicea and Retama monosperma boiss. in an Atlantic sand dunes forest. J. Sustain. For. 2019, 38, 1–17. [Google Scholar] [CrossRef]
- Gao, D.; Pan, X.; Zhou, X.; Wei, Z.; Li, N.; Wu, F. Phosphorus fertilization and intercropping interactively affect tomato and potato onion growth and rhizosphere arbuscular mycorrhizal fungal community. Arch. Agron. Soil Sci. 2020. [Google Scholar] [CrossRef]
- Rubio, R.; Borie, F.; Schalchli, C.; Castillo, C.; Azcón, R. Occurrence and effect of arbuscular mycorrhizal propagules in wheat as affected by the source and amount of phosphorus fertilizer and fungal inoculation. Appl. Soil Ecol. 2003, 23, 245–255. [Google Scholar] [CrossRef]
- Cuéllar, A.E.; Martinez, L.R.; Espinosa, R.R.; Cuéllar, E.E. The Inoculation with an Effecient AMF Strain Decreases the Phosphoric Fertilizer Requirements in Ipomea Batata (L.), Lam in Dry Period. J. Chem. Environ. Biol. Eng. 2019, 3, 13. [Google Scholar] [CrossRef]
- Effendy, M.; Wijayani, B.W. Estimation of Available Phosphorus in Soil Using the Population of Arbuscular Mycorrhizal Fungi Spores. J. Trop. Soils 2019, 16, 225–232. [Google Scholar] [CrossRef]
- Abdel-Fattah, G.M.; El-Dohlob, A.F.; El-Dohlob, G.M.; El-Haddad, S.M.; Hafez, S.A.; Rashad, Y.M. An ecological view of arbuscular mycorrhizal status in some Egyptian plants. J. Environ. Sci. 2010, 37, 123–136. [Google Scholar]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A. Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth: New Paradigms from Cellular to Ecosystem Scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, J.R.; Rillig, M.C. Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol. 2018, 220, 1059–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fakhech, A.; Ouahmane, L.; Hafidi, M. Analysis of symbiotic microbial status of Atlantic sand dunes forest and its effects on Acacia gummifera and Retama monosperma (Fabaceae) to be used in reforestation. J. For. Res. 2019. [Google Scholar] [CrossRef]
- Ait-El-Mokhtar, M.; Fakhech, A.; Anli, M.; Ben-Laouane, R.; Boutasknit, A.; Wahbi, S.; Meddich, A. Infectivity of the palm groves arbuscular mycorrhizal fungi under arid and semi-arid climate and its edaphic determinants towards efficient ecological restoration. Rhizosphere 2020, 100220. [Google Scholar] [CrossRef]
- Tucker, M. Effects of Phosphorus on Mycorrhizal Ccolonization of Wetland Plants under Natural and Controlled Conditions. Master’s Thesis, Wilfrid Laurier University, Waterloo, Belgium, 2020. [Google Scholar]
- Linderman, R.G. The mycorrhizosphere phenomenon. In Proceedings of the International Symposium “Mycorrhiza for Plant Vitality” and the Joint Meeting of Working Groups 1–4 of COST Action 870, Hannover, Germany, 3–5 October 2007; pp. 341–355. [Google Scholar]
- Nuccio, E.E.; Hodge, A.; Pett-Ridge, J.; Herman, D.J.; Weber, P.K.; Firestone, M.K. An arbuscular mycorrhizal fungus significantly modifies the soil bacterial community and nitrogen cycling during litter decomposition: AMF alters soil bacterial community and N cycling. Environ. Microbiol. 2013, 15, 1870–1881. [Google Scholar] [CrossRef]
- Lin, C.; Wang, Y.; Liu, M.; Li, Q.; Xiao, W.; Song, X. Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia lanceolata). Sci Rep. 2020, 10, 12260. [Google Scholar] [CrossRef]
- Liu, C.; Ravnskov, S.; Liu, F.; Rubæk, G.H.; Andersen, M.N. Arbuscular mycorrhizal fungi alleviate abiotic stresses in potato plants caused by low phosphorus and deficit irrigation/partial root-zone drying. J. Agric. Sci. 2018, 156, 46–58. [Google Scholar] [CrossRef]
- Camenzind, T.; Homeier, J.; Dietrich, K.; Hempel, S.; Hertel, D.; Krohn, A.; Rillig, M.C. Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradient in tropical montane forests. Soil Biol. Biochem. 2016, 94, 37–47. [Google Scholar] [CrossRef]
- Xiao, D.; Che, R.; Liu, X.; Tan, Y.; Yang, R.; Zhang, W.; Wang, K. Arbuscular mycorrhizal fungi abundance was sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems. Biol. Fertil. Soils 2019, 55, 457–469. [Google Scholar] [CrossRef]
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Petit, S.; Watkins, C. Forgotten peasant practices: Tree pollarding in Great Britain. Etudes Rural. 2004, 169, 197–214. [Google Scholar] [CrossRef]
- Genin, D.; Aumeerudy-Thomas, Y.; Balent, G.; Nasi, R. The multiple dimensionsof rural forests: Lessons from a comparative analysis. Ecol. Soc. 2013, 18, 27. [Google Scholar] [CrossRef] [Green Version]
- Ferrini, F. Pollarding and its effects on tree physiology: A look to mature and senescent tree management in Italy. In Pollard Trees in Europe, European Meeting on Pollard Trees; Dumont, E., Ed.; Maison Botanique: Boursay, France, 2006; pp. 71–79. [Google Scholar]
- Van Noordwijk, M.; Purnomosidhi, P. Root architecture in relation to tree-soil-crop interactions and shoot pruning in agroforestry. Agrofor. Syst. 1995, 30, 161–173. [Google Scholar] [CrossRef]
- Connell, J. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Siebert, S.F.; Belsky, J.M. Historic livelihoods and land uses as ecological disturbances and their role in enhancing biodiversity: An example from Bhutan. Biol. Conserv. 2014, 177, 82–89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ANOVA p-Values | Tukey’s HSD p-Values | |||||||
|---|---|---|---|---|---|---|---|---|
| F-Value | Pr (>F) | An–Na | An–Fo | An–Nr | Na–Fo | Na–Nr | Fo–Nr | |
| M | 321.9 | <2 × 10−16 *** | 3.13 × 10−5 | 0 | 0 | 0.06 | 0 | 0 |
| Sn | 92.38 | <2 × 10−16 *** | 0 | 0 | 0 | 0.09 | 2.4 × 10−6 | 0.003 |
| P | 142.4 | <2 × 10−16 *** | 0 | 0 | 0 | 0.07 | 1 × 10−7 | 0.0001 |
| TKN | 95.32 | <2 × 10−16 *** | 4 × 10−7 | 0 | 0 | 0.86 | 0 | 0 |
| TOC | 68.31 | 6.24 × 10−15 *** | 4 × 10−7 | 0 | 0 | 0.86 | 0 | 5 × 10−7 |
| C/N | 20.36 | 6.9 × 10−8 *** | 0.9 | 0.9 | 4 × 10−7 | 0.9 | 2.6 × 10−6 | 2.1 × 10−6 |
| pH | 0.82 | 0.49 | - | - | - | - | - | - |
| EC | 0.65 | 0.59 | - | - | - | - | - | - |
| M | Sn | P | TKN | TOC | C/N | pH | EC | |
|---|---|---|---|---|---|---|---|---|
| M | 0.01 | 0.01 | 0.07 | 0.05 | 0.99 | 0.3 | 0.19 | |
| Sn | 0.76 | 0.01 | 0.02 | 0 | 0.72 | 0.59 | 0.28 | |
| P | 0.77 | 0.78 | 0.04 | 0.09 | 0.63 | 0.71 | 0.34 | |
| TKN | 0.59 | 0.73 | 0.66 | 0.06 | 0.14 | 0.53 | 0.35 | |
| TOC | 0.63 | 0.91 | 0.56 | 0.61 | 0.28 | 0.44 | 0.47 | |
| C/N | 0.01 | 0.13 | −0.17 | −0.5 | 0.38 | 0.07 | 0.8 | |
| pH | −0.36 | −0.2 | 0.14 | 0.22 | −0.28 | −0.59 | 0.91 | |
| EC | 0.45 | 0.38 | 0.34 | 0.33 | 0.26 | −0.09 | 0.04 |
| M | Sn | P | TKN | TOC | C/N | pH | EC | |
|---|---|---|---|---|---|---|---|---|
| M | 0.13 | 0 | 0.01 | 0.04 | 0.07 | 0.49 | 0.25 | |
| Sn | 0.51 | 0.22 | 0.26 | 0.18 | 0.44 | 0.28 | 0.26 | |
| P | 0.94 | 0.43 | 0.04 | 0.07 | 0.19 | 0.63 | 0.4 | |
| TKN | 0.79 | 0.39 | 0.66 | 0.25 | 0 | 0.44 | 0.18 | |
| TOC | 0.66 | 0.46 | 0.6 | 0.4 | 0.95 | 0.66 | 0.29 | |
| C/N | −0.59 | −0.28 | −0.46 | −0.9 | 0.02 | 0.28 | 0.4 | |
| pH | 0.25 | 0.38 | 0.17 | 0.28 | −0.16 | −0.38 | 0.95 | |
| EC | 0.4 | −0.39 | 0.3 | 0.46 | 0.37 | −0.3 | −0.02 |
| M | Sn | P | TKN | TOC | C/N | pH | EC | |
|---|---|---|---|---|---|---|---|---|
| M | 0 | 0.01 | 0.53 | 0.13 | 0.69 | 0.53 | 0.89 | |
| Sn | 0.82 | 0.29 | 0.62 | 0.2 | 0.61 | 0.58 | 0.99 | |
| P | 0.77 | 0.37 | 0.2 | 0.23 | 0.65 | 0.15 | 0.73 | |
| TKN | 0.23 | 0.18 | 0.44 | 0.4 | 0.03 | 0.84 | 0.52 | |
| TOC | 0.51 | 0.44 | 0.42 | 0.3 | 0.17 | 0.24 | 0.09 | |
| C/N | 0.14 | 0.18 | −0.16 | −0.69 | 0.47 | 0.61 | 0.04 | |
| pH | −0.22 | 0.2 | −0.49 | −0.08 | −0.41 | −0.18 | 0.86 | |
| EC | 0.05 | 0 | 0.13 | 0.23 | −0.57 | −0.66 | 0.06 |
| Sn | P | TKN | TOC | C/N | pH | EC | |
|---|---|---|---|---|---|---|---|
| Sn | 0.1 | 0.54 | 0.42 | 0.85 | 0.52 | 0.81 | |
| P | 0.55 | 0.66 | 0.29 | 0.86 | 0.26 | 0.25 | |
| TKN | 0.22 | −0.16 | 0.92 | 0 | 0.92 | 0.23 | |
| TOC | 0.29 | −0.37 | −0.04 | 0.14 | 0.33 | 0.41 | |
| C/N | 0.07 | 0.06 | −0.85 | 0.5 | 0.54 | 0.58 | |
| pH | −0.23 | 0.4 | 0.04 | −0.35 | −0.22 | 0.41 | |
| EC | −0.09 | −0.4 | 0.41 | 0.3 | −0.2 | 0.29 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fakhech, A.; Genin, D.; Ait-El-Mokhtar, M.; Outamamat, E.M.; M’Sou, S.; Alifriqui, M.; Meddich, A.; Hafidi, M. Traditional Pollarding Practices for Dimorphic Ash Tree (Fraxinus dimorpha) Support Soil Fertility in the Moroccan High Atlas. Land 2020, 9, 334. https://doi.org/10.3390/land9090334
Fakhech A, Genin D, Ait-El-Mokhtar M, Outamamat EM, M’Sou S, Alifriqui M, Meddich A, Hafidi M. Traditional Pollarding Practices for Dimorphic Ash Tree (Fraxinus dimorpha) Support Soil Fertility in the Moroccan High Atlas. Land. 2020; 9(9):334. https://doi.org/10.3390/land9090334
Chicago/Turabian StyleFakhech, Abdessamad, Didier Genin, Mohamed Ait-El-Mokhtar, El Mustapha Outamamat, Soufiane M’Sou, Mohamed Alifriqui, Abdelilah Meddich, and Mohamed Hafidi. 2020. "Traditional Pollarding Practices for Dimorphic Ash Tree (Fraxinus dimorpha) Support Soil Fertility in the Moroccan High Atlas" Land 9, no. 9: 334. https://doi.org/10.3390/land9090334