Bentonite-Based Organic Amendment Enriches Microbial Activity in Agricultural Soils


 ,
,  , ,
, ,  , ,
, ,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
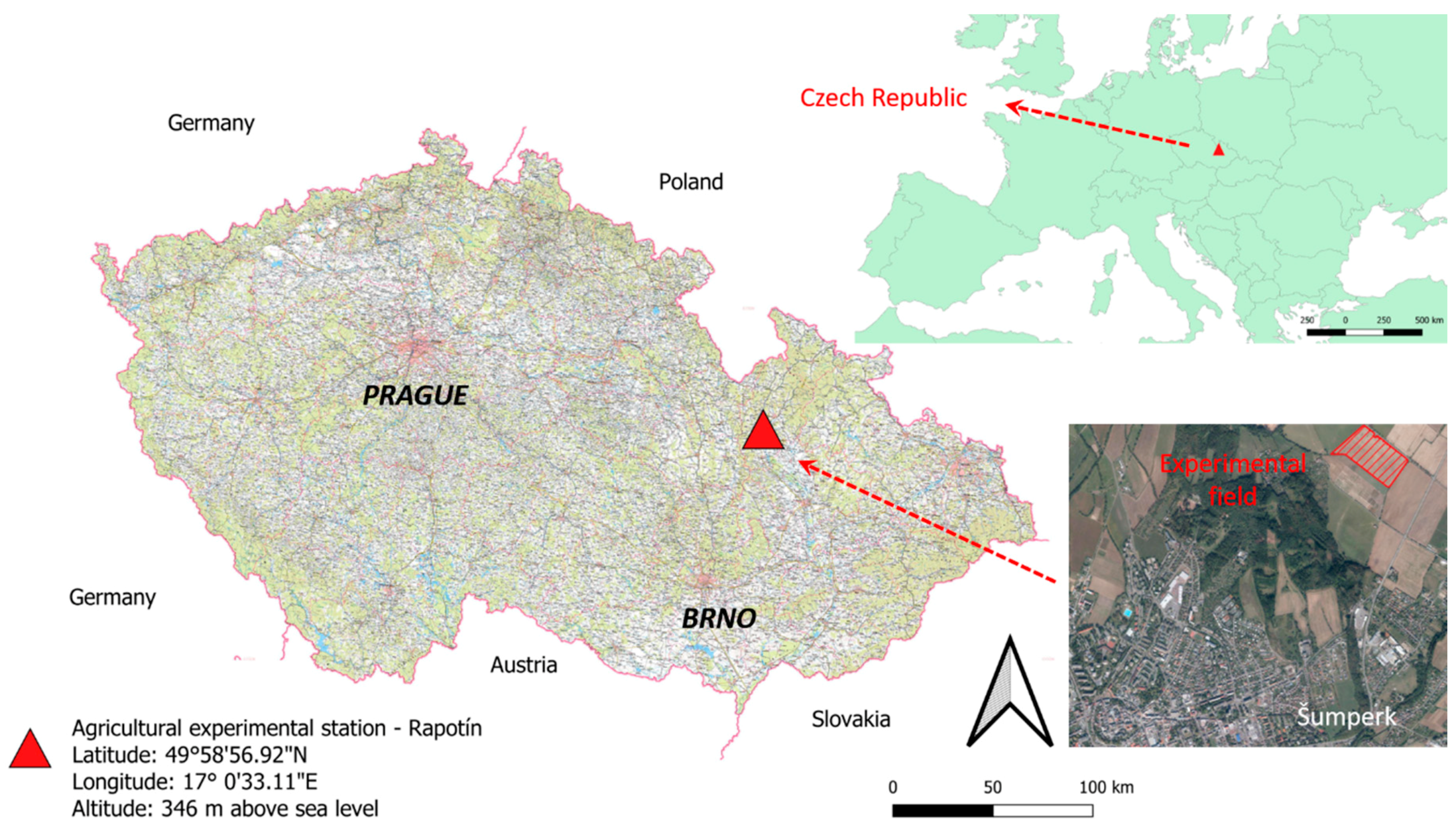
2.1. Experimental Field Site and Experiment Design
2.2. Soil and Amendments Characteristics
2.2.1. Soil Properties
2.2.2. Bentonite Properties
2.2.3. Manure Production and Properties
2.2.4. Combined Manure and Bentonite Production and Properties
2.3. Sample Collection
2.4. Microbial Biomass Carbon
2.5. Dehydrogenase Activities
2.6. DNA Extraction and Real-Time qPCR
2.7. Quantification of Microbial Biomass
2.8. Statistical Analyses
3. Results
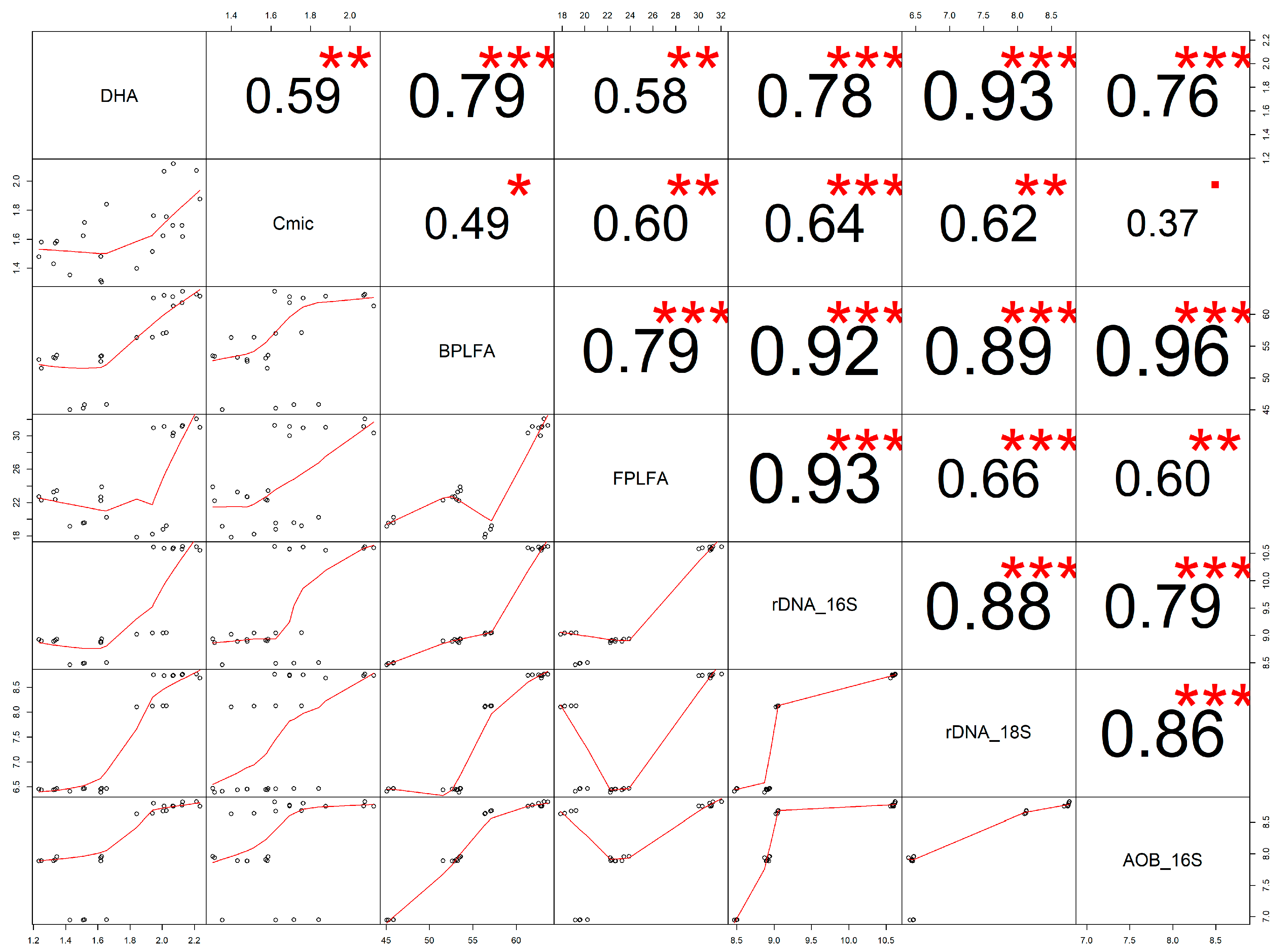
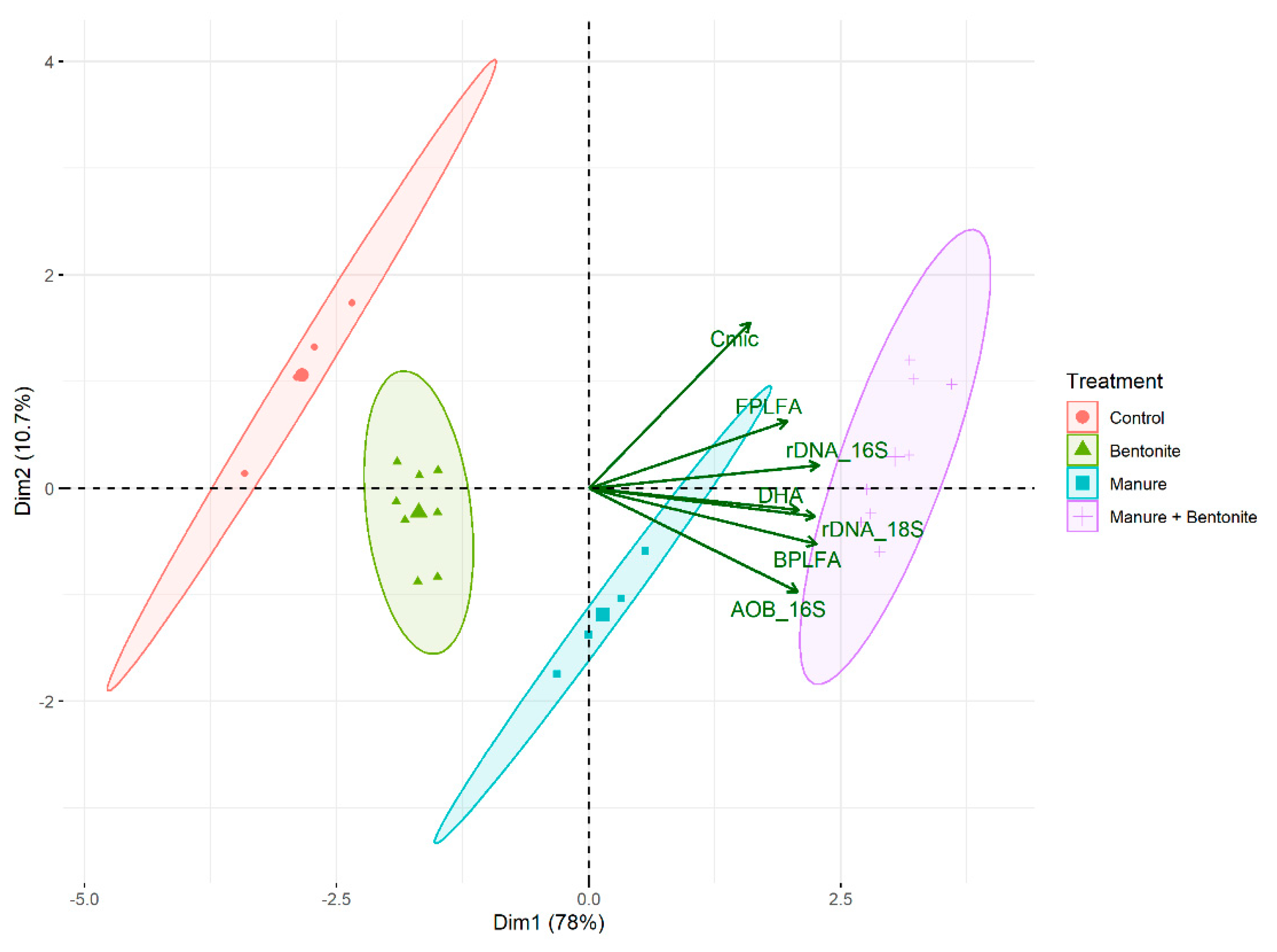
3.1. Dependences of Microbial Indexes on the Treatment Applied
3.2. Dehydrogenase
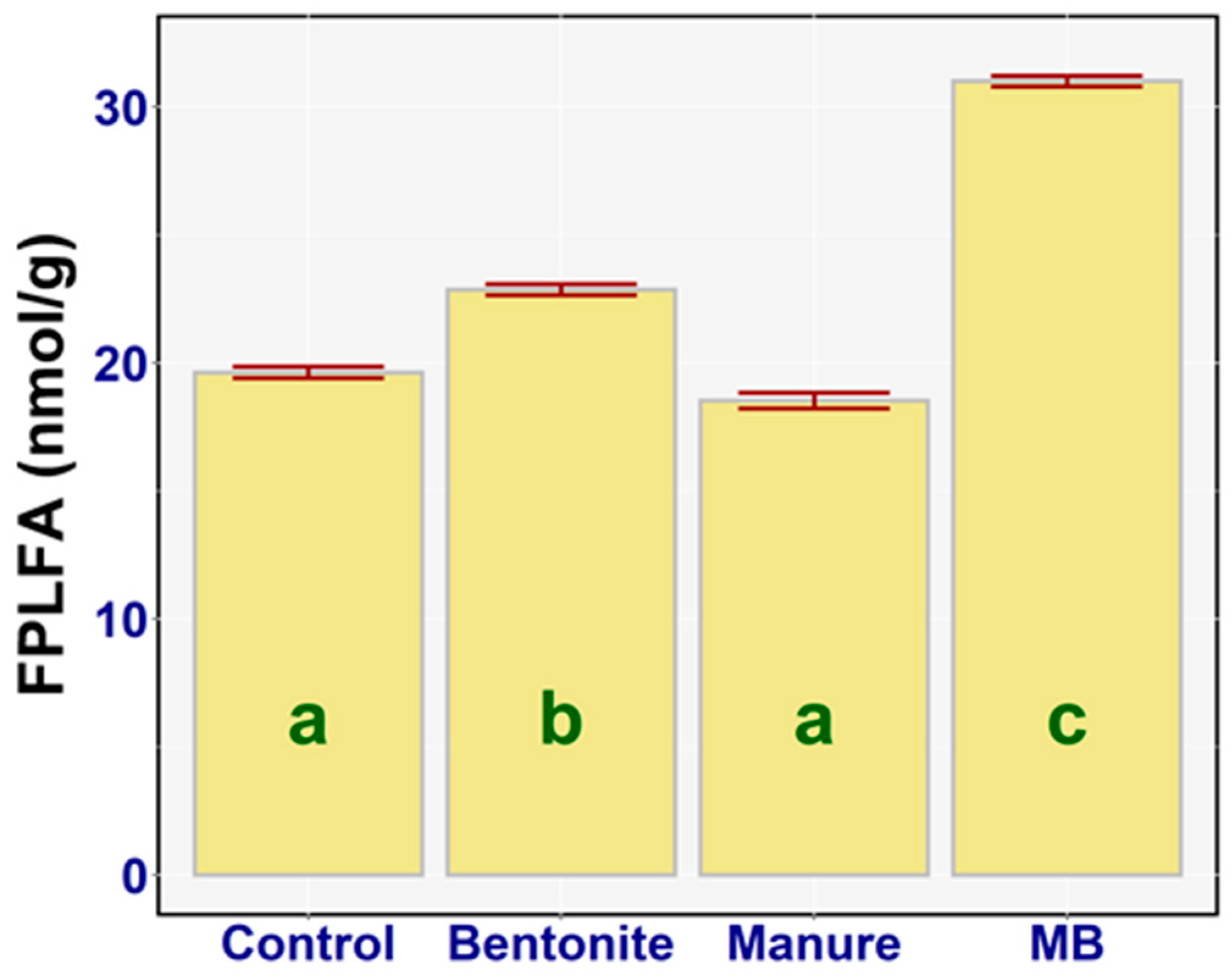
3.3. Soil Phospholipid Fatty Acid Analysis
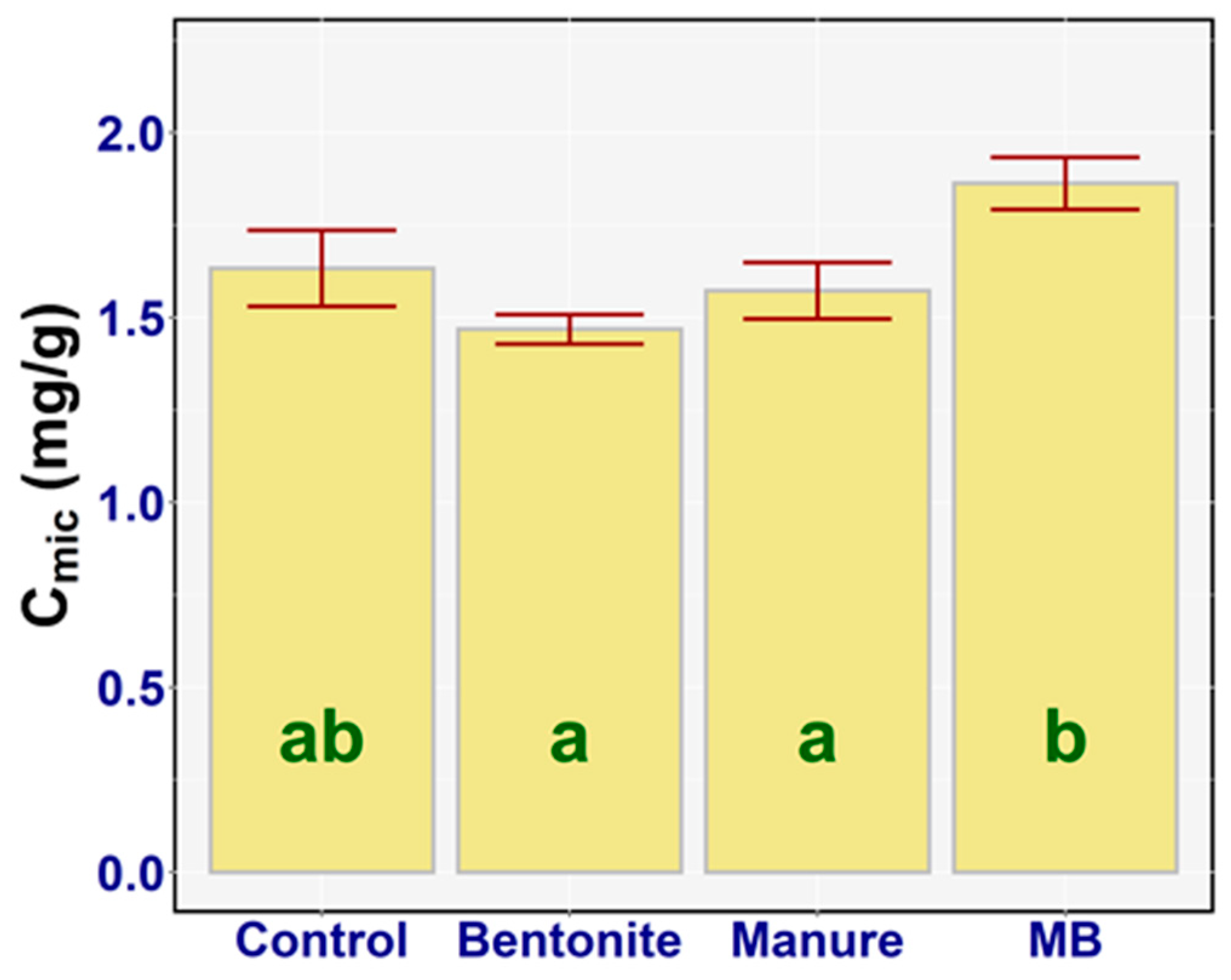
3.4. Microbial Biomass Carbon
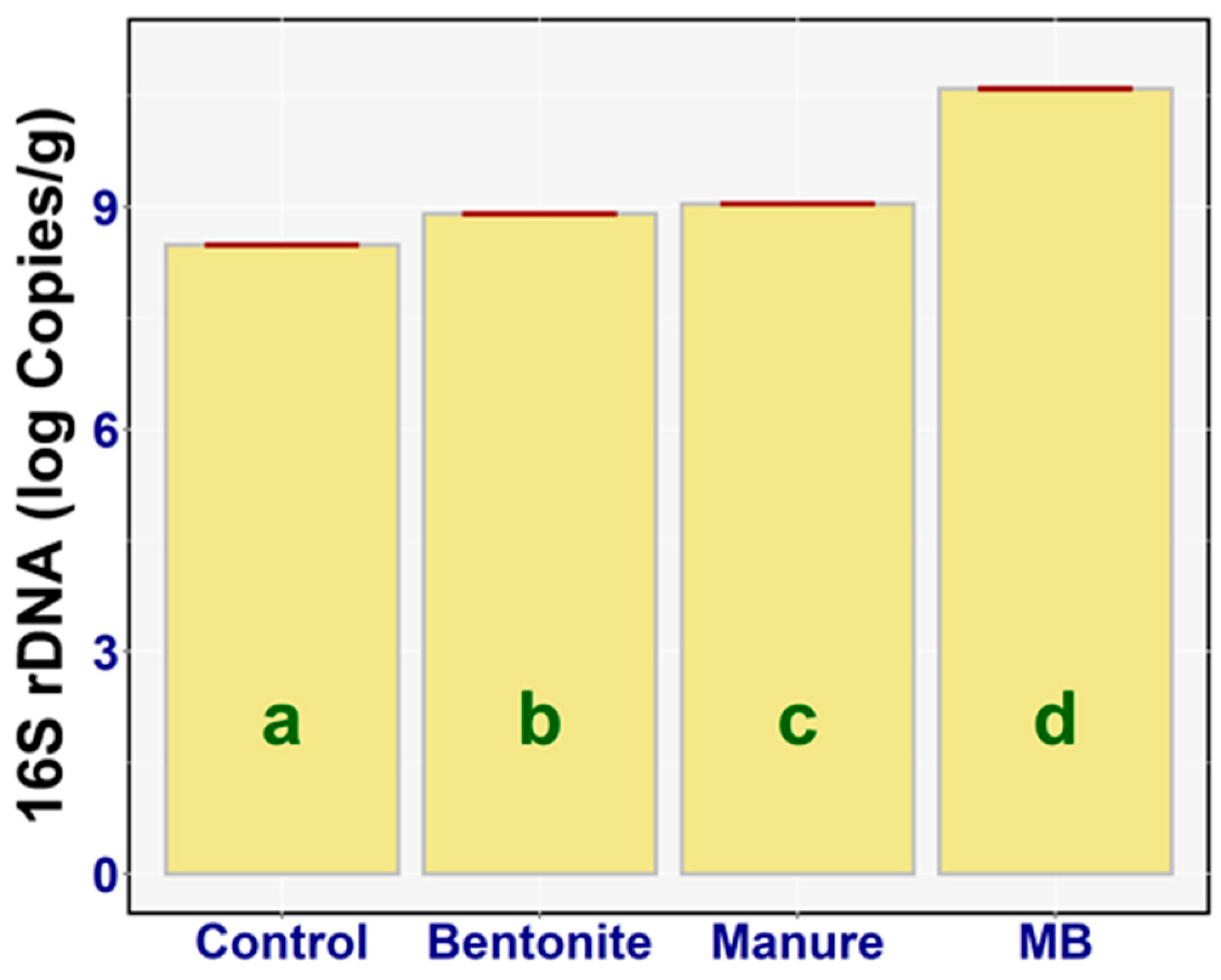
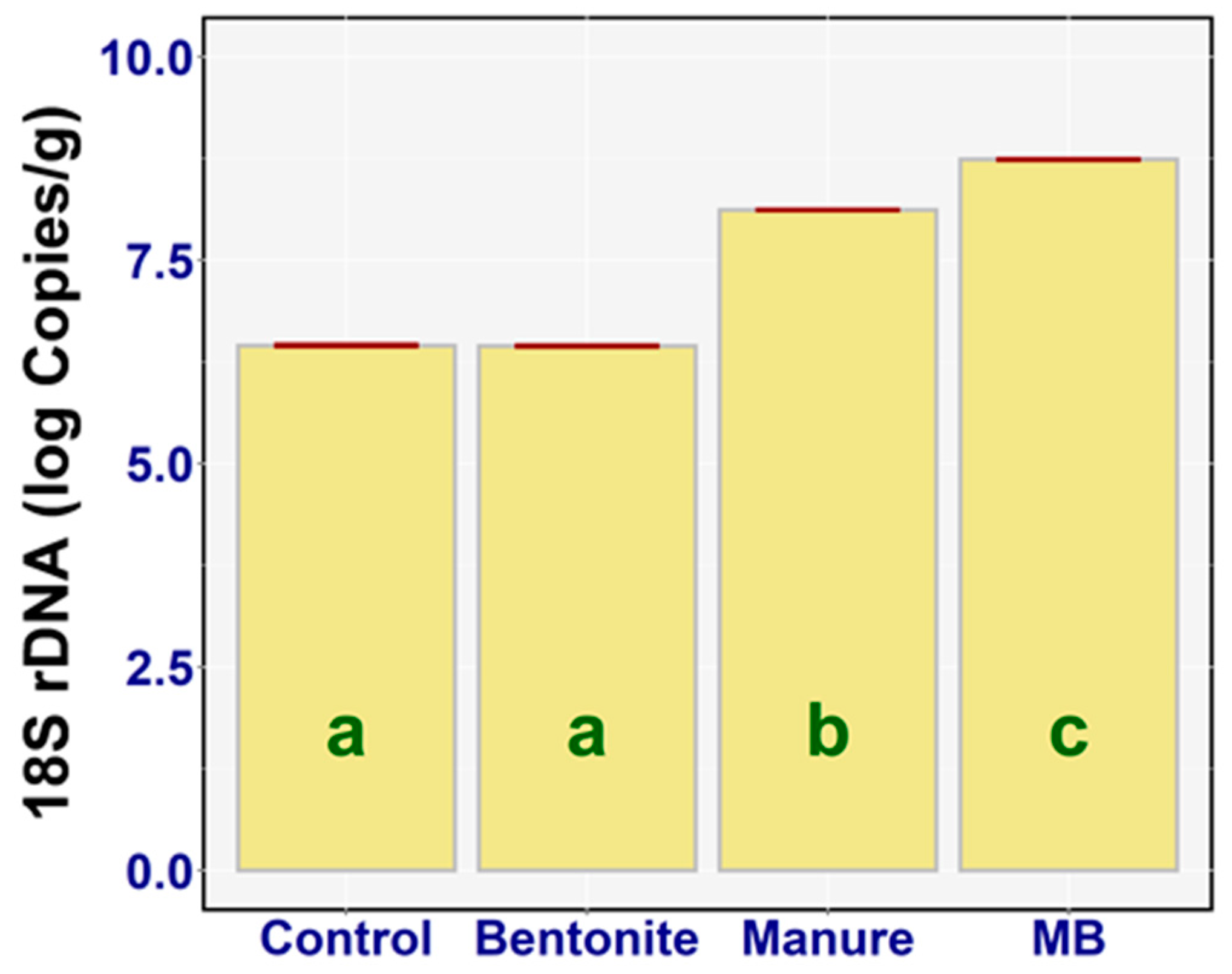
3.5. DNA Extraction and Real-Time qPCR
3.5.1. 16S rDNA
3.5.2. 18S rDNA
3.5.3. Ammonium Oxidation in Soil (16S AOB-rDNA)
4. Discussion
4.1. Dehydrogenase
4.2. Soil Phospholipid Fatty Acid Analysis
4.3. Microbial Biomass Carbon
4.4. DNA Extraction and Real-time qPCR
4.4.1. 16S rDNA
4.4.2. 18S rDNA
4.4.3. 16S AOB-rDNA
4.5. Potential for the Reclamation of Polluted Soils
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foley, J.A. Global Consequences of Land Use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef]
- Hernández, T.; Chocano, C.; Moreno, J.-L.; García, C. Use of compost as an alternative to conventional inorganic fertilizers in intensive lettuce (Lactuca sativa L.) crops—Effects on soil and plant. Soil Tillage Res. 2016, 160, 14–22. [Google Scholar] [CrossRef]
- Davies, J.E.D.; Jabeen, N. The adsorption of herbicides and pesticides on clay minerals and soils. Part 1. Isoproturon. J. Incl. Phenom. Macrocycl. Chem. 2002, 43, 329–336. [Google Scholar] [CrossRef]
- Genç, N.; Dogan, E.C. Adsorption kinetics of the antibiotic ciprofloxacin on bentonite, activated carbon, zeolite, and pumice. Desalin. Water Treat. 2013, 53, 785–793. [Google Scholar] [CrossRef]
- Baker, L.R.; White, P.M.; Pierzynski, G.M. Changes in microbial properties after manure, lime, and bentonite application to a heavy metal-contaminated mine waste. Appl. Soil Ecol. 2011, 48, 1–10. [Google Scholar] [CrossRef]
- Li, J.; Sun, X.; Li, S. Effects of Garden Waste Compost and Bentonite on Muddy Coastal Saline Soil. Sustainability 2020, 12, 3602. [Google Scholar] [CrossRef]
- Myasnikov, S.K.; Tikhonov, A.Y.; Chipryakova, A.P.; Kulov, N.N. Removal of heavy metal ions from water by an combined sorption–crystallization process using activated clays. Theor. Found. Chem. Eng. 2016, 50, 366–382. [Google Scholar] [CrossRef]
- Bouabid, R.; Badraoui, M.; Bloom, P.R. Potassium Fixation and Charge Characteristics of Soil Clays. Soil Sci. Soc. Am. J. 1991, 55, 1493–1498. [Google Scholar] [CrossRef]
- Sequi, P. Le funzioni agronomiche della sostanza organica. In Chimica del suolo; Patron editore: Bologna, Italy, 1989; pp. 279–292. [Google Scholar]
- Paustian, K.; Larson, E.; Kent, J.; Marx, E.; Swan, A. Soil C Sequestration as a Biological Negative Emission Strategy. Available online: https://www.frontiersin.org/articles/10.3389/fclim.2019.00008/full (accessed on 15 July 2020).
- Wacal, C.; Ogata, N.; Basalirwa, D.; Sasagawa, D.; Ishigaki, T.; Handa, T.; Kato, M.; Tenywa, M.M.; Masunaga, T.; Yamamoto, S.; et al. Imbalanced Soil Chemical Properties and Mineral Nutrition in Relation to Growth and Yield Decline of Sesame on Different Continuously Cropped Upland Fields Converted Paddy. Agronomy 2019, 9, 184. [Google Scholar] [CrossRef]
- Li, R.; Wang, J.J.; Zhang, Z.; Shen, F.; Zhang, G.; Qin, R.; Li, X.; Xiao, R. Nutrient transformations during composting of pig manure with bentonite. Bioresour. Technol. 2012, 121, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.S.; Pandey, V.C.; Singh, D.P. Coal fly ash and farmyard manure amendments in dry-land paddy agriculture field: Effect on N-dynamics and paddy productivity. Appl. Soil Ecol. 2011, 47, 133–140. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982. [Google Scholar] [CrossRef]
- Casida, L.E.J.; Klein, D.A.; Santoro, D. Soil Dehydrogenase Activity. Soil Sci. Annu. 1964, 98, 371–378. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.; Hantula, J. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycol. Res. 2000, 104, 927–936. [Google Scholar] [CrossRef]
- Hermansson, A.; Lindgren, P.-E. Quantification of Ammonia-Oxidizing Bacteria in Arable Soil by Real-Time PCR. Appl. Environ. Microbiol. 2001, 67, 972–976. [Google Scholar] [CrossRef] [PubMed]
- Brtnicky, M.; Dokulilova, T.; Holatko, J.; Pecina, V.; Kintl, A.; Latal, O.; Vyhnanek, T.; Prichystalova, J.; Datta, R. Long-Term Effects of Biochar-Based Organic Amendments on Soil Microbial Parameters. Agronomy 2019, 9, 747. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A RAPID METHOD OF TOTAL LIPID EXTRACTION AND PURIFICATION. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Oravecz, O.; Elhottova, D.; Kristufek, V.; Sustr, V.; Frouz, J.; Triska, J.; Marialigeti, K. Application of ARDRA and PLFA analysis in characterizing the bacterial communities of the food, gut and excrement of saprophagous larvae of Penthetria holosericea (Diptera: Bibionidae): A pilot study. Folia Microbiol. 2004, 49, 83–93. [Google Scholar] [CrossRef]
- R_CORE_TEAM. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H. Ggplot2: Elegant graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. Available online: https://rpkgs.datanovia.com/factoextra/ (accessed on 15 July 2020).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Mendiburu, d.F. Agricolae: Statistical Procedures for Agricultural Research. Available online: https://cran.r-project.org/web/packages/agricolae/index.html (accessed on 15 July 2020).
- Beaujean, A.A. R Package for Baylor University Educational Psychology Quantitative Courses. Available online: https://rdrr.io/cran/BaylorEdPsych/ (accessed on 15 July 2020).
- Ukaoma, A.A.; Ukaoma, V.O.; Opara, F.N.; Osuala, F.O.U. Inhibition of Dehydrogenase Activity in Pathogenic Bacteria Isolates by Aqueous Extract of Curcuma Longa (Turmeric) Rhizome. J. Phytopharm. 2013, 2, 9–17. [Google Scholar]
- Walls-Thumma, D. Dehydrogenase Activity in Soil Bacteria. Available online: https://www.intechopen.com/books/dehydrogenases/dehydrogenase-activity-in-the-soil-environment (accessed on 15 July 2020).
- Haderlein, S.B.; Weissmahr, K.W.; Schwarzenbach, R.P. Specific Adsorption of Nitroaromatic Explosives and Pesticides to Clay Minerals. Environ. Sci. Technol. 1996, 30, 612–622. [Google Scholar] [CrossRef]
- Chaerun, S.K.; Tazaki, K. How kaolinite plays an essential role in remediating oil-polluted seawater. In ENERGYO; De Gruyter: Berlin, Germany, 2018. [Google Scholar] [CrossRef]
- Makádi, M.; Tomócsik, A.; Orosz, V.; Bogdányi, Z.; Biró, B. Effect of a biogas-digestate and bentonite on some enzyme activities of the amended soils. Cereal Res. Commun. 2007, 35, 741–744. [Google Scholar] [CrossRef]
- Sarkar, B.; Megharaj, M.; Shanmuganathan, D.; Naidu, R. Toxicity of organoclays to microbial processes and earthworm survival in soils. J. Hazard. Mater. 2013, 261, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.Y.; Lin, X.G.; Fujii, T.; Morimoto, S.; Yagi, K.; Hu, J.L.; Zhang, J.B. Soil microbial biomass, dehydrogenase activity, bacterial community structure in response to long-term fertilizer management. Soil Biol. Biochem. 2007, 39, 2971–2976. [Google Scholar] [CrossRef]
- Tejada, M.; Hernandez, M.T.; Garcia, C. Application of two organic amendments on soil restoration: Effects on the soil biological properties. J. Environ. Qual. 2006, 35, 1010–1017. [Google Scholar] [CrossRef]
- Lu, Q.; Feng, X.; Sun, K.; Liao, Z. Study on the use of polymer/bentonite composites for controlled release. Plant Nutr. Fertil. Sci. 2005, 2, 183. [Google Scholar]
- Wangwang, L.; Chengcheng, H.; Huixing, X.; Yafan, B. Study on the Release Property of Nitrogen in Sustained-release Fertilizer with Carrier of Bentonite. In Proceedings of the 2013 Third International Conference on Intelligent System Design and Engineering Applications, Hong Kong, China, 16–18 January 2013. [Google Scholar]
- Li, H.F.; Chen, M.Q.; Fu, B.A.; Liang, B. Evaluation on the thermal and moisture diffusion behavior of sand/bentonite. Appl. Therm. Eng. 2019, 151, 55–65. [Google Scholar] [CrossRef]
- Kizilkaya, R.; Akça, İ.; Aşkın, T.; Olekhov, V.; Samofalova, I.; Mudrykh, N. Effect of Soil contamination with azadirachtin on dehydrogenase and catalase activity of soil. Eurasian J. Soil Sci. 2012, 1, 93–108. [Google Scholar]
- Bach, E.M.; Baer, S.G.; Meyer, C.K.; Six, J. Soil texture affects soil microbial and structural recovery during grassland restoration. Soil Biol. Biochem. 2010, 42, 2182–2191. [Google Scholar] [CrossRef]
- Hemkemeyer, M.; Pronk, G.J.; Heister, K.; Koegel-Knabner, I.; Martens, R.; Tebbe, C.C. Artificial soil studies reveal domain-specific preferences of microorganisms for the colonisation of different soil minerals and particle size fractions. FEMS Microbiol. Ecol. 2014, 90, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Lutzow, M.v.; Kogel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Müller, T.; Höper, H. Soil organic matter turnover as a function of the soil clay content: Consequences for model applications. Soil Biol. Biochem. 2004, 36, 877–888. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Q.; Lü, Y.; Zhang, X.; Liang, W. Contributions of soil biota to C sequestration varied with aggregate fractions under different tillage systems. Soil Biol. Biochem. 2013, 62, 147–156. [Google Scholar] [CrossRef]
- Oades, J.M. The retention of organic matter in soils. Biogeochemistry 1988, 5, 35–70. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K. Aggregate-associated soil organic matter as an ecosystem property and a measurement tool. Soil Biol. Biochem. 2014, 68, A4–A9. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Haney, R.L.; Hons, F.M.; Zuberer, D.A. Active fractions of organic matter in soils with different texture. Soil Biol. Biochem. 1996, 28, 1367–1372. [Google Scholar] [CrossRef]
- Dilustro, J.J.; Collins, B.; Duncan, L.; Crawford, C. Moisture and soil texture effects on soil CO2 efflux components in southeastern mixed pine forests. For. Ecol. Manag. 2005, 204, 87–97. [Google Scholar] [CrossRef]
- Baath, E.; Anderson, T.H. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 2003, 35, 955–963. [Google Scholar] [CrossRef]
- Fontaine, S.; Barot, S.; Barré, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef]
- Harris, J.A. Measurements of the soil microbial community for estimating the success of restoration. Eur. J. Soil Sci. 2003, 54, 801–808. [Google Scholar] [CrossRef]
- Alekseev, A.; Alekseeva, T.; Kalinin, P.; Hajnos, M. Soils response to the land use and soil climatic gradients at ecosystem scale: Mineralogical and geochemical data. Soil Tillage Res. 2018, 180, 38–47. [Google Scholar] [CrossRef]
- Vogel, C.; Heister, K.; Buegger, F.; Tanuwidjaja, I.; Haug, S.; Schloter, M.; Kogel-Knabner, I. Clay mineral composition modifies decomposition and sequestration of organic carbon and nitrogen in fine soil fractions. Biol. Fertil. Soils 2015, 51, 427–442. [Google Scholar] [CrossRef]
- Gonod, L.V.; Jones, D.L.; Chenu, C. Sorption regulates the fate of the amino acids lysine and leucine in soil aggregates. Eur. J. Soil Sci. 2006, 57, 320–329. [Google Scholar] [CrossRef]
- Morrissey, E.M.; McHugh, T.A.; Preteska, L.; Hayer, M.; Dijkstra, P.; Hungate, B.A.; Schwartz, E. Dynamics of extracellular DNA decomposition and bacterial community composition in soil. Soil Biol. Biochem. 2015, 86, 42–49. [Google Scholar] [CrossRef]
- Spence, A.; Robinson, C.; Hanson, R.E. The effects of microstructural changes on montmorillonite–microbial interactions. J. Mol. Struct. 2014, 1056–1057, 157–165. [Google Scholar] [CrossRef]
- Yang, H.; Tong, M.; Kim, H. Influence of Bentonite Particles on Representative Gram Negative and Gram Positive Bacterial Deposition in Porous Media. Environ. Sci. Technol. 2012, 46, 11627–11634. [Google Scholar] [CrossRef]
- Černá, K.; Ševců, A.; Steinová, J.; Burkartová, K. Microbial Diversity in Aged Bentonites; Technical University of Liberec: Liberec, Czechia, 2018. [Google Scholar]
- Stroes-Gascoyne, S.; Haveman, S.A.; Hamon, C.J.; Delaney, T.-L.; Pedersen, K.; Arlinger, J.; Ekendahl, S.; Hallbeck, L.; Jahromi, N.; Dekeyser, K.; et al. Occurrence and identification of microorganisms in compacted clay-based buffer material designed for use in a nuclear fuel waste disposal vault. Can. J. Microbiol. 1997, 43, 1133–1146. [Google Scholar] [CrossRef]
- Stroes-Gascoyne, S.; West, J.M. An overview of microbial research related to high-level nuclear waste disposal with emphasis on the Canadian concept for the disposal of nuclear fuel waste. Can. J. Microbiol. 1996, 42, 349–366. [Google Scholar] [CrossRef]
- Haynes, R.J.; Naidu, R. Influence of lime, fertilizer and manure applications on soil organic matter content and soil physical conditions: A review. Nutr. Cycl. Agroecosyst. 1998, 51, 123–137. [Google Scholar] [CrossRef]
- Engel, K.; Ford, S.E.; Coyotzi, S.; McKelvie, J.; Diomidis, N.; Slater, G.; Neufeld, J.D. Stability of Microbial Community Profiles Associated with Compacted Bentonite from the Grimsel Underground Research Laboratory. MSphere 2019, 4, e00601–e00619. [Google Scholar] [CrossRef] [PubMed]
- Marschner, P.; Kandeler, E.; Marschner, B. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biol. Biochem. 2003, 35, 453–461. [Google Scholar] [CrossRef]
- Poll, C.; Thiede, A.; Wermbter, N.; Sessitsch, A.; Kandeler, E. Micro-scale distribution of microorganisms and microbial enzyme activities in a soil with long-term organic amendment. Eur. J. Soil Sci. 2003, 54, 715–724. [Google Scholar] [CrossRef]
- Moosavi, M. Bentonite Clay as a Natural Remedy: A Brief Review. Iran. J. Public Health 2017, 46, 1176–1183. [Google Scholar] [PubMed]
- Rättö, M.; Itävaara, M.; Nol, E. Microbial Activity in Bentonite Buffers Literature Study; Technical University of Liberec: Liberec, Czechia, 2012. [Google Scholar]
- Schutter, M.; Dick, R. Microbial Community Profiles and Activities among Aggregates of Winter Fallow and Cover-Cropped Soil. Soil Sci. Soc. Am. J. SSSAJ 2002, 66, 142–153. [Google Scholar] [CrossRef]
- Tisdall, J.; Oades, J. Organic Matter and Water-stable Aggregates in Soils. J. Soil Sci. 2006, 33, 141–163. [Google Scholar] [CrossRef]
- Birkhofer, K.; Bezemer, T.M.; Bloem, J.; Bonkowski, M.; Christensen, S.; Dubois, D.; Ekelund, F.; Fliessbach, A.; Gunst, L.; Hedlund, K.; et al. Long-term organic farming fosters below and aboveground biota: Implications for soil quality, biological control and productivity. Soil Biol. Biochem. 2008, 40, 2297–2308. [Google Scholar] [CrossRef]
- Miller, J.D. Fungi as contaminants in indoor air. Atmos. Environ. Part A. Gen. Top. 1992, 26, 2163–2172. [Google Scholar] [CrossRef]
- Whalen, J.; Chang, C. Macroaggregate Characteristics in Cultivated Soils after 25 Annual Manure Applications. Soil Sci. Soc. Am. J. 2002, 66, 1637–1647. [Google Scholar] [CrossRef]
- Sawhney, B.L. Selective Sorption and Fixation of Cations by Clay Minerals: A Review. Clays Clay Miner. 1972, 20, 93–100. [Google Scholar] [CrossRef]
- Plaza, C.; Giannetta, B.; Fernández, J.M.; López-de-Sá, E.G.; Polo, A.; Gascó, G.; Méndez, A.; Zaccone, C. Response of different soil organic matter pools to biochar and organic fertilizers. Agric. Ecosyst. Environ. 2016, 225, 150–159. [Google Scholar] [CrossRef]
- Hallin, S.; Jones, C.; Schloter, M.; Philippot, L. Relationship between N-cycling communities and ecosystem functioning in a 50-year-old fertilization experiment. ISME J. 2009, 3, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, M.I. Soil Clay Mineralogies in Relation to Fertility Management: Effect of Clay Mineral Types on Ammonium Fixation under Conditions of Wetland Rice Culture1. Agron. J. 1907, 74, 143–144. [Google Scholar] [CrossRef]
- Meunier, A.; Velde, B.D. Illite-Origins, Evolution and Metamorphism, 1st ed.; Springer: Berlin, Germany, 2004. [Google Scholar] [CrossRef]
- Barthod, J. Innovative Waste Treatment by Composting with Minerals and Worms: Effects on Carbon Storage, Soil Properties and Plant Growth. Ph.D. Thesis, Université Pierre et Marie Curie, Paris, France, 2017. [Google Scholar]
- Skála, J.; Vácha, R.; Hofman, J.; Horváthová, V.; Sáňka, M.; Čechmánková, J. Spatial differentiation of ecosystem risks of soil pollution in floodplain areas of the Czech Republic. Soil Water Res. 2017, 12, 1–9. [Google Scholar] [CrossRef]
- Gutarowska, B.; Matusiak, K.; Borowski, S.; Rajkowska, A.; Brycki, B. Removal of odorous compounds from poultry manure by microorganisms on perlite-bentonite carrier. J. Environ. Manag. 2014, 141, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Biswas, B.; Sarkar, B.; Rusmin, R.; Naidu, R. Bioremediation of PAHs and VOCs: Advances in clay mineral-microbial interaction. Environ. Int. 2015, 85, 168–181. [Google Scholar] [CrossRef]
- Briones, R.M.; Sarmah, A.K. Insight into the sorption mechanism of metformin and its transformation product guanylurea in pastoral soils and model sorbents. Sci. Total Environ. 2018, 645, 1323–1333. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Physical Properties | Soil Chemical Properties | ||||
|---|---|---|---|---|---|
| Parameter | Value | Unit | Parameter | Value | Unit |
| Bulk density | 1.38 | g/cm3 | C:N ratio | 5.89 | - |
| Porosity | 47.7 | % | K:Mg ratio | 0.58 | - |
| Maximum capillary capacity | 35.8 | % | Ntot | 1.9 | g/kg |
| Minimum airiness | 12.0 | % | Available P | 95.9 | mg/kg |
| Available N | 238 | mg/kg | |||
| Content of particles (<0.01 mm) | 27 | % | Exchange capacity | 122 | mmol/kg |
| Mg | 145 | mg/kg | |||
| Ca | 2.6 | mg/kg | |||
| Corg | 11.2 | g/kg | |||
| Humic and fulvic acid ratio | 0.93 | - | |||
| pH (KCl) | 5.13 | - | |||
| Soil Microbial Properties | Descriptive Statistics | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SEM | SD | Min | 5% | 25% | 50% | 75% | 95% | Max | |
| DHA | 1.75 | 0.07 | 0.33 | 1.23 | 1.26 | 1.49 | 1.75 | 2.03 | 2.20 | 2.23 |
| Cmic | 1.65 | 0.05 | 0.23 | 1.31 | 1.32 | 1.48 | 1.62 | 1.76 | 2.07 | 2.12 |
| BPLFA | 55.6 | 1.26 | 6.16 | 45.1 | 45.4 | 52.8 | 55.0 | 62.0 | 63.1 | 63.6 |
| FPLFA | 24.3 | 1.05 | 5.12 | 17.9 | 18.3 | 19.6 | 22.7 | 30.5 | 31.3 | 32.0 |
| 16S rDNA | 9.42 | 0.18 | 0.86 | 8.47 | 8.49 | 8.89 | 8.98 | 10.6 | 10.6 | 10.6 |
| 18S rDNA | 7.49 | 0.22 | 1.08 | 6.39 | 6.42 | 6.46 | 7.29 | 8.73 | 8.76 | 8.76 |
| 16S AOB | 8.16 | 0.14 | 0.67 | 6.95 | 6.95 | 7.89 | 8.29 | 8.75 | 8.81 | 8.82 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Datta, R.; Holatko, J.; Latal, O.; Hammerschmiedt, T.; Elbl, J.; Pecina, V.; Kintl, A.; Balakova, L.; Radziemska, M.; Baltazar, T.; et al. Bentonite-Based Organic Amendment Enriches Microbial Activity in Agricultural Soils. Land 2020, 9, 258. https://doi.org/10.3390/land9080258
Datta R, Holatko J, Latal O, Hammerschmiedt T, Elbl J, Pecina V, Kintl A, Balakova L, Radziemska M, Baltazar T, et al. Bentonite-Based Organic Amendment Enriches Microbial Activity in Agricultural Soils. Land. 2020; 9(8):258. https://doi.org/10.3390/land9080258
Chicago/Turabian StyleDatta, Rahul, Jiri Holatko, Oldrich Latal, Tereza Hammerschmiedt, Jakub Elbl, Vaclav Pecina, Antonin Kintl, Ludmila Balakova, Maja Radziemska, Tivadar Baltazar, and et al. 2020. "Bentonite-Based Organic Amendment Enriches Microbial Activity in Agricultural Soils" Land 9, no. 8: 258. https://doi.org/10.3390/land9080258
APA StyleDatta, R., Holatko, J., Latal, O., Hammerschmiedt, T., Elbl, J., Pecina, V., Kintl, A., Balakova, L., Radziemska, M., Baltazar, T., Skarpa, P., Danish, S., Zafar-ul-Hye, M., Vyhnanek, T., & Brtnicky, M. (2020). Bentonite-Based Organic Amendment Enriches Microbial Activity in Agricultural Soils. Land, 9(8), 258. https://doi.org/10.3390/land9080258