Is There an Equivalence between Measures of Landscape Structural and Functional Connectivity for Plants in Conservation Assessments of the Cerrado?

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
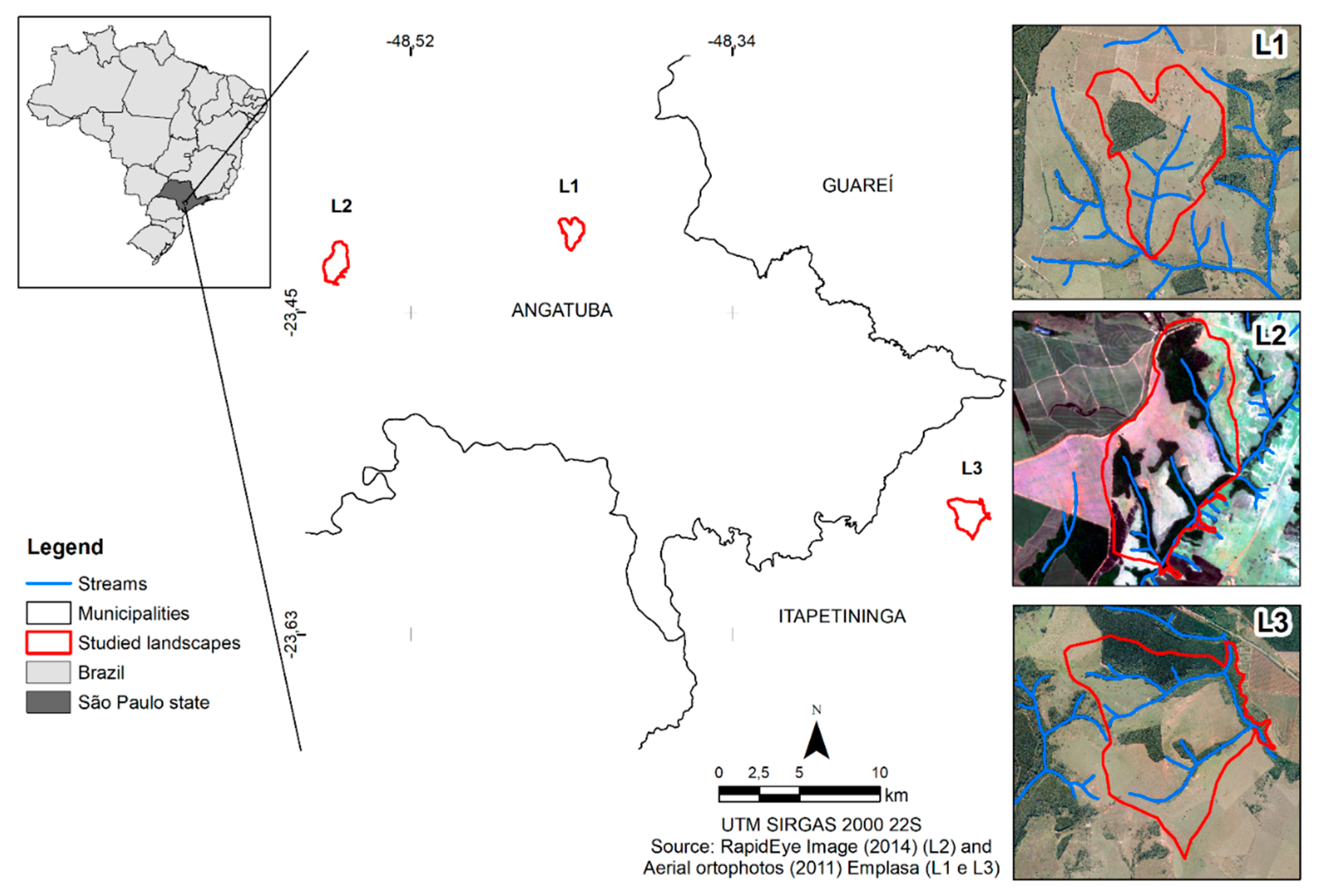
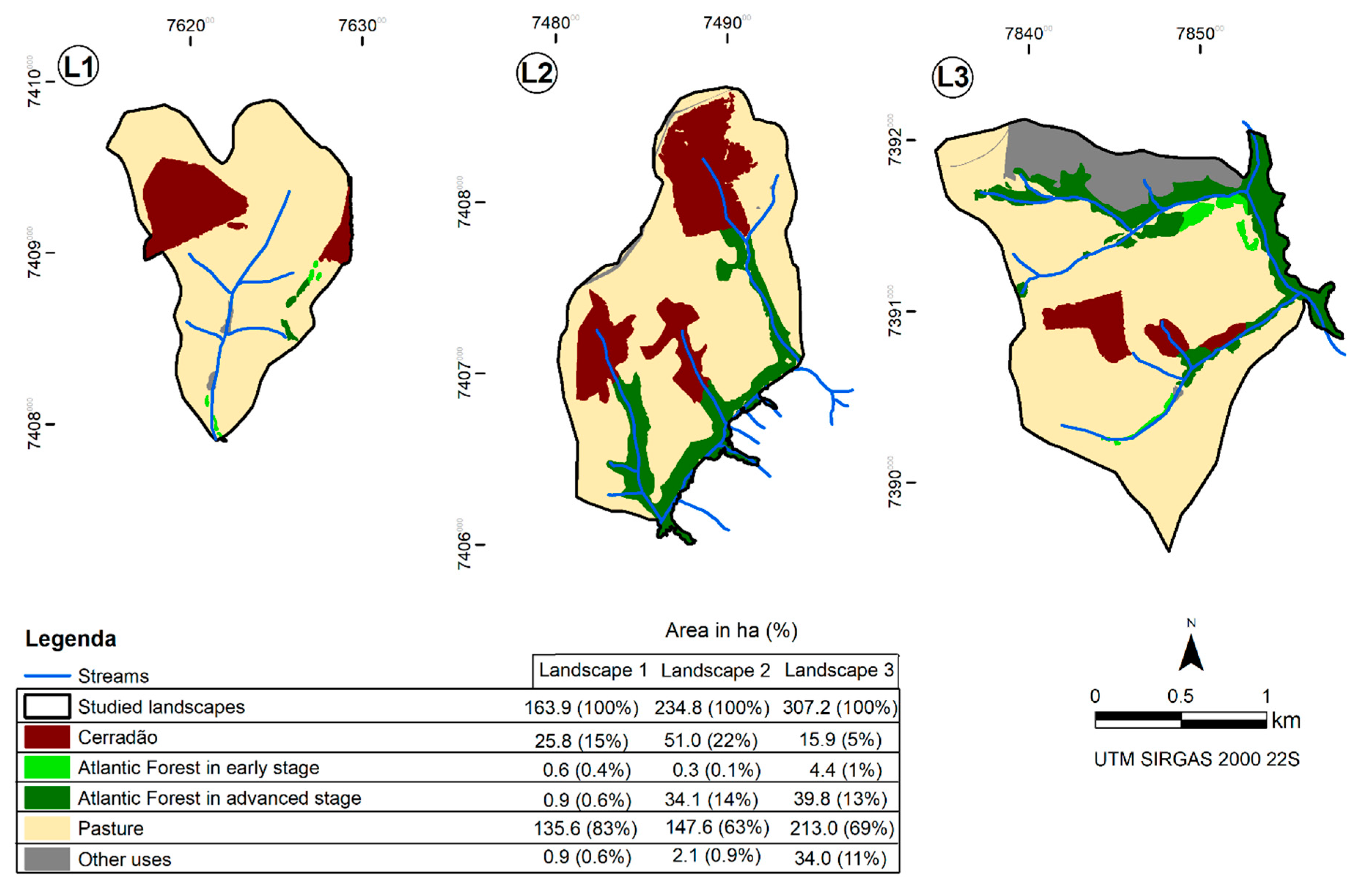
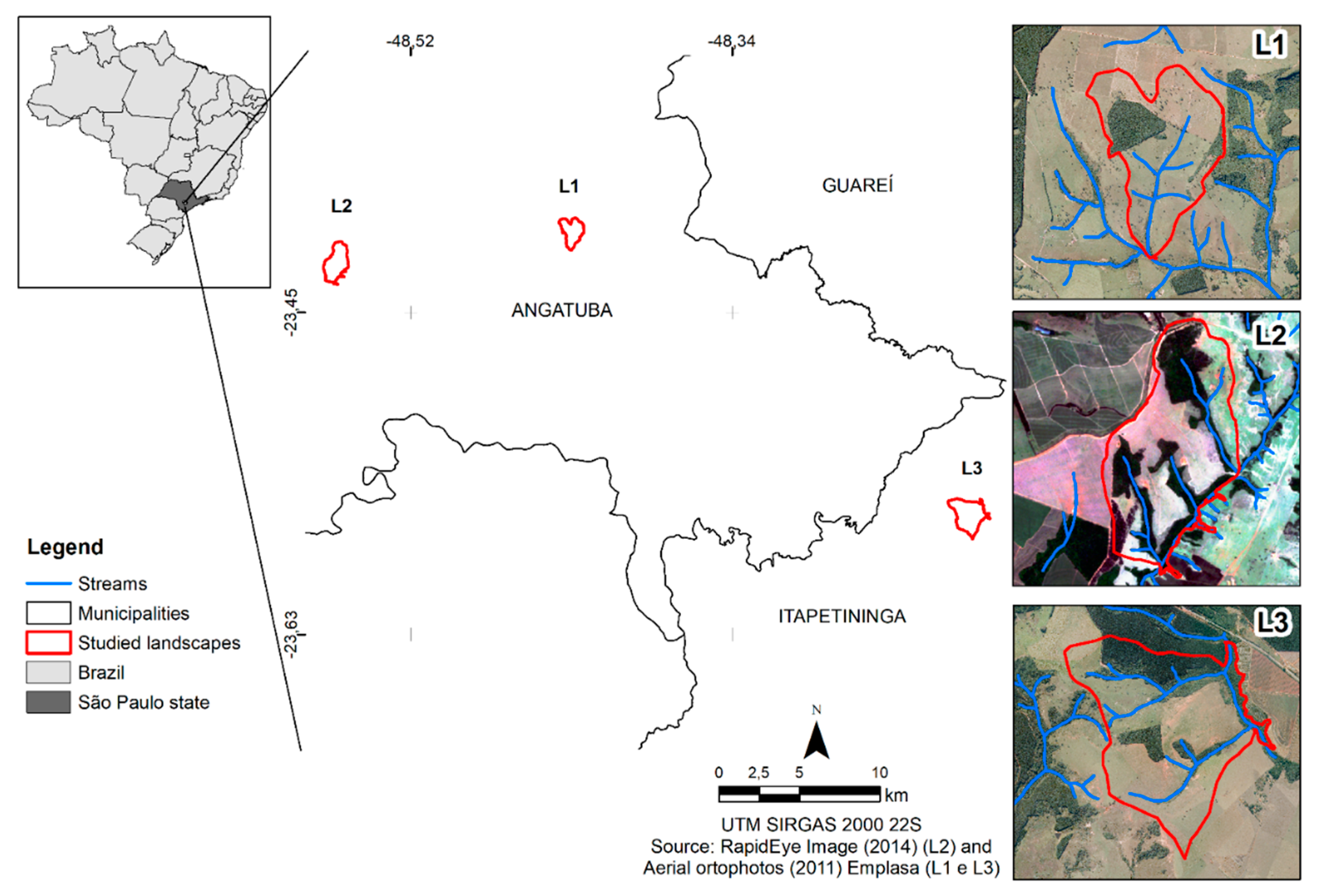
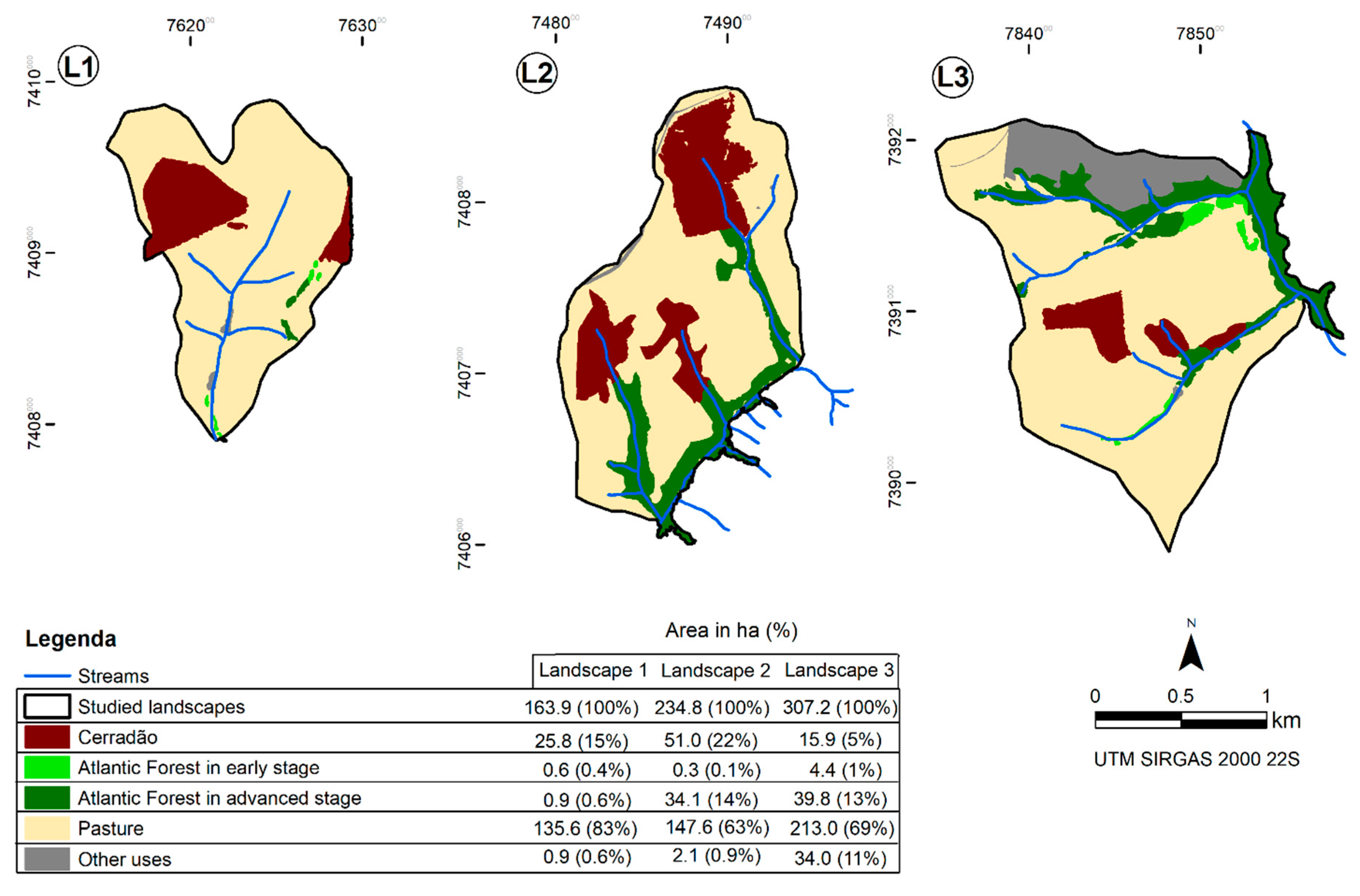
2.1. Selection and Mapping of the Study Area
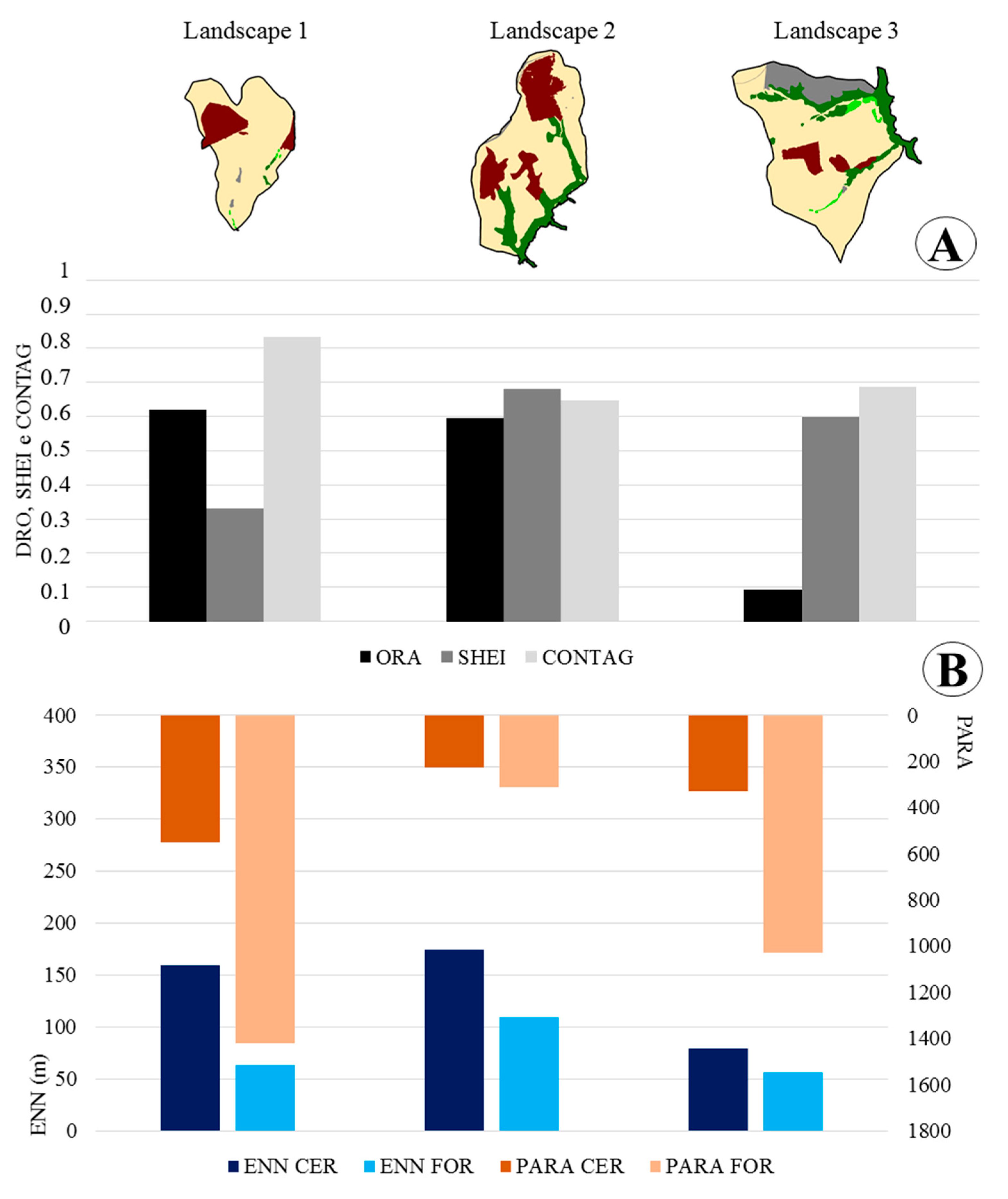
2.2. Measures of Landscape Structure
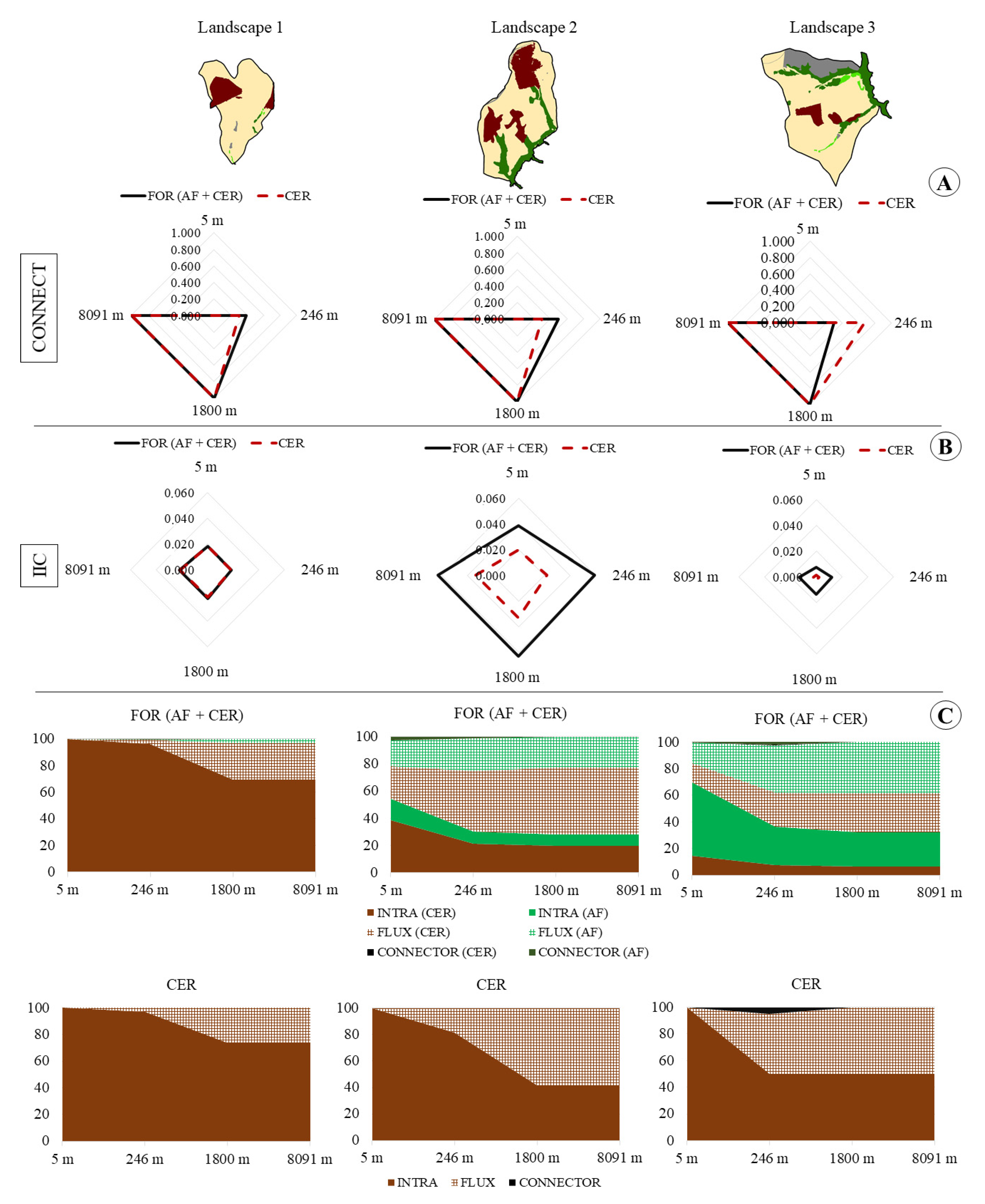
2.3. Measures of Landscape Functional Connectivity
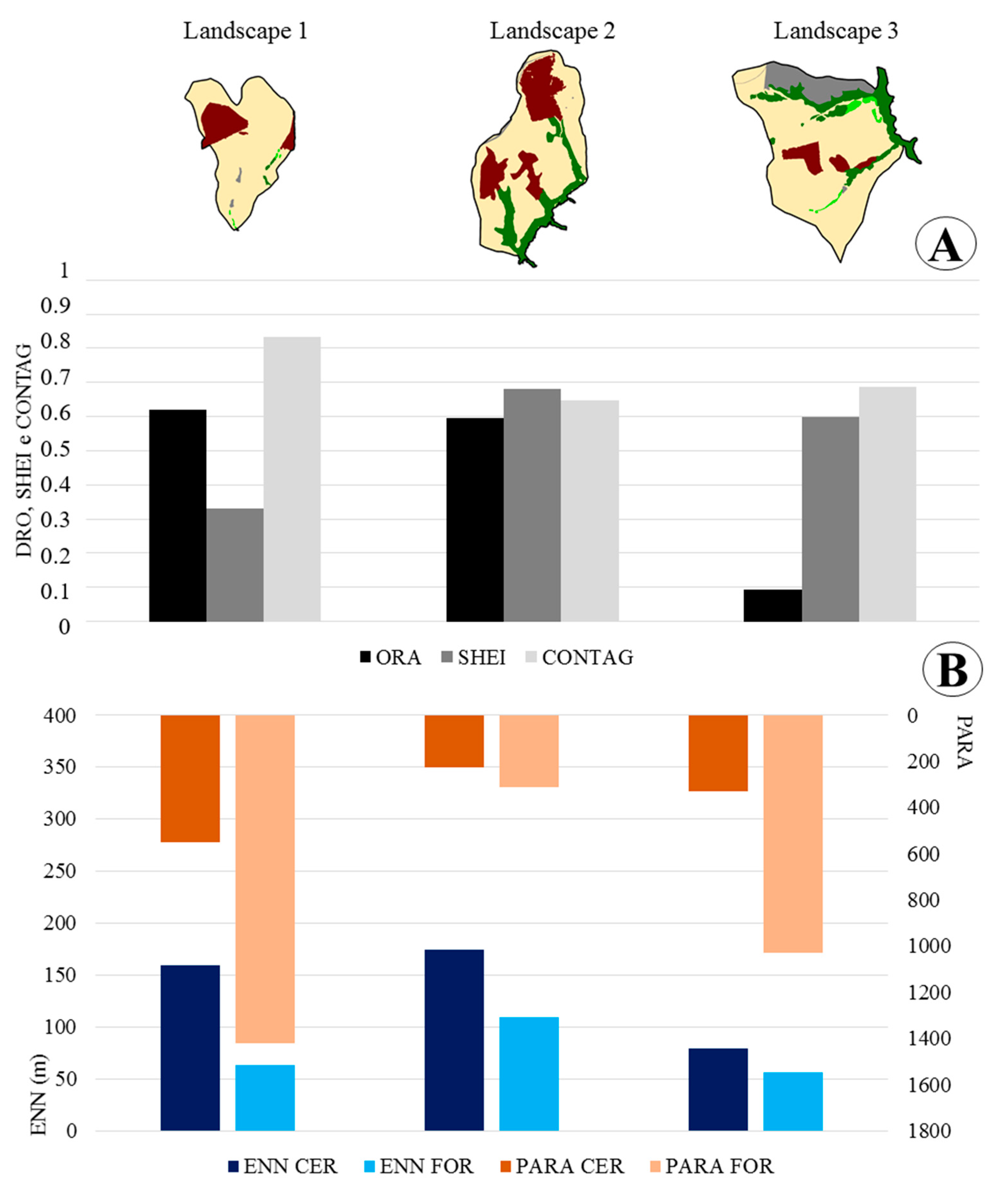
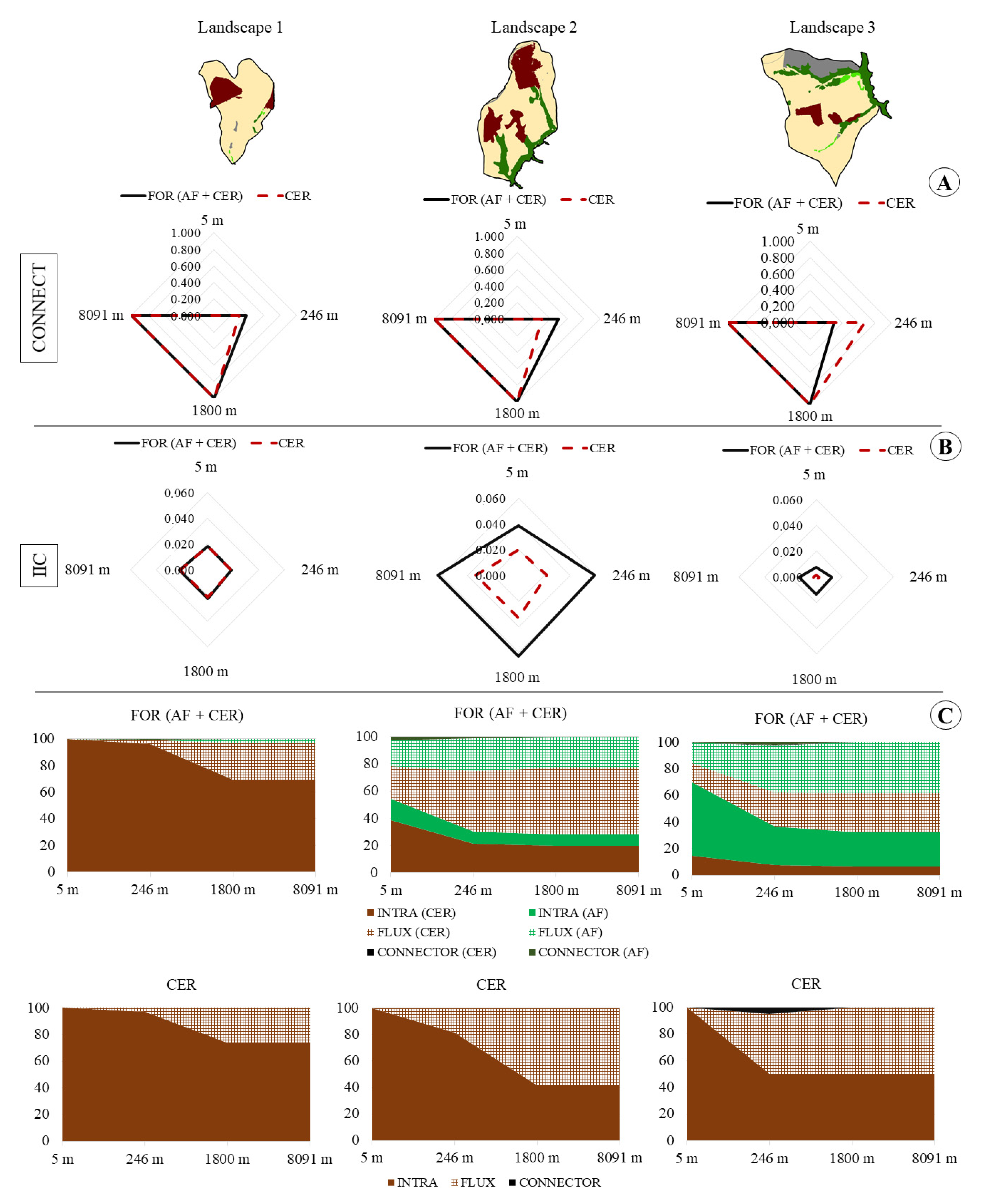
3. Results
4. Discussion
5. Practical Implications for Biodiversity Conservation Planning
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landscape 1 | Landscape 2 | Landscape 3 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LANDSCAPE STRUCTURE | ||||||||||||
| ORAFOR | 0.62 | 0.59 | 0.09 | |||||||||
| NPFOR | 11 | 5 | 15 | |||||||||
| NPCER | 3 | 3 | 3 | |||||||||
| PARAFOR | 1419.40 | 311.99 | 1029.51 | |||||||||
| PARACER | 552.87 | 228.59 | 330.64 | |||||||||
| ENN_MNFOR | 62.92 | 109.32 | 55.91 | |||||||||
| ENNCER | 159.41 | 174.26 | 79.24 | |||||||||
| SHEI | 0.53 | 0.94 | 0.98 | |||||||||
| CONTAG | 83.19 | 64.67 | 68.86 | |||||||||
| FUNCTIONAL CONNECTIVITY | ||||||||||||
| 5 m | 246 m | 1800 m | 8091 m | 5 m | 246 m | 1800 m | 8091 m | 5 m | 246 m | 1800 m | 8091 m | |
| CONNECTFOR | 0 | 0.38 | 1 | 1 | 0 | 0.5 | 1 | 1 | 0 | 0.30 | 1 | 1 |
| CONNECTCER | 0 | 0.33 | 1 | 1 | 0 | 0.33 | 1 | 1 | 0 | 0.67 | 1 | 1 |
| IICFOR | 0.018 | 0.019 | 0.022 | 0.022 | 0.038 | 0.059 | 0.063 | 0.063 | 0.008 | 0.012 | 0.014 | 0.014 |
| IICCER | 0.018 | 0.018 | 0.021 | 0.021 | 0.020 | 0.022 | 0.033 | 0.033 | 0.0014 | 0.0020 | 0.0020 | 0.0020 |
| Variation (%) | 0.03 | 0.48 | 3.57 | 3.57 | 49.00 | 63.11 | 47.04 | 47.04 | 83.05 | 83.93 | 85.06 | 85.06 |
| Intra FOR | 100 | 98.15 | 81.87 | 81.87 | 71.77 | 46.70 | 43.73 | 43.73 | 82.29 | 54.03 | 48.50 | 48.50 |
| Flux FOR | 0.0039 | 3.70 | 36.25 | 36.25 | 56.46 | 106.60 | 112.54 | 112.54 | 35.41 | 91.93 | 103.00 | 103.00 |
| Connector FOR | 0 | 0.033 | 0 | 0 | 3.80 | 1.91 | 0 | 0 | 0.76 | 3.50 | 0 | 0 |
| Intra CER | 100 | 98.59 | 84.88 | 84.88 | 100 | 89.95 | 58.67 | 58.67 | 100 | 69.26 | 66.89 | 66.89 |
| Flux CER | 0 | 2.81 | 30.23 | 30.23 | 0 | 20.10 | 82.65 | 82.65 | 0 | 61.48 | 66.22 | 66.22 |
| Connector CER | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7.08 | 0 | 0 |
References
- De la Cruz, M.; Maestre, F.T. Avances en el Análisis Espacial de Datos Ecológicos: Aspectos Metodológicos y Aplicados (In Spainish); Ecological Asociation of Terrerstrial Ecology-AEET: Madrid, Spain, 2013; ISBN 9788461634484. [Google Scholar]
- Hardt, E.; Borgomeo, E.; dos Santos, R.F.; Pinto, L.F.G.; Metzger, J.P.; Sparovek, G. Does certification improve biodiversity conservation in Brazilian coffee farms? For. Ecol. Manage. 2015, 357, 181–194. [Google Scholar] [CrossRef]
- With, K.A.; Gardner, R.H.; Turner, M.G. Landscape Connectivity and Population Distributions in Heterogeneous Environments. Oikos 1997, 78, 151–169. [Google Scholar] [CrossRef] [Green Version]
- Boscolo, D.; Ferreira, P.A.; Lopes, L.E. Da matriz à matiz: Em busca de uma abordagem funcional na Ecologia de Paisagens. Filos. e História da Biol. 2016, 11, 157–187. [Google Scholar]
- Herrmann, J.D.; Haddad, N.M.; Levey, D.J. Testing the relative importance of local resources and landscape connectivity on Bombus impatiens (Hymenoptera, Apidae) colonies. Apidologie 2017, 48, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Mikoláš, M.; Tejkal, M.; Kuemmerle, T.; Griffiths, P.; Svoboda, M.; Hlásny, T.; Leitão, P.J.; Morrissey, R.C. Forest management impacts on capercaillie (Tetrao urogallus) habitat distribution and connectivity in the Carpathians. Landsc. Ecol. 2017, 32, 163–179. [Google Scholar] [CrossRef]
- Velázquez, J.; Gutiérrez, J.; Hernando, A.; García-Abril, A. Evaluating landscape connectivity in fragmented habitats: Cantabrian capercaillie (Tetrao urogallus cantabricus) in northern Spain. For. Ecol. Manage. 2017, 389, 59–67. [Google Scholar] [CrossRef]
- Ramirez-Reyes, C.; Bateman, B.L.; Radeloff, V.C. Effects of habitat suitability and minimum patch size thresholds on the assessment of landscape connectivity for jaguars in the Sierra Gorda, Mexico. Biol. Conserv. 2016, 204, 296–305. [Google Scholar] [CrossRef]
- de la Torre, J.A.; Núñez, J.M.; Medellín, R.A. Habitat availability and connectivity for jaguars (Panthera onca) in the Southern Mayan Forest: Conservation priorities for a fragmented landscape. Biol. Conserv. 2016, 206, 270–282. [Google Scholar] [CrossRef]
- Brudvig, L.A.; Damschen, E.I.; Tewksbury, J.J.; Haddad, N.M.; Levey, D.J. Landscape connectivity promotes plant biodiversity spillover into non-target habitats. Proc. Natl. Acad. Sci. USA 2009, 106, 9328–9332. [Google Scholar] [CrossRef] [Green Version]
- Auffret, A.G.; Rico, Y.; Bullock, J.M.; Hooftman, D.A.P.; Pakeman, R.J.; Soons, M.B.; Suárez-Esteban, A.; Traveset, A.; Wagner, H.H.; Cousins, S.A.O. Plant functional connectivity–integrating landscape structure and effective dispersal. J. Ecol. 2017, 105, 1648–1656. [Google Scholar] [CrossRef]
- Damschen, E.I.; Baker, D.V.; Bohrer, G.; Nathan, R.; Orrock, J.L.; Turner, J.R.; Brudvig, L.A.; Haddad, N.M.; Levey, D.J.; Tewksbury, J.J. How fragmentation and corridors affect wind dynamics and seed dispersal in open habitats. Proc. Natl. Acad. Sci. USA 2014, 111, 3484–3489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trakhtenbrot, A.; Katul, G.G.; Nathan, R. Mechanistic modeling of seed dispersal by wind over hilly terrain. Ecol. Modell. 2014. [Google Scholar] [CrossRef]
- McConkey, K.R.; Prasad, S.; Corlett, R.T.; Campos-Arceiz, A.; Brodie, J.F.; Rogers, H.; Santamaria, L. Seed dispersal in changing landscapes. Biol. Conserv. 2012, 146, 1–13. [Google Scholar] [CrossRef]
- McGarigal, K.; Marks, B.J. FRAGSTATS: Spatial Pattern Analysis Program for Quantifying Landscape Structure; US Department of Agriculture, Forest Service, Pacific Northwest Research Station: Corvallis, OR, USA, 1995. [Google Scholar]
- Guo, X.; Coops, N.C.; Gergel, S.E.; Bater, C.W.; Nielsen, S.E.; Stadt, J.J.; Drever, M. Integrating airborne lidar and satellite imagery to model habitat connectivity dynamics for spatial conservation prioritization. Landsc. Ecol. 2018, 33, 491–511. [Google Scholar] [CrossRef]
- Saura, S.; Torné, J. Conefor Sensinode 2.2: A software package for quantifying the importance of habitat patches for landscape connectivity. Environ. Model. Softw. 2009, 24, 135–139. [Google Scholar] [CrossRef]
- Saura, S.; Rubio, L.S. A common currency for the different ways in which patches and links can contribute to habitat availability and connectivity in the landscape. Ecography (Cop.) 2010, 33, 523–537. [Google Scholar] [CrossRef]
- Gatica-Saavedra, P.; Echeverría, C.; Nelson, C.R. Ecological indicators for assessing ecological success of forest restoration: A world review. Restor. Ecol. 2017, 25, 850–857. [Google Scholar] [CrossRef]
- IBGE Mapa de Biomas; Instituto Brasileiro de Geografia e Estatística: Rio de Janeiro, Brazil. 2004. Available online: https://www.ibge.gov.br/geociencias/informacoes-ambientais/15842-biomas.html?=&t=downloads (accessed on 1 February 2020).
- Strassburg, B.B.N.; Brooks, T.; Feltran-Barbieri, R.; Iribarrem, A.; Crouzeilles, R.; Loyola, R.D.; Latawiec, A.E.; Oliveira Filho, F.J.B.; De Scaramuzza, C.A.M.; Scarano, F.R.; et al. Moment of truth for the Cerrado hotspot. Nat. Ecol. Evol. 2017, 1, 1–3. [Google Scholar] [CrossRef]
- IBGE Censo agropecuário: Resultados preliminares. Inst. Bras. Geogr. e Estatística. 2017, 7, 1–180.
- Taylor, P.D.; Fahrig, L.; With, K.A. Landscape connectivity: A return to the basics. In Connectivity Conservation; Cambridge University Press: Cambridge, UK, 2010; pp. 29–43. [Google Scholar] [CrossRef]
- Kronka, F.J.N. Inventário Florestal da Vegetação Natural do Estado de São Paulo; Secretaria do Meio Ambiente: Instituto Florestal. Impensa Oficial: São Paulo, Brazil, 2005; p. 200. [Google Scholar]
- Attanasio, C.M.; Lima, W.D.P.; Gandolfi, S.; Zakia, M.J.; Veniziani Júnior, J.C.T. Método para a identificação da zona ripária: Microbacia hidrográfica do Ribeirão São João (Mineiros do Tietê, SP). Sci. Florestalis 2006, 71, 131–140. [Google Scholar]
- Nascimbene, J.; Marini, L.; Ivan, D.; Zottini, M. Management Intensity and Topography Determined Plant Diversity in Vineyards. PLoS ONE 2013, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheesman, A.W.; Preece, N.D.; van Oosterzee, P.; Erskine, P.D.; Cernusak, L.A. The role of topography and plant functional traits in determining tropical reforestation success. J. Appl. Ecol. 2018, 55, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Penrod, K.; Cabañero, C.; Beier, P.; Luke, C.; Spencer, W.; Rubin, E.; Sauvajot, R.; Riley, S.; Kamradt, D. South Coast Missing Linkages Project: A Linkage Design for the Santa Monica-Sierra Madre Connection; South Coast Wildlands: Idyllwild, CA, USA, 2006. [Google Scholar]
- Inventário Florestal. Apresenta a Distribuição dos Remanescentes de Vegetação Natural Existentes no Estado de São Paulo, Classificados por Fitofisionomia. Mapeamento Realizado Com Imagens de Satélite 2008 e 2009, Escala de Interpretação 1:25.000. 2010. Available online: http://snif.florestal.gov.br/pt-br/ (accessed on 1 May 2019).
- SMA Arquivos Shapefile Disponibilizados pela Coordenadoria de Planejamento Ambiental: São Paulo, Brazil. 2018. Available online: https://www.infraestruturameioambiente.sp.gov.br/cpla/> (accessed on 1 November 2018).
- Mendonça, R.A.; Costa, C.G. O negligenciado cerrado paulista. Rev. Bras. Geogr. 2018, 63, 129–155. [Google Scholar] [CrossRef] [Green Version]
- MMA Mapeamento do uso e cobertura do Cerrado: Projeto TerraClass Cerrado 2013; MMA: São Paulo, Brazil, 2015; ISBN 978-85-7738-255-2.
- Brasil Lei no 12.651, de 25 de maio de 2012. Dispõe Sobre a Proteção da Vegetação Nativa; Altera as Leis Nos 6.938, de 31 de Agosto de 1981, 9.393, de 19 de Dezembro de 1996, e 11.428, de 22 de Dezembro de 2006; Revoga as Leis nos 4.771, de 15 de Setembro de 1965. Casa Civil. 2012. Available online: http://www.planalto.gov.br/ccivil_03/_Ato2011-2014/2012/Lei/L12651.htm (accessed on 30 January 2019).
- Hardt, E.; Ferreira dos Santos, R.; Lopes Pereira-Silva, E.F. Landscape changes in Serra do Japi: Legal protection or scientific expectation? Bosque (Valdivia) 2012, 33, 37–38. [Google Scholar] [CrossRef] [Green Version]
- Issii, T.M.; Romero, A.C.; Pereira-Silva, E.F.L.; Attanasio, M.R.; Hardt, E. The role of legal protection in forest conservation in an urban matrix. Land Use Policy 2020, 91, 104366. [Google Scholar] [CrossRef]
- Alvim, A.T.B.; Kato, V.R.C.; Rosin, J.R.D.G. A urgência das águas: Intervenções urbanas em áreas de mananciais. Cad. Metrópole 2015, 17, 83–107. [Google Scholar] [CrossRef]
- Alvim, A.T.B.; Bruna, G.C.; Kato, V.R.C. Políticas ambientais e urbanas em áreas de mananciais: Interfaces e conflitos. Cad. Metrópole 2008, 143, 143–164. [Google Scholar]
- IDEA-SP Infraestrutura de Dados Espaciais Ambientais do Estado de São Paulo: Base Territorial Ambiental Unificada (Geoportal–DataGEO); 2019. Available online: https://datageo.ambiente.sp.gov.br/ (accessed on 30 January 2019).
- Hardt, E.; Dos Santos, R.F.; Pereira-Silva, E.F.L. Evaluating the ecological effects of social agent scenarios for a housing development in the Atlantic Forest. Ecol. Indic. 2014, 36, 120–130. [Google Scholar] [CrossRef]
- Martinez del Castillo, E.; García-Martin, A.; Longares Aladrén, L.A.; de Luis, M. Evaluation of forest cover change using remote sensing techniques and landscape metrics in Moncayo Natural Park (Spain). Appl. Geogr. 2015, 62, 247–255. [Google Scholar] [CrossRef]
- Romero, A.C.; Issii, T.M.; Pereira-Silva, E.F.L.; Hardt, E. Effects of Urban Sprawl on Forest Conservation in a Metropolitan Water Source Area. Rev. Árvore 2018, 42. [Google Scholar] [CrossRef]
- Hardt, E.; dos Santos, R.F.; de Pablo, C.L.; de Agar, P.M.; Pereira-Silva, E.F.L. Utility of landscape mosaics and boundaries in forest conservation decision making in the Atlantic Forest of Brazil. Landsc. Ecol. 2013, 28, 385–399. [Google Scholar] [CrossRef]
- Tarazi, R.; Sebbenn, A.M.; Kageyama, P.Y.; Vencovsky, R. Long-distance dispersal in a fire- and livestock-protected savanna. Ecol. Evol. 2013, 3, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Moraes, M.A.; Kubota, T.Y.K.; Rossini, B.C.; Marino, C.L.; Freitas, M.L.M.; Moraes, M.L.T.; Da Silva, A.M.; Cambuim, J.; Sebbenn, A.M. Long-distance pollen and seed dispersal and inbreeding depression in Hymenaea stigonocarpa (Fabaceae: Caesalpinioideae) in the Brazilian savannah. Int. J. Bus. Innov. Res. 2018, 17, 7800–7816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golin, V.; Santos-Filho, M.; Pereira, M.J.B. Dispersal and predation of araticum seeds in the Cerrado of Mato Grosso, Brazil. Cienc. Rural 2011, 41, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Flora do Brasil 2020 em Construção. Jardim Botânico do Rio de Janeiro: Rio de Janeiro, Brazil. 2020. Available online: http://floradobrasil.jbrj.gov.br/reflora/listaBrasil/ConsultaPublicaUC/ConsultaPublicaUC.do (accessed on 25 May 2020).
- Vittoz, P.; Engler, R. Seed dispersal distances: A typology based on dispersal modes and plant traits. Bot. Helv. 2007, 117, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Saura, S.; Torne, J. CONEFOR 2.6 User Manual. 2012, pp. 1–19. Available online: http://www.conefor.org/files/usuarios/Manual_Conefor_26.pdf (accessed on 18 November 2020).
- Urban, D.; Keitt, T. Landscape Connectivity: A Graph-Theoretic Perspective. Ecology 2001, 82, 1205. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Wilson, M.C.; Chen, X.Y.; Corlett, R.T.; Didham, R.K.; Ding, P.; Holt, R.D.; Holyoak, M.; Hu, G.; Hughes, A.C.; Jiang, L.; et al. Habitat fragmentation and biodiversity conservation: Key findings and future challenges. Landsc. Ecol. 2016, 31, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Wu, J. Key concepts and research topics in landscape ecology revisited: 30 years after the Allerton Park workshop. Landsc. Ecol. 2013, 28, 1–11. [Google Scholar] [CrossRef]
- Andrén, H. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: A review. Oikos 1994, 71, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. Rethinking patch size and isolation effects: The habitat amount hypothesis. J. Biogeogr. 2013, 40, 1649–1663. [Google Scholar] [CrossRef]
- Rocha-Santos, L.; Pessoa, M.S.; Cassano, C.R.; Talora, D.C.; Orihuela, R.L.L.; Mariano-neto, E.; Morante-filho, J.C.; Faria, D.; Cazetta, E. The shrinkage of a forest: Landscape-scale deforestation leading to overall changes in local forest structure. BIOC 2016, 196, 1–9. [Google Scholar] [CrossRef]
- Yin, D.; Leroux, S.J.; He, F. Methods and models for identifying thresholds of habitat loss. Ecography (Cop.) 2017, 40, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Boesing, A.L.; Nichols, E.; Metzger, J.P. Biodiversity extinction thresholds are modulated by matrix type. Ecography (Cop.) 2018, 41, 1520–1533. [Google Scholar] [CrossRef] [Green Version]
- Huggett, A.J. The concept and utility of “ecological thresholds” in biodiversity conservation. Biol. Conserv. 2005, 124, 301–310. [Google Scholar] [CrossRef]
- Panetta, F.; Gooden, B. Managing for biodiversity: Impact and action thresholds for invasive plants in natural ecosystems. NeoBiota 2017, 34, 53–66. [Google Scholar] [CrossRef] [Green Version]
- Evju, M.; Sverdrup-Thygeson, A. Spatial configuration matters: A test of the habitat amount hypothesis for plants in calcareous grasslands. Landsc. Ecol. 2016, 31, 1891–1902. [Google Scholar] [CrossRef]
- Haddad, N.M.; Gonzalez, A.; Brudvig, L.A.; Burt, M.A.; Levey, D.J.; Damschen, E.I. Experimental evidence does not support the Habitat Amount Hypothesis. Ecography (Cop.) 2017, 40, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Rybicki, J.; Hanski, I. Species-area relationships and extinctions caused by habitat loss and fragmentation. Ecol. Lett. 2013, 16, 27–38. [Google Scholar] [CrossRef]
- Coudrain, V.; Schüepp, C.; Herzog, F.; Albrecht, M.; Entling, M.H. Habitat amount modulates the effect of patch isolation on host-parasitoid interactions. Front. Environ. Sci. 2014, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.F.; Whigham, D.F. Effects of forest fragmentation on breeding bird communities in Maryland, USA. Biol. Conserv. 1984, 28, 287–324. [Google Scholar] [CrossRef]
- Pivello, V.R.; Petenon, D.; De Jesus, F.M.; Meirelles, S.T.; Vidal, M.M.; Alonso, R.D.A.S.; Franco, G.A.D.C.; Metzger, J.P. Chuva de sementes em fragmentos de Floresta Atlântica (São Paulo, SP, Brasil), sob diferentes situações de conectividade, estrutura florestal e proximidade da borda. Acta Bot. Brasilica 2006, 20, 845–859. [Google Scholar] [CrossRef] [Green Version]
- Gurrutxaga, M.; Rubio, L.; Saura, S. Key connectors in protected forest area networks and the impact of highways: A transnational case study from the Cantabrian Range to the Western Alps (SW Europe). Landsc. Urban Plan. 2011, 101, 310–320. [Google Scholar] [CrossRef]
- McRae, B.H.; Dickson, B.G.; Keitt, T.H.; Shah, V.B. Using circuit theory to model connectivity in ecology, evolution, and conservation. Ecology 2008, 89, 2712–2724. [Google Scholar] [CrossRef] [PubMed]
- Dickson, B.G.; Albano, C.M.; Anantharaman, R.; Beier, P.; Fargione, J.; Graves, T.A.; Gray, M.E.; Hall, K.R.; Lawler, J.J.; Leonard, P.B.; et al. Circuit-theory applications to connectivity science and conservation. Conserv. Biol. 2018, 33, 239–249. [Google Scholar] [CrossRef]
- McRae, B.H. Isolation By Resistance. Evolution (N. Y). 2006, 60, 1551. [Google Scholar] [CrossRef]
- McRae, B.H.; Beier, P. Circuit theory predicts gene flow in plant and animal populations. Proc. Natl. Acad. Sci. USA 2007, 104, 19885–19890. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, D.A.; Banks, S.C.; Barton, P.S.; Lindenmayer, D.B.; Smith, A.L. Conceptual domain of the matrix in fragmented landscapes. Trends Ecol. Evol. 2013, 28, 605–613. [Google Scholar] [CrossRef]
- Pascual-Hortal, L.; Saura, S. Comparison and development of new graph-based landscape connectivity indices: Towards the priorization of habitat patches and corridors for conservation. Landsc. Ecol. 2006, 21, 959–967. [Google Scholar] [CrossRef]
- Mix, C.; Arens, P.F.P.; Rengelink, R.; Smulders, M.J.M.; Van Groenendael, J.M.; Ouborg, N.J. Regional gene flow and population structure of the wind-dispersed plant species Hypochaeris radicata (Asteraceae) in an agricultural landscape. Mol. Ecol. 2006. [Google Scholar] [CrossRef]
- Donatti, C.I.; Galetti, M.; Pizo, M.A.; Guimarães, P.R.; Jordano, P. Living in the land of ghosts: Fruit traits and the importance of large mammals as seed dispersers in the Pantanal, Brazil. In Seed Dispersal Theory Its Appl. a Chang. World; CAB International: Wallinford, UK, 2009; pp. 104–123. [Google Scholar] [CrossRef] [Green Version]
- Croteau, E.K. Causes and Consequences of Dispersal in Plants and Animals. Nat. Educ. Knowl. 2010, 3, 12. [Google Scholar]
- ICMBio. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção: Volume II-Mamíferos. In Livro Vermelho da Fauna Bras. Ameaçada Extinção; ICMBio: Jaú National Park: Novo Airão - AM, Brazil, 2018; p. 622. Available online: https://www.icmbio.gov.br/portal/images/stories/comunicacao/publicacoes/publicacoes-diversas/livro_vermelho_2018_vol2.pdf (accessed on 25 January 2020).
- Monteiro, C.H.B.; do Prado, B.H.; Dias, A.C. Plano de Manejo da Estação Ecológica de Angatuba; Páginas & Letras-Editora e Gráfica Ltda: Belenzinho, São Paulo, Brazil, 2009; ISBN 978-85-8191-044-0. Available online: http://arquivo.ambiente.sp.gov.br/consema/2011/11/oficio_consema_2009_056/Plano_de_Manejo_Estacao_Ecologica_Angatuba.pdf (accessed on 25 January 2020).
- Gheler-Costa, C.; Verdade, L.M.; Lyra-Jorge, M.C.; Martin, P.S.; Calaboni, A.; Pinto, A.P.R. Mamíferos da região de Angatuba; Edição do autor.: São Paulo, Brazil, 2018. [Google Scholar]
- Passos, F.C.; Silva, W.R.; Pedro, W.A.; Bonin, M.R. Frugivoria em morcegos (Mammalia, Chiroptera) no Parque Estadual Intervales, sudeste do Brasil. Rev. Bras. Zool. 2003, 20, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Bueno, A.D.A.; Motta-Junior, J.C. Food habits of two syntopic canids, the maned wolf (Chrysocyon brachyurus) and the crab-eating fox (Cerdocyon thous), in southeastern Brazil. Rev. Chil. História Nat. 2004, 77, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Gottsberger, G.; Silberbauer-Gottsberger, I. Life in the Cerrado: A South American Tropical Seasonal Ecosystem; Reta Verlag: Berlin, Germany, 2006; ISBN 9783000179280. [Google Scholar]
- Klimešová, J.; Klimeš, L. Bud banks and their role in vegetative regeneration-A literature review and proposal for simple classification and assessment. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 115–129. [Google Scholar] [CrossRef]
- Ferreira, M.C.; Walter, B.M.T.; Vieira, D.L.M. Topsoil translocation for Brazilian savanna restoration: Propagation of herbs, shrubs, and trees. Restor. Ecol. 2015, 23, 723–728. [Google Scholar] [CrossRef]
- Cava, M.G.D.B.; Isernhagen, I.; de Mendonça, A.H.; Durigan, G. Comparação de técnicas para restauração da vegetação lenhosa de Cerrado em pastagens abandonadas. Hoehnea 2016, 43, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Hardt, E.; Pereira-Silva, E.F.L.; Dos Santos, R.F.; Tamashiro, J.Y.; Ragazzi, S.; Lins, D.B.D.S. The influence of natural and anthropogenic landscapes on edge effects. Landsc. Urban Plan. 2013, 120, 59–69. [Google Scholar] [CrossRef]
- Ries, L.; Sisk, T.D. A predictive model of edge effects. Ecology 2004, 85, 2917–2926. [Google Scholar] [CrossRef] [Green Version]
- Schneider-Maunoury, L.; Lefebvre, V.; Ewers, R.M.; Medina-Rangel, G.F.; Peres, C.A.; Somarriba, E.; Urbina-Cardona, N.; Pfeifer, M. Abundance signals of amphibians and reptiles indicate strong edge effects in Neotropical fragmented forest landscapes. Biol. Conserv. 2016, 200, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Levey, D.J.; Bolker, B.M.; Tewksbury, J.J.; Sargent, S.; Haddad, N.M. Effects of landscape corridors on seed dispersal by birds. Science (80-. ) 2005, 309, 146–148. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Hernández, C.G.; Vergara, P.M.; Saura, S.; Hernández, J. Do corridors promote connectivity for bird-dispersed trees? The case of Persea lingue in Chilean fragmented landscapes. Landsc. Ecol. 2014, 30, 77–90. [Google Scholar] [CrossRef]
- Gilbert-Norton, L.; Wilson, R.; Stevens, J.R.; Beard, K.H. A Meta-Analytic Review of Corridor Effectiveness. Conserv. Biol. 2010, 24, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Doerr, V.; Doerr, E.; Davies, M. Does structural connectivity facilitate dispersal of native species in Australia’s fragmented terrestrial landscapes. CEE Rev. 2010, 7, 08-007. [Google Scholar] [CrossRef] [Green Version]
- Zanirato, R.B.D.C.A.; de Pádua Melo, C.E.S. Sistema reprodutivo de Anadenanthera peregrina (L.) Speg e Vochysia haenkiana (Spreng.) Mart. em fragmento de cerrado na Chapada dos Guimarães-MT. Ciência Rural 2003, 33, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Bonjorne de Almeida, L.; Galetti, M. Seed dispersal and spatial distribution of Attalea geraensis (Arecaceae) in two remnants of Cerrado in Southeastern Brazil. Acta Oecologica 2007, 32, 180–187. [Google Scholar] [CrossRef]
- Fichera, C.R.; Laudari, L.; Modica, G. Application, validation and comparison in different geographical contexts of an integrated model for the design of ecological networks. J. Agric. Eng. 2015, 46, 52. [Google Scholar] [CrossRef] [Green Version]
- de la Fuente, B.; Mateo-Sánchez, M.C.; Rodríguez, G.; Gastón, A.; Pérez de Ayala, R.; Colomina-Pérez, D.; Melero, M.; Saura, S. Natura 2000 sites, public forests and riparian corridors: The connectivity backbone of forest green infrastructure. Land Use Policy 2018, 75, 429–441. [Google Scholar] [CrossRef]
- D’Eon, R.G.; Glenn, S.M.; Parfitt, I.; Fortin, M.-J. Landscape Connectivity as a Function of Scale and Organism Vagility in a Real Forested Landscape. Conserv. Ecol. 2002, 6, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Borda-Niño, M.; Hernández-Muciño, D.; Ceccon, E. Planning restoration in human-modified landscapes: New insights linking different scales. Appl. Geogr. 2017, 83, 118–129. [Google Scholar] [CrossRef]
- Molin, P.G.; Chazdon, R.; Frosini de Barros Ferraz, S.; Brancalion, P.H.S. A landscape approach for cost-effective large-scale forest restoration. J. Appl. Ecol. 2018, 55, 2767–2778. [Google Scholar] [CrossRef]
- Herrera, L.P.; Sabatino, M.C.; Jaimes, F.R.; Saura, S. Landscape connectivity and the role of small habitat patches as stepping stones: An assessment of the grassland biome in South America. Biodivers. Conserv. 2017, 26, 3465–3479. [Google Scholar] [CrossRef]
- Yeong, K.L.; Reynolds, G.; Hill, J.K. Enrichment planting to improve habitat quality and conservation value of tropical rainforest fragments. Biodivers. Conserv. 2016, 25, 957–973. [Google Scholar] [CrossRef]
- Tjørve, E. How to resolve the SLOSS debate: Lessons from species-diversity models. J. Theor. Biol. 2010, 264, 604–612. [Google Scholar] [CrossRef]
- Diniz, M.F.; Machado, R.B.; Bispo, A.A.; Júnior, P.D.M. Can we face different types of storms under the same umbrella? Efficiency and consistency of connectivity umbrellas across different patchy landscape patterns. Landsc. Ecol. 2018, 33, 1911–1923. [Google Scholar] [CrossRef]
- Miller-Rushing, A.J.; Primack, R.B.; Devictor, V.; Corlett, R.T.; Cumming, G.S.; Loyola, R.; Maas, B.; Pejchar, L. How does habitat fragmentation affect biodiversity? A controversial question at the core of conservation biology. Biol. Conserv. 2019, 232, 271–273. [Google Scholar] [CrossRef]
- Adams, V.M.; Mills, M.; Weeks, R.; Segan, D.B.; Pressey, R.L.; Gurney, G.G.; Groves, C.; Davis, F.W.; Álvarez-Romero, J.G. Implementation strategies for systematic conservation planning. Ambio 2019, 48, 139–152. [Google Scholar] [CrossRef]
- Botts, E.A.; Pence, G.; Holness, S.; Sink, K.; Skowno, A.; Driver, A.; Harris, L.R.; Desmet, P.; Escott, B.; Lötter, M.; et al. Practical actions for applied systematic conservation planning. Conserv. Biol. 2019, 33, 1235–1246. [Google Scholar] [CrossRef]
- Margules, C.R.; Pressey, R.L. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef]
- Alfaya, P.; de Pablo, C.T.L.; de Agar, P.M.; Alonso, G. Assessing the influence of ecological interaction patterns among habitat types on species distribution: Studying the Iberian lynx (Lynx pardinus Temminck 1827) in central Spain. Landsc. Ecol. 2020, 35, 1923–1944. [Google Scholar] [CrossRef]
- Hansen, A.I.; Di Castri, F. Landscape Boundaries: Consequences for Biological Diversity and Ecological Flows; Springer: New York, NY, USA, 1992. [Google Scholar]
- Mander, U.; Kull, A.; Kuusemets, V. Nutrient flows and land use change in a rural catchment: A modeling approach. Landsc. Ecol. 2000, 15, 187–199. [Google Scholar] [CrossRef]
- Hood, E.W.; Williams, M.W.; Caine, N. Landscape Controls on Organic and Inorganic Nitrogen Leaching across an Alpine/Subalpine Ecotone, Green Lakes Valley, Colorado Front Range. Ecosystems 2003, 6, 0031–0045. [Google Scholar] [CrossRef]
- Ecosystem Function in Heterogeneous Landscapes; Lovett, G.M.; Turner, M.G.; Jones, C.G.; Weathers, K.C. (Eds.) Springer New York: New York, NY, USA, 2005; ISBN 978-0-387-24089-3. [Google Scholar]
- López-Barrera, F.; Armesto, J.J.; Williams-Linera, G.; Smith-Ramírez, C.; Manson, R.H. Fragmentation and edge effects on plant-animal interactions, ecological processes and biodiversity. In Biodiversity Loss and Conservation in Fragmented Forest Landscapes: The Forests of Montane Mexico and Temperate South America; CABI: Wallingford, UK, 2007; pp. 69–101. [Google Scholar]
- Bastian, O.; Grunewald, K.; Khoroshev, A.V. The significance of geosystem and landscape concepts for the assessment of ecosystem services: Exemplified in a case study in Russia. Landsc. Ecol. 2015, 30, 1145–1164. [Google Scholar] [CrossRef]
- Martín de Agar, P.; Ortega, M.; de Pablo, C.L. A procedure of landscape services assessment based on mosaics of patches and boundaries. J. Environ. Manage. 2016, 180, 214–227. [Google Scholar] [CrossRef] [PubMed]
- Marull, J.; Herrando, S.; Brotons, L.; Melero, Y.; Pino, J.; Cattaneo, C.; Pons, M.; Llobet, J.; Tello, E. Building on Margalef: Testing the links between landscape structure, energy and information flows driven by farming and biodiversity. Sci. Total Environ. 2019, 674, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.M. Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography (Cop.) 2003, 26, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Gómez, J.M.; Puerta-Piñero, C.; Schupp, E.W. Effectiveness of rodents as local seed dispersers of Holm oaks. Oecologia 2008, 155, 529–537. [Google Scholar] [CrossRef]
| Metric | Equation | Description of Measures | |
|---|---|---|---|
| Forest resource availability | Optimal resource availability—ORA (varies from 0 to 1) Refs. [39,42] with adaptations | (1) FShi for edge of 60 m [42]; CSi varies from 1 to 4: 1 (≤1 ha), 2 (1–10 ha), 3 (10–50 ha), and 4 (≥50 ha); * CFi varies from 1 to 3: 1 (AF in early stages), 2 (AF at intermediate/advanced stages), and 3 (CER) | Potential optimal resource availability of forest in the landscape based on the shape of the patchesi (FShi), size coefficient (CSi), and physiognomic importance (CFi) in relation to the best resource conditions with maximum potential (CSmáx = 4 and CFmáx = 3) for the total landscape area (TA) (in ArcGIS®) |
| Forest fragmentation | Number of patches—NP (varies from 0 to ∞) | (2) | Subdivision or fragmentation of the landscape given by the quantity (n) of forest class patchesi (in Fragstats®) |
| Perimeter/area ratio—PARA (>0–∞) | (3) | Shape complexity of the forest class is expressed by the mean of the perimeter (Pi) by area (Ai) ratio in relation to the number of patchesi (ni). High values: more uneven or elongated shapes; low values: more uniform shapes (in Fragstats®) | |
| Euclidean distance from nearest neighbor—ENN_MN (>0–∞) | (4) | Isolation of the forest class calculated using the sum of the distance in meters (Hij) from the edge of the patchi to the nearest edge of the patchj divided by the number of patches (n) (in Fragstats®) | |
| Landscape heterogeneity and aggregation | Shannon evenness index (varies from 0 to 1) | (5) ln = Napierian logarithm | Landscape heterogeneity calculated using the ratio between the mean of the landscape occupied by the type of classi (Pi) ratio and the number of types of patches (classes) (m) present in the landscape. Close to 0: uneven distribution of classes that may indicate a dominance of a class in a homogeneous landscape. 1: class distribution is perfectly uniform, and the landscape is heterogeneous (in Fragstats®) |
| Contagion index—CONTAG (varies from 0 to 100%) | (6) ln = Napierian logarithm | Landscape aggregation is expressed as a percentage based on the mean product of the proportion of the landscape occupied by classi (Pi) and the number of joinings between classesik (gik) in relation to the number of landscape classes (m). 0: classes totally interspersed and disaggregated; 100: fully aggregated classes (in Fragstats®) | |
| Functional landscape connectivity | Connectivity index—CONNECT (varies from 0 to 100) | (7) | Functional connections at species-specific distance thresholds (m) between patchesjk (Cijk) of the same classi in relation to the number of patches (n) in the landscape. 0: null functional connectivity; 100: landscape with fully connected patches (in Fragstats®) |
| Integral index of connectivity—IIC (varies from 0 to 1) | (8) | Functional landscape connectivity based on the number of patches (n) in the landscape, the quality of the patchesij (ai and aj), the number of functional connections at species-specific distance thresholds between patchesij (nlij) divided by the total landscape area (AL) multiplied by 3, since the habitat patches had this maximum weight. 0: null functional connectivity; 1: hypothetical landscape completely covered by habitat (in Conefor®) | |
| Dispersal Capacity | Dispersal Syndrome | Indicator Species | N | References | |
|---|---|---|---|---|---|
| Category | Maximum Distance (m) | ||||
| Short | 5 | autochory | Temperate climate species | variable | [47] |
| Medium | 246 | mammaliochory and ornithochory | Copaifera langsdorffii (Fabaceae) | 340 | [43] |
| Medium–long | 1800 | mammaliochory | Annona crassiflora (Annonaceae) | 40 | [45] |
| Long | 8091 | mammaliochory and ornithochory | Hymenaea stigonocarpa (Fabaceae: Caesalpinioideae) | 450 | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Issii, T.M.; Pereira-Silva, E.F.L.; López de Pablo, C.T.; Ferreira dos Santos, R.; Hardt, E. Is There an Equivalence between Measures of Landscape Structural and Functional Connectivity for Plants in Conservation Assessments of the Cerrado? Land 2020, 9, 459. https://doi.org/10.3390/land9110459
Issii TM, Pereira-Silva EFL, López de Pablo CT, Ferreira dos Santos R, Hardt E. Is There an Equivalence between Measures of Landscape Structural and Functional Connectivity for Plants in Conservation Assessments of the Cerrado? Land. 2020; 9(11):459. https://doi.org/10.3390/land9110459
Chicago/Turabian StyleIssii, Thais Martins, Erico Fernando Lopes Pereira-Silva, Carlos Tomás López de Pablo, Rozely Ferreira dos Santos, and Elisa Hardt. 2020. "Is There an Equivalence between Measures of Landscape Structural and Functional Connectivity for Plants in Conservation Assessments of the Cerrado?" Land 9, no. 11: 459. https://doi.org/10.3390/land9110459
APA StyleIssii, T. M., Pereira-Silva, E. F. L., López de Pablo, C. T., Ferreira dos Santos, R., & Hardt, E. (2020). Is There an Equivalence between Measures of Landscape Structural and Functional Connectivity for Plants in Conservation Assessments of the Cerrado? Land, 9(11), 459. https://doi.org/10.3390/land9110459