Quantifying Climate-Wise Connectivity across a Topographically Diverse Landscape


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
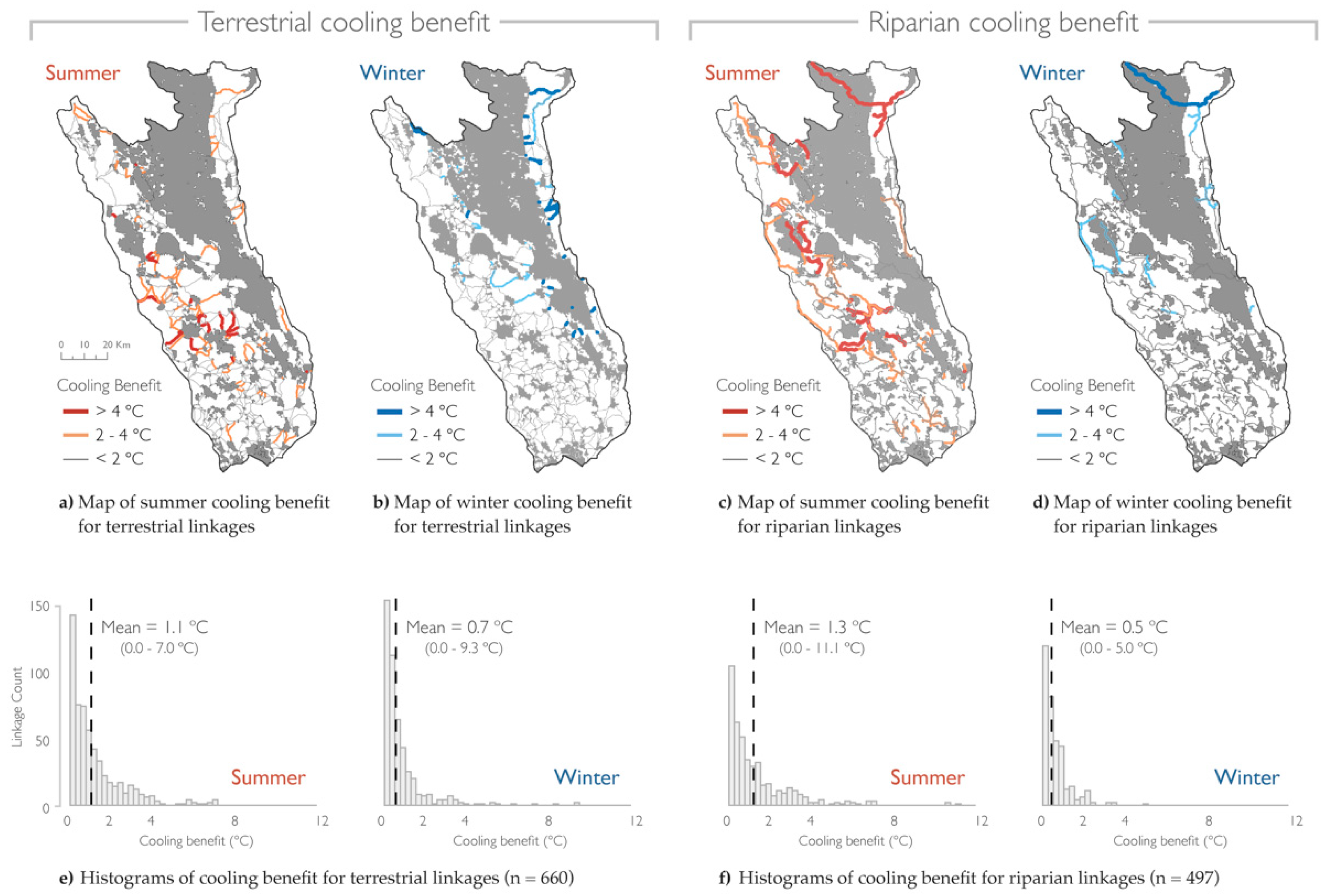{kind=link}
1. Introduction
2. Materials and Methods
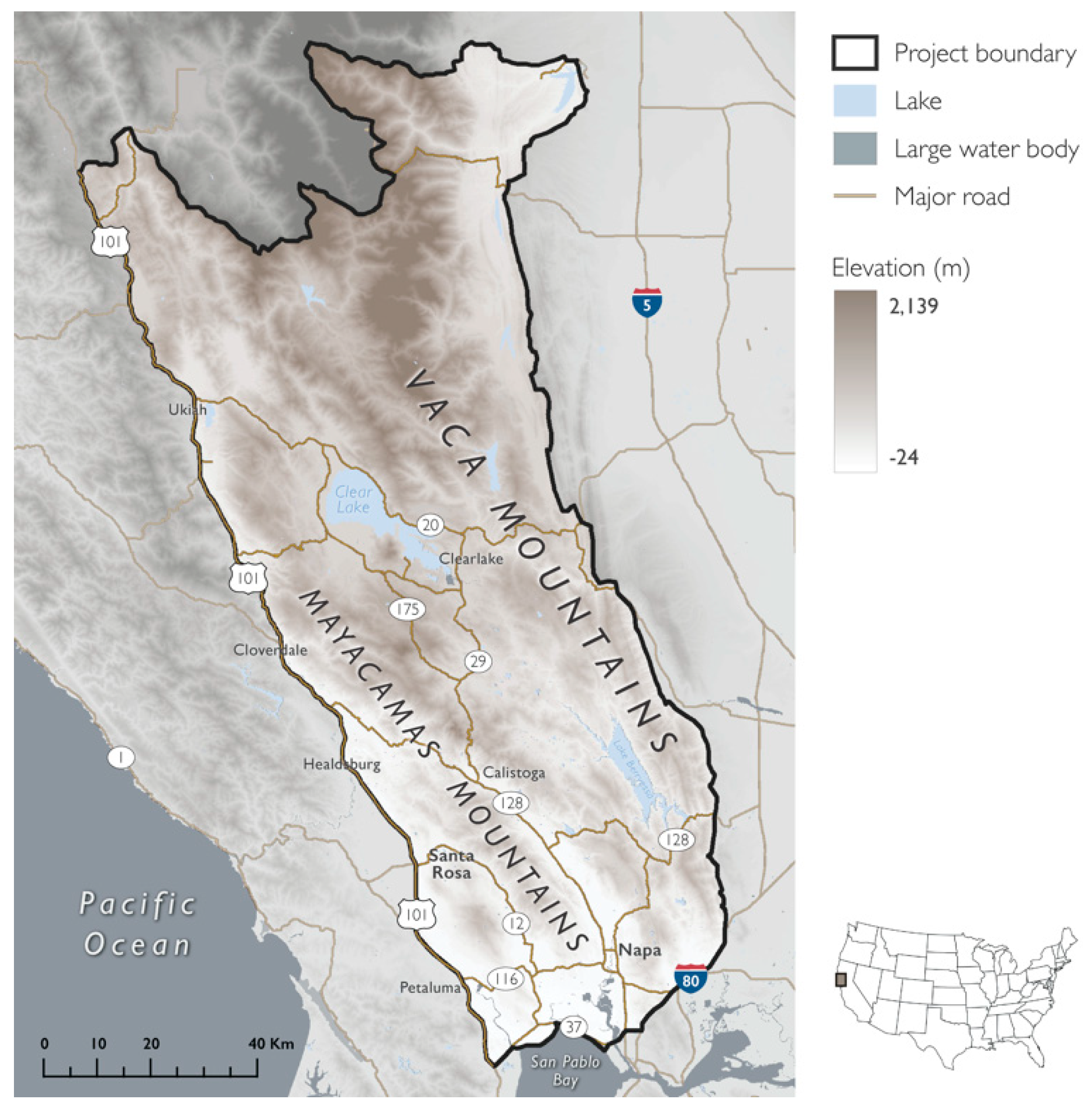
2.1. Project Area
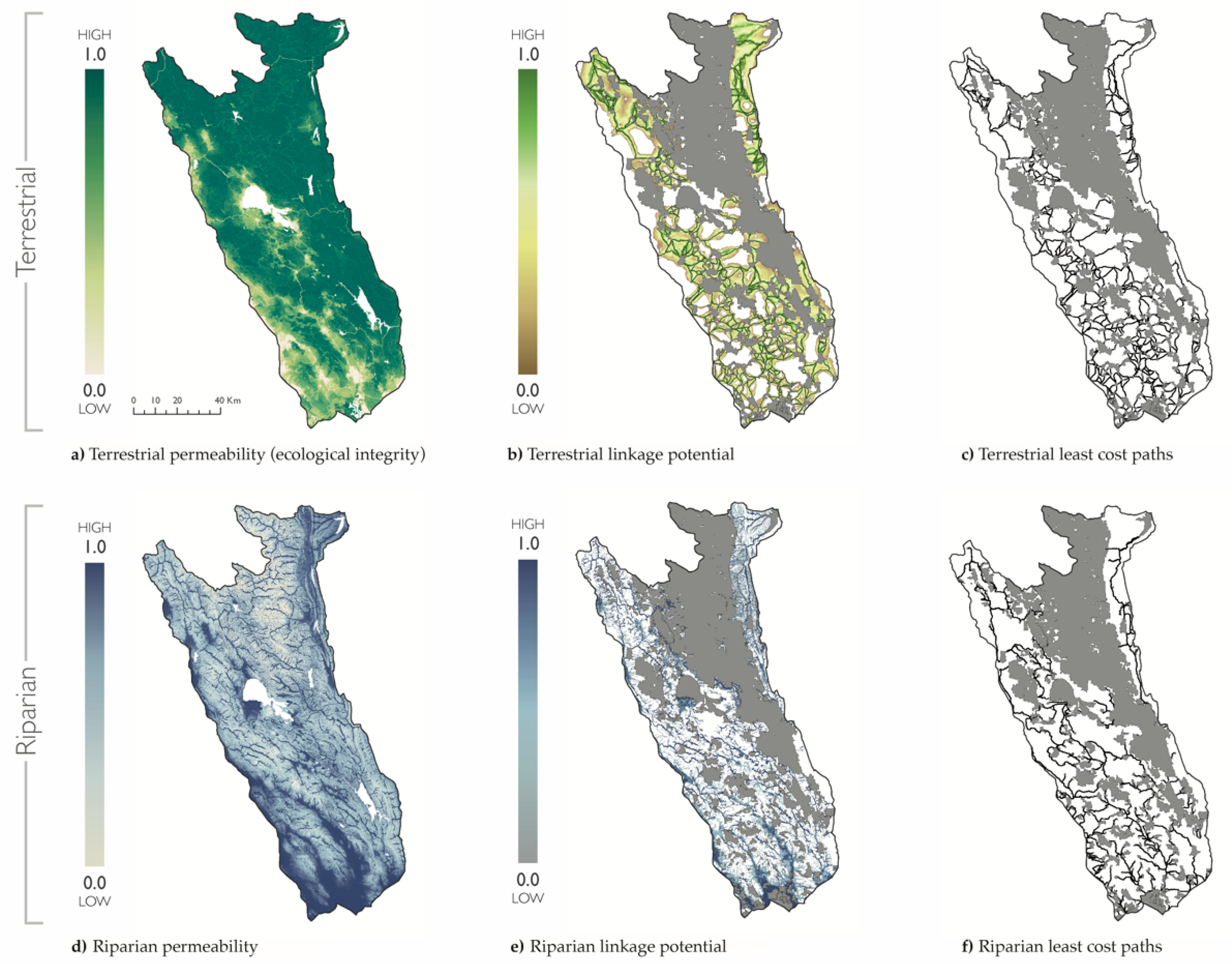
2.2. Terrestrial and Riparian Connectivity
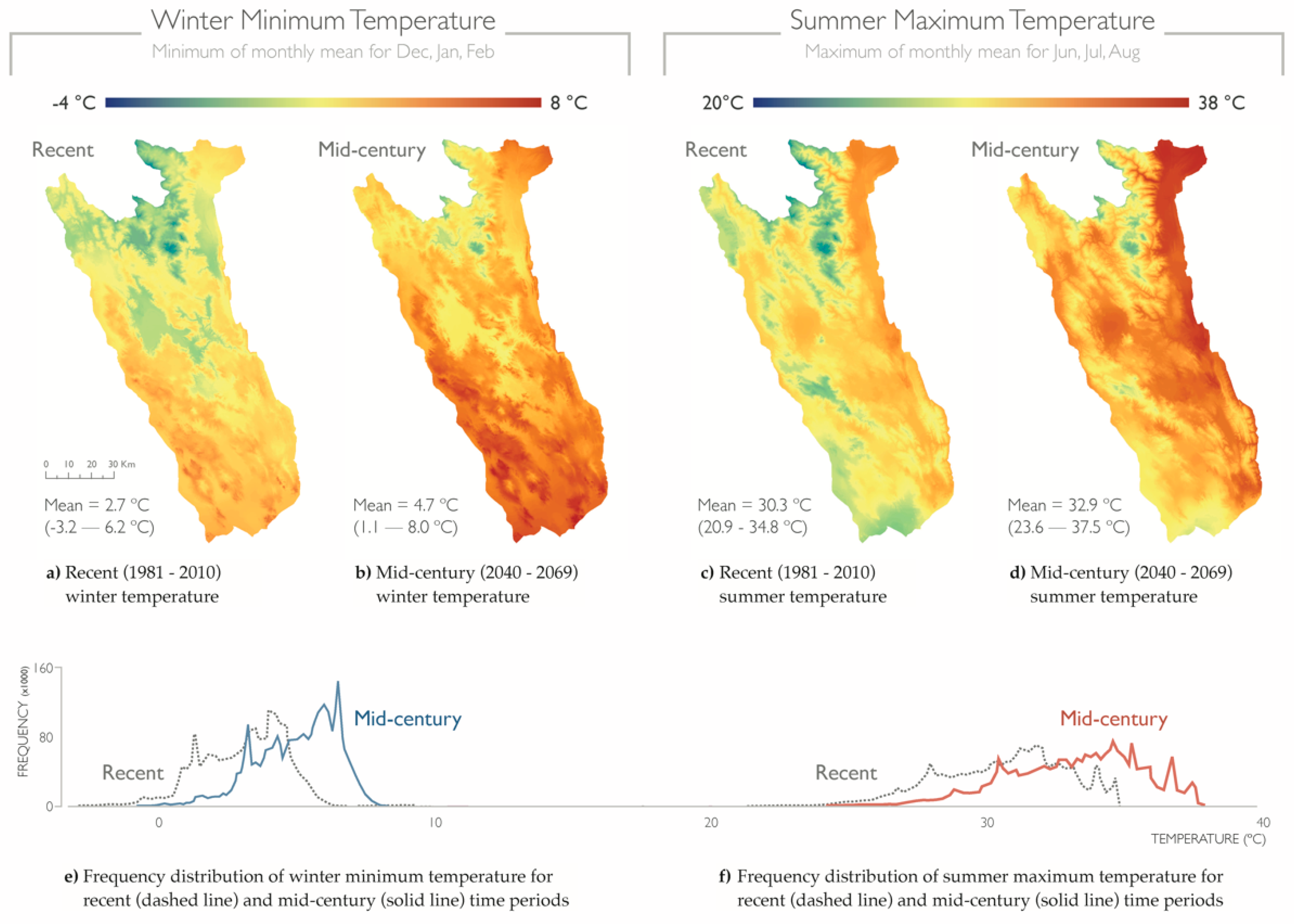
2.3. Cooling Benefit for Seasonal Temperature Extremes
3. Results
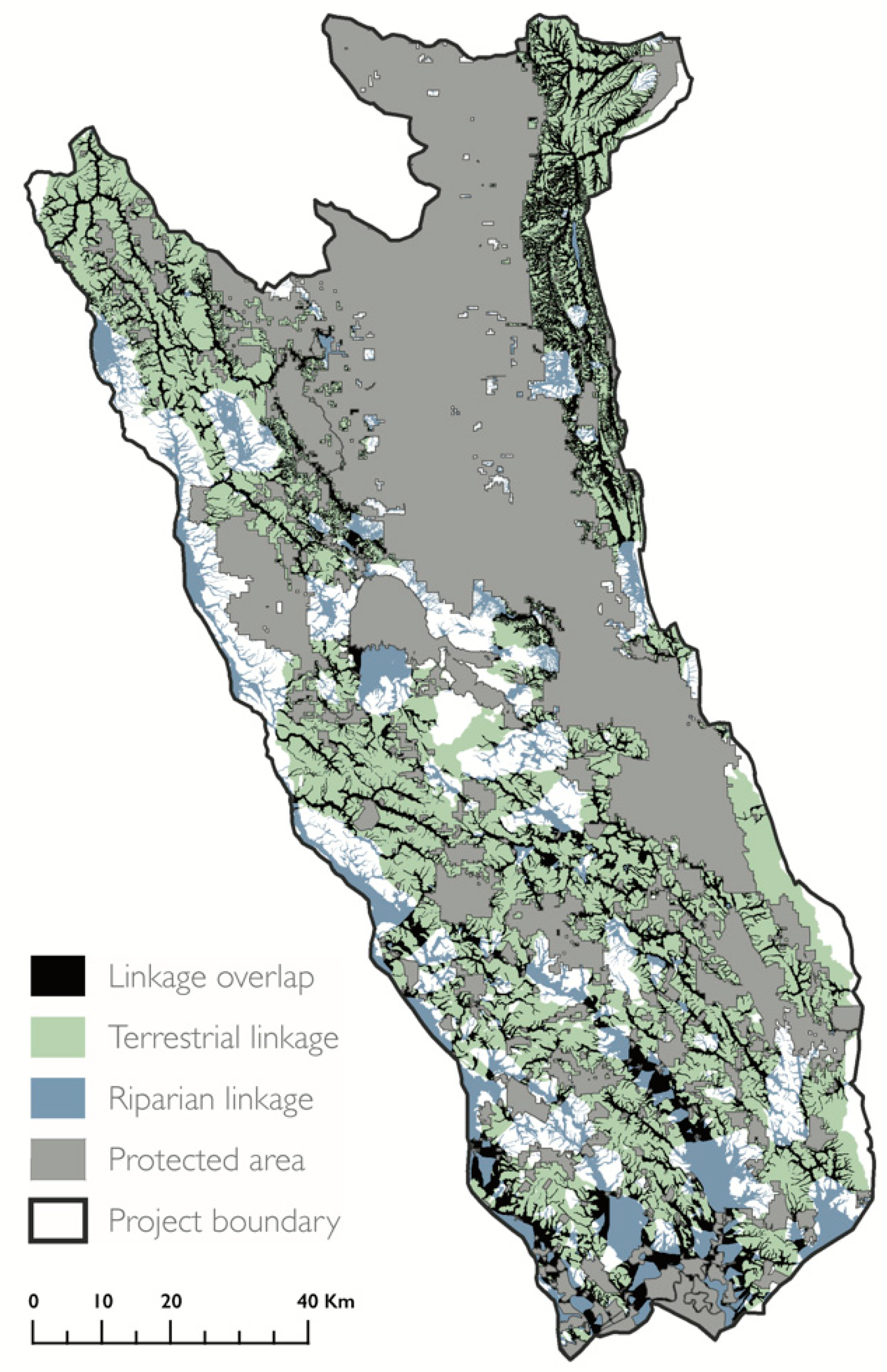
3.1. Terrestrial and Riparian Connectivity
3.2. Cooling Benefit for Seasonal Temperature Extremes
4. Discussion
4.1. Terrestrial and Riparian Connectivity
4.2. Cooling Benefit for Seasonal Temperature Extremes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. (Supplement) Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355. [Google Scholar] [CrossRef]
- Berg, M.P.; Toby Kiers, E.; Driessen, G.; van der Heijden, M.; Kooi, B.W.; Kuenen, F.; Liefting, M.; Verhoef, H.A.; Ellers, J. Adapt or disperse: Understanding species persistence in a changing world. Glob. Chang. Biol. 2010, 16, 587–598. [Google Scholar] [CrossRef]
- Krosby, M.; Tewksbury, J.; Haddad, N.; Hoekstra, J. Ecological Connectivity for a Changing Climate. Conserv. Biol. 2010, 24, 1686–1689. [Google Scholar] [CrossRef] [PubMed]
- Burrows, M.T.; Schoeman, D.S.; Richardson, A.J.; Molinos, J.G.; Hoffmann, A.; Buckley, L.B.; Moore, P.J.; Brown, C.J.; Bruno, J.F.; Duarte, C.M.; et al. Geographical limits to species-range shifts are suggested by climate velocity. Nature 2014, 507, 492–495. [Google Scholar] [CrossRef]
- Chen, I.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Fox, R.; Thomas, C.D. The distributions of a wide range of taxonomic groups are expanding polewards. Glob. Chang. Biol. 2006, 12, 450–455. [Google Scholar] [CrossRef]
- Fei, S.; Desprez, J.M.; Potter, K.M.; Jo, I.; Knott, J.A.; Oswalt, C.M. Divergence of species responses to climate change. Sci. Adv. 2017, 3, e1603055. [Google Scholar] [CrossRef]
- Moritz, C.; Patton, J.L.; Conroy, C.J.; Parra, J.L.; White, G.C.; Beissinger, S.R. Impact of a Century of Climate Change on Small-Mammal Communities in Yosemite National Park, USA. Science 2008, 322, 261–264. [Google Scholar] [CrossRef]
- Tingley, M.W.; Koo, M.S.; Moritz, C.; Rush, A.C.; Beissinger, S.R. The push and pull of climate change causes heterogeneous shifts in avian elevational ranges. Glob. Chang. Biol. 2012, 18, 3279–3290. [Google Scholar] [CrossRef]
- Roelke, M.E.; Martenson, J.S.; O’Brien, S.J.; Jablonski, D.; Stanley, S.M.; Myers, N.; Elliot, D.K.; Centre, T.I.C.M.; Service, U.S.F.W.; Service, U.S.F.W.; et al. The consequences of demographic reduction and genetic depletion in the endangered Florida panther. Curr. Biol. 1993, 3, 340–350. [Google Scholar] [CrossRef]
- Fahrig, L.; Merriam, G. Conservation of fragmented populations. Conserv. Biol. 1994, 8, 50–59. [Google Scholar] [CrossRef]
- Epps, C.W.; Palsbøll, P.J.; Wehausen, J.D.; Roderick, G.K.; Ramey, R.R.; McCullough, D.R. Highways block gene flow and cause a rapid decline in genetic diversity of desert bighorn sheep. Ecol. Lett. 2005, 8, 1029–1038. [Google Scholar] [CrossRef]
- Charlesworth, D.; Charlesworth, D.; Willis, J.H.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef]
- Hannah, L. Climate Change, Connectivity, and Conservation Success. Conserv. Biol. 2011, 25, 1139–1142. [Google Scholar] [CrossRef] [PubMed]
- Hilty, J.A.; Lidicker, W.Z.; Merenlender, A. Corridor Ecology: The Science and Practice of Linking Landscapes for Biodiversity Conservation; Island Press: Washington, DC, USA, 2006; ISBN 1597265934. [Google Scholar]
- Heller, N.; Zavaleta, E. Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biol. Conserv. 2009, 142, 14–32. [Google Scholar] [CrossRef]
- Tilman, D.; Clark, M.; Williams, D.R.; Kimmel, K.; Polasky, S.; Packer, C. Future threats to biodiversity and pathways to their prevention. Nature 2017, 546, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Jayadevan, A.; Nayak, R.; Karanth, K.K.; Krishnaswamy, J.; DeFries, R.; Karanth, K.U.; Vaidyanathan, S. Navigating paved paradise: Evaluating landscape permeability to movement for large mammals in two conservation priority landscapes in India. Biol. Conserv. 2020, 247, 108613. [Google Scholar] [CrossRef]
- Hilty, J.A.; Keeley, A.T.H.; Lidicker, W.Z.; Merenlender, A.M. Corridor Ecology: Linking Landscapes for Biodiversity Conservation and Climate Adaptation, 2nd ed.; Island Press: Washington, DC, USA, 2019. [Google Scholar]
- Keeley, A.; Ackerly, D.; Cameron, D.; Heller, N.; Huber, P.; Schloss, C.; Thorne, J.; Merenlender, A. New concepts, models, and assessments of climate-wise connectivity. Environ. Res. Lett. 2018, 13. [Google Scholar] [CrossRef]
- Phillips, S.J.; Williams, P.; Midgley, G.; Archer, A. Optimizing Dispersal Corridors for the Cape Proteaceae Using Network Flow. Ecol. Appl. 2008, 18, 1200–1211. [Google Scholar] [CrossRef]
- Lawler, J.J.; Ruesch, A.S.; Olden, J.D.; McRae, B.H. Projected climate-driven faunal movement routes. Ecol. Lett. 2013, 16, 1014–1022. [Google Scholar] [CrossRef]
- Alagador, D.; Cerdeira, J.O.; Araújo, M.B. Climate change, species range shifts and dispersal corridors: An evaluation of spatial conservation models. Methods Ecol. Evol. 2016, 7, 853–866. [Google Scholar] [CrossRef]
- Rose, N.-A.; Burton, P.J. Using bioclimatic envelopes to identify temporal corridors in support of conservation planning in a changing climate. For. Ecol. Manag. 2009, 258, S64–S74. [Google Scholar] [CrossRef]
- Pellatt, M.G.; Goring, S.J.; Bodtker, K.M.; Cannon, A.J. Using a down-scaled bioclimate envelope model to determine long-term temporal connectivity of garry oak (Quercus garryana) habitat in Western North America: Implications for protected area planning. Environ. Manag. 2012, 49, 802–815. [Google Scholar] [CrossRef] [PubMed]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The velocity of climate change. Nature 2009, 462, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- McKelvey, K.S.; Copeland, J.P.; Schwartz, M.K.; Littell, J.S.; Aubry, K.B.; Squires, J.R.; Parks, S.A.; Elsner, M.M.; Mauger, G.S. Climate change predicted to shift wolverine distributions, connectivity, and dispersal corridors. Ecol. Appl. 2011, 21, 2882–2897. [Google Scholar] [CrossRef]
- Howard, T.; Schlesinger, M.D. Wildlife habitat connectivity in the changing climate of New York’s Hudson Valley. Available online: https://nyaspubs.onlinelibrary.wiley.com/doi/10.1111/nyas.12172 (accessed on 25 September 2020).
- Drake, J.C.; Griffis-Kyle, K.; McIntyre, N.E. Using nested connectivity models to resolve management conflicts of isolated water networks in the Sonoran Desert. Ecosphere 2017, 8. [Google Scholar] [CrossRef]
- Littlefield, C.E.; McRae, B.H.; Michalak, J.; Lawler, J.J.; Carroll, C. Connecting today’s climates to future analogs to facilitate species movement under climate change. Conserv. Biol. 2017, 1–31. [Google Scholar] [CrossRef]
- Pressey, R.L. Conservation Planning for a Changing Climate. 2007. Available online: https://researchonline.jcu.edu.au/25280/1/15948.pdf (accessed on 25 September 2020).
- Noss, R. Beyond Kyoto: Forest management in a time of rapid climate change. Conserv. Biol. 2001, 15, 578–590. [Google Scholar] [CrossRef]
- Rouget, M.; Cowling, R.M.; Lombard, A.T.; Knight, A.T.; Kerley, G.I.H. Designing Large-Scale Conservation Corridors for Pattern and Process. Conserv. Biol. 2006, 20, 549–561. [Google Scholar] [CrossRef]
- Morelli, T.L.; Daly, C.; Dobrowski, S.Z.; Dulen, D.M.; Ebersole, J.L.; Jackson, S.T.; Lundquist, J.D.; Millar, C.I.; Maher, S.P.; Monahan, W.B.; et al. Managing climate change refugia for climate adaptation. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Heller, N.E.; Kreitler, J.; Ackerly, D.D.; Weiss, S.B.; Recinos, A.; Branciforte, R.; Flint, L.E.; Flint, A.L.; Micheli, E. Targeting climate diversity in conservation planning to build resilience to climate change. Ecosphere 2015, 6, 65. [Google Scholar] [CrossRef]
- Keppel, G.; Van Niel, K.P.; Wardell-Johnson, G.W.; Yates, C.J.; Byrne, M.; Mucina, L.; Schut, A.G.T.; Hopper, S.D.; Franklin, S.E. Refugia: Identifying and understanding safe havens for biodiversity under climate change. Glob. Ecol. Biogeogr. 2012, 21, 393–404. [Google Scholar] [CrossRef]
- Anderson, M.G.; Barnett, A.; Clark, M.; Sheldon, A.O.; Prince, J.; Vickery, B. Resilient and Connected Landscapes for Terrestrial Conservation. Available online: https://easterndivision.s3.amazonaws.com/Resilient_and_Connected_Landscapes_For_Terrestial_Conservation.pdf (accessed on 25 September 2020).
- Fremier, A.K.; Kiparsky, M.; Gmur, S.; Aycrigg, J.; Craig, R.K.; Svancara, L.K.; Goble, D.D.; Cosens, B.; Davis, F.W.; Scott, J.M. A riparian conservation network for ecological resilience. Biol. Conserv. 2015, 191, 29–37. [Google Scholar] [CrossRef]
- Capon, S.; Nally, R. Mac; Davies, P.M. Riparian Ecosystems in the 21st Century: Hotspots for Climate Change Adaptation? Environmental Water Needs for the Fitzroy River View project An Integrated Approach for Assessing Vulnerability and Potential Adaptation Options for a Coastal Water Supply and Demand System Subject to Climatic and Non-Climatic Changes View project. Ecosystems 2013, 16, 359–381. [Google Scholar] [CrossRef]
- Theobald, D.M.; Harrison-Atlas, D.; Monahan, W.B.; Albano, C.M. Ecologically-relevant maps of landforms and physiographic diversity for climate adaptation planning. PLoS ONE 2015, 10, 1–17. [Google Scholar] [CrossRef]
- Jeong, S.; Kim, H.G.; Thorne, J.H.; Lee, H.; Cho, Y.H.; Lee, D.K.; Park, C.H.; Seo, C. Evaluating connectivity for two mid-sized mammals across modified riparian corridors with wildlife crossing monitoring and species distribution modeling. Glob. Ecol. Conserv. 2018, 16, e00485. [Google Scholar] [CrossRef]
- Dickson, B.; Jenness, J.; Beier, P. Influence of vegetation, topography, and roads on cougar movement in southern California. J. Wildl. Manag. 2005, 69, 262–276. [Google Scholar] [CrossRef]
- Gillies, C.S.; St. Clair, C.C. Riparian corridors enhance movement of a forest specialist bird in fragmented tropical forest. Proc. Natl. Acad. Sci. USA 2008, 105, 19774–19779. [Google Scholar] [CrossRef]
- Hilty, J.A.; Merenlender, A.M. Use of Riparian Corridors and Vineyards by Mammalian Predators in Northern California. Conserv. Biol. 2004, 18, 126–135. [Google Scholar] [CrossRef]
- Sabo, J.L.; Sponseller, R.; Dixon, M.; Gade, K.; Harms, T.; Heffernan, J.; Jani, A.; Katz, G.; Soykan, C.; Watts, J.; et al. Riparian zones increase regional species richness by harboring different, not more, species. Ecology 2005, 86, 56–62. [Google Scholar] [CrossRef]
- Naiman, R.J.; Decamps, H.; Pollock, M. The Role of Riparian Corridors in Maintaining Regional Biodiversity. Ecol. Appl. 1993, 3, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Parrish, J.D.; Braun, D.P.; Unnasch, R.S. Are We Conserving What We Say We Are? Measuring Ecological Integrity within Protected Areas; Oxford Academic: Oxford, UK, 2003; Volume 53. [Google Scholar]
- Belote, T.R.; Dietz, M.S.; McRae, B.H.; Theobald, D.M.; McClure, M.L.; Hugh Irwin, G.; McKinley, P.S.; Gage, J.A.; Aplet, G.H. Identifying corridors among large protected areas in the United States. PLoS ONE 2016, 11, e0154223. [Google Scholar] [CrossRef] [PubMed]
- Belote, T.R.; Dietz, M.S.; Jenkins, C.N.; McKinley, P.S.; Irwin, G.H.; Fullman, T.J.; Leppi, J.C.; Aplet, G.H. Wild, connected, and diverse: Building a more resilient system of protected areas. Ecol. Appl. 2017, 27, 1050–1056. [Google Scholar] [CrossRef] [PubMed]
- Theobald, D.M. A general model to quantify ecological integrity for landscape assessments and US application. Landsc. Ecol. 2013, 28, 1859–1874. [Google Scholar] [CrossRef]
- Dickson, B.G.; Albano, C.M.; Anantharaman, R.; Beier, P.; Fargione, J.; Graves, T.A.; Gray, M.E.; Hall, K.R.; Lawler, J.J.; Leonard, P.B.; et al. Circuit-theory applications to connectivity science and conservation. Conserv. Biol. 2019, 33, 239–249. [Google Scholar] [CrossRef]
- Crooks, K.R.; Burdett, C.L.; Theobald, D.M.; King, S.R.B.; Di Marco, M.; Rondinini, C.; Boitani, L. Quantification of habitat fragmentation reveals extinction risk in terrestrial mammals. Proc. Natl. Acad. Sci. USA 2017, 114, 7635–7640. [Google Scholar] [CrossRef]
- McGuire, J.L.; Lawler, J.J.; McRae, B.H.; Nuñez, T.A.; Theobald, D.M. Achieving climate connectivity in a fragmented landscape. Proc. Natl. Acad. Sci. USA 2016, 113, 7195–7200. [Google Scholar] [CrossRef]
- Game, E.T.; Lipsett-Moore, G.; Saxon, E.; Peterson, N.; Sheppard, S. Incorporating climate change adaptation into national conservation assessments. Glob. Chang. Biol. 2011, 17, 3150–3160. [Google Scholar] [CrossRef]
- Ohlemüller, R.; Anderson, B.J.; Araújo, M.B.; Butchart, S.H.M.; Kudrna, O.; Ridgely, R.S.; Thomas, C.D. The coincidence of climatic and species rarity: High risk to small-range species from climate change. Biol. Lett. 2008, 4, 568–572. [Google Scholar] [CrossRef]
- Williams, J.W.; Jackson, S.T.; Kutzbach, J.E. Projected distributions of novel and disappearing climates by 2100 AD. Proc. Natl. Acad. Sci. USA 2007, 104, 5738–5742. [Google Scholar] [CrossRef]
- Wright, S.J.; Muller-Landau, H.; Schipper, J. The Future of Tropical Species on a Warmer Planet. Conserv. Biol. 2009, 23, 1418–1426. [Google Scholar] [CrossRef] [PubMed]
- Iwamura, T.; Guisan, A.; Wilson, K.A.; Possingham, H.P. How robust are global conservation priorities to climate change? Glob. Environ. Chang. 2013, 23, 1277–1284. [Google Scholar] [CrossRef]
- Watson, J.E.M.; Iwamura, T.; Butt, N. Mapping vulnerability and conservation adaptation strategies under climate change. Nat. Clim. Chang. 2013, 3, 989–994. [Google Scholar] [CrossRef]
- Morelli, T.L.; Smith, A.B.; Kastely, C.R.; Mastroserio, I.; Moritz, C.; Beissinger, S.R. Anthropogenic refugia ameliorate the severe climate-related decline of a montane mammal along its trailing edge. Proc. R. Soc. B Biol. Sci. 2012, 279, 4279–4286. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.M.; Henry, H.A.; Sinclair, B.J. Cold truths: How winter drives responses of terrestrial organisms to climate change. Biol. Rev. 2015, 90, 214–235. [Google Scholar] [CrossRef]
- Dolanc, C.R.; Thorne, J.H.; Safford, H.D. Widespread shifts in the demographic structure of subalpine forests in the Sierra Nevada, California, 1934 to 2007. Glob. Ecol. Biogeogr. 2013, 22, 264–276. [Google Scholar] [CrossRef]
- Morelli, T.L.; Maher, S.P.; Lim, M.C.W.; Kastely, C.; Eastman, L.M.; Flint, L.E.; Flint, A.L.; Beissinger, S.R.; Moritz, C. Climate change refugia and habitat connectivity promote species persistence. Clim. Chang. Responses 2017, 4, 1–12. [Google Scholar] [CrossRef]
- Blondel, J.; Aronson, J.; Bodiou, J.; Boeuf, G. The Mediterranean Region: Biological Diversity in Space and Time; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Zittis, G. Observed rainfall trends and precipitation uncertainty in the vicinity of the Mediterranean, Middle East and North Africa. Appl. Clim. 2018, 134, 1207–1230. [Google Scholar] [CrossRef]
- Cook, B.I.; Anchukaitis, K.J.; Touchan, R.; Meko, D.M.; Cook, E.R. Spatiotemporal drought variability in the mediterranean over the last 900 years. J. Geophys. Res. 2016, 121, 2060–2074. [Google Scholar] [CrossRef]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Lionello, P.; Scarascia, L. The relation between climate change in the Mediterranean region and global warming. Reg. Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Western Regional Climate Center Western Regional Climate Center: California Climate Tracker. Available online: https://wrcc.dri.edu/Climate/Tracker/CA/ (accessed on 25 September 2020).
- Office of Environmental Health Hazard Assessment. Indicators of Climate Change in California; Office of Environmental Health Hazard Assessment: Sacramento, CA, USA, 2018. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Bay Area Open Space Council. The Conservation Lands Network 2.0 Report; Bay Area Open Space Council: Berkeley, CA, USA, 2019. [Google Scholar]
- Gray, M.; Micheli, E.R.; Comendant, T.; Merenlender, A.M. Sustained stakeholder engagement promotes use of co-produced climate-wise connectivity knowledge by a practitioner network. Land 2020. under review. [Google Scholar]
- Beier, P. Conceptualizing and designing corridors for climate change. Ecol. Restor. 2012, 30, 312–319. [Google Scholar] [CrossRef]
- Rouget, M.; Cowling, R.M.; Pressey, R.L.; Richardson, D.M. Identifying spatial components of ecological and evolutionary processes for regional conservation planning in the Cape Floristic Region, South Africa. Divers. Distrib. 2003, 9, 191–210. [Google Scholar] [CrossRef]
- Stella, J.C.; Rodríguez-González, P.M.; Dufour, S.; Bendix, J.; Stella, J.C.; Rodríguez-González, P.M.; Dufour, S.; Bendix, J. Riparian vegetation research in Mediterranean-climate regions: Common patterns, ecological processes, and considerations for management. Hydrobiologia 2013, 719, 291–315. [Google Scholar] [CrossRef]
- Flint, L.E.; Flint, A.L.; Thorne, J.H.; Boynton, R. Fine-scale hydrologic modeling for regional landscape applications: The California Basin Characterization Model development and performance. Ecol. Process. 2013, 2, 1–21. [Google Scholar] [CrossRef]
- California Department of Forestry and Fire Protection Vegetation (fveg)—CALFIRE FRAP. [ds1327]; CALFIRE FRAP: Sacramento, CA, USA, 2015. [Google Scholar]
- Greeninfo Network California Protected Areas Database (Version 2017a). 2017. Available online: https://www.lacounts.org/dataset/california-protected-areas-database-2017a (accessed on 25 September 2020).
- Greeninfo Network California Conservation Easement Database (Version 2016). 2016. Available online: https://www.greeninfo.org/work/project/cpad-the-california-protected-areas-database (accessed on 25 September 2020).
- Esri ArcGIS Pro, Version 10.4.1; 2016. Available online: https://desktop.arcgis.com/en/arcmap/10.4/get-started/setup/arcgis-desktop-quick-start-guide.htm (accessed on 25 September 2020).
- McRae, B.; Kavanagh, D. Linkage Mapper Connectivity Analysis Software. 2011. Available online: https://circuitscape.org/linkagemapper/ (accessed on 25 September 2020).
- Jewitt, D.; Goodman, P.S.; Erasmus, B.F.N.; O’Connor, T.G.; Witkowski, E.T.F. Planning for the Maintenance of Floristic Diversity in the Face of Land Cover and Climate Change. Environ. Manag. 2017, 59, 792–806. [Google Scholar] [CrossRef]
- Riley, S.J.; DeGloria, S.D.; Elliot, R. A Terrain Ruggedness Index that Quantifies Topographic Heterogeneity. Intermt. J. Sci. 1999, 5, 23–27. [Google Scholar]
- Voldoire, A.; Sanchez-Gomez, E.; Salas y Mélia, D.; Decharme, B.; Cassou, C.; Sénési, S.; Valcke, S.; Beau, I.; Alias, A.; Chevallier, M.; et al. The CNRM-CM5.1 global climate model: Description and basic evaluation. Clim. Dyn. 2013, 40, 2091–2121. [Google Scholar] [CrossRef]
- Riahi, K.; Rao, S.; Krey, V.; Cho, C.; Chirkov, V.; Fischer, G.; Kindermann, G.; Nakicenovic, N.; Rafaj, P. RCP 8.5-A scenario of comparatively high greenhouse gas emissions. Clim. Chang. 2011, 109, 33–57. [Google Scholar] [CrossRef]
- Townsend, P.; Masters, K.L.; Townsend, P.A.; Masters, K.L. Lattice-work corridors for climate change: A conceptual framework for biodiversity conservation and social-ecological resilience in a tropical elevational gradient. Ecol. Soc. 2015. [Google Scholar] [CrossRef]
- McRae, B.H.; Popper, K.; Jones, A.; Schindel, M.; Buttrick, S.; Hall, K.R.; Unnasch, B.; Platt, J.; Unnasch, R.S.; Platt, J. Conserving Nature’s Stage: Mapping Omnidirectional Connectivity for Resilient Terrestrial Landscapes in the Pacific Northwest; The Nature Conservancy: Portland, OR, USA, 2016; p. 47. [Google Scholar] [CrossRef]
- The Nature Conservancy Omniscape 2018. Available online: https://omniscape.codefornature.org/ (accessed on 25 September 2020).
- Marrec, R.; Abdel Moniem, H.; Iravani, M.; Hricko, B.; Kariyeva, J.; Wagner, H.H. Conceptual framework and uncertainty analysis for large-scale, species-agnostic modelling of landscape connectivity across Alberta, Canada. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.D.; Williams, B.K.; Hodges, K.E.; Gov, E. Ecological integrity assessment as a metric of biodiversity: Are we measuring what we say we are? Biodivers. Conserv. 2016, 25, 1011–1035. [Google Scholar] [CrossRef]
- Martinuzzi, S.S.; Radeloff, V.C.; Joppa, L.N.; Hamilton, C.M.; Helmers, D.P.; Plantinga, A.J.; Lewis, D.J. Scenarios of future land use change around United States’ protected areas. Biol. Conserv. 2015, 184, 446–455. [Google Scholar] [CrossRef]
- Klausmeyer, K.R.; Shaw, M.R. Climate change, habitat loss, protected areas and the climate adaptation potential of species in Mediterranean ecosystems wordwide. PLoS ONE 2009, 4, 1–9. [Google Scholar] [CrossRef]
- Araújo, M.B.; Alagador, D.; Cabeza, M.; Nogués-Bravo, D.; Thuiller, W. Climate change threatens European conservation areas. Ecol. Lett. 2011, 14, 484–492. [Google Scholar] [CrossRef]
- Regos, A.; D’Amen, M.; Titeux, N.; Herrando, S.; Guisan, A.; Brotons, L. Predicting the future effectiveness of protected areas for bird conservation in Mediterranean ecosystems under climate change and novel fire regime scenarios. Divers. Distrib. 2016, 22, 83–96. [Google Scholar] [CrossRef]
- Jetz, W.; Wilcove, D.S.; Dobson, A.P. Projected impacts of climate and land-use change on the global diversity of birds. PLoS Biol. 2007, 5, 1211–1219. [Google Scholar] [CrossRef]
- Elsen, P.R.; Monahan, W.B.; Merenlender, A.M. Topography and human pressure in mountain ranges alter expected species responses to climate change. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Osland, M.J.; Feher, L.C. Winter climate change and the poleward range expansion of a tropical invasive tree (Brazilian pepper—Schinus terebinthifolius). Glob. Chang. Biol. 2020, 26, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, J.G.; Diamond, S.E.; Buckley, L.B. Heat stress and the fitness consequences of climate change for terrestrial ectotherms. Funct. Ecol. 2013, 27, 1415–1423. [Google Scholar] [CrossRef]
- Polade, S.D.; Gershunov, A.; Cayan, D.R.; Dettinger, M.D.; Pierce, D.W. Precipitation in a warming world: Assessing projected hydro-climate changes in California and other Mediterranean climate regions. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The Community Climate System Model Version 4. Artic. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Watanabe, S.; Hajima, T.; Sudo, K.; Nagashima, T.; Takemura, T.; Okajima, H.; Nozawa, T.; Kawase, H.; Abe, M.; Yokohata, T.; et al. MIROC-ESM 2010: Model description and basic results of CMIP5-20c3m experiments. Geosci. Model Dev. 2011, 4, 845–872. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gray, M.; Micheli, E.; Comendant, T.; Merenlender, A. Quantifying Climate-Wise Connectivity across a Topographically Diverse Landscape. Land 2020, 9, 355. https://doi.org/10.3390/land9100355
Gray M, Micheli E, Comendant T, Merenlender A. Quantifying Climate-Wise Connectivity across a Topographically Diverse Landscape. Land. 2020; 9(10):355. https://doi.org/10.3390/land9100355
Chicago/Turabian StyleGray, Morgan, Elisabeth Micheli, Tosha Comendant, and Adina Merenlender. 2020. "Quantifying Climate-Wise Connectivity across a Topographically Diverse Landscape" Land 9, no. 10: 355. https://doi.org/10.3390/land9100355
APA StyleGray, M., Micheli, E., Comendant, T., & Merenlender, A. (2020). Quantifying Climate-Wise Connectivity across a Topographically Diverse Landscape. Land, 9(10), 355. https://doi.org/10.3390/land9100355