
Historical and Current Niche Construction in an Anthropogenic Biome: Old Cultural Landscapes in Southern Scandinavia

Abstract
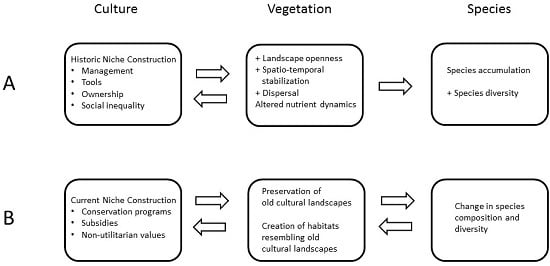
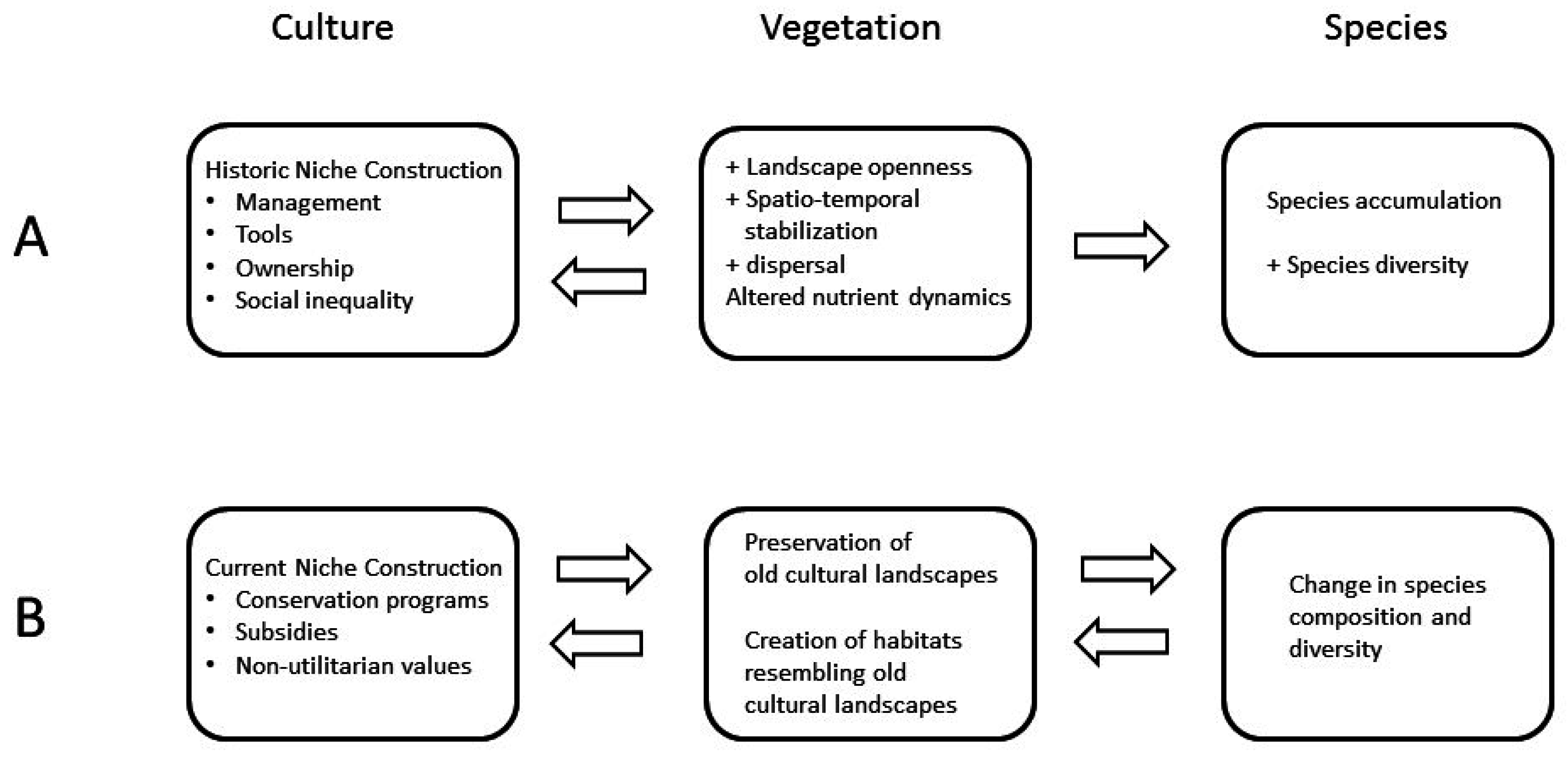
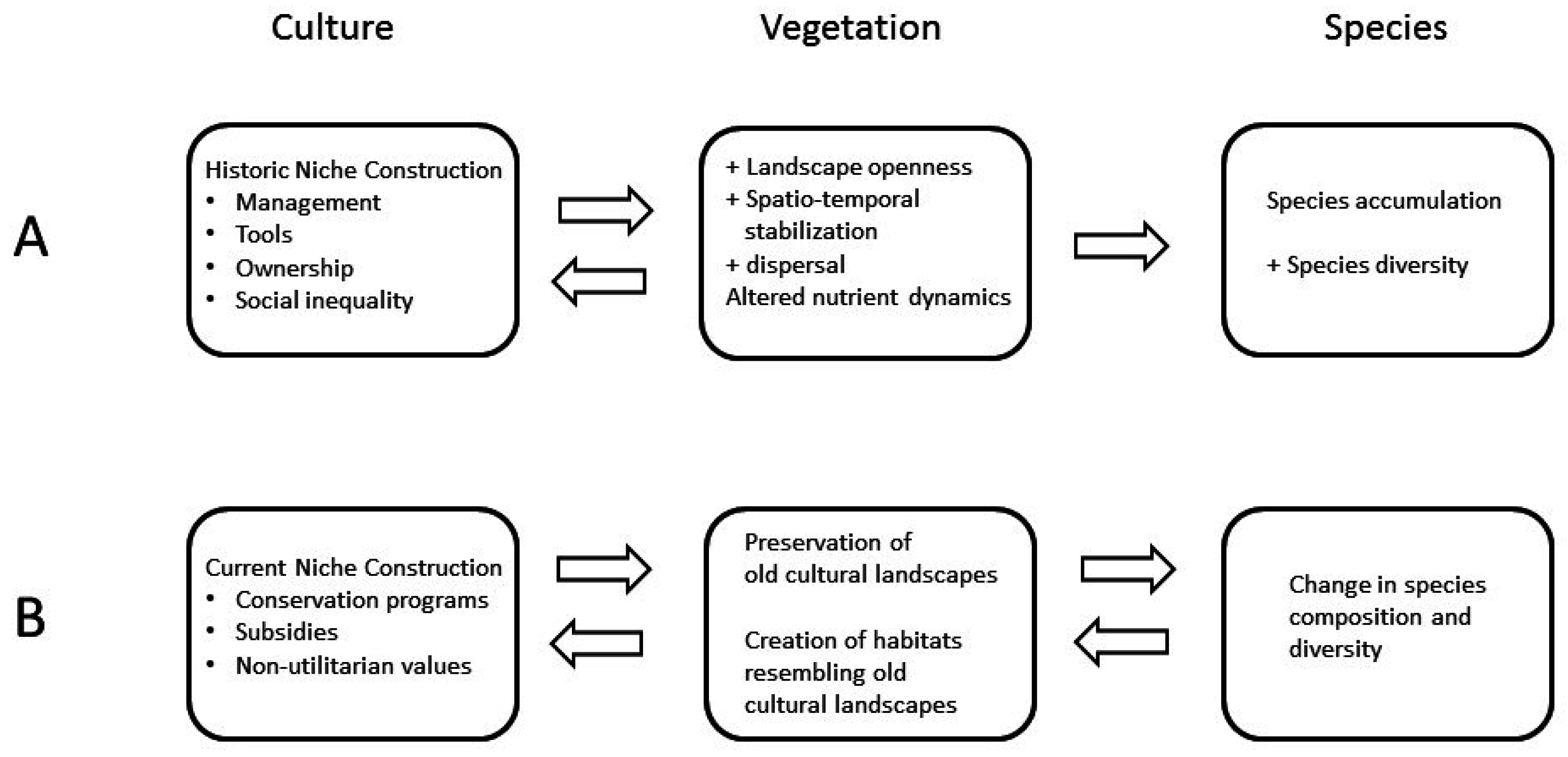
:
1. Introduction
2. Historical Context and Species Richness
3. Historical Human Niche Construction in Cultural Landscapes
4. Current Human Niche Construction Affecting Old Cultural Landscapes
4.1. Utilitarian and Non-Utilitarian Motivators for Maintaining Old Cultural Landscapes
4.2. Effects of Current Niche Construction on Remaining Old Cultural Landscapes
4.3. New Landscapes Mimicking Features of Old Cultural Landscapes
- (1)
- The area and connectivity of semi-natural grasslands may be extended using ex-arable fields. These fields are former crop fields, thus subjected to previous ploughing and fertilization, which have been recently transferred to pastures. Initially, they are species poor, but over time, they are colonized by plants typical for semi-natural grasslands [95]. Seed sowing may speed-up the process [96]. Ex-arable fields are not likely to reach the same species composition and density as well-managed semi-natural grasslands [95], but they may approach a considerable diversity. If ex-arable fields have not been too heavily fertilized previously, they will, for most people, appear indistinguishable from semi-natural grasslands, and they will contribute to maintaining a generally open landscape, rich in flowers and associated insects. In addition to considerably increasing the grazed area and the connectivity among grazed areas, these grasslands also have the advantage of being more profitable economically, thus increasing the likelihood that farmers will maintain the grazing regime [97]. Thus, by applying a landscape perspective, one may maintain features of the old cultural landscapes even within a modern landscape matrix [78,98].
- (2)
- Road verges and other constructed linear grassland landscape elements, if properly managed, may harbor plant species otherwise dependent on semi-natural grasslands and grassland-forest mosaics [99] and enhance the dispersal of these species [84]. Such linear grasslands have also been found favorable for butterflies [45,100]. However, road verges are also favorable for several invasive plants, for example lupines (Lupinus polyphyllus). Lupines are often regarded as beautiful when flowering along the motor-ways. Their ecological effects are more controversial. As they may develop mono-specific stands, lupines may be negative for native flora [89]. However, their massive flower production may also have positive effects on pollinators, such as bumble-bees, and thereby also on the pollination of other plants [101]. Road verges undoubtedly open space for the development of new plant communities, but it is still an open question how these communities will be perceived and valuated in the future.
- (3)
- Park landscapes may hold considerable biological and aesthetic qualities and may function as refuges for a range of temperate forest species [102,103]. Species diversity is dependent on the age of the park, however, suggesting that if recently created, the park landscapes may need extensive time periods to reach their potential as diversity hotspots. Even though the parks that currently have high diversity are old, they were once created, and there is no practical limitation to create new park landscapes. A long-term management strategy will thus be needed in order to utilize parks for this purpose. As the fraction of urban woodland in Swedish cities is on average 20% of urban area [104], there is a potential for creating biologically diverse landscapes even in strongly transformed landscapes.
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ellis, E.C.; Ramankutty, N. Putting people on the map: Anthropogenic biomes of the world. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Ellis, E.C. Ecology in an anthropogenic biosphere. Ecol. Monogr. 2015, 85, 287–331. [Google Scholar] [CrossRef]
- Walter, H. Vegetation of the Earth and Ecological Systems of the Geo-biosphere, 2nd ed.; Springer: New York, NY, USA, 1979. [Google Scholar]
- Lewis, S.L.; Maslin, M.A. Defining the anthropocene. Nature 2015, 519, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, R.J.; Higgs, E.; Harris, J.A. Novel ecosystems: Implications for conservation and restoration. Trends Ecol. Evol. 2009, 24, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Kareiva, P.; Marvier, M. What is conservation science? BioScience 2012, 62, 962–969. [Google Scholar]
- Doak, D.F.; Bakker, V.J.; Goldstein, B.E.; Hale, B. What is the future of conservation? Trends Ecol. Evol. 2014, 29, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Marris, E. Rambunctious Garden: Saving Nature in a Post-Wild World; Bloomsbury Publishing: New York, NY, USA, 2011. [Google Scholar]
- Mace, G.M. Whose conservation? Science 2014, 345, 1558–1560. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J.; Quinn, J.E.; Ellis, E.C.; Shaw, M.R.; Dorning, M.A.; Hallett, L.M.; Heller, N.E.; Hobbs, R.J.; Kraft, C.E.; Law, E.; et al. Conservation opportunities across the world’s anthromes. Divers. Distrib. 2014, 20, 745–755. [Google Scholar] [CrossRef]
- Miller, B.; Soulé, M.E.; Terborgh, J. ”New conservation” or surrender to development? Animal Conserv. 2014, 17, 509–515. [Google Scholar] [CrossRef]
- Wuerthner, G.; Crist, E.; Butler, T. Keeping the Wild: Against the Domestication of Earth; Island Press: Washington, DC, USA, 2014. [Google Scholar]
- Corlett, R.T. The Anthropocene concept in ecology and conservation. Trends Ecol. Evol. 2015, 30, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Odling-Smee, F.J.; Laland, K.N.; Feldman, M.W. Niche Construction: The Neglected Process in Evolution; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- Kendal, J.; Tehrani, J.J.; Odling-Smee, J. Human niche construction in interdisciplinary focus: Introduction. Philos. Trans. R. Soc. Biol. Sci. 2011, 366, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Laland, K.N.; O’Brien, M.J. Cultural niche construction: An introduction. Biol. Theory 2011, 6, 191–202. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Laland, K.N. Genes, culture and agriculture: An example of human niche construction. Curr. Anthropol. 2012, 53, 434–470. [Google Scholar] [CrossRef]
- Smith, B.D. Neo-Darwinism, niche construction theory, and the initial domestication of plants and animals. Evol. Ecol. 2016, 30, 307–324. [Google Scholar] [CrossRef]
- Pelletier, F.; Garant, D.; Hendry, A.P. Eco-evolutionary dynamics. Philos. Trans. R. Soc. Biol. Sci. 2009, 364, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Schoener, T.W. The newest synthesis: Understanding the interplay of evolutionary and ecological dynamics. Science 2011, 331, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.K.; Schweitzer, J.A.; Úbeda, F.; Koricheva, J.; LeRoy, C.J.; Madritch, M.D.; Rehill, B.J.; Bangert, R.K.; Fischer, D.G.; Allan, G.J.; et al. From genes to ecosystems: A synthesis of the effects of plant genetic factors across levels of organization. Philos. Trans. R. Soc. Biol. Sci. 2009, 364, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, O. Vegetation change and eco-evolutionary dynamics. J. Veg. Sci. 2014, 25, 1141–1147. [Google Scholar] [CrossRef]
- Erickson, C.L. The domesticated landscapes of the Bolivian Amazon. In Time and Complexity in Historical Ecology; Balée, W., Erickson, C.L., Eds.; Columbia University Press: New York, NY, USA, 2006; pp. 235–278. [Google Scholar]
- Widgren, M. Landscape research in a world of domesticated landscapes: The role of values, theory, and concepts. Quat. Int. 2012, 251, 117–124. [Google Scholar] [CrossRef]
- Antrop, M. Why landscapes of the past are important for the future. Landsc. Urban Plan. 2005, 70, 21–34. [Google Scholar] [CrossRef]
- European Landscape Convention. Council of Europe, 2000. Available online: http://www.coe.int/EuropeanLandscapeConvention (accessed on 29 April 2016).
- Eriksson, O. Species pools in cultural landscapes—Niche construction, ecological opportunity and niche shifts. Ecography 2013, 36, 403–413. [Google Scholar] [CrossRef]
- Eriksson, O.; Arnell, M. Niche construction, entanglement and landscape domestication in Scandinavian infield systems. Landsc. Res. 2016. [Google Scholar] [CrossRef]
- Sjörs, H. The background: Geology, climate and vegetation. In Swedish Plant Geography; Rydin, H., Snoeijs, P., Diekmann, M., Eds.; Acta Phytogeographica Suecica 84; Svenska Vaxtgeografiska Sallskapet: Uppsala, Sweden, 1999; pp. 5–14. [Google Scholar]
- Larsson, J. The expansion and decline of a transhumance system in Sweden, 1550-1920. Hist. Agrar. 2012, 56, 11–39. [Google Scholar]
- Brännlund, I.; Axelsson, P. Reindeer management during the colonization of Sami lands: A long-term perspective of vulnerability and adaptation strategies. Glob. Environ. Chang. 2011, 21, 1095–1105. [Google Scholar] [CrossRef]
- Berglund, B.E. The cultural landscape during 6000 years in southern Sweden—The Ystad Project. Ecol. Bull. 1991, 41, 1–495. [Google Scholar]
- Eriksson, O.; Cousins, S.A.O. Historical landscape perspectives on grasslands in Sweden and the Baltic region. Land 2014, 3, 300–321. [Google Scholar] [CrossRef]
- Emanuelsson, U. The Rural Landscapes of Europe: How Man has shaped European Nature; Swedish Research Council Formas: Stockholm, Sweden, 2009. [Google Scholar]
- Pedersen, E.A.; Widgren, M. Agriculture in Sweden, 800 BC–AD 1000. In The Agrarian History of Sweden: From 4000 BC to AD 2000; Morell, M., Myrdal, J., Eds.; Nordic Academic Press: Lund, Sweden, 2011; pp. 46–71. [Google Scholar]
- Berglund, B.E.; Kitigawa, J.; Lagerås, P.; Nakamura, K.; Sasaki, N.; Yasuda, Y. Traditional farming landscapes for sustainable living in Scandinavia and Japan: Global revival through the Satoyama initiative. Ambio 2014, 43, 559–578. [Google Scholar] [CrossRef] [PubMed]
- Slotte, H. Harvesting of leaf-hay shapes the Swedish landscape. Landsc. Ecol. 2001, 16, 691–702. [Google Scholar] [CrossRef]
- Widgren, M. Climate and causation in the Swedish Iron Age: Learning from the present to understand the past. Geogr. Tidsskr. Dan. J. Geogr. 2012, 112, 126–134. [Google Scholar] [CrossRef]
- Cousins, S.A.O.; Auffret, A.G.; Lindgren, J.; Tränk, L. Regional-scale land-cover change during the 20th century and its consequences for biodiversity. Ambio 2015, 44, S17–S27. [Google Scholar] [CrossRef] [PubMed]
- Swedish Board of Agriculture. Betesmarker och slåtterängar med miljöersättning; Jordbruksverket Rapport: Jönköping, Sweden, 2012. [Google Scholar]
- Eriksson, O.; Wikström, S.; Eriksson, Å.; Lindborg, R. Species-rich Scandinavian grasslands are inherently open to invasion. Biol. Invasions 2006, 8, 355–363. [Google Scholar] [CrossRef]
- Öster, M.; Eriksson, O. Recruitment in species-rich grasslands: The effects of functional traits and propagule pressure. J. Plant Ecol. 2012, 5, 260–269. [Google Scholar] [CrossRef]
- Vessby, K.; Söderström, B.; Glimskär, A.; Svensson, B. Species-richness correlations of six different taxa in Swedish seminatural grasslands. Conserv. Biol. 2002, 16, 430–439. [Google Scholar] [CrossRef]
- Öster, M. Low congruence between the diversity of waxcap (Hygrocybe spp.) fungi and vascular plants in semi-natural grasslands. Basic Appl. Ecol. 2008, 9, 514–522. [Google Scholar] [CrossRef]
- Berg, Å.; Ahrné, K.; Öckinger, E.; Svensson, R.; Söderström, B. Butterfly distribution and abundance is affected by variation in the Swedish forest-farmland landscape. Biol. Conserv. 2011, 144, 2819–2831. [Google Scholar] [CrossRef]
- Paltto, H.; Thomasson, I.; Nordén, B. Multispecies and multiscale conservation planning: Setting quantitative targets for red-listed lichens on ancient oaks. Conserv. Biol. 2010, 24, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Milberg, P.; Bergman, K.O.; Johansson, H.; Jansson, N. Low host-tree preferences among saproxylic beetles: A comparison of four deciduous species. Insect Conserv. Divers. 2014, 7, 508–522. [Google Scholar] [CrossRef]
- Wretenberg, J.; Lindström, Å.; Svensson, S.; Thierfelder, T.; Pärt, T. Population trends of farmland birds in Sweden and England: Similar trende but different patterns of agricultural intensification. J. Appl. Ecol. 2006, 43, 1110–1120. [Google Scholar] [CrossRef]
- Sandström, J.; Bjelke, U.; Carlberg, T.; Sundberg, S. Tillstånd och trender för arter och deras livsmiljöer—rödlistade arter i Sverige 2015; Artdatabanken Rapporterar 17; The Swedish Species Information Centre (SLU): Uppsala, Sweden, 2015. [Google Scholar]
- Lindborg, R.; Eriksson, O. Historical landscape connectivity affects present plant species diversity. Ecology 2004, 85, 1840–1845. [Google Scholar] [CrossRef]
- Auffret, A.G.; Plue, J.; Cousins, S.A.O. The spatial and temporal components of functional connectivity in fragmented landscapes. Ambio 2015, 44, S51–S59. [Google Scholar] [CrossRef] [PubMed]
- Vera, F.W.M. Grazing Ecology and Forest History; CABI Publishing: Wallingford, UK, 2000. [Google Scholar]
- Svenning, J.C. A review of natural vegetation openness in north-western Europe. Biol. Conserv. 2002, 104, 133–148. [Google Scholar] [CrossRef]
- Mitchell, F.J.G. How open were European primeval forests? Hypothesis testing using palaeoecological data. J. Ecol. 2005, 93, 168–177. [Google Scholar] [CrossRef]
- Whitehouse, N.J.; Smith, D. How fragmented was the British Holocene wildwood? Perspectives on the “Vera” grazing debate from the fossil beetle record. Quat. Sci. Rev. 2010, 29, 539–553. [Google Scholar] [CrossRef]
- Sandom, C.J.; Ejrnaes, R.; Hansen, M.D.D.; Svenning, J.C. High herbivore density associated with vegetation diversity in interglacial ecosystems. Proc. Natl. Acad. Sci. USA 2014, 111, 4162–4167. [Google Scholar] [CrossRef] [PubMed]
- Berglund, B.E.; Gaillard, M.J.; Björkman, L.; Persson, T. Long-term changes in floristic diversity in southern Sweden: Palynological richness, vegetation dynamics and land-use. Veg. Hist. Archaeobot. 2008, 17, 573–583. [Google Scholar] [CrossRef]
- Pärtel, M.; Helm, A.; Reitalu, T.; Liira, J.; Zobel, M. Grassland diversity related to the Late Iron Age human population density. J. Ecol. 2007, 95, 574–582. [Google Scholar] [CrossRef]
- Veen, P.; Jefferson, R.; de Smidt, J.; van der Straaten, J. Grasslands in Europe of High Nature Value; KNNV Publishing: Den Haag, The Netherlands, 2009. [Google Scholar]
- Chapron, G.; Kaczenskyand, P.; Linnell, J.D.C.; von Arx, M.; Huber, D.; Andrén, H.; López-Bao, J.V.; Adamec, M.; Álvares, F.; Anders, O.; et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef] [PubMed]
- Stenseke, M. Local participation in cultural landscape maintenance: Lessons from Sweden. Land Use Policy 2009, 26, 214–223. [Google Scholar] [CrossRef]
- Balée, W. The research program of historical ecology. Annu. Rev. Anthropol. 2006, 35, 75–98. [Google Scholar]
- Szabó, P.; Hedl, R. Advancing the integration of history and ecology for conservation. Conserv. Biol. 2011, 25, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Higgs, E.; Falk, D.A.; Guerrini, A.; Hall, M.; Harris, J.; Hobbs, R.J.; Jackson, S.T.; Rhemtulla, J.M.; Throop, W. The changing role of history in restoration ecology. Front. Ecol. Environ. 2014, 12, 499–506. [Google Scholar] [CrossRef]
- Loreau, M. Reconciling utilitarian and non-utilitarian approaches to biodiversity conservation. Ethics Sci. Eviron. Polit. 2014, 14, 27–32. [Google Scholar] [CrossRef]
- Millenium Ecosystem Assessment. Ecosystems and Human Well-Being: Synthesis; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Rader, R.; Birkhofer, K.; Schmucki, R.; Smith, H.G.; Stjernman, M.; Lindborg, R. Organic farming and heterogeneous landscapes positively affect different measures of plant diversity. J. Appl. Ecol. 2014, 51, 1544–1553. [Google Scholar] [CrossRef]
- Samnegård, U.; Persson, A.S.; Smith, H.G. Gardens benefit bees and enhance pollination in intensively managed farmland. Biol. Conserv. 2011, 144, 2602–2606. [Google Scholar] [CrossRef]
- Queiroz, C.; Meacham, M.; Richter, K.; Norström, A.V.; Andersson, E.; Norberg, J.; Peterson, G. Mapping bundles of ecosystem services reveals distinct types of multifunctionality within a Swedish landscape. Ambio 2015, 44, S89–S101. [Google Scholar] [CrossRef] [PubMed]
- Stenseke, M. Biodiversity and the local context: Linking seminatural grasslands and their future use to social aspects. Environ. Sci. Policy 2006, 9, 350–359. [Google Scholar] [CrossRef]
- Lindborg, R.; Stenseke, M.; Cousins, S.A.O.; Bengtsson, J.; Berg, Å.; Gustafsson, T.; Sjödin, N.E.; Eriksson, O. Investigating biodiversity trajectories using scenarios: Lessons from two contrasting agricultural landscapes. J. Environ. Manag. 2009, 91, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Schama, S. Landscape and Memory; Vintage Books: New York, NY, USA, 1995. [Google Scholar]
- Hobsbawm, E. Introduction: Inventing traditions. In The Invention of Tradition; Hobsbawm, E., Ranger, T., Eds.; Cambridge University Press: Cambridge, UK, 1983; pp. 1–14. [Google Scholar]
- Graham, B.; Howard, P. Heritage and identity. In The Ashgate Research Companion to Heritage and Identity; Graham, B.J., Howard, P., Eds.; Ashgate Publishing: Aldershot, UK, 2008; pp. 1–15. [Google Scholar]
- Storm, A. Hope and Rust: Reinterpreting the Industrial Place in the Late 20th Century. Ph.D. Thesis, Royal Institute of Technology, Stockholm, Sweden, 2008. [Google Scholar]
- Braaksma, P.J.; Jacobs, M.H.; van der Zande, A.N. The production of local landscape heritage: A case study in the Netherlands. Landsc. Res. 2016, 41, 64–78. [Google Scholar] [CrossRef]
- Lindborg, R.; Bengtsson, J.; Berg, Å.; Cousins, S.A.O.; Eriksson, O.; Gustafsson, T.; Hasund, K.P.; Lenoir, L.; Pihlgren, A.; Sjödin, E.; et al. A landscape perspective on conservation of semi-natural grasslands. Agric. Ecosyst. Environ. 2008, 125, 213–222. [Google Scholar] [CrossRef]
- Orians, G.H.; Heerwagen, J.H. Evolved responses to landscapes. In The Adapted Mind: Evolutionary Psychology and the Generation of Culture; Barkow, J.H., Cosmides, L., Tooby, J., Eds.; Oxford University Press: Oxford, UK, 1992; pp. 555–579. [Google Scholar]
- Kaplan, S. Environmental preference in a knowledge-seeking, knowledge-using organism. In The Adapted Mind: Evolutionary Psychology and the Generation of Culture; Barkow, J.H., Cosmides, L., Tooby, J., Eds.; Oxford University Press: Oxford, UK, 1992; pp. 581–598. [Google Scholar]
- Dutton, D. The Art Instinct: Beauty, Pleasure and Human Evolution; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Portera, M. Toward an integrated science of aesthetics: Getting rid of the main misunderstandings in evolutionary aesthetics. Aisthesis. Pratiche, linguaggi e saperi dell’estetico 2015, 8, 194–203. [Google Scholar]
- Poschlod, P.; Bonn, S. Changing dispersal processes in the central European landscape since the last Ice Age: An explanation for the actual decrease of plant species richness in different habitats? Acta Bot. Neerl. 1998, 47, 27–44. [Google Scholar]
- Auffret, A.G.; Berg, J.; Cousins, S.A.O. The geography of human-mediated dispersal. Divers. Distrib. 2014, 20, 1450–1456. [Google Scholar] [CrossRef]
- Cousins, S.A.O. Extinction debt in fragmented grasslands: Paid or not? J. Veg. Sci. 2009, 20, 3–7. [Google Scholar] [CrossRef]
- Dahlström, A.; Iuga, A.M.; Lennartsson, T. Managing biodiversity rich hay meadows in the EU: A comparison of Swedish and Romanian grasslands. Environ. Conserv. 2013, 40, 194–205. [Google Scholar] [CrossRef]
- Eriksson, O.; Bolmgren, K.; Westin, A.; Lennartsson, T. Historic hay cutting dates from Sweden 1873–1951 and their implications for conservation management of species-rich meadows. Biol. Conserv. 2015, 184, 100–107. [Google Scholar] [CrossRef]
- Kiviniemi, K.; Eriksson, O. Size-related deterioration of semi-natural grassland fragments in Sweden. Divers. Distrib. 2002, 8, 21–29. [Google Scholar] [CrossRef]
- Ramula, S.; Pihlaja, K. Plant communities and the reproductive success of native plants after the invasion of an ornamental herb. Biol. Invasions 2012, 14, 2079–2090. [Google Scholar] [CrossRef]
- Cousins, S.A.O.; Eriksson, O. After the hotspots are gone: Land use history and grassland plant species diversity in a strongly transformed agricultural landscape. Appl. Veg. Sci. 2008, 11, 365–374. [Google Scholar] [CrossRef]
- Saltzman, K.; Head, L.; Stenseke, M. Do cows belong in nature? The cultural basis of agriculture in Sweden and Australia. J. Rural Stud. 2011, 27, 54–62. [Google Scholar] [CrossRef]
- Proctor, J.D. The social construction of nature: Relativist accusations, pragmatist and critical realist responses. Ann. Assoc. Am. Geogr. 1998, 88, 352–376. [Google Scholar] [CrossRef]
- Eriksson, O. Human niche construction and the rural environment. Rural Landsc. Soc. Environ. Hist. 2014, 1, 1–4. [Google Scholar] [CrossRef]
- Plieninger, T.; Hartel, T.; Martín-López, B.; Beaufoy, G.; Bergmeier, E.; Kirby, K.; Montero, M.J.; Moreno, G.; Oteros-Rozas, E.; Van Uytvanck, J. Wood-pastures of Europe: Geographic coverage, social-ecological values, conservation management, and policy implications. Biol. Conserv. 2015, 190, 70–79. [Google Scholar] [CrossRef]
- Öster, M.; Ask, K.; Cousins, S.A.O.; Eriksson, O. Dispersal and establishment limitation reduces the potential for successful restoration of semi-natural grassland communities on former arable fields. J. Appl. Ecol. 2009, 46, 1266–1274. [Google Scholar] [CrossRef]
- Marteinsdóttir, B. Seed rain and seed bank reveal that seed limitation strongly influences plant community assembly in grasslands. PlosOne 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Kumm, K.I. Sustainable management of Swedish seminatural pastures with high species diversity. J. Nat. Conserv. 2003, 11, 117–125. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Higgs, E.; Hall, C.M.; Bridgewater, P.; Chapin, F.S., III; Ellis, E.C.; Ewel, J.J.; Hallett, L.M.; Harris, J.; Hulvey, K.B.; et al. Managing the whole landscape: Historical, hybrid, and novel ecosystems. Front. Ecol. Environ. 2014, 12, 557–564. [Google Scholar] [CrossRef]
- Auestad, I.; Rydgren, K.; Austad, I. Road verges: Potential refuges for declining grassland species despite remnant vegetation dynamics. Ann. Bot. Fenn. 2011, 48, 289–303. [Google Scholar] [CrossRef]
- Bubová, T.; Vrabec, V.; Kulma, M.; Nowicki, P. Land management impacts on European butterflies of conservation concern: A review. J. Insect Conserv. 2015, 19, 805–821. [Google Scholar] [CrossRef]
- Jakobsson, A.; Padrón, B. Does the invasive Lupinus polyphyllus increase pollinator visitation to a native herb through effects on pollinator population sizes? Oecologia 2014, 174, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Lõhmus, K.; Liira, J. Old rural parks support higher biodiversity than forest remnants. Basic Appl. Ecol. 2013, 14, 165–173. [Google Scholar] [CrossRef]
- Lõhmus, K.; Paal, T.; Liira, J. Long-term colonization ecology of forest-dwelling species in a fragmented rural landscape: Dispersal versus establishment. Ecol. Evol. 2014, 4, 3113–3126. [Google Scholar] [CrossRef] [PubMed]
- Hedblom, M.; Söderström, B. Woodlands across Swedish urban gradients: Status, structure and management implications. Landsc. Urban Plan. 2008, 84, 62–73. [Google Scholar] [CrossRef]
- Odling-Smee, J.; Erwin, D.H.; Palkovacs, E.P.; Feldman, M.W.; Laland, K.N. Niche construction theory: A practical guide for ecologists. Quart. Rev. Biol. 2013, 88, 3–28. [Google Scholar] [CrossRef]
- Papworth, S.K.; Rist, J.; Coad, L.; Milner-Gulland, E.J. Evidence for shifting baseline syndrome in conservation. Conserv. Lett. 2009, 2, 93–100. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components of Current Niche Construction | Comparison with Historical Niche Construction | Effects on Species Composition and Richness |
|---|---|---|
| Landscape configuration |
|
|
| Dispersal |
|
|
| Management |
|
|
| Inflow of species from dominating vegetation, and non-native species |
|
|
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eriksson, O. Historical and Current Niche Construction in an Anthropogenic Biome: Old Cultural Landscapes in Southern Scandinavia. Land 2016, 5, 42. https://doi.org/10.3390/land5040042
Eriksson O. Historical and Current Niche Construction in an Anthropogenic Biome: Old Cultural Landscapes in Southern Scandinavia. Land. 2016; 5(4):42. https://doi.org/10.3390/land5040042
Chicago/Turabian StyleEriksson, Ove. 2016. "Historical and Current Niche Construction in an Anthropogenic Biome: Old Cultural Landscapes in Southern Scandinavia" Land 5, no. 4: 42. https://doi.org/10.3390/land5040042
APA StyleEriksson, O. (2016). Historical and Current Niche Construction in an Anthropogenic Biome: Old Cultural Landscapes in Southern Scandinavia. Land, 5(4), 42. https://doi.org/10.3390/land5040042