
Mapping Vegetation Morphology Types in Southern Africa Savanna Using MODIS Time-Series Metrics: A Case Study of Central Kalahari, Botswana


Abstract
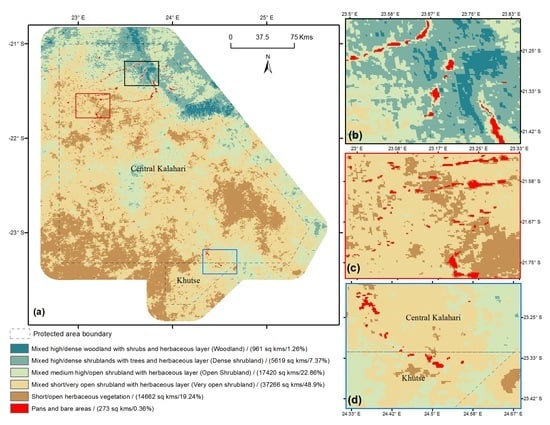
:
1. Introduction
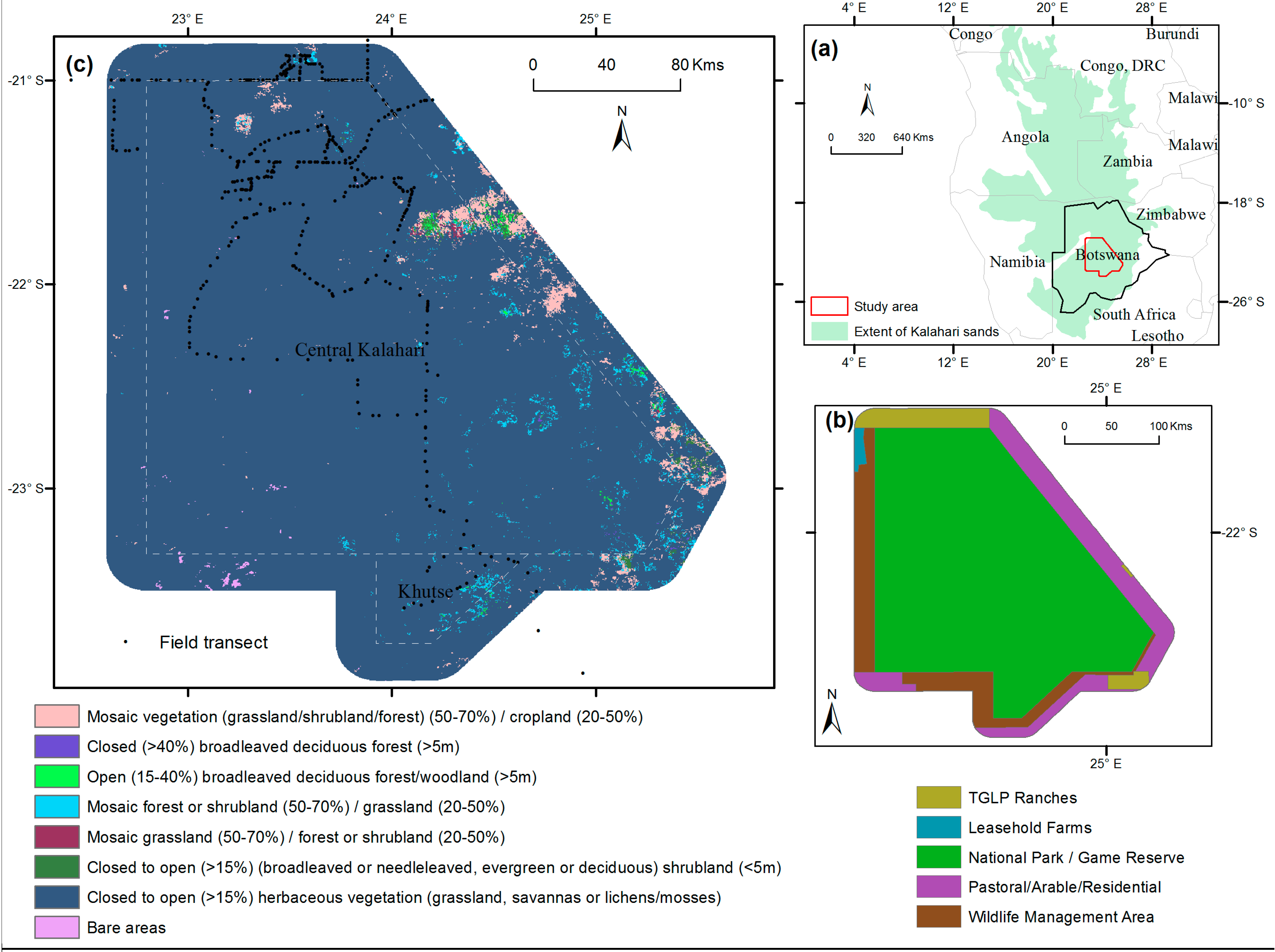
2. Study Area
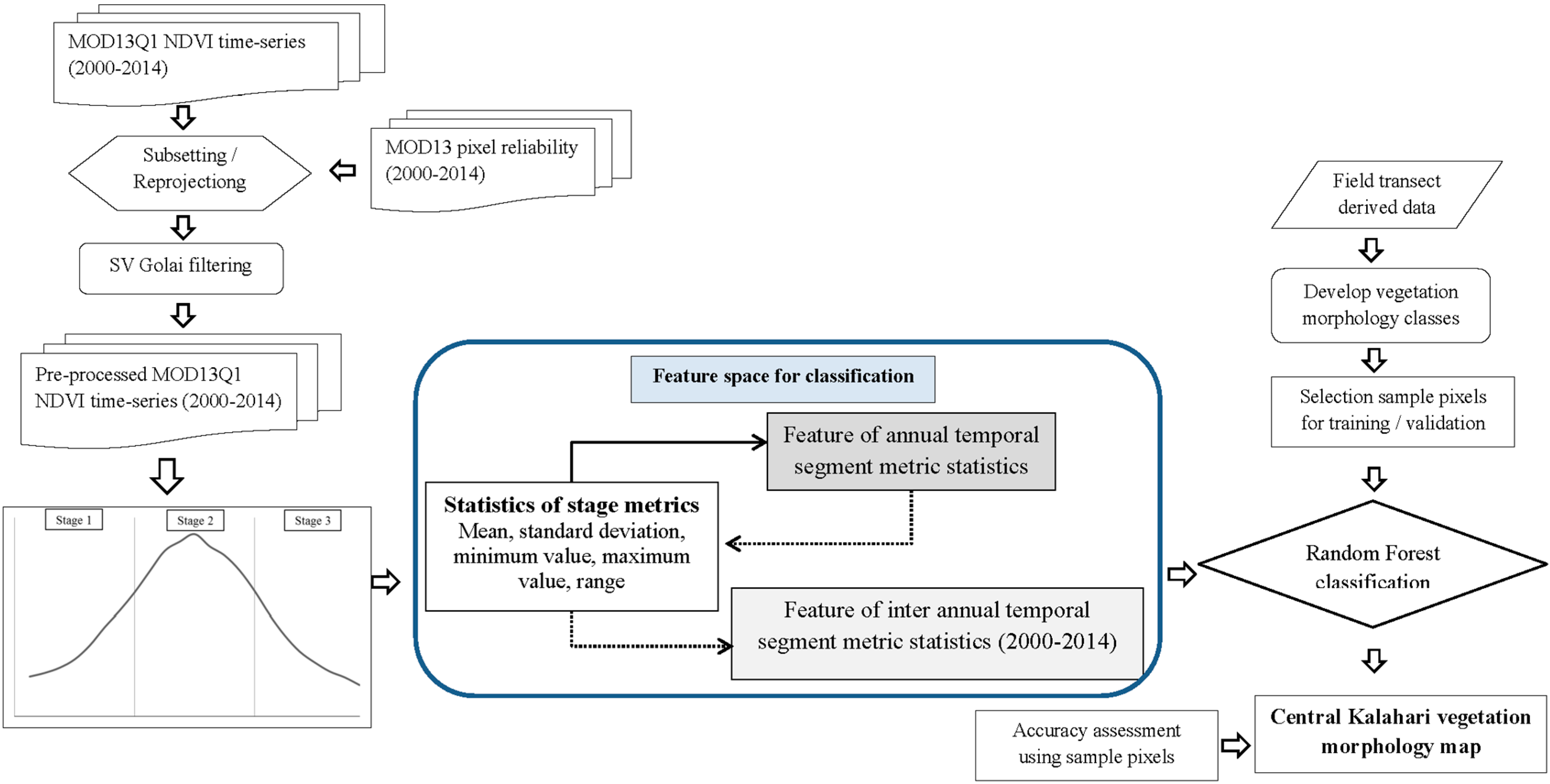
3. Data and Methods
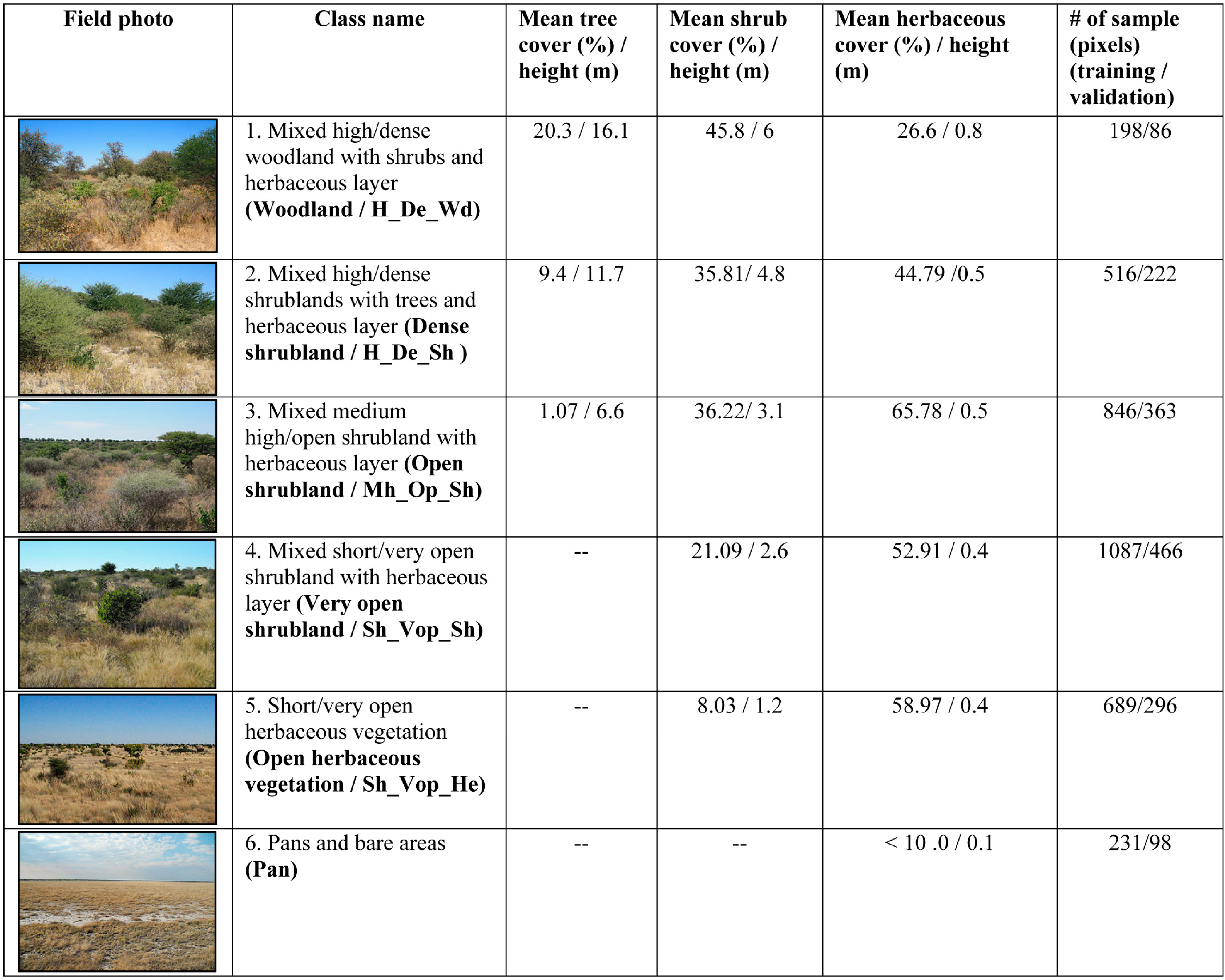
3.1. Field Data Processing and Development of Vegetation Morphology Class
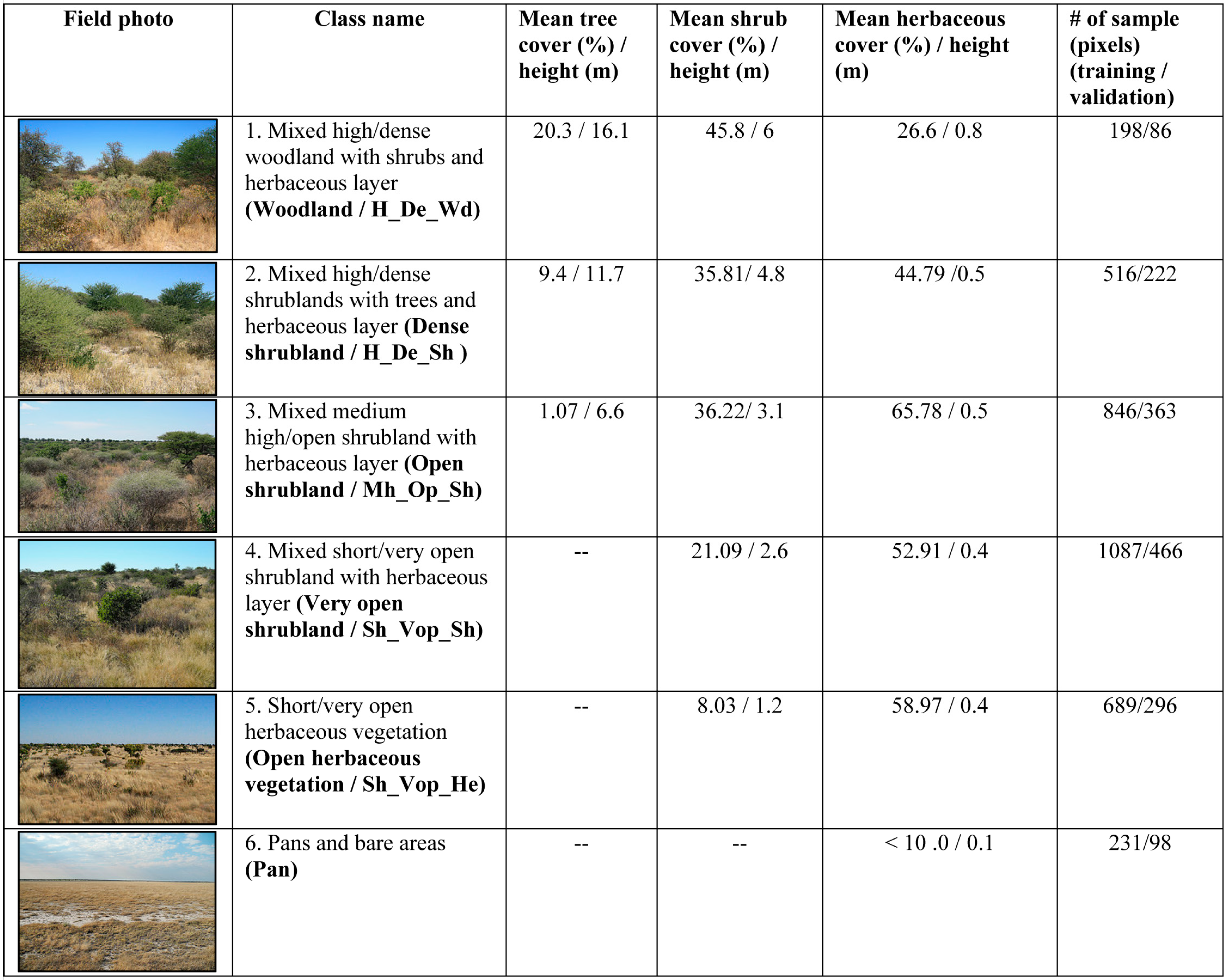
3.2. Remotely Sensed Data and Pre-Processing
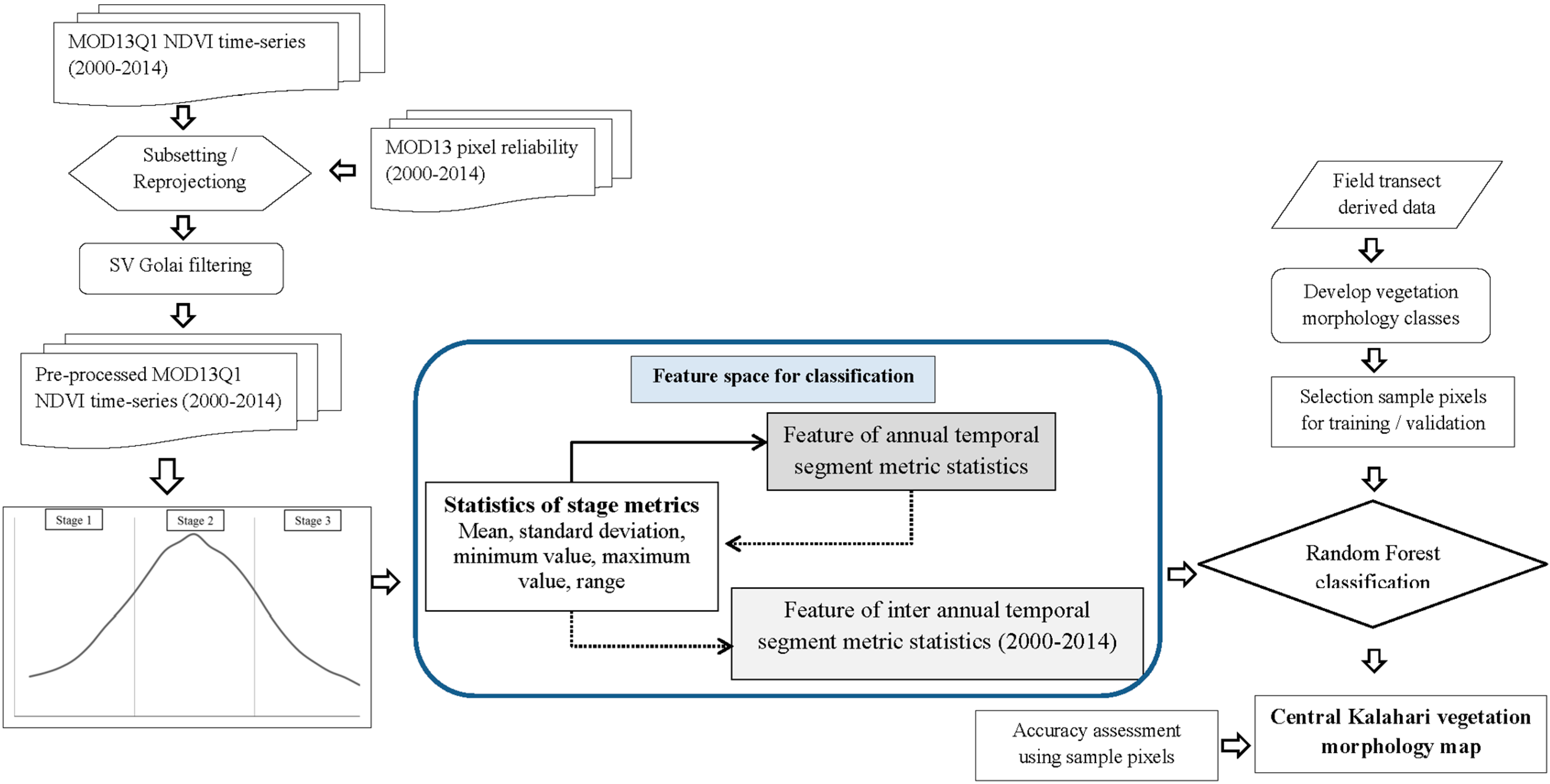
3.3. Calculating Time-Series Metrics, Separability Analysis and Random Forest Classification
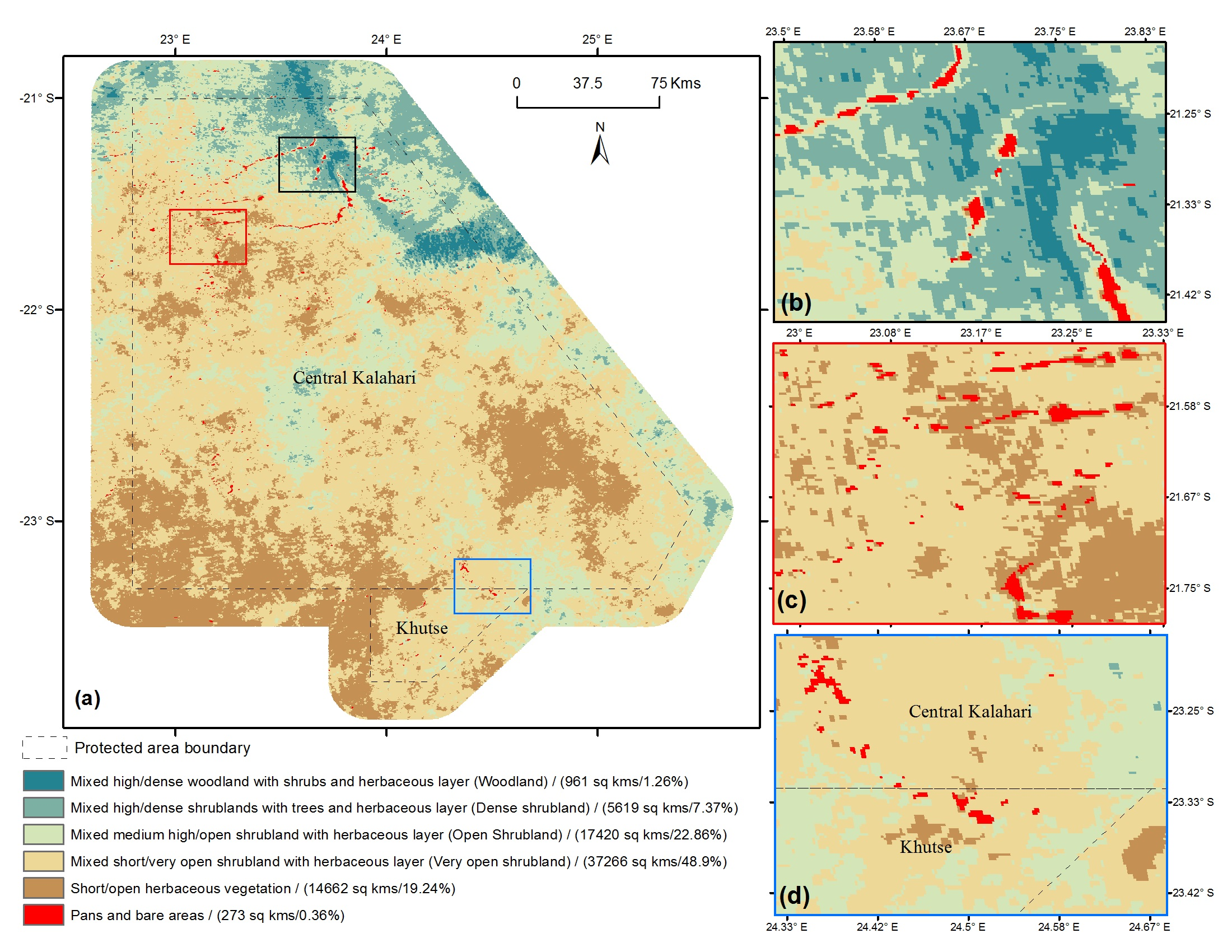
4. Results and Discussion
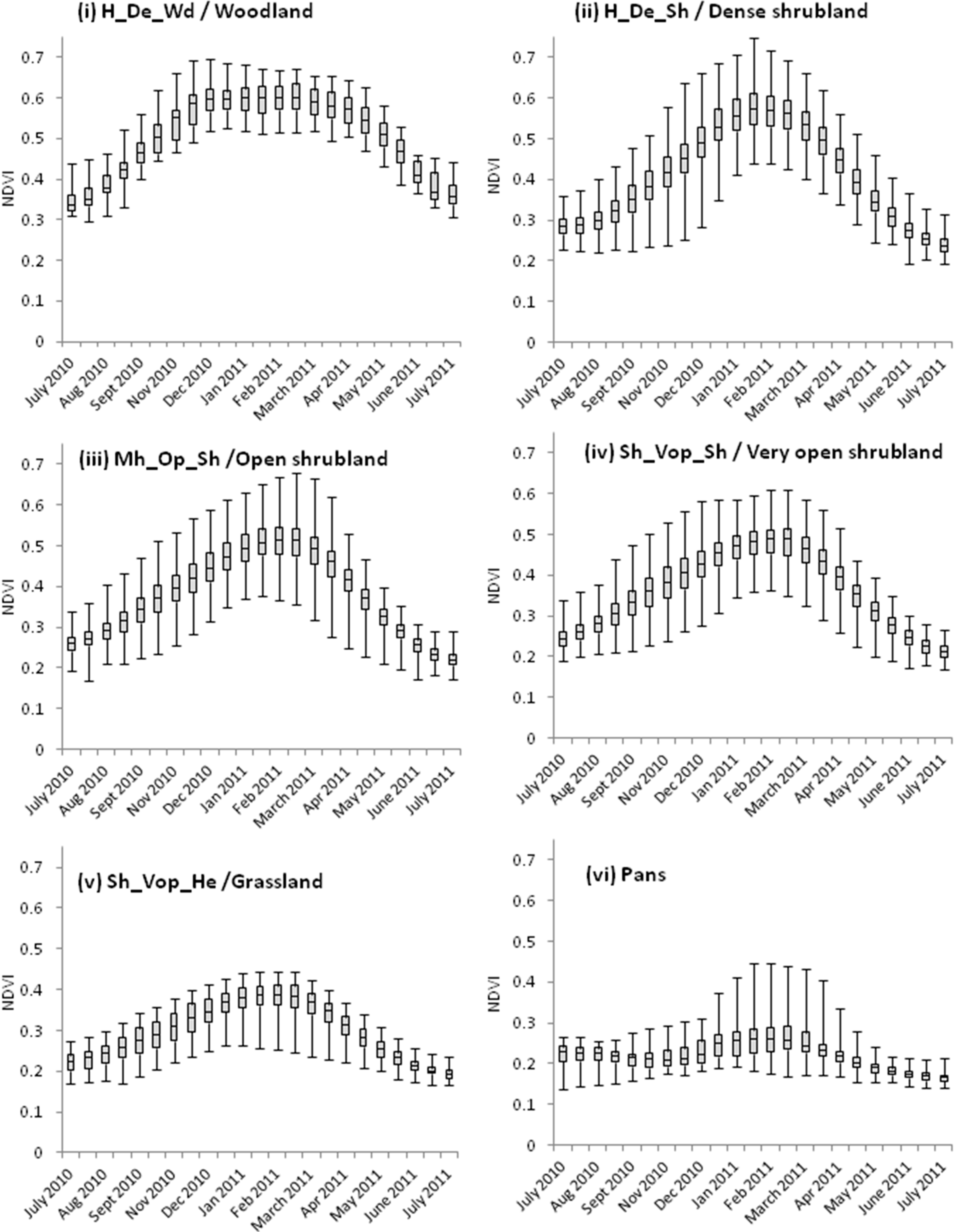

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
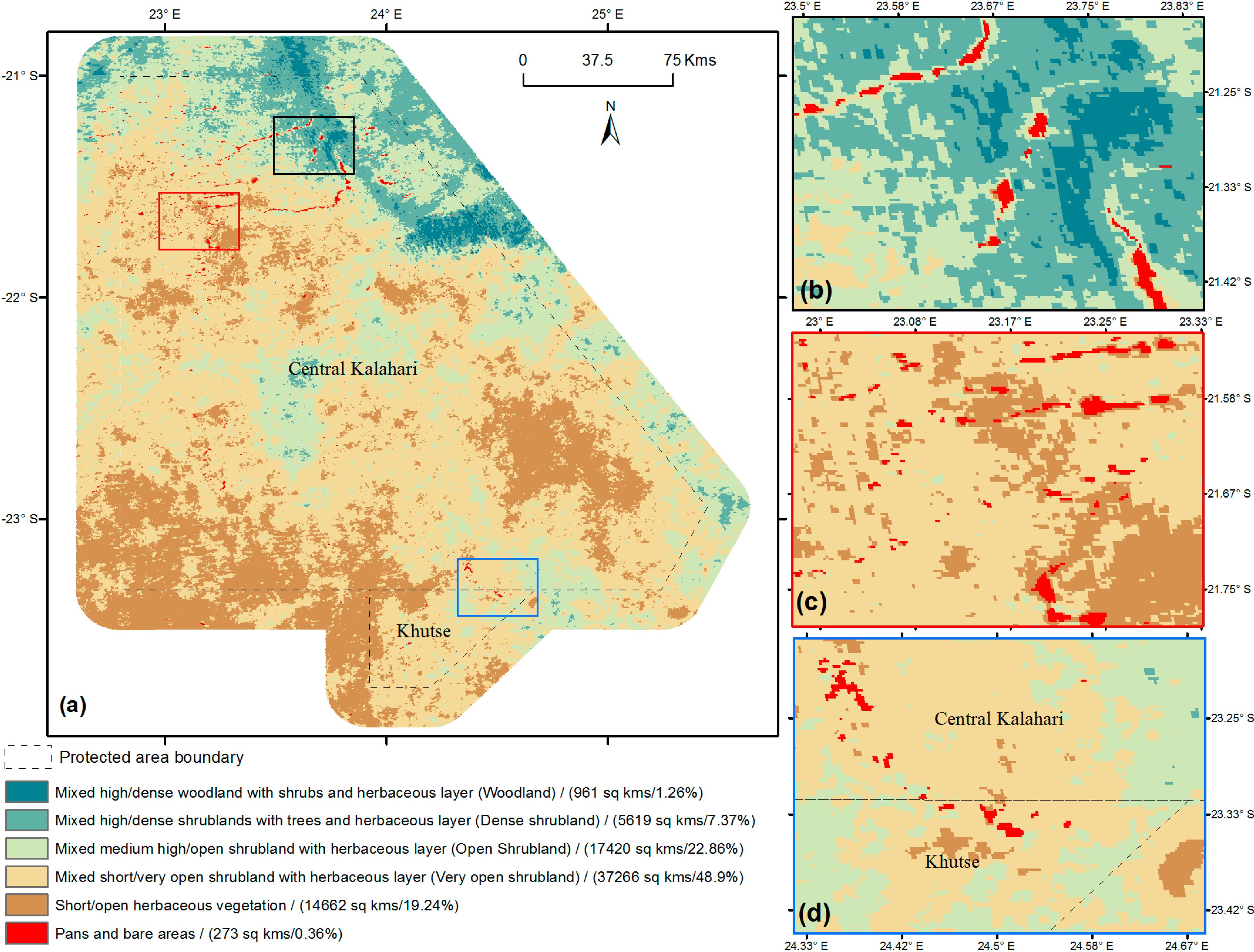
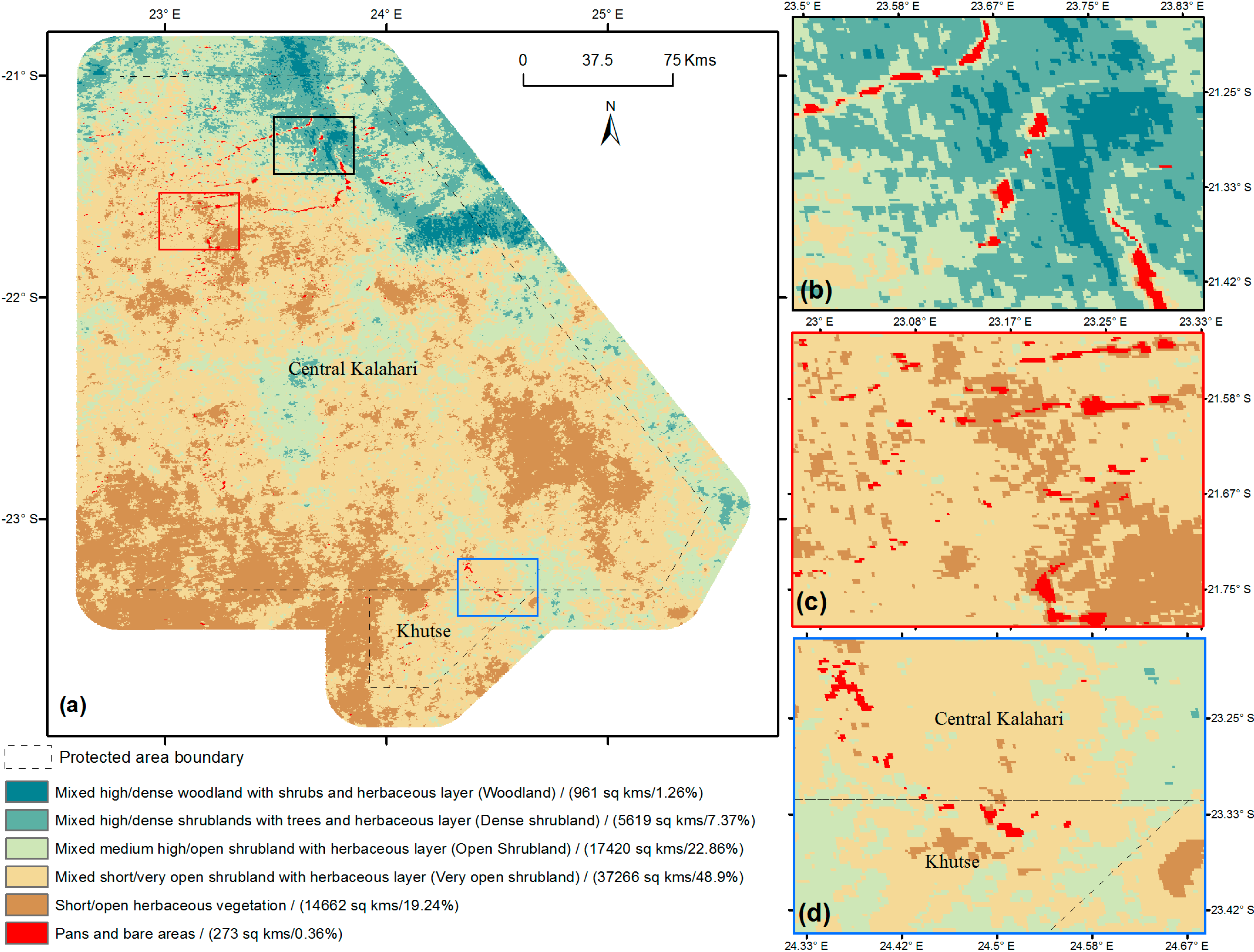{kind=link}
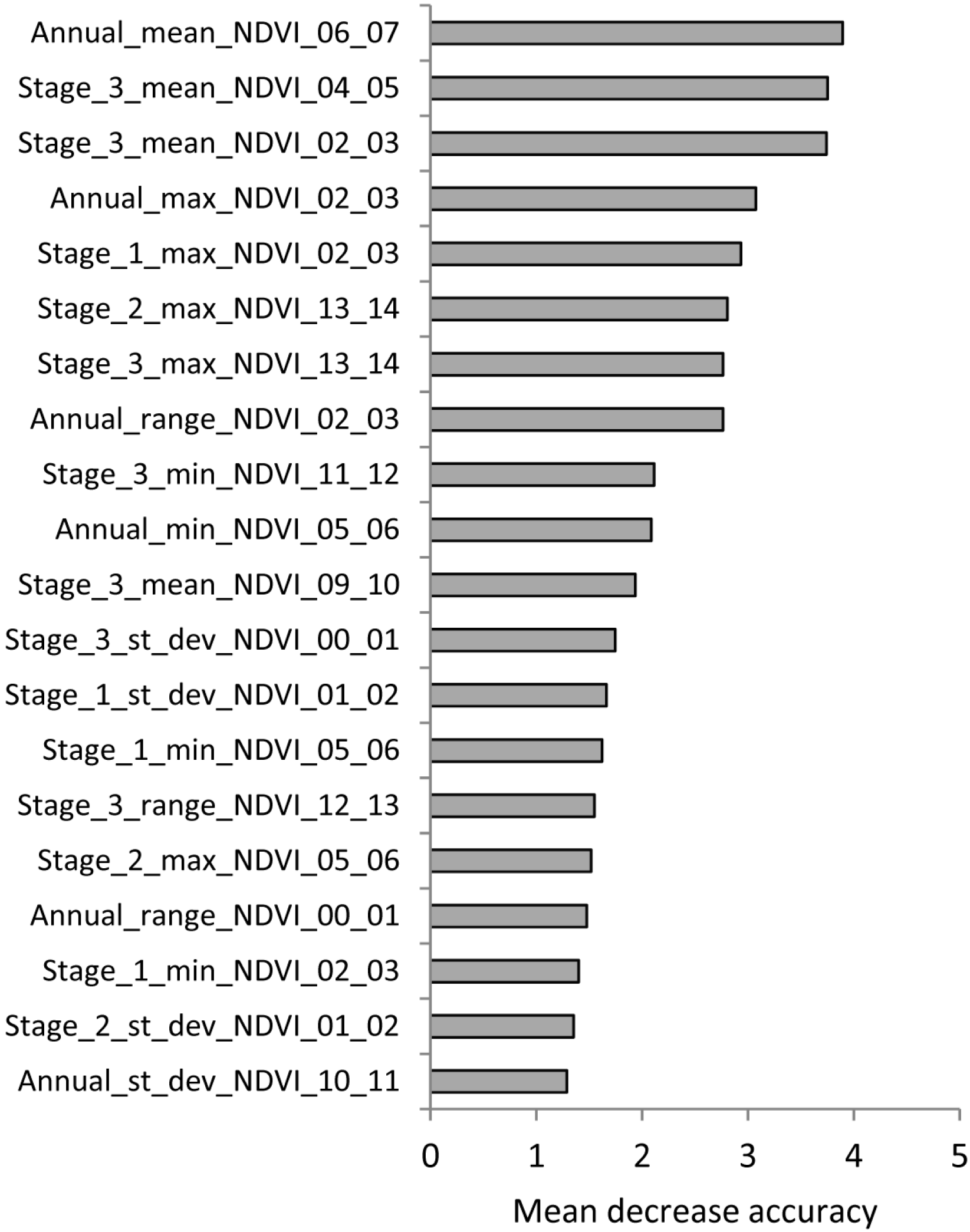
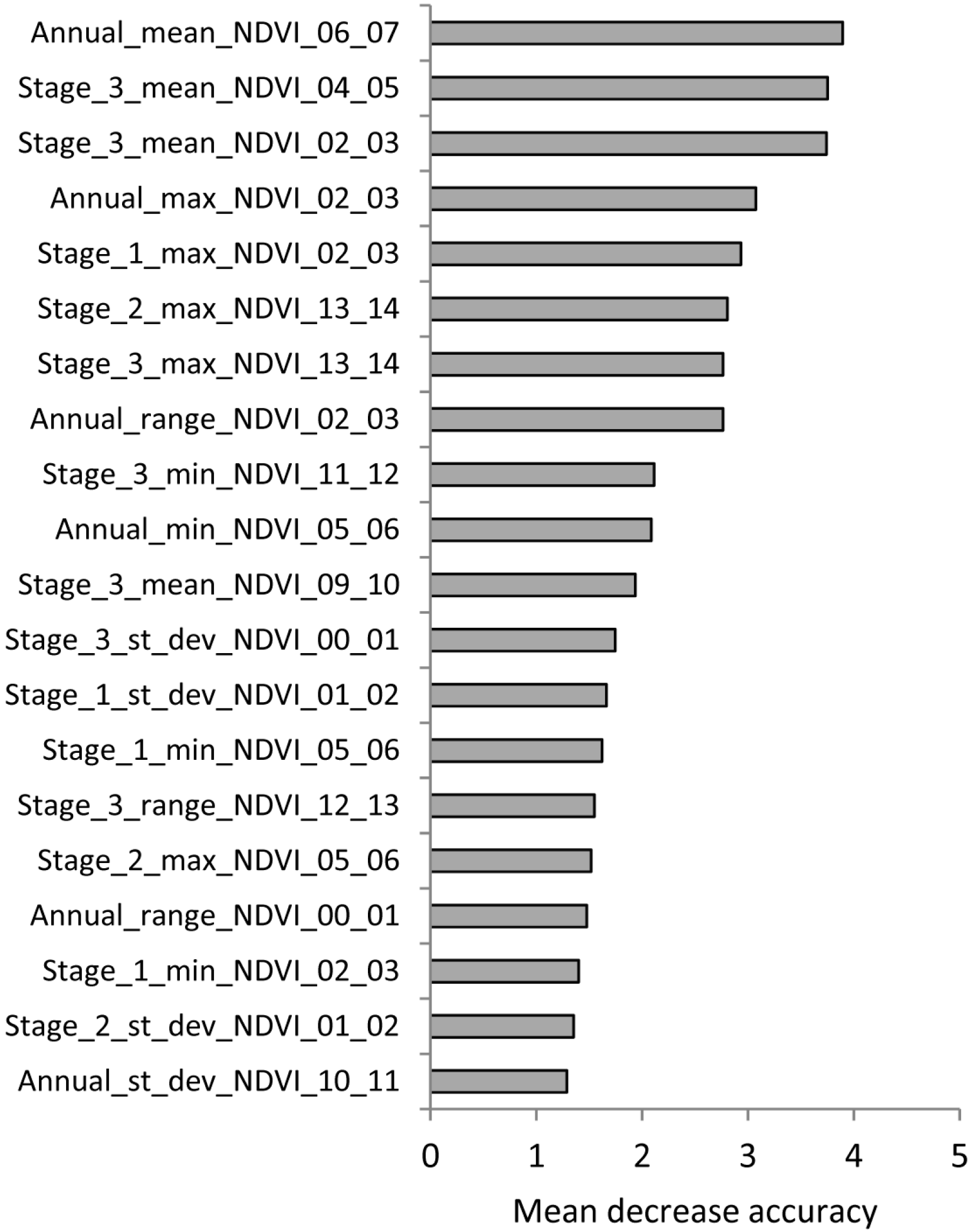{kind=link}
| H_De_Wd (Woodland) | H_De_Sh (Dense Shrubland) | Mh_Op_Sh (Open Shrubland) | Sh_Vop_Sh (Very Open Shrubland) | Sh_Vop_He (Very Open Herbaceous Vegetation) | Pans | |
|---|---|---|---|---|---|---|
| H_De_Wd (Woodland) | -- | 1.88 | 1.91 | 1.99 | 1.99 | 1.99 |
| H_De_Sh (Dense shrubland) | -- | 1.86 | 1.88 | 1.99 | 1.99 | |
| Mh_Op_Sh (Open shrubland) | -- | 1.91 | 1.95 | 1.99 | ||
| Sh_Vop_Sh (Very open shrubland) | -- | 1.92 | 1.99 | |||
| Sh_Vop_He (Very open herbaceous vegetation) | -- | 1.96 | ||||
| Pans | -- |
| Class Name | H_De_Wd (Woodland) | H_De_Sh (Dense Shrubland) | Mh_Op_Sh (Open Shrubland) | Sh_Vop_Sh (Very Open Shrubland) | Sh_Vop_He (Very Open Herbaceous Vegetation) | Pans | Producer’s Accuracy (%) |
|---|---|---|---|---|---|---|---|
| H_De_Wd (Woodland) | 79 | 4 | 3 | - | - | - | 91.86 |
| H_De_Sh (Dense shrubland) | 14 | 201 | 5 | 2 | - | - | 90.54 |
| Mh_Op_Sh (Open shrubland) | 1 | 14 | 336 | 10 | 2 | - | 92.56 |
| Sh_Vop_Sh (Very open shrubland) | - | 22 | 10 | 429 | 5 | - | 92.06 |
| Sh_Vop_He (Very open herbaceous vegetation) | - | - | 8 | 17 | 268 | 3 | 90.54 |
| Pans | - | - | - | - | 3 | 94 | 95.91 |
| User’s accuracy (%) | 84.04 | 83.4 | 92.81 | 93.66 | 95.37 | 96.9 |
5. Summary
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Balmford, A.; Moore, J.L.; Brooks, T.; Burgess, N.; Hansen, L.A.; Williams, P.; Rahbek, C. Conservation conflicts across africa. Science 2001, 291, 2616–2619. [Google Scholar] [CrossRef] [PubMed]
- Von Richter, W. The status of wildlife conservation in botswana. Koedoe 2014, 20, 143. [Google Scholar]
- Thomas, D.S.G.; Twyman, C. Good or bad rangeland? Hybrid knowledge, science, and local understandings of vegetation dynamics in the Kalahari. Land Degrad. Dev. 2004, 15, 215–231. [Google Scholar] [CrossRef]
- Winterbach, H.E.; Winterbach, C.W.; Somers, M.J. Landscape suitability in botswana for the conservation of its six large African carnivores. PLoS One 2014, 9, e100202. [Google Scholar] [CrossRef] [PubMed]
- Dougill, A.J.; Thomas, D.S.G.; Heathwaite, A.L. Environmental change in the kalahari: Integrated land degradation studies for nonequilibrium dryland environments. Ann. Assoc. Am. Geog. 1999, 89, 420–442. [Google Scholar] [CrossRef]
- Ferguson, K.; Hanks, J. The effects of protected area and veterinary fencing on wildlife conservation in southern Africa. PARKS 2012, 18, 49. [Google Scholar]
- Deaprtment of Wildlife and National Park (DWNP), Government of Botswana. Central Kalahari and Kutse Game Reserve Management Plan; DWNP: Gaborone, Botswana, 2003.
- Kesch, K.M.; Bauer, D.T.; Loveridge, A.J. Undermining game fences: Who is digging holes in Kalahari sands? Afr. J. Ecol. 2014, 52, 144–150. [Google Scholar] [CrossRef]
- Vanderpost, C.; Ringrose, S.; Matheson, W.; Arntzen, J. Satellite based long-term assessment of rangeland condition in semi-arid areas: An example from Botswana. J. Arid Environ. 2011, 75, 383–389. [Google Scholar] [CrossRef]
- Ringrose, S.; Matheson, W.; Wolski, P.; Huntsman-Mapila, P. Vegetation cover trends along the Botswana Kalahari transect. J. Arid Environ. 2003, 54, 297–317. [Google Scholar] [CrossRef]
- Moleele, N.; Ringrose, S.; Matheson, W.; Vanderpost, C. More woody plants? The status of bush encroachment in Botswana’S grazing areas. J. Environ. Manag. 2002, 64, 3–11. [Google Scholar] [CrossRef]
- Solomon, S.D.; Qin, M.; Manning, Z.; Chen, M.; Marquis, K.B.; Averyt, M.; Tignor, H.L. Climate Change: The Physical Science Basis, Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge Univeristy Press: Cambridge, UK, 2007. [Google Scholar]
- Thomas, D.S.G.; Knight, M.; Wiggs, G.F.S. Remobilization of southern African desert dune systems by Twenty-First Century global warming. Nature 2005, 435, 1218–1221. [Google Scholar] [CrossRef] [PubMed]
- Scholes, R.J.; Walker, B.H. Cambridge Studies in Applied Ecology and Resource Management—An African Savanna: Synthesis of the Nylsvley Study; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Van Rooyen, N.; Bezuidenhout, D.; Theron, G.K.; Bothma, J.D.P. Monitoring of the vegetation around artificial watering points (windmills) in the Kalahari Gemsbok National Park. Koedoe 1990, 33, 63–88. [Google Scholar] [CrossRef]
- Loarie, S.R.; Tambling, C.J.; Asner, G.P. Lion hunting behaviour and vegetation structure in an African savanna. Anim. Behav. 2013, 85, 899–906. [Google Scholar] [CrossRef]
- Pettorelli, N.; Laurance, W.F.; O’Brien, T.G.; Wegmann, M.; Nagendra, H.; Turner, W. Satellite remote sensing for applied ecologists: Opportunities and challenges. J. Appl. Ecol. 2014, 51, 839–848. [Google Scholar] [CrossRef]
- Huttich, C.; Gessner, U.; Herold, M.; Strohbach, B.J.; Schmidt, M.; Keil, M.; Dech, S. On the suitability of MODIS time series metrics to map vegetation types in dry savanna ecosystems: A case study in the Kalahari of Ne Namibia. Remote Sens. 2009, 1, 620–643. [Google Scholar] [CrossRef]
- Thomas, D.S.G. Sand, grass, thorns, and ... cattle: The modern Kalahari environment. In Sustainable Livelihoods in Kalahari. Environments: Contributions to Global Debates; Sporton, D., Thomas, D.S.G., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 39–66. [Google Scholar]
- Mishra, N.B. Characterizing Ecosystem Structural and Functional Properties in the Central Kalahari Using Multi-Scale Remote Sensing. Docter’s Thesis, University of Texas, Austin, TX, USA, 2014. [Google Scholar]
- Ringrose, S.; Jellema, A.; Huntsman-Mapila, P.; Baker, L.; Brubaker, K. Use of remotely sensed data in the analysis of soil-vegetation changes along a drying gradient peripheral to the Okavango Delta, Botswana. Int. J. Remote Sens. 2005, 26, 4293–4319. [Google Scholar] [CrossRef]
- Mishra, N.B.; Crews, K.A. Mapping vegetation morphology types in a dry savanna ecosystem: Integrating hierarchical object-based image analysis with random forest. Int. J. Remote Sens. 2014, 35, 1175–1198. [Google Scholar] [CrossRef]
- Bontemps, S.; Defourny, P.; Bogaert, E.V.; Arino, O.; Kalogirou, V.; Perez, J.R. Globcover 2009-Products Description and Validation Report. 2011. Available online: http://dup.esrin.esa.int/files/p68/GLOBCOVER2009_Validation_Report_2.2.pdf (accessed on 3 September 2014).
- Hill, M.J.; Román, M.O.; Schaaf, C.B. Dynamics of vegetation indices in tropical and subtropical savannas defined by ecoregions and moderate resolution imaging spectroradiometer (MODIS) land cover. Geocarto Int. 2012, 27, 153–191. [Google Scholar] [CrossRef]
- Wu, J.; Marceau, D. Modeling complex ecological systems: An introduction. Ecol. Model. 2002, 153, 1–6. [Google Scholar] [CrossRef]
- Elmore, A.J.; Mustard, J.F.; Manning, S.J.; Lobell, D.B. Quantifying vegetation change in semiarid environments: Precision and accuracy of spectral mixture analysis and the normalized difference vegetation index. Remote Sens. Environ. 2000, 73, 87–102. [Google Scholar] [CrossRef]
- Jensen, J.R. Remote Sensing of the Environment: An Earth Resource Perspective 2/e; Pearson Education India: Chennai, India, 2009. [Google Scholar]
- De Fries, R.; Hansen, M.; Townshend, J.; Sohlberg, R. Global land cover classifications at 8 km spatial resolution: The use of training data derived from Landsat imagery in decision tree classifiers. Int. J. Remote Sens. 1998, 19, 3141–3168. [Google Scholar] [CrossRef]
- Hansen, M.; DeFries, R.; Townshend, J.; Carroll, M.; Dimiceli, C.; Sohlberg, R. Global percent tree cover at a spatial resolution of 500 m: First results of the MODIS vegetation continuous fields algorithm. Earth Interact. 2003, 7, 1–15. [Google Scholar] [CrossRef]
- Jacquin, A.; Sheeren, D.; Lacombe, J.-P. Vegetation cover degradation assessment in madagascar savanna based on trend analysis of MODIS NDVI time series. Int. J. Appl. Earth Observ. Geoinf. 2010, 12, S3–S10. [Google Scholar] [CrossRef]
- Okin, G.S.; Roberts, D.A. Remote sensing in arid regions: Challenges and opportunities. In Manual of Remote Sensing—Volume 4, Remote Sensing for Natural Resource Management and Environmental Monitoring, 3rd ed.; Ustin, S., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2004; pp. 111–146. [Google Scholar]
- Colditz, R.; Schmidt, M.; Conrad, C.; Hansen, M.; Dech, S. Land cover classification with coarse spatial resolution data to derive continuous and discrete maps for complex regions. Remote Sens. Environ. 2011, 115, 3264–3275. [Google Scholar] [CrossRef]
- Friedl, M.; Zhang, X.; Strahler, A. Characterizing global land cover type and seasonal land cover dynamics at moderate spatial resolution with MODIS data. In Land Remote Sensing and Global Environmental Change; Springer: Berlin, Germany, 2011; pp. 709–724. [Google Scholar]
- Clark, M.L.; Aide, T.M.; Grau, H.R.; Riner, G. A scalable approach to mapping annual land cover at 250 m using MODIS time series data: A case study in the dry chaco ecoregion of South America. Remote Sens. Environ. 2010, 114, 2816–2832. [Google Scholar] [CrossRef]
- Klein, I.; Gessner, U.; Kuenzer, C. Regional land cover mapping and change detection in central asia using MODIS time-series. Appl. Geogr. 2012, 35, 219–234. [Google Scholar] [CrossRef]
- Ghimire, B.; Rogan, J.; Galiano, V.R.; Panday, P.; Neeti, N. An evaluation of bagging, boosting, and random forests for land-cover classification in Cape Cod, Massachusetts, USA. GISci. Remote Sens. 2012, 49, 623–643. [Google Scholar] [CrossRef]
- Waske, B.; Benediktsson, J.A.; Sveinsson, J.R. Chapter 18 Random forest classification of remote sensing data. In Signal Image Processing for Remote Sensing; CRC Press: Boca Raton, FL, USA, 2012; pp. 365–374. [Google Scholar]
- Verikas, A.; Gelzinis, A.; Bacauskiene, M. Mining data with random forests: A survey and results of new tests. Pattern Recognit. 2011, 44, 330–349. [Google Scholar] [CrossRef]
- Mendelsohn, J.; El Obeid, S. Okavango River: The Flow of A Lifeline; Struik Publishers: Cape Town, South Africa, 2004. [Google Scholar]
- Scholes, R.J.; Dowty, P.R.; Caylor, K.; Parsons, D.A.B.; Frost, P.G.H.; Shugart, H.H. Trends in savanna structure and composition along an aridity gradient in the Kalahari. J. Veg. Sci. 2002, 13, 419–428. [Google Scholar] [CrossRef]
- Makhabu, S.W.; Marotsi, B.; Perkins, J. Vegetation gradients around artificial water points in the central Kalahari game reserve of Botswana. Afr. J. Ecol. 2002, 40, 103–109. [Google Scholar] [CrossRef]
- Thomas, D.S.G.; Shaw, P.A. The Kalahari Environment; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Fagan, W.F. Connectivity, fragmentation, and extinction risk in dendritic metapopulations. Ecology 2002, 83, 3243–3249. [Google Scholar] [CrossRef]
- Mishra, N.B.; Crews, K.A. Estimating fractional land cover in semi-arid central kalahari: The impact of mapping method (spectral unmixing versus object based image analysis) and vegetation morphology. Geocarto Int. 2014, 29, 860–877. [Google Scholar] [CrossRef]
- Thompson, M. Standard land-cover classification scheme for remote-sensing applications in South Africa. South. Afr. J. Sci. 1996, 29, 36–42. [Google Scholar]
- Groffman, P.M.; Tiedje, T.M.; Mokma, D.L.; Simkins, S. Regional-scale analysis of denitrification in north temperate forest soils. Landsc. Ecol. 1992, 7, 45–53. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. Timesat—A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Laliberte, A.; Browning, D.; Rango, A. A comparison of three feature selection methods for object-based classification of sub-decimeter resolution UltraCam-l imagery. Int. J. Appl. Earth Observ. Geoinf. 2012, 15, 70–78. [Google Scholar] [CrossRef]
- Friedl, M.A.; Brodley, C.E. Decision tree classification of land cover from remotely sensed data. Remote Sens. Environ. 1997, 61, 399–409. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Liaw, A.; Weiener, M. Classification and regression by randomforest. R News 2002, 2, 18–22. [Google Scholar]
- Levick, S.R.; Asner, G.P.; Kennedy-Bowdoin, T.; Knapp, D.E. The relative influence of fire and herbivory on savanna three-dimensional vegetation structure. Biol. Conserv. 2009, 142, 1693–1700. [Google Scholar] [CrossRef]
- Sankaran, M.; Ratnam, J.; Hanan, N. Woody cover in African savannas: The role of resources, fire and herbivory. Glob. Ecol. Biogeogr. 2008, 17, 236–245. [Google Scholar] [CrossRef]
- Vanacker, V.; Linderman, M.; Lupo, F.; Flasse, S.; Lambin, E. Impact of short-term rainfall fluctuation on interannual land cover change in Sub-Saharan Africa. Glob. Ecol. Biogeogr. 2005, 14, 123–135. [Google Scholar] [CrossRef]
- Archibald, S.; Scholes, R. Leaf green-up in a semi-arid African savanna-separating tree and grass responses to environmental cues. J. Veg. Sci. 2007, 18, 583–594. [Google Scholar]
- Gessner, U.; Machwitz, M.; Conrad, C.; Dech, S. Estimating the fractional cover of growth forms and bare surface in savannas. A multi-resolution approach based on regression tree ensembles. Remote Sens. Environ. 2013, 129, 90–102. [Google Scholar] [CrossRef]
- Mishra, N.B.; Crews, K.A.; Okin, G.S. Relating spatial patterns of fractional land cover to savanna vegetation morphology using multi-scale remote sensing in the central Kalahari. Int. J. Remote Sens. 2014, 35, 2082–2104. [Google Scholar] [CrossRef]
- Guerschman, J.P.; Hill, M.J.; Renzullo, L.J.; Barrett, D.J.; Marks, A.S.; Botha, E.J. Estimating fractional cover of photosynthetic vegetation, non-photosynthetic vegetation and bare soil in the Australian tropical savanna region upscaling the EO-1 hyperion and MODIS sensors. Remote Sens. Environ. 2009, 113, 928–945. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, N.B.; Crews, K.A.; Miller, J.A.; Meyer, T. Mapping Vegetation Morphology Types in Southern Africa Savanna Using MODIS Time-Series Metrics: A Case Study of Central Kalahari, Botswana. Land 2015, 4, 197-215. https://doi.org/10.3390/land4010197
Mishra NB, Crews KA, Miller JA, Meyer T. Mapping Vegetation Morphology Types in Southern Africa Savanna Using MODIS Time-Series Metrics: A Case Study of Central Kalahari, Botswana. Land. 2015; 4(1):197-215. https://doi.org/10.3390/land4010197
Chicago/Turabian StyleMishra, Niti B., Kelley A. Crews, Jennifer A. Miller, and Thoralf Meyer. 2015. "Mapping Vegetation Morphology Types in Southern Africa Savanna Using MODIS Time-Series Metrics: A Case Study of Central Kalahari, Botswana" Land 4, no. 1: 197-215. https://doi.org/10.3390/land4010197
APA StyleMishra, N. B., Crews, K. A., Miller, J. A., & Meyer, T. (2015). Mapping Vegetation Morphology Types in Southern Africa Savanna Using MODIS Time-Series Metrics: A Case Study of Central Kalahari, Botswana. Land, 4(1), 197-215. https://doi.org/10.3390/land4010197