
Quantitative Analysis and Nonlinear Response of Vegetation Dynamic to Driving Factors in Arid and Semi-Arid Regions of China


Abstract
1. Introduction
2. Materials and Methods
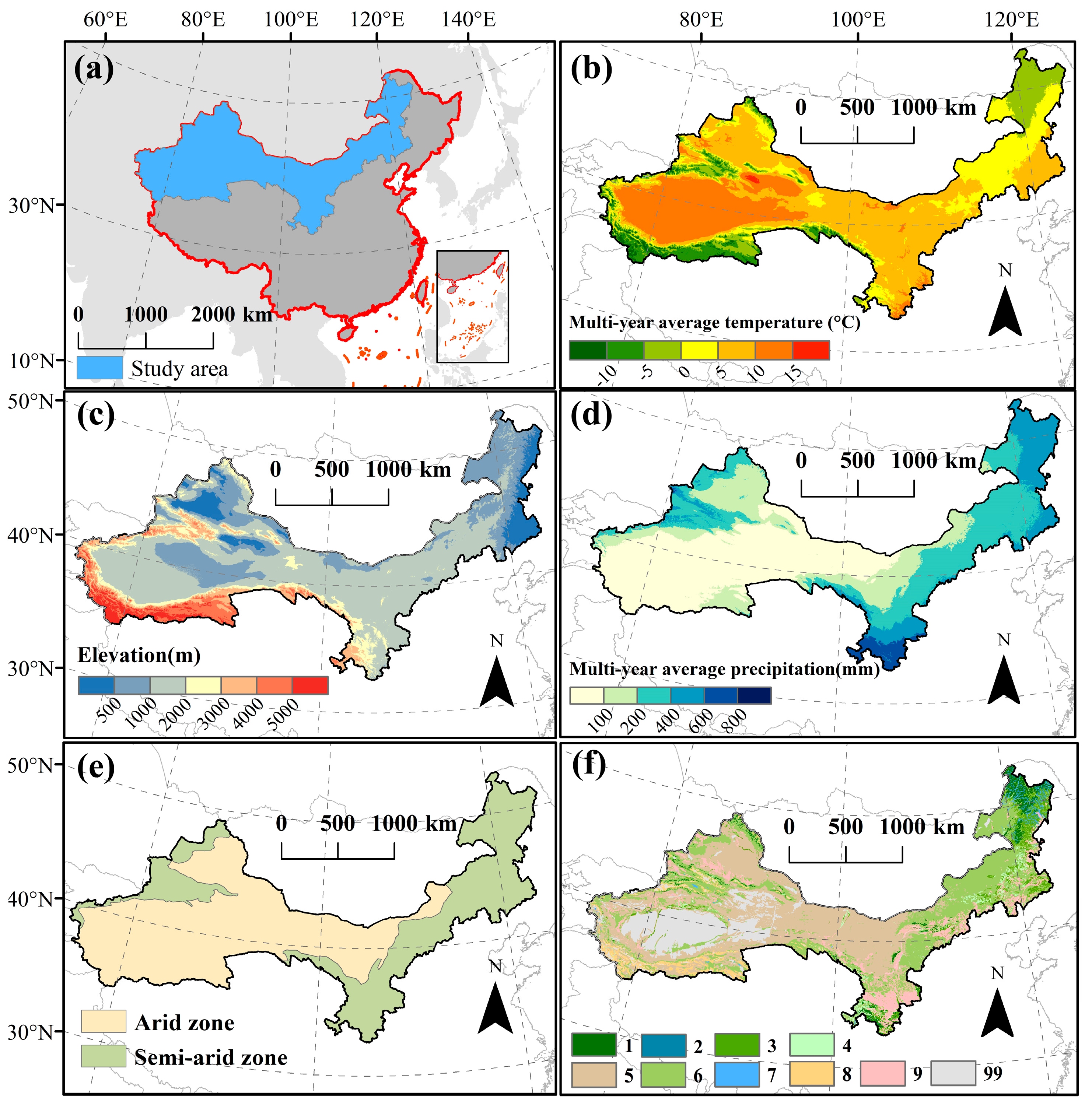
2.1. Study Area
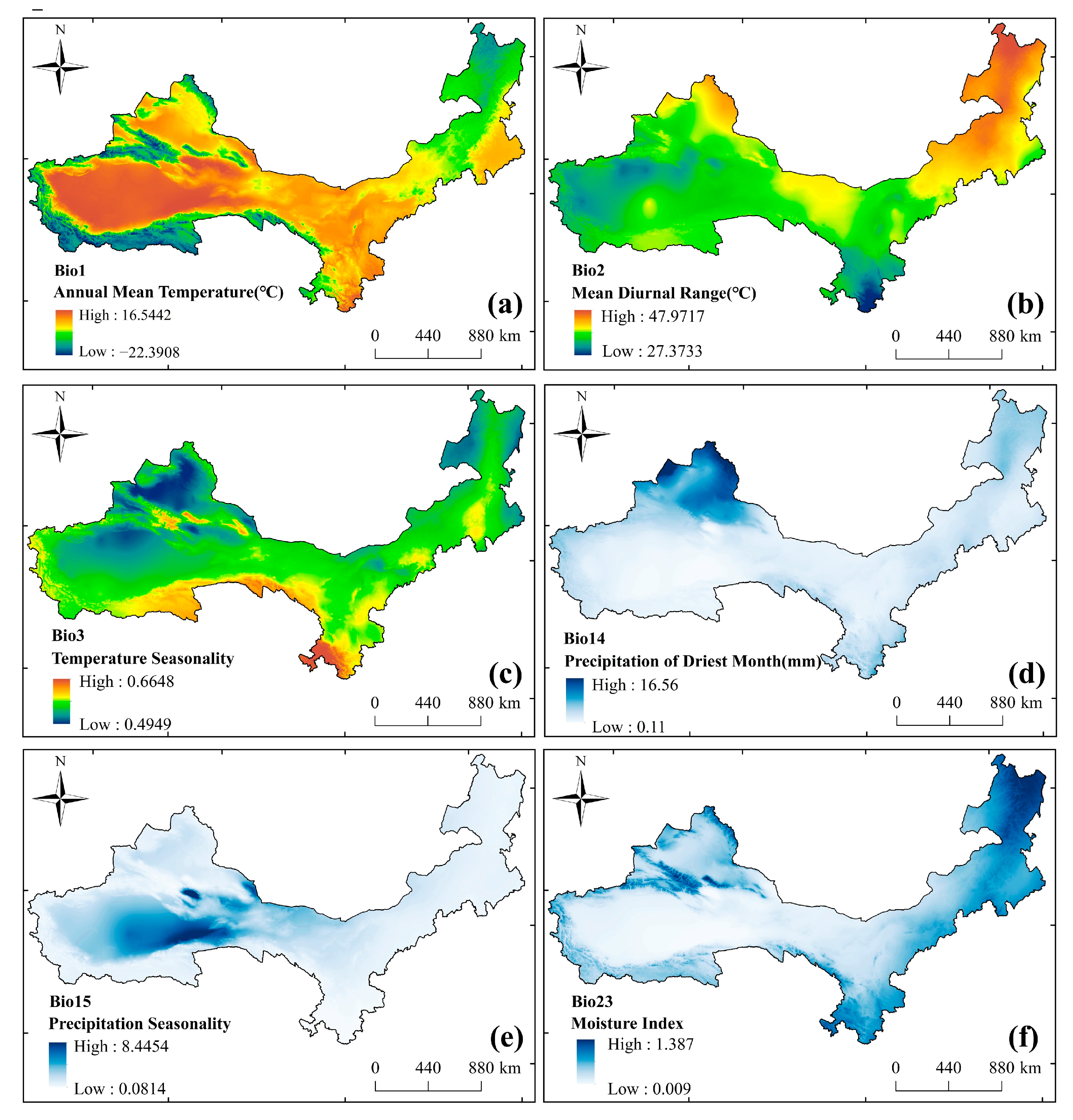
2.2. Data Sources
2.3. Methodology
2.3.1. Calculation of kNDVI
2.3.2. MK and Sen Trend Analysis
2.3.3. XGBoost
2.3.4. SHAP
3. Results
3.1. Spatial and Temporal Characteristics of kNDVI
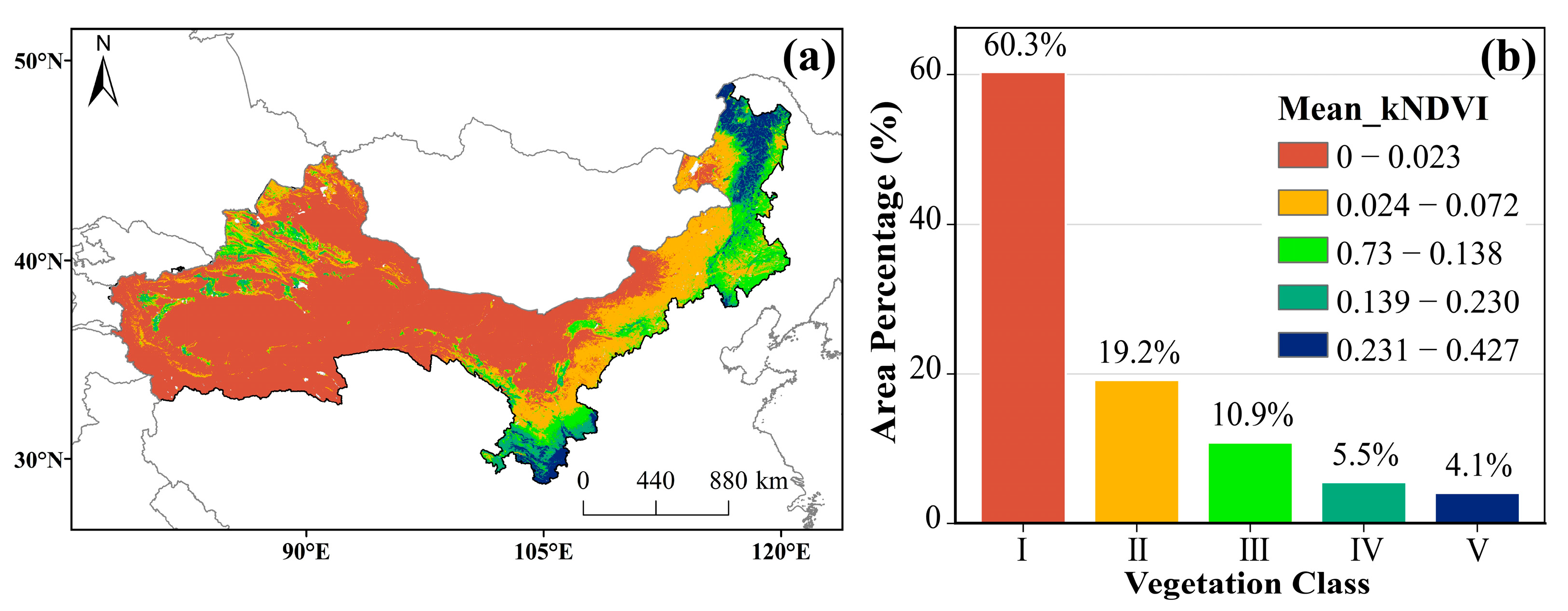
3.1.1. Spatial Distribution Characteristics of kNDVI
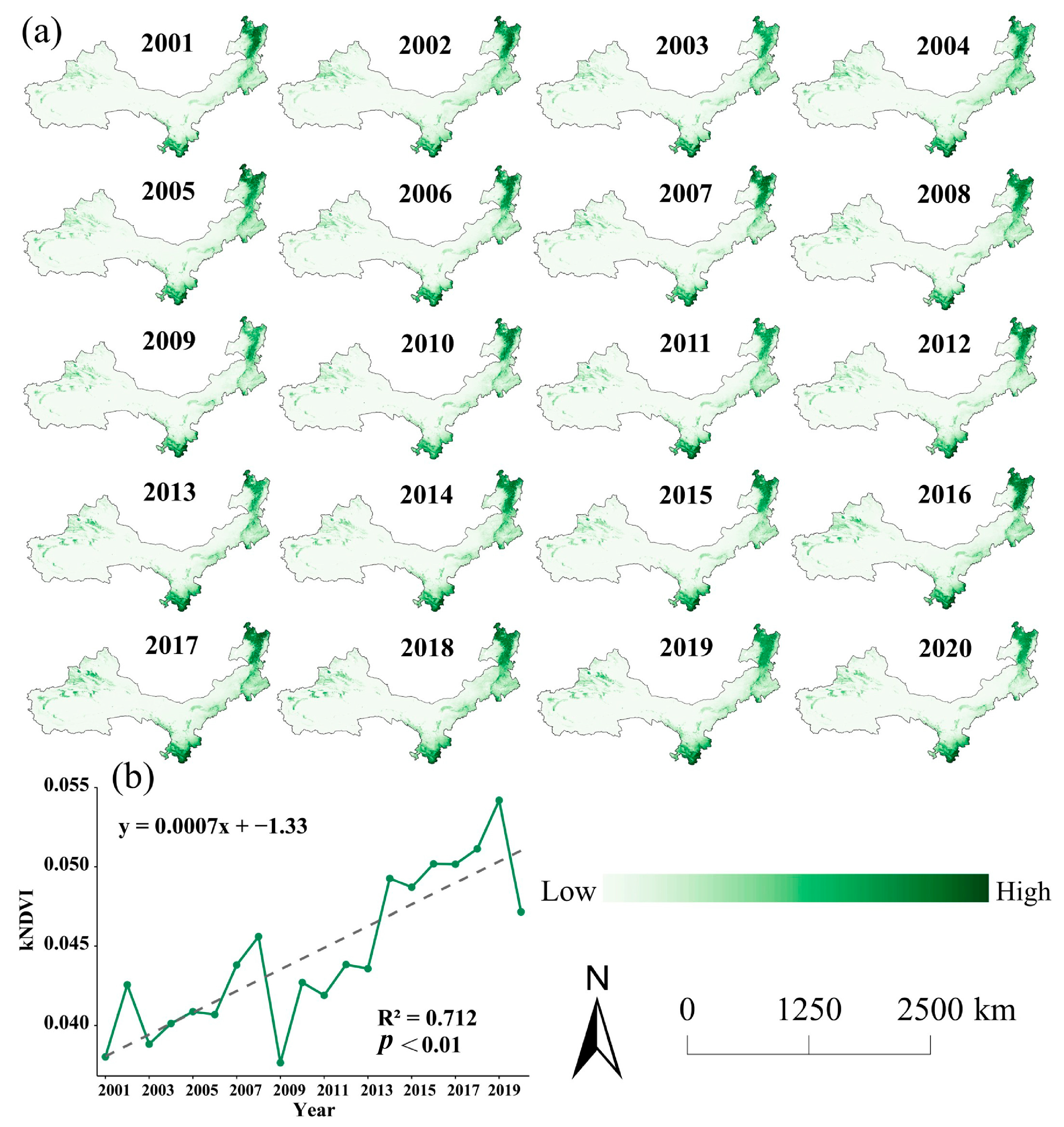
3.1.2. Trends in Spatial and Temporal Patterns of kNDVI
3.1.3. Spatial Distribution of kNDVI Trends
3.2. Drivers of Vegetation Dynamics Change in the ASAR
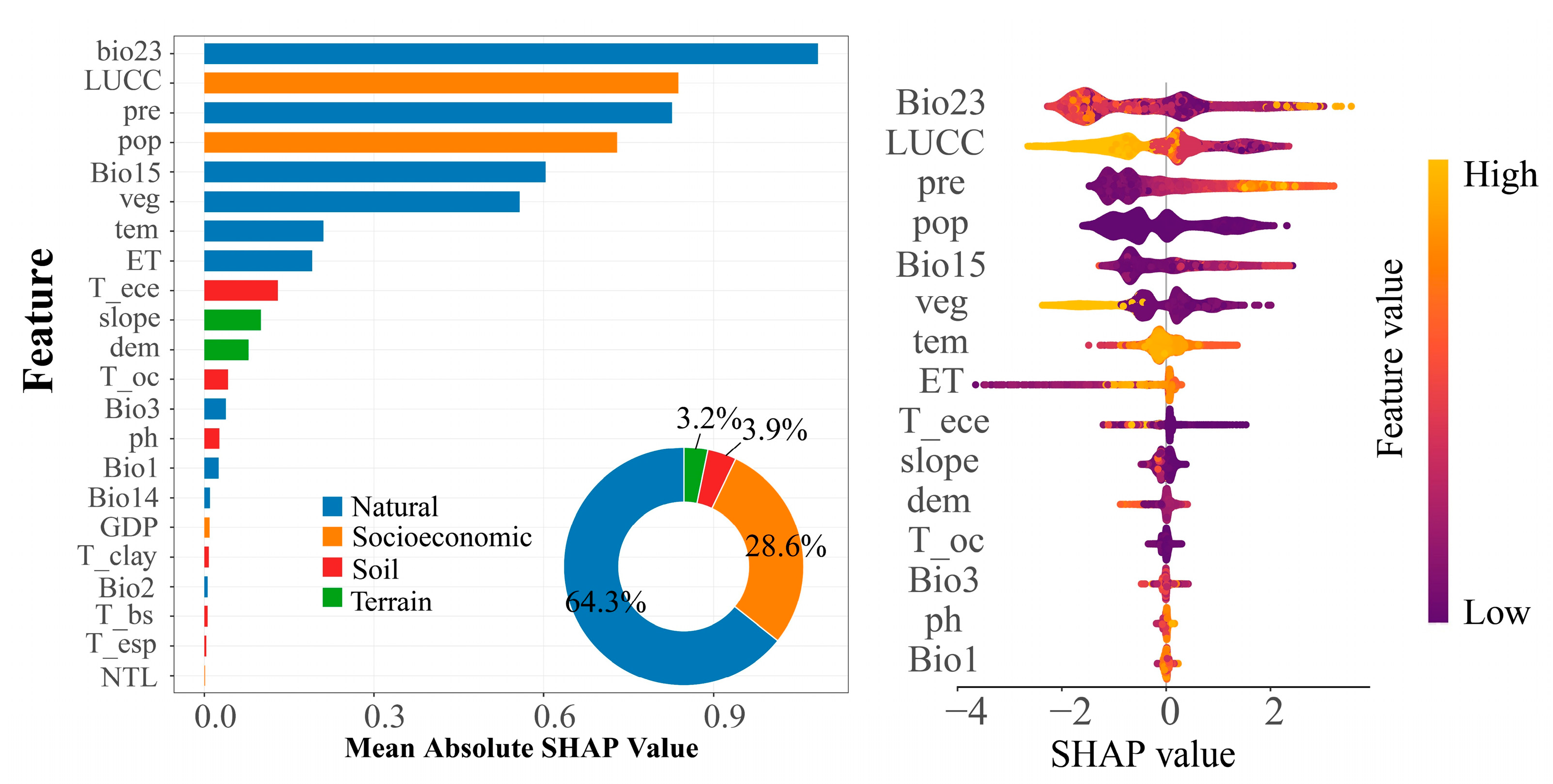
3.2.1. Degree of Relative Contribution of Drivers to kNDVI
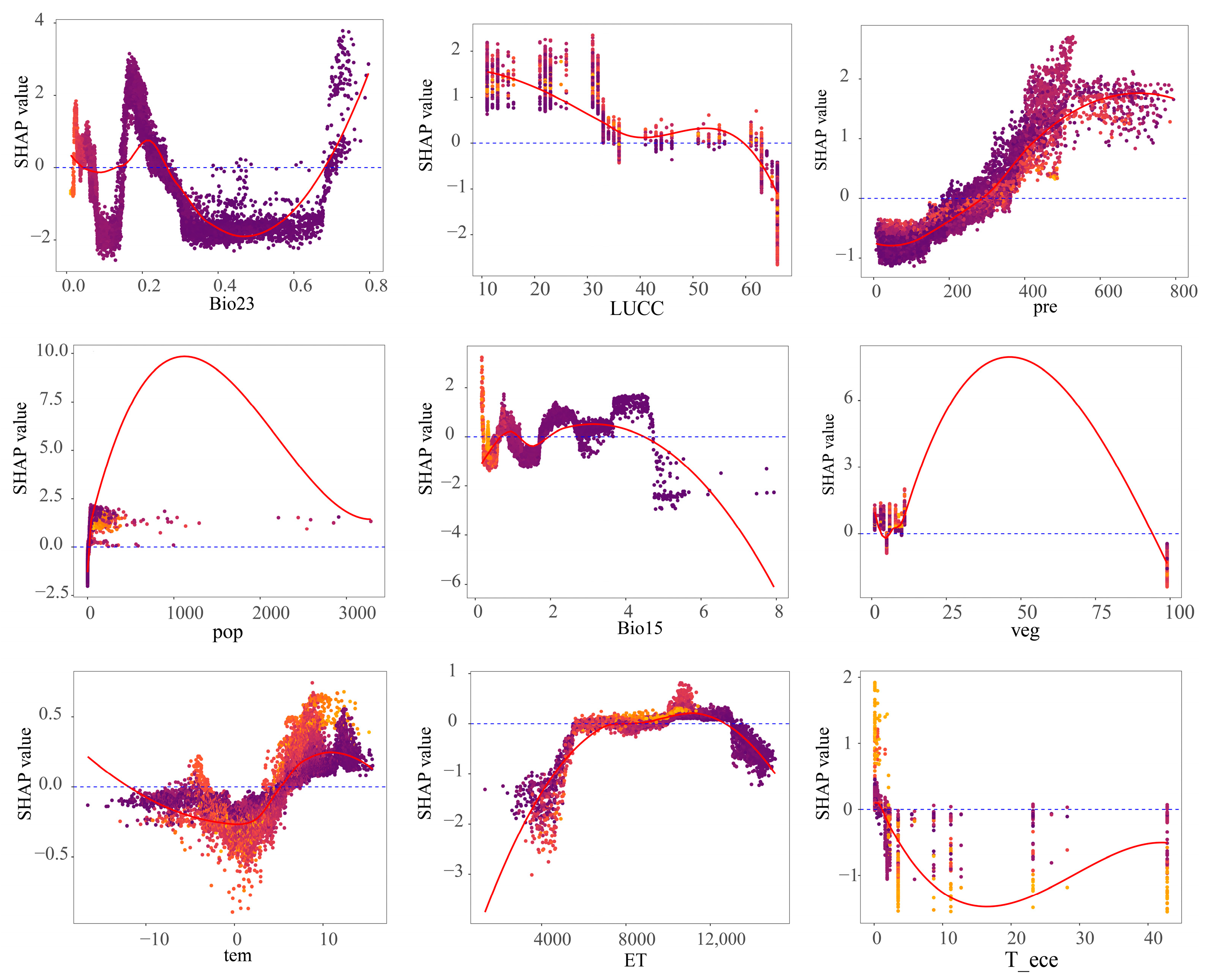
3.2.2. Nonlinear Mechanisms of Action of Vegetation Dynamics Change Drivers
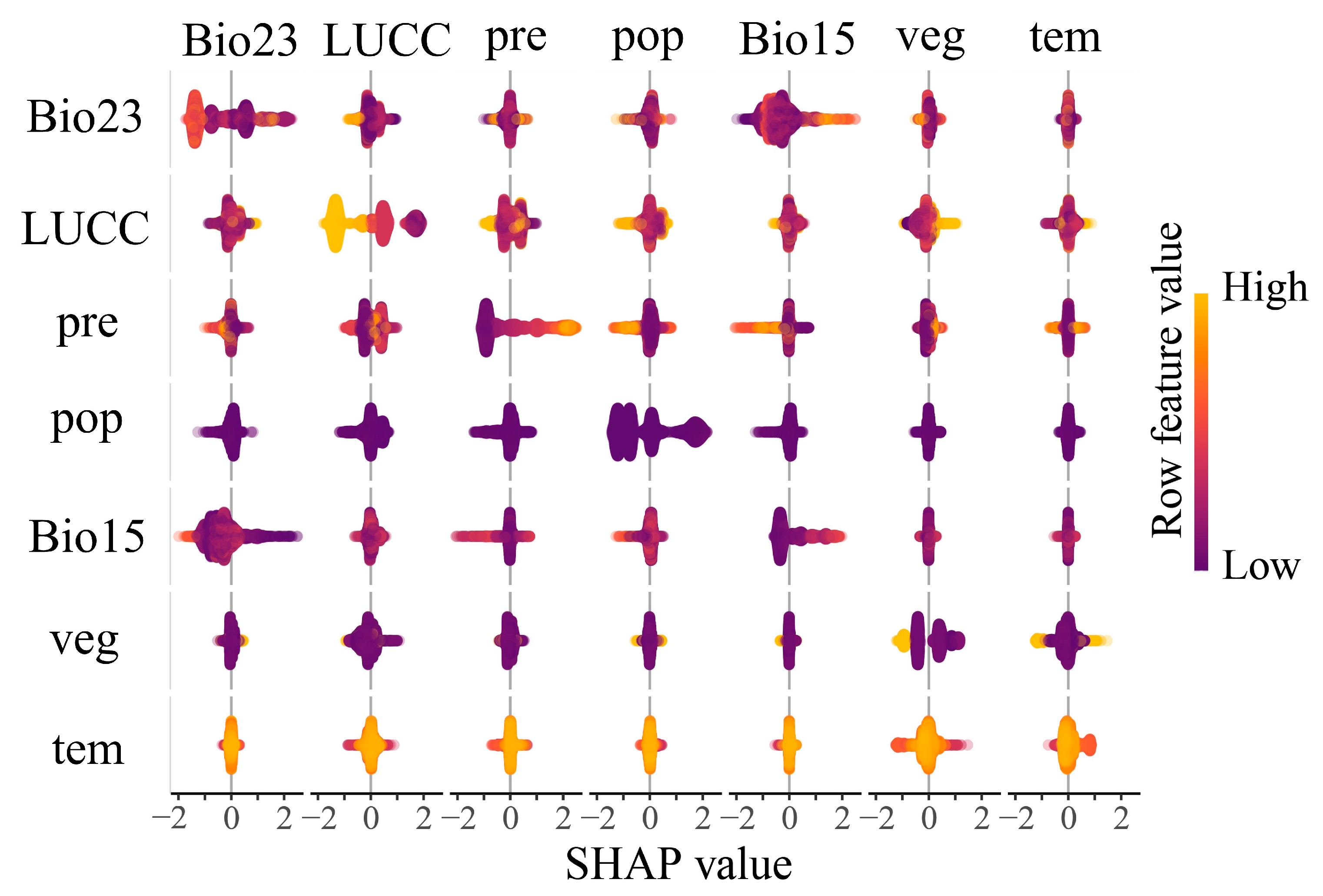
3.2.3. Interaction Mechanisms of kNDVI Drivers
4. Discussion
4.1. Factors Influencing Changes in kNDVI
4.2. Nonlinear Response of Vegetation Dynamics Trends to Driving Factors
4.3. Policy Implications
4.4. Limitations and Future Research
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vieira, F.R.; Christofaro, C. Contributions of the Vegetation Index (NDVI) in Water Quality Prediction Models in a Semi-Arid Tropical Watershed. J. Arid Environ. 2024, 220, 105122. [Google Scholar] [CrossRef]
- Xu, Y.; Dai, Q.-Y.; Zou, B.; Xu, M.; Feng, Y.-X. Tracing Climatic and Human Disturbance in Diverse Vegetation Zones in China: Over 20 Years of NDVI Observations. Ecol. Indic. 2023, 156, 111170. [Google Scholar] [CrossRef]
- Nolan, C.; Overpeck, J.T.; Allen, J.R.M.; Anderson, P.M.; Betancourt, J.L.; Binney, H.A.; Brewer, S.; Bush, M.B.; Chase, B.M.; Cheddadi, R.; et al. Past and Future Global Transformation of Terrestrial Ecosystems under Climate Change. Science 2018, 361, 920–923. [Google Scholar] [CrossRef]
- Piao, S.; Friedlingstein, P.; Ciais, P.; Zhou, L.; Chen, A. Effect of Climate and CO2 Changes on the Greening of the Northern Hemisphere over the Past Two Decades. Geophys. Res. Lett. 2006, 33, 028205. [Google Scholar] [CrossRef]
- Lin, M.; Hou, L.; Qi, Z.; Wan, L. Impacts of Climate Change and Human Activities on Vegetation NDVI in China’s Mu Us Sandy Land during 2000–2019. Ecol. Indic. 2022, 142, 109164. [Google Scholar] [CrossRef]
- Chaolin, G.U.; Liya, W.U.; Cook, I. Progress in Research on Chinese Urbanization. Front. Archit. Res. 2012, 1, 101–149. [Google Scholar] [CrossRef]
- Qi, H.; Gao, X.; Lei, J.; Meng, X.; Hu, Z. Transforming Desertification Patterns in Asia: Evaluating Trends, Drivers, and Climate Change Impacts from 1990 to 2022. Ecol. Indic. 2024, 161, 111948. [Google Scholar] [CrossRef]
- Xu, D.; Li, C.; Zhuang, D.; Pan, J. Assessment of the Relative Role of Climate Change and Human Activities in Desertification: A Review. J. Geogr. Sci. 2011, 21, 926–936. [Google Scholar] [CrossRef]
- Liu, X.; Feng, X.; Ciais, P.; Fu, B.; Hu, B.; Sun, Z. GRACE Satellite-Based Drought Index Indicating Increased Impact of Drought over Major Basins in China during 2002–2017. Agric. For. Meteorol. 2020, 291, 108057. [Google Scholar] [CrossRef]
- Yang, D.; Liu, Y. Human Activities Unevenly Disturbed Climatic Impacts on Vegetation Dynamics across Natural-Anthropogenic-Integrated Ecosystem Types in China. Habitat Int. 2025, 162, 103451. [Google Scholar] [CrossRef]
- Guo, B.; Lu, M.; Fan, Y.; Wu, H.; Yang, Y.; Wang, C. A Novel Remote Sensing Monitoring Index of Salinization Based on Three-Dimensional Feature Space Model and Its Application in the Yellow River Delta of China. Geomat. Nat. Hazards Risk 2023, 14, 95–116. [Google Scholar] [CrossRef]
- He, J.; Zhou, W.; Qian, M.; Zha, E.; Cao, A.; Shi, X. Unveiling Differentiation Characteristics of Vegetation Restoration Potential for Browning Areas in China’s Hilly and Gully Region. Land 2025, 14, 52. [Google Scholar] [CrossRef]
- Chen, T.; Bao, A.; Jiapaer, G.; Guo, H.; Zheng, G.; Jiang, L.; Chang, C.; Tuerhanjiang, L. Disentangling the Relative Impacts of Climate Change and Human Activities on Arid and Semiarid Grasslands in Central Asia during 1982–2015. Sci. Total Environ. 2019, 653, 1311–1325. [Google Scholar] [CrossRef]
- Zhan, Y.; Ma, C.; Yan, Y.; Zhu, J.; Ji, Y.; Ma, C.; Luo, Y. Spatial Differentiation Characteristics of Vegetation Greening Rates and Climate Attribution in China’s Arid and Semi-Arid Regions. Glob. Ecol. Conserv. 2023, 46, e02563. [Google Scholar] [CrossRef]
- Guo, J.; Fan, L.; Feng, P.; Sun, X.; Xue, S. Response of Vegetation Evapotranspiration to Landscape Pattern Changes in an Arid Region: A Case Study of the Loess Plateau, China. CATENA 2025, 252, 108878. [Google Scholar] [CrossRef]
- Jia, Z.; Zhang, Z.; Cheng, Y.; Buhebaoyin; Borjigin, S.; Quan, Z. Grassland Biomass Spatiotemporal Patterns and Response to Climate Change in Eastern Inner Mongolia Based on XGBoost Model Estimates. Ecol. Indic. 2024, 158, 111554. [Google Scholar] [CrossRef]
- Zeng, Q.; Lu, X.; Chen, S.; Cui, X.; Zhang, H.; Zhang, Q. Comparing the Performance of Vegetation Indices for Improving Urban Vegetation GPP Estimation via Eddy Covariance Flux Data and Landsat 5/7 Data. Ecol. Inform. 2025, 86, 103023. [Google Scholar] [CrossRef]
- Gupta, R.K.; Prasad, S.; Nadham, T.S.V.; Rao, G.H. Relative Sensitivity of District Mean RVI and NDVI over an Agrometeorological Zone. Adv. Space Res. 1993, 13, 261–264. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, J.; Tong, X.; Zhang, J.; Meng, P.; Li, J.; Liu, P.; Yu, P. NIRv and SIF Better Estimate Phenology than NDVI and EVI: Effects of Spring and Autumn Phenology on Ecosystem Production of Planted Forests. Agric. For. Meteorol. 2022, 315, 108819. [Google Scholar] [CrossRef]
- Mänd, P.; Hallik, L.; Peñuelas, J.; Nilson, T.; Duce, P.; Emmett, B.A.; Beier, C.; Estiarte, M.; Garadnai, J.; Kalapos, T.; et al. Responses of the Reflectance Indices PRI and NDVI to Experimental Warming and Drought in European Shrublands along a North–South Climatic Gradient. Remote Sens. Environ. 2010, 114, 626–636. [Google Scholar] [CrossRef]
- Chen, L.; Li, Z.; Zhang, C.; Fu, X.; Ma, J.; Zhou, M.; Peng, J. Spatiotemporal Changes of Vegetation in the Northern Foothills of Qinling Mountains Based on kNDVI Considering Climate Time-Lag Effects and Human Activities. Environ. Res. 2025, 270, 120959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hao, X.; Liu, Y.; Li, X.; Liang, Q.; Sun, F.; Ci, M.; Li, Y. Vegetation Greening Does Not Significantly Enhance Ecosystem Resilience in the Northern Hemisphere. Ecol. Indic. 2025, 177, 113762. [Google Scholar] [CrossRef]
- Camps-Valls, G.; Campos-Taberner, M.; Moreno-Martínez, Á.; Walther, S.; Duveiller, G.; Cescatti, A.; Mahecha, M.D.; Muñoz-Marí, J.; García-Haro, F.J.; Guanter, L.; et al. A Unified Vegetation Index for Quantifying the Terrestrial Biosphere. Sci. Adv. 2021, 7, eabc7447. [Google Scholar] [CrossRef] [PubMed]
- Schiefer, F.; Schmidtlein, S.; Frick, A.; Frey, J.; Klinke, R.; Zielewska-Büttner, K.; Junttila, S.; Uhl, A.; Kattenborn, T. UAV-Based Reference Data for the Prediction of Fractional Cover of Standing Deadwood from Sentinel Time Series. ISPRS Open J. Photogramm. Remote Sens. 2023, 8, 100034. [Google Scholar] [CrossRef]
- Xu, Y.; Dai, Q.-Y.; Lu, Y.-G.; Zhao, C.; Huang, W.-T.; Xu, M.; Feng, Y.-X. Identification of Ecologically Sensitive Zones Affected by Climate Change and Anthropogenic Activities in Southwest China through a NDVI-Based Spatial-Temporal Model. Ecol. Indic. 2024, 158, 111482. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, R.; Jia, M.; Wang, Z.; Zhao, C.; Guo, X.; Li, H.; Xue, Z.; Zhou, C.; Xiao, X. Significant Greening of China’s Mangrove Forests during the Last Three Decades. Ecol. Indic. 2025, 176, 113632. [Google Scholar] [CrossRef]
- Wang, Z.; Fu, B.; Wu, X.; Wang, S.; Li, Y.; Zhang, L.; Chen, N.; Wu, X. Exploring the Interdependencies of Ecosystem Services and Social-Ecological Factors on the Loess Plateau through Network Analysis. Sci. Total Environ. 2025, 960, 178362. [Google Scholar] [CrossRef]
- Pang, J.; Wang, M.; Zhang, H.; Dong, L.; Li, J.; Ding, Y.; Zhu, Z.; Yan, F. Study on the Driving Mechanism of Spatio-Temporal Non-Stationarity of Vegetation Dynamics in the Taihangshan-Yanshan Region. Ecol. Indic. 2025, 170, 113084. [Google Scholar] [CrossRef]
- Zhang, Y.; He, Y.; Li, Y.; Jia, L. Spatiotemporal Variation and Driving Forces of NDVI from 1982 to 2015 in the Qinba Mountains, China. Environ. Sci. Pollut. Res. 2022, 29, 52277–52288. [Google Scholar] [CrossRef]
- Song, Y.; Xie, W.; Wu, F.; Cui, X.; Yan, X.; Song, S.; Ren, J.; Bai, H.; Zhang, Y.; Pang, W.; et al. Tree Density Has Been Underestimated in the Mountainous Regions of Northeast China. Ecol. Indic. 2025, 176, 113655. [Google Scholar] [CrossRef]
- Guo, B.; Yang, F.; Fan, Y.; Zang, W. The Dominant Driving Factors of Rocky Desertification and Their Variations in Typical Mountainous Karst Areas of Southwest China in the Context of Global Change. CATENA 2023, 220, 106674. [Google Scholar] [CrossRef]
- Li, C.; Fu, B.; Wang, S.; Stringer, L.C.; Wang, Y.; Li, Z.; Liu, Y.; Zhou, W. Drivers and Impacts of Changes in China’s Drylands. Nat. Rev. Earth Environ. 2021, 2, 858–873. [Google Scholar] [CrossRef]
- Li, Z. Extracting Spatial Effects from Machine Learning Model Using Local Interpretation Method: An Example of SHAP and XGBoost. Comput. Environ. Urban Syst. 2022, 96, 101845. [Google Scholar] [CrossRef]
- Wang, Y.; Cai, L.; Liu, G.; Lyu, Y. Spatiotemporal Variation in Net Primary Productivity and Factor Detection in Arid and Semiarid Regions of China. Ecol. Front. 2025, 16, 001. [Google Scholar] [CrossRef]
- Qi, T.; Ren, Q.; He, C.; Zhang, X. Dual Effects on Vegetation from Urban Expansion in the Drylands of Northern China: A Multiscale Investigation Using the Vegetation Disturbance Index. Sci. Total Environ. 2024, 928, 172481. [Google Scholar] [CrossRef]
- Huang, X.; Liang, S.; Ziegler, A.D.; Zeng, Z. Decoupling Vegetation and Soil-Moisture Interaction in Evapotranspiration Interannual Variability. iScience 2025, 28, 113008. [Google Scholar] [CrossRef]
- Yang, L.; Shen, F.; Zhang, L.; Cai, Y.; Yi, F.; Zhou, C. Quantifying Influences of Natural and Anthropogenic Factors on Vegetation Changes Using Structural Equation Modeling: A Case Study in Jiangsu Province, China. J. Clean. Prod. 2021, 280, 124330. [Google Scholar] [CrossRef]
- Bai, Q.; Wang, T.; Han, Q.; Li, X. Vegetation Dynamics Induced by Climate Change and Human Activities: Implications for Coastal Wetland Restoration. J. Environ. Manag. 2025, 384, 125594. [Google Scholar] [CrossRef]
- Li, J.; Li, X.; Li, D. Impact of Vegetation Phenology on Anisotropy of Artificial Light at Night—Evidence from Multi-Angle Satellite Observations. Remote Sens. Environ. 2025, 317, 114525. [Google Scholar] [CrossRef]
- Feng, X.; Tian, J.; Wang, Y.; Wu, J.; Liu, J.; Ya, Q.; Li, Z. Spatio-Temporal Variation and Climatic Driving Factors of Vegetation Coverage in the Yellow River Basin from 2001 to 2020 Based on kNDVI. Forests 2023, 14, 620. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, H.; Li, N.; Zhu, J.; Pan, Z.; Qin, F. Spatial and Temporal Characteristics and Driving Forces of Vegetation Changes in the Huaihe River Basin from 2003 to 2018. Sustainability 2020, 12, 2198. [Google Scholar] [CrossRef]
- Fan, Q.; Zhang, L.; Wu, H.; Cheng, J.; Shi, M.; Fan, Y.; Zhao, J. Vegetation Dynamics and Driving Mechanisms in the Northwest Desert Zone Based on Long-Term kNDVI Data. Ecol. Front. 2025, 45, 836–848. [Google Scholar] [CrossRef]
- Güçlü, Y.S. Multiple Şen-Innovative Trend Analyses and Partial Mann-Kendall Test. J. Hydrol. 2018, 566, 685–704. [Google Scholar] [CrossRef]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016; Association for Computing Machinery: New York, NY, USA, 2016; pp. 785–794. [Google Scholar]
- Wang, S.; Peng, H.; Hu, Q.; Jiang, M. Analysis of Runoff Generation Driving Factors Based on Hydrological Model and Interpretable Machine Learning Method. Journal of Hydrology. Reg. Stud. 2022, 42, 101139. [Google Scholar] [CrossRef]
- Ouyang, N.; Rui, X.; Zhang, X.; Tang, H.; Xie, Y. Spatiotemporal Evolution of Ecosystem Health and Its Driving Factors in the Southwestern Karst Regions of China. Ecol. Indic. 2024, 166, 112530. [Google Scholar] [CrossRef]
- Ming, L.; Wang, Y.; Liu, G.; Meng, L.; Chen, X. Analysis of Vegetation Dynamics from 2001 to 2020 in China’s Ganzhou Rare Earth Mining Area Using Time Series Remote Sensing and SHAP-Enhanced Machine Learning. Ecol. Inform. 2024, 84, 102887. [Google Scholar] [CrossRef]
- Wen, H.; Liu, B.; Di, M.; Li, J.; Zhou, X. A SHAP-Enhanced XGBoost Model for Interpretable Prediction of Coseismic Landslides. Adv. Space Res. 2024, 74, 3826–3854. [Google Scholar] [CrossRef]
- Zhang, Z.; Hu, Y.; Batunacun. Analysis of the Driving Mechanism of Grassland Degradation in Inner Mongolia Grassland from 2015 to 2020 Using Interpretable Machine Learning Methods. Land 2025, 14, 386. [Google Scholar] [CrossRef]
- Zhang, G.; Li, X.; Zhang, L.; Wei, X. Dynamics and Causes of Cropland Non-Agriculturalization in Typical Regions of China: An Explanation Based on Interpretable Machine Learning. Ecol. Indic. 2024, 166, 112348. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Liu, X.; Lian, Z.; Zhang, G.; Liu, Z.; An, S.; Ren, Y.; Li, Y.; Liu, S. Dynamic Monitoring and Drivers of Ecological Environmental Quality in the Three-North Region, China: Insights Based on Remote Sensing Ecological Index. Ecol. Inform. 2025, 85, 102936. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, J.; Zhang, Z.; Tang, H.; Liu, L.; Liu, X.; Ma, R.; Zhang, M.; Zhang, X. The Difference in Ecological Environmental Quality Impact Factors between Human Activity Zone and Non-Human Activity Zone in Arid Regions: A Case Study of the Northern Slope of the Tianshan Mountains. Ecol. Indic. 2025, 171, 113226. [Google Scholar] [CrossRef]
- Lv, X.; Chen, G.; Wang, Q.; Chen, T. Exploring the Resilience of Global Vegetation Ecosystem: Nonlinearity, Driving Forces, and Management. J. Environ. Manag. 2025, 377, 124634. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, C.; Liu, Y.; Wang, T.; Tian, Z.; Ding, L.; Liu, Z.; Feng, T.; Niu, Q.; Mao, X.; et al. The Change Pattern and Spatiotemporal Transition of Land Use Carbon Emissions in China’s Three-North Shelterbelt Program Region. Ecol. Eng. 2025, 219, 107680. [Google Scholar] [CrossRef]
- Chu, X.; Zhan, J.; Li, Z.; Zhang, F.; Qi, W. Assessment on Forest Carbon Sequestration in the Three-North Shelterbelt Program Region, China. J. Clean. Prod. 2019, 215, 382–389. [Google Scholar] [CrossRef]
- Burrell, A.L.; Evans, J.P.; De Kauwe, M.G. Anthropogenic Climate Change Has Driven over 5 Million Km2 of Drylands towards Desertification. Nat. Commun. 2020, 11, 3853. [Google Scholar] [CrossRef]
- Eastman, J.R.; Sangermano, F.; Machado, E.A.; Rogan, J.; Anyamba, A. Global Trends in Seasonality of Normalized Difference Vegetation Index (NDVI), 1982–2011. Remote Sens. 2013, 5, 4799–4818. [Google Scholar] [CrossRef]
- Xu, X.; Liu, H.; Jiao, F.; Gong, H.; Lin, Z. Nonlinear Relationship of Greening and Shifts from Greening to Browning in Vegetation with Nature and Human Factors along the Silk Road Economic Belt. Sci. Total Environ. 2021, 766, 142553. [Google Scholar] [CrossRef]
- Vagnon, C.; Olden, J.D.; Boulêtreau, S.; Bruel, R.; Chevalier, M.; Garcia, F.; Holtgrieve, G.; Jackson, M.; Thebault, E.; Tedesco, P.A.; et al. Ecosystem Synchrony: An Emerging Property to Elucidate Ecosystem Responses to Global Change. Trends Ecol. Evol. 2024, 39, 1080–1089. [Google Scholar] [CrossRef]
- Pandey, R.; Yangchen, C.; Thiyaharajan, M.; Kishwan, J. Attributable Factors for Climate Change Adaptation among Urban Informal Settlers of a Least Developed Country, Bhutan. Habitat Int. 2023, 136, 102817. [Google Scholar] [CrossRef]
- Liu, H.; Jiao, F.; Yin, J.; Li, T.; Gong, H.; Wang, Z.; Lin, Z. Nonlinear Relationship of Vegetation Greening with Nature and Human Factors and Its Forecast—A Case Study of Southwest China. Ecol. Indic. 2020, 111, 106009. [Google Scholar] [CrossRef]
- Wang, K.; Fu, C.; Zhao, C.; Zhou, T.; Lian, Y.; Wang, G. Vegetation Change Mechanism and Key Driving Factors in Complex Terrains, China. Sci. Total Environ. 2025, 979, 179424. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, W.; Wang, H.; Wang, D. Spatiotemporal Evolution and Driving Factors Analysis of Fractional Vegetation Coverage in the Arid Region of Northwest China. Sci. Total Environ. 2024, 954, 176271. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Piao, S.; Zhou, L.; He, J.; Wei, F.; Myneni, R.B.; Tucker, C.J.; Tan, K. Precipitation Patterns Alter Growth of Temperate Vegetation. Geophys. Res. Lett. 2005, 32, 024231. [Google Scholar] [CrossRef]
- Zhang, J.; Guan, Q.; Zhang, Z.; Shao, W.; Zhang, E.; Kang, T.; Xiao, X.; Liu, H.; Luo, H. Characteristics of Spatial and Temporal Dynamics of Vegetation and Its Response to Climate Extremes in Ecologically Fragile and Climate Change Sensitive Areas—A Case Study of Hexi Region. CATENA 2024, 239, 107910. [Google Scholar] [CrossRef]
- Wu, D.; Zhao, X.; Liang, S.; Zhou, T.; Huang, K.; Tang, B.; Zhao, W. Time-Lag Effects of Global Vegetation Responses to Climate Change. Glob. Change Biol. 2015, 21, 3520–3531. [Google Scholar] [CrossRef]
- Yan, W.; He, Y.; Cai, Y.; Qu, X.; Cui, X. Relationship between Extreme Climate Indices and Spatiotemporal Changes of Vegetation on Yunnan Plateau from 1982 to 2019. Glob. Ecol. Conserv. 2021, 31, e01813. [Google Scholar] [CrossRef]
- Lei, T.; Feng, J.; Lv, J.; Wang, J.; Song, H.; Song, W.; Gao, X. Net Primary Productivity Loss under Different Drought Levels in Different Grassland Ecosystems. J. Environ. Manag. 2020, 274, 111144. [Google Scholar] [CrossRef]
- Song, Y.; Penuelas, J.; Ciais, P.; Wang, S.; Zhang, Y.; Gentine, P.; McCabe, M.F.; Wang, L.; Li, X.; Li, F.; et al. Recent Water Constraints Mediate the Dominance of Climate and Atmospheric CO2 on Vegetation Growth Across China. Earth’s Future 2024, 12, e2023EF004395. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Bevacqua, E.; Zscheischler, J.; Keenan, T.F.; Lian, X.; Zhou, S.; Zhang, H.; He, M.; Piao, S. Future Increase in Compound Soil Drought-Heat Extremes Exacerbated by Vegetation Greening. Nat. Commun. 2024, 15, 10875. [Google Scholar] [CrossRef] [PubMed]
- Zhao, A.; Zhang, A.; Liu, J.; Feng, L.; Zhao, Y. Assessing the Effects of Drought and “Grain for Green” Program on Vegetation Dynamics in China’s Loess Plateau from 2000 to 2014. CATENA 2019, 175, 446–455. [Google Scholar] [CrossRef]
- Ma, L.; Ma, J.; Yan, P.; Tian, F.; Peñuelas, J.; Rao, M.P.; Fu, Y.; Hu, Z. Planted Forests in China Have Higher Drought Risk Than Natural Forests. Glob. Change Biol. 2025, 31, e70055. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.M.; Hewlett, J.D. A Review of Catchment Experiments to Determine the Effect of Vegetation Changes on Water Yield and Evapotranspiration. J. Hydrol. 1982, 55, 3–23. [Google Scholar] [CrossRef]
- Gao, L.; Fan, F.; He, Y.; Wei, C.; Xu, H.; Liu, X.; Lu, X.; Bardgett, R.D. Contrasting Responses of Plant, Soil Fungal and above-Ground Arthropod Communities to Plant Invasion across Latitudes. J. Ecol. 2024, 112, 2333–2343. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Zhang, X.; Gui, J.; Xue, J. Vegetation Variations and Its Driving Factors in the Transition Zone between Tibetan Plateau and Arid Region. Ecol. Indic. 2022, 141, 109101. [Google Scholar] [CrossRef]
- Piao, S.; Yin, G.; Tan, J.; Cheng, L.; Huang, M.; Li, Y.; Liu, R.; Mao, J.; Myneni, R.B.; Peng, S.; et al. Detection and Attribution of Vegetation Greening Trend in China over the Last 30 Years. Glob. Change Biol. 2015, 21, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Ruichen, M.; Jinxi, S.; Bin, T.; Wenjin, X.; Feihe, K.; Haotian, S.; Yuxin, L. Vegetation Variation Regulates Soil Moisture Sensitivity to Climate Change on the Loess Plateau. J. Hydrol. 2023, 617, 128763. [Google Scholar] [CrossRef]
- Li, H.; Wu, Y.; Liu, S.; Xiao, J. Regional Contributions to Interannual Variability of Net Primary Production and Climatic Attributions. Agric. For. Meteorol. 2021, 303, 108384. [Google Scholar] [CrossRef]
- Liu, J.; Wei, L.; Zheng, Z.; Du, J. Vegetation Cover Change and Its Response to Climate Extremes in the Yellow River Basin. Sci. Total Environ. 2023, 905, 167366. [Google Scholar] [CrossRef]
- Xiao, X.; Wang, Q.; Guan, Q.; Zhang, Z.; Yan, Y.; Mi, J.; Yang, E. Quantifying the Nonlinear Response of Vegetation Greening to Driving Factors in Longnan of China Based on Machine Learning Algorithm. Ecol. Indic. 2023, 151, 110277. [Google Scholar] [CrossRef]
- Li, J.; Peng, S.; Li, Z. Detecting and Attributing Vegetation Changes on China’s Loess Plateau. Agric. For. Meteorol. 2017, 247, 260–270. [Google Scholar] [CrossRef]
- Qu, S.; Liu, J.; Li, B.; Zhao, L.; Li, X.; Zhang, Z.; Yuan, M.; Niu, Z.; Lin, A. Unveiling the Driver behind China’s Greening Trend: Urban vs. Rural Areas. Environ. Res. Lett. 2023, 18, 084027. [Google Scholar] [CrossRef]
- Zhou, Y.; Yue, D.; Guo, J.; Chao, Z.; Meng, X. Assessing the Impact of Land Conversion and Management Measures on the Net Primary Productivity in the Bailong River Basin, in China. CATENA 2021, 207, 105672. [Google Scholar] [CrossRef]
- Ogutu, B.O.; D’Adamo, F.; Dash, J. Impact of Vegetation Greening on Carbon and Water Cycle in the African Sahel-Sudano-Guinean Region. Glob. Planet. Change 2021, 202, 103524. [Google Scholar] [CrossRef]
- Strain, E.M.A.; Morris, R.L.; Bishop, M.J.; Tanner, E.; Steinberg, P.; Swearer, S.E.; MacLeod, C.; Alexander, K.A. Building Blue Infrastructure: Assessing the Key Environmental Issues and Priority Areas for Ecological Engineering Initiatives in Australia’s Metropolitan Embayments. J. Environ. Manag. 2019, 230, 488–496. [Google Scholar] [CrossRef]
- Dey, D.C.; Gardiner, E.S.; Kabrick, J.M.; Stanturf, J.A.; Douglass, F.J. Innovations in Afforestation of Agricultural Bottomlands to Restore Native Forests in the Eastern USA. Scand. J. For. Res. 2010, 25, 31–42. [Google Scholar] [CrossRef]
- Monteiro, A.T.; Fava, F.; Hiltbrunner, E.; Marianna, G.D.; Bocchi, S. Assessment of Land Cover Changes and Spatial Drivers behind Loss of Permanent Meadows in the Lowlands of Italian Alps. Landsc. Urban Plan. 2011, 100, 287–294. [Google Scholar] [CrossRef]
- Hermoso, V.; Morán-Ordóñez, A.; Lanzas, M.; Brotons, L. Designing a Network of Green Infrastructure for the EU. Landsc. Urban Plan. 2020, 196, 103732. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India Lead in Greening of the World through Land-Use Management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- DeFries, R.; Nagendra, H. Ecosystem Management as a Wicked Problem. Science 2017, 356, 265–270. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Type | Time | Resolution/Data Format | Data Sources |
|---|---|---|---|
| Climate data | 2001–2020 | 1000 m, Tiff | National Qinghai-Tibet Plateau Science Data Center (https://data.tpdc.ac.cn/, accessed on 2 May 2025) |
| Remote sensing imagery | 2001–2020 | 500 m, Tiff | Earth Data (https://lpdaac.usgs.gov/, accessed on 4 May 2025) |
| Soil attribute data | 2009 | 1000 m, Tiff | National Qinghai-Tibet Plateau Science Data Center (https://data.tpdc.ac.cn/, accessed on 4 May 2025) |
| Topographic data | - | 30 m, Tiff | NASA SRTM1 v3.0 (https://www.earthdata.nasa.gov/, accessed on 6 May 2025) |
| GDP | 2000–2020 | 1000 m, Tiff | Resource and Environmental Science Data Center (https://www.resdc.cn/, accessed on 6 May 2025) |
| Population density | 2001–2023 | 1000 m, Tiff | LandScan Global (https://landscan.ornl.gov/, accessed on 6 May 2025) |
| Nighttime lighting data | 2000–2020 | 1000 m, Tiff | Resource and Environmental Science Data Center (https://www.resdc.cn/, accessed on 6 May 2025) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Yang, D.; Zhang, X.; Liu, F. Quantitative Analysis and Nonlinear Response of Vegetation Dynamic to Driving Factors in Arid and Semi-Arid Regions of China. Land 2025, 14, 1575. https://doi.org/10.3390/land14081575
Liu S, Yang D, Zhang X, Liu F. Quantitative Analysis and Nonlinear Response of Vegetation Dynamic to Driving Factors in Arid and Semi-Arid Regions of China. Land. 2025; 14(8):1575. https://doi.org/10.3390/land14081575
Chicago/Turabian StyleLiu, Shihao, Dazhi Yang, Xuyang Zhang, and Fangtian Liu. 2025. "Quantitative Analysis and Nonlinear Response of Vegetation Dynamic to Driving Factors in Arid and Semi-Arid Regions of China" Land 14, no. 8: 1575. https://doi.org/10.3390/land14081575
APA StyleLiu, S., Yang, D., Zhang, X., & Liu, F. (2025). Quantitative Analysis and Nonlinear Response of Vegetation Dynamic to Driving Factors in Arid and Semi-Arid Regions of China. Land, 14(8), 1575. https://doi.org/10.3390/land14081575