



Spaceborne LiDAR Reveals Anthropogenic and Biophysical Drivers Shaping the Spatial Distribution of Forest Aboveground Biomass in Eastern Himalayas

 ,
,  , , ,
, , ,  , , and
, , and 
Abstract
1. Introduction
2. Methods
2.1. Study Area
2.2. Data Collection
2.3. Model Development and Validation
3. Results
3.1. Descriptive Statistics and Data Preprocessing
3.2. Model Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ao, A.; Changkija, S.; Tripathi, S.K. Stand Structure, Community Composition and Tree Species Diversity of Sub-Tropical Forest of Nagaland, Northeast India. Trop. Ecol. 2021, 62, 549–562. [Google Scholar] [CrossRef]
- Gogoi, B.; Lahon, D. Impact of Climate Change on Biodiversity of Northeast India: An Overview. Indian J. Appl. Pure Biol. 2022, 37, 322–331. [Google Scholar]
- Jain, A.; Das, A. North-East India: A Unique Biodiversity Paradise Unexplored or Lost? Int. J. Sci. Res. Publ. IJSRP 2022, 12, 112. [Google Scholar] [CrossRef]
- Laskar, B.I. The Sacred Groves of Meghalaya. In Environmental Intimacies from India’s North East; Routledge: New Delhi, India, 2024; ISBN 978-1-003-50487-0. [Google Scholar]
- Tiwari, B.K.; Tynsong, H.; Lynser, M.B. Forest Management Practices of the Tribal People of Meghalaya, North-East India. J. Trop. For. Sci. 2010, 22, 329–342. [Google Scholar]
- Wangpan, T.; Tangjang, S. Significance of Mixed-Cropping in Jhum Based Traditional Agroforestry in Tirap and Longding District of Arunachal Pradesh, India. J. Bioresour. Bioprod. 2015, 2, 62–73. [Google Scholar]
- Fernandes, W. Land, Environmental Degradation and Conflicts in Northeastern India. In Agriculture and a Changing Environment in Northeastern India; Routledge: New Delhi, India, 2020; pp. 119–134. [Google Scholar]
- Semy, K.; Singh, M.R. Changes in Plant Diversity and Community Attributes of Coal Mine Affected Forest in Relation to a Community Reserve Forest of Nagaland, Northeast India. Trop. Ecol. 2024, 65, 16–25. [Google Scholar] [CrossRef]
- Dikshit, K.R.; Dikshit, J.K. Agriculture in North-East India: Past and Present. In North-East India: Land, People and Economy; Advances in Asian Human-Environmental Research; Springer: Dordrecht, The Netherlands, 2014; pp. 587–637. ISBN 978-94-007-7054-6. [Google Scholar]
- Nad, C.; Roy, R.; Roy, T.B. Human Elephant Conflict in Changing Land-Use Land-Cover Scenario in and Adjoining Region of Buxa Tiger Reserve, India. Environ. Chall. 2022, 7, 100384. [Google Scholar] [CrossRef]
- Parida, B.R.; Mahato, T.; Ghosh, S. Monitoring Tea Plantations during 1990–2022 Using Multi-Temporal Satellite Data in Assam (India). Trop. Ecol. 2024, 65, 387–398. [Google Scholar] [CrossRef]
- Pasha, S.V.; Dadhwal, V.K.; Reddy, C.S. Rubber Expansion and Age-Class Mapping in the State of Tripura (India) 1990–2021 Using Multi-Year and Multi-Sensor Data. Environ. Monit. Assess. 2023, 195, 348. [Google Scholar] [CrossRef]
- Lotha, T.N.; Ritse, V.; Nakro, V.; Rudithongru, L.; Hazarika, N.; Jamir, L. Climate Change Impact and Traditional Adaptation Practices in Northeast India: A Review. Curr. World Environ. 2024, 19, 558–575. [Google Scholar] [CrossRef]
- Rai, P.K.; Singh, J.S. Plant Invasion in Protected Areas, the Indian Himalayan Region, and the North East India: Progress and Prospects. Proc. Indian Natl. Sci. Acad. 2021, 87, 19–35. [Google Scholar] [CrossRef]
- FSI India State of Forest Report 2023; Ministry of Environment, Forest and Climate Change, Government of India: New Delhi, India, 2023.
- NEDFi NE—General Information|NER DATABANK. Available online: https://databank.nedfi.com/node/105 (accessed on 13 July 2025).
- Adhikari, D.; Singh, P.P.; Tiwary, R.; Barik, S.K. Forest Carbon Stock-Based Bioeconomy: Mixed Models Improve Accuracy of Tree Biomass Estimates. Biomass Bioenergy 2024, 183, 107142. [Google Scholar] [CrossRef]
- Yanai, R.D.; Wayson, C.; Lee, D.; Espejo, A.B.; Campbell, J.L.; Green, M.B.; Zukswert, J.M.; Yoffe, S.B.; Aukema, J.E.; Lister, A.J. Improving Uncertainty in Forest Carbon Accounting for REDD+ Mitigation Efforts. Environ. Res. Lett. 2020, 15, 124002. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, L.; She, G. Estimating Forest Structural Parameters Using Canopy Metrics Derived from Airborne LiDAR Data in Subtropical Forests. Remote Sens. 2017, 9, 940. [Google Scholar] [CrossRef]
- Ouyang, S.; Xiang, W.; Wang, X.; Xiao, W.; Chen, L.; Li, S.; Sun, H.; Deng, X.; Forrester, D.I.; Zeng, L.; et al. Effects of Stand Age, Richness and Density on Productivity in Subtropical Forests in China. J. Ecol. 2019, 107, 2266–2277. [Google Scholar] [CrossRef]
- Zou, K.; Duan, G.; Wu, Y.; Wang, Z.; Liu, X. Site Index Model for Southern Subtropical Masson Pine Forests Using Stand Dominant Height. Forests 2023, 15, 87. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.; Chhin, S.; Zhang, J.; Duan, A. Disentangling the Effects of Stand and Climatic Variables on Forest Productivity of Chinese Fir Plantations in Subtropical China Using a Random Forest Algorithm. Agric. For. Meteorol. 2021, 304–305, 108412. [Google Scholar] [CrossRef]
- Zhang, J.; Li, M.; Cheng, R.; Ge, Z.; Liu, C.; Zhang, Z. Stand Structure and Functional Traits Determine Productivity of Larix Principis-Rupprechtii Forests. Forests 2024, 15, 924. [Google Scholar] [CrossRef]
- Jucker, T.; Bongalov, B.; Burslem, D.F.R.P.; Nilus, R.; Dalponte, M.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Coomes, D.A. Topography Shapes the Structure, Composition and Function of Tropical Forest Landscapes. Ecol. Lett. 2018, 21, 989–1000. [Google Scholar] [CrossRef]
- Huang, J.; Haider, F.U.; Huang, W.; Liu, S.; Mwangi, B.N.; Suba, V.; Sikuku, L.; Tang, X.; Zhang, Q.; Chu, G. Influence of Photosynthetic Active Radiation on Sap Flow Dynamics across Forest Succession Stages in Dinghushan Subtropical Forest Ecosystem. Heliyon 2024, 10, e37530. [Google Scholar] [CrossRef] [PubMed]
- Kanniah, K.D.; Beringer, J.; North, P.; Hutley, L. Control of Atmospheric Particles on Diffuse Radiation and Terrestrial Plant Productivity: A Review. Prog. Phys. Geogr. Earth Environ. 2012, 36, 209–237. [Google Scholar] [CrossRef]
- Quesada, C.A.; Lloyd, J.; Anderson, L.O.; Fyllas, N.M.; Schwarz, M.; Czimczik, C.I. Soils of Amazonia with Particular Reference to the RAINFOR Sites. Biogeosciences 2011, 8, 1415–1440. [Google Scholar] [CrossRef]
- Asner, G.P.; Mascaro, J. Mapping Tropical Forest Carbon: Calibrating Plot Estimates to a Simple LiDAR Metric. Remote Sens. Environ. 2014, 140, 614–624. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- AIPP; IWGIA. Drivers of Deforestation? Facts to Be Considered Regarding the Impact of Shifting Cultivation in Asia; AIPP: Chiang Mai, Thailand, 2012. [Google Scholar]
- Bruun, T.B.; Hepp, C.M. Carbon Footprint of Shifting Cultivation Landscapes: Current Knowledge, Assumptions and Data Gaps. Carbon Footpr. 2023, 2, 11. [Google Scholar] [CrossRef]
- Mukul, S.A.; Herbohn, J.; Firn, J. Tropical Secondary Forests Regenerating after Shifting Cultivation in the Philippines Uplands Are Important Carbon Sinks. Sci. Rep. 2016, 6, 22483. [Google Scholar] [CrossRef]
- Batar, A.; Watanabe, T.; Kumar, A. Assessment of Land-Use/Land-Cover Change and Forest Fragmentation in the Garhwal Himalayan Region of India. Environments 2017, 4, 34. [Google Scholar] [CrossRef]
- Graham, V.; Geldmann, J.; Adams, V.M.; Negret, P.J.; Sinovas, P.; Chang, H.-C. Southeast Asian Protected Areas Are Effective in Conserving Forest Cover and Forest Carbon Stocks Compared to Unprotected Areas. Sci. Rep. 2021, 11, 23760. [Google Scholar] [CrossRef]
- Collins, M.B.; Mitchard, E.T.A. A Small Subset of Protected Areas Are a Highly Significant Source of Carbon Emissions. Sci. Rep. 2017, 7, 41902. [Google Scholar] [CrossRef]
- Banerjee, A.K. Asia-Pacific Forestry Sector Outlook Study. In Decentralization and Devolution of Forest Management in Asia and the Pacific; Working Paper No.: APFSOS/WP/21; RAP: Saitama, Japan, 1997. [Google Scholar]
- Lainé, N. Effects of the 1996 Timber Ban in Northeast India: The Case of the Khamtis of Lohit District, Arunachal Pradesh. Nat. Environ. Soc. Conserv. Gov. Transform. India 2012, 73–93, xvii–xxiii. [Google Scholar]
- Ahirwal, J.; Nath, A.; Brahma, B.; Deb, S.; Sahoo, U.K.; Nath, A.J. Patterns and Driving Factors of Biomass Carbon and Soil Organic Carbon Stock in the Indian Himalayan Region. Sci. Total Environ. 2021, 770, 145292. [Google Scholar] [CrossRef]
- Sharma, K.; Saikia, A.; Goswami, S.; Borthakur, M. Aboveground Biomass Estimation and Carbon Stock Assessment along a Topographical Gradient in the Forests of Manipur, Northeast India. Arab. J. Geosci. 2020, 13, 443. [Google Scholar] [CrossRef]
- Das, B.; Bordoloi, R.; Deka, S.; Paul, A.; Pandey, P.K.; Singha, L.B.; Tripathi, O.P.; Mishra, B.P.; Mishra, M. Above Ground Biomass Carbon Assessment Using Field, Satellite Data and Model Based Integrated Approach to Predict the Carbon Sequestration Potential of Major Land Use Sector of Arunachal Himalaya, India. Carbon Manag. 2021, 12, 201–214. [Google Scholar] [CrossRef]
- Tashi, S.; Singh, B.; Keitel, C.; Adams, M. Soil Carbon and Nitrogen Stocks in Forests along an Altitudinal Gradient in the Eastern Himalayas and a Meta-analysis of Global Data. Glob. Change Biol. 2016, 22, 2255–2268. [Google Scholar] [CrossRef]
- Traoré, S.; Zo-Bi, I.C.; Piponiot, C.; Aussenac, R.; Hérault, B. Fragmentation Is the Main Driver of Residual Forest Aboveground Biomass in West African Low Forest-High Deforestation Landscapes. Trees For. People 2024, 15, 100477. [Google Scholar] [CrossRef]
- Van Der Laan, C.; Verweij, P.A.; Quiñones, M.J.; Faaij, A.P. Analysis of Biophysical and Anthropogenic Variables and Their Relation to the Regional Spatial Variation of Aboveground Biomass Illustrated for North and East Kalimantan, Borneo. Carbon Balance Manag. 2014, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yamaguchi, Y. Relative Importance of Climatic and Anthropogenic Drivers on the Dynamics of Aboveground Biomass across Agro-Ecological Zones on the Mongolian Plateau. Sustainability 2018, 10, 3435. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Li, C.; Liu, Z. Forest Aboveground Biomass Estimation Using Landsat 8 and Sentinel-1A Data with Machine Learning Algorithms. Sci. Rep. 2020, 10, 9952. [Google Scholar] [CrossRef]
- Shen, W.; Li, M.; Huang, C.; Wei, A. Quantifying Live Aboveground Biomass and Forest Disturbance of Mountainous Natural and Plantation Forests in Northern Guangdong, China, Based on Multi-Temporal Landsat, PALSAR and Field Plot Data. Remote Sens. 2016, 8, 595. [Google Scholar] [CrossRef]
- Pham, T.D.; Yoshino, K.; Bui, D.T. Biomass Estimation of Sonneratia Caseolaris (L.) Engler at a Coastal Area of Hai Phong City (Vietnam) Using ALOS-2 PALSAR Imagery and GIS-Based Multi-Layer Perceptron Neural Networks. GISci. Remote Sens. 2017, 54, 329–353. [Google Scholar] [CrossRef]
- Ranglong, A.; Dutta Roy, A.; Das, S.; Deb, S. Estimating Aboveground Biomass and Biodiversity of a Protected Area in Moist Tropical Forest Ecosystem Using Remote Sensing Application. J. Nat. Conserv. 2025, 84, 126823. [Google Scholar] [CrossRef]
- Liu, Z.; Peng, C.; Work, T.; Candau, J.-N.; DesRochers, A.; Kneeshaw, D. Application of Machine-Learning Methods in Forest Ecology: Recent Progress and Future Challenges. Environ. Rev. 2018, 26, 339–350. [Google Scholar] [CrossRef]
- Zhao, Q.; Yu, S.; Zhao, F.; Tian, L.; Zhao, Z. Comparison of Machine Learning Algorithms for Forest Parameter Estimations and Application for Forest Quality Assessments. For. Ecol. Manag. 2019, 434, 224–234. [Google Scholar] [CrossRef]
- Chen, L.; Ren, C.; Zhang, B.; Wang, Z.; Xi, Y. Estimation of Forest Above-Ground Biomass by Geographically Weighted Regression and Machine Learning with Sentinel Imagery. Forests 2018, 9, 582. [Google Scholar] [CrossRef]
- Dutta Roy, A.; Pitumpe Arachchige, P.S.; Watt, M.S.; Kale, A.; Davies, M.; Heng, J.E.; Daneil, R.; Galgamuwa, G.A.P.; Moussa, L.G.; Timsina, K.; et al. Remote Sensing-Based Mangrove Blue Carbon Assessment in the Asia-Pacific: A Systematic Review. Sci. Total Environ. 2024, 938, 173270. [Google Scholar] [CrossRef]
- López-Serrano, P.M.; Cárdenas Domínguez, J.L.; Corral-Rivas, J.J.; Jiménez, E.; López-Sánchez, C.A.; Vega-Nieva, D.J. Modeling of Aboveground Biomass with Landsat 8 OLI and Machine Learning in Temperate Forests. Forests 2019, 11, 11. [Google Scholar] [CrossRef]
- Wai, P.; Su, H.; Li, M. Estimating Aboveground Biomass of Two Different Forest Types in Myanmar from Sentinel-2 Data with Machine Learning and Geostatistical Algorithms. Remote Sens. 2022, 14, 2146. [Google Scholar] [CrossRef]
- Adrah, E.; Wan Mohd Jaafar, W.S.; Omar, H.; Bajaj, S.; Leite, R.V.; Mazlan, S.M.; Silva, C.A.; Chel Gee Ooi, M.; Mohd Said, M.N.; Abdul Maulud, K.N. Analyzing Canopy Height Patterns and Environmental Landscape Drivers in Tropical Forests Using NASA’s GEDI Spaceborne LiDAR. Remote Sens. 2022, 14, 3172. [Google Scholar] [CrossRef]
- Wang, S.; Liu, C.; Li, W.; Jia, S.; Yue, H. Hybrid Model for Estimating Forest Canopy Heights Using Fused Multimodal Spaceborne LiDAR Data and Optical Imagery. Int. J. Appl. Earth Obs. Geoinf. 2023, 122, 103431. [Google Scholar] [CrossRef]
- Brown, S. Estimating Biomass and Biomass Change of Tropical Forests: A Primer; Food & Agriculture Organization: Rome, Italy, 1997; Volume 134. [Google Scholar]
- Duncanson, L.; Kellner, J.R.; Armston, J.; Dubayah, R.; Minor, D.M.; Hancock, S.; Healey, S.P.; Patterson, P.L.; Saarela, S.; Marselis, S.; et al. Aboveground Biomass Density Models for NASA’s Global Ecosystem Dynamics Investigation (GEDI) Lidar Mission. Remote Sens. Environ. 2022, 270, 112845. [Google Scholar] [CrossRef]
- Kellner, J.R.; Armston, J.; Duncanson, L. Algorithm Theoretical Basis Document for GEDI Footprint Aboveground Biomass Density. Earth Space Sci. 2023, 10, e2022EA002516. [Google Scholar] [CrossRef]
- Dubayah, R.O.; Armston, J.; Kellner, J.R.; Duncanson, L.; Healey, S.P.; Patterson, P.L.; Hancock, S.; Tang, H.; Bruening, J.; Hofton, M.A.; et al. GEDI L4A Footprint Level Aboveground Biomass Density, Version 2.1; Global Ecosystem Dynamics Investigation (GEDI); NASA: Washington, DC, USA, 2022. [Google Scholar]
- Beck, H.E.; McVicar, T.R.; Vergopolan, N.; Berg, A.; Lutsko, N.J.; Dufour, A.; Zeng, Z.; Jiang, X.; Van Dijk, A.I.J.M.; Miralles, D.G. High-Resolution (1 Km) Köppen-Geiger Maps for 1901–2099 Based on Constrained CMIP6 Projections. Sci. Data 2023, 10, 724. [Google Scholar] [CrossRef]
- Negi, V.S.; Pandey, A.; Singh, A.; Bahukhandi, A.; Pharswan, D.S.; Gaira, K.S.; Wani, Z.A.; Bhat, J.A.; Siddiqui, S.; Yassin, H.M. Elevation Gradients Alter Vegetation Attributes in Mountain Ecosystems of Eastern Himalaya, India. Front. For. Glob. Change 2024, 7, 1381488. [Google Scholar] [CrossRef]
- Roy, A.; Das, S.K.; Tripathi, A.K.; Singh, N.U.; Barman, H.K. Biodiversity in North East India and Their Conservation. Progress. Agric. 2015, 15, 182. [Google Scholar] [CrossRef]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R. An Ecoregion-Based Approach to Protecting Half the Terrestrial Realm. BioScience 2017, 67, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved Allometric Models to Estimate the Aboveground Biomass of Tropical Trees. Glob. Change Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Bruening, J.; May, P.; Armston, J.; Dubayah, R. Precise and Unbiased Biomass Estimation from GEDI Data and the US Forest Inventory. Front. For. Glob. Change 2023, 6, 1149153. [Google Scholar] [CrossRef]
- Silva, C.A.; Duncanson, L.; Hancock, S.; Neuenschwander, A.; Thomas, N.; Hofton, M.; Fatoyinbo, L.; Simard, M.; Marshak, C.Z.; Armston, J.; et al. Fusing Simulated GEDI, ICESat-2 and NISAR Data for Regional Aboveground Biomass Mapping. Remote Sens. Environ. 2021, 253, 112234. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef]
- NASA JPL. NASADEM Merged DEM Global 1 Arc Second V001. 2020. Available online: https://www.earthdata.nasa.gov/data/catalog/lpcloud-nasadem-hgt-001 (accessed on 25 March 2024).
- Donchyts, G.; Winsemius, H.; Schellekens, J.; Erickson, T.; Gao, H.; Savenije, H.; van de Giesen, N. Global 30m Height Above the Nearest Drainage. 2016. Available online: https://www.researchgate.net/publication/301559649_Global_30m_Height_Above_the_Nearest_Drainage?channel=doi&linkId=5719ed8f08ae30c3f9f2cc88&showFulltext=true (accessed on 25 March 2024).
- Funk, C.; Peterson, P.; Landsfeld, M.; Pedreros, D.; Verdin, J.; Shukla, S.; Husak, G.; Rowland, J.; Harrison, L.; Hoell, A.; et al. The Climate Hazards Infrared Precipitation with Stations—A New Environmental Record for Monitoring Extremes. Sci. Data 2015, 2, 150066. [Google Scholar] [CrossRef]
- Wan, Z.; Hook, S.; Hulley, G. MOD11A2 MODIS/Terra Land Surface Temperature/Emissivity 8-Day L3 Global 1km SIN Grid V006. 2015. Available online: https://www.earthdata.nasa.gov/data/catalog/lpcloud-mod11a2-006 (accessed on 25 March 2024).
- Wang, D. MODIS/Terra+Aqua Photosynthetically Active Radiation Daily/3-Hour L3 Global 0.05Deg CMG V062. 2024. Available online: https://www.earthdata.nasa.gov/data/catalog/lpcloud-mcd18c2-062 (accessed on 25 March 2024).
- Friedl, M.; Sulla-Menashe, D. MODIS/Terra+Aqua Land Cover Type Yearly L3 Global 500m SIN Grid V061. 2022. Available online: https://www.earthdata.nasa.gov/data/catalog/lpcloud-mcd12q1-061 (accessed on 25 March 2024).
- WII. Protected Area in India (National Park and Wildlife Sanctuary). 2020. Available online: https://geo.ejatlas.org/layers/geonode_data:geonode:Protected_Area_India_Final (accessed on 8 March 2025).
- Hengl, T.; Gupta, S. Soil Water Content (Volumetric %) for 33 kPa and 1500 kPa Suctions Predicted at 6 Standard Depths (0, 10, 30, 60, 100 and 200 cm) at 250 m Resolution. 2019. Available online: https://zenodo.org/records/2784001 (accessed on 25 March 2024).
- Hengl, T. Soil pH in H2O at 6 Standard Depths (0, 10, 30, 60, 100 and 200 cm) at 250 m Resolution. 2018. Available online: https://zenodo.org/records/2525664 (accessed on 25 March 2024).
- Hengl, T. Soil Bulk Density (Fine Earth) 10 × kg / m-cubic at 6 Standard Depths (0, 10, 30, 60, 100 and 200 cm) at 250 m Resolution. 2018. Available online: https://zenodo.org/records/2525665 (accessed on 25 March 2024).
- Hengl, T. Soil Texture Classes (USDA System) for 6 Soil Depths (0, 10, 30, 60, 100 and 200 cm) at 250 m. 2018. Available online: https://zenodo.org/records/2525817 (accessed on 25 March 2024).
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Darst, B.F.; Malecki, K.C.; Engelman, C.D. Using Recursive Feature Elimination in Random Forest to Account for Correlated Variables in High Dimensional Data. BMC Genet. 2018, 19, 65. [Google Scholar] [CrossRef]
- Hanberry, B.B. Practical Guide for Retaining Correlated Climate Variables and Unthinned Samples in Species Distribution Modeling, Using Random Forests. Ecol. Inform. 2024, 79, 102406. [Google Scholar] [CrossRef]
- Dutta Roy, A.; Mohan, M.; Hendy, I.; AlMealla, R.; Watt, M.S.; Burt, J.A.; Torres-Florez, J.P.; Almansoori, A.; Alzahlawi, N.; Abdullah, M.; et al. Optimizing Mangrove Afforestation Site Selection in Gulf Cooperation Council Nations Using Remote Sensing and Machine Learning. Sci. Total Environ. 2025, 988, 179805. [Google Scholar] [CrossRef]
- FSI India State of Forest Report 2021; Ministry of Environment, Forest and Climate Change, Government of India: New Delhi, India, 2021.
- Wang, C.; Wang, J.; He, Z.; Feng, M. Detecting Mountain Forest Dynamics in the Eastern Himalayas. Remote Sens. 2022, 14, 3638. [Google Scholar] [CrossRef]
- Bordoloi, R.; Das, B.; Tripathi, O.P.; Sahoo, U.K.; Nath, A.J.; Deb, S.; Das, D.J.; Gupta, A.; Devi, N.B.; Charturvedi, S.S.; et al. Satellite Based Integrated Approaches to Modelling Spatial Carbon Stock and Carbon Sequestration Potential of Different Land Uses of Northeast India. Environ. Sustain. Indic. 2022, 13, 100166. [Google Scholar] [CrossRef]
- Bhutia, Y.; Gudasalamani, R.; Ganesan, R.; Saha, S. Assessing Forest Structure and Composition along the Altitudinal Gradient in the State of Sikkim, Eastern Himalayas, India. Forests 2019, 10, 633. [Google Scholar] [CrossRef]
- Singh, D.K.; Pusalkar, P.K. Floristic Diversity of the Indian Himalaya. In Biodiversity of the Himalaya: Jammu and Kashmir State; Dar, G.H., Khuroo, A.A., Eds.; Springer: Singapore, 2020; pp. 93–126. ISBN 978-981-329-174-4. [Google Scholar]
- Reddy, C.S.; Jha, C.S.; Diwakar, P.G.; Dadhwal, V.K. Nationwide Classification of Forest Types of India Using Remote Sensing and GIS. Environ. Monit. Assess. 2015, 187, 777. [Google Scholar] [CrossRef]
- Shukla, J.B.; Dubey, B.; Freedman, H.I. Effect of Changing Habitat on Survival of Species. Ecol. Model. 1996, 87, 205–216. [Google Scholar] [CrossRef]
- Murthy, I.K.; Sharma, N.; Nijavalli, H.R. Harnessing REDD+ Opportunities for Forest Conservation and Carbon Stock Enhancement in the Northeastern States of India. Nat. Sci. 2013, 05, 349–358. [Google Scholar] [CrossRef]
- Nath, A.J.; Brahma, B.; Lal, R.; Das, A.K. Jhum: Cultivation. In Encyclopedia of Soil Science, 3rd ed.; Lal, R., Ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 1273–1280. [Google Scholar] [CrossRef]
- Thong, P.; Sahoo, U.K.; Thangjam, U.; Pebam, R. Pattern of Forest Recovery and Carbon Stock Following Shifting Cultivation in Manipur, North-East India. PLoS ONE 2020, 15, e0239906. [Google Scholar] [CrossRef]
- KC, Y.B.; Liu, Q.; Saud, P.; Xu, C.; Gaire, D.; Adhikari, H. Driving Factors and Spatial Distribution of Aboveground Biomass in the Managed Forest in the Terai Region of Nepal. Forests 2024, 15, 663. [Google Scholar] [CrossRef]
- Gogoi, A.; Sahoo, U.K.; Singh, S. Assessment of Biomass and Total Carbon Stock in a Tropical Wet Evergreen Rainforest of Eastern Himalaya along a Disturbance Gradient. J. Plant Biol. Soil Health 2017, 4, 1–8. [Google Scholar]
- Reddy, C.S.; Rakesh, F.; Jha, C.S.; Athira, K.; Singh, S.; Alekhya, V.V.L.P.; Rajashekar, G.; Diwakar, P.G.; Dadhwal, V.K. Geospatial Assessment of Long-Term Changes in Carbon Stocks and Fluxes in Forests of India (1930–2013). 2016, 143, 50–65. [CrossRef]
- De Miranda, S.D.C.; Bustamante, M.; Palace, M.; Hagen, S.; Keller, M.; Ferreira, L.G. Regional Variations in Biomass Distribution in Brazilian Savanna Woodland. Biotropica 2014, 46, 125–138. [Google Scholar] [CrossRef]
- Wang, G.; Guan, D.; Xiao, L.; Peart, M.R. Forest Biomass-Carbon Variation Affected by the Climatic and Topographic Factors in Pearl River Delta, South China. J. Environ. Manag. 2019, 232, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Sanaei, A.; Chahouki, M.A.Z.; Ali, A.; Jafari, M.; Azarnivand, H. Abiotic and Biotic Drivers of Aboveground Biomass in Semi-Steppe Rangelands. Sci. Total Environ. 2018, 615, 895–905. [Google Scholar] [CrossRef]
- Xu, M.; Ji, H.; Zhuang, S. Carbon Stock of Moso Bamboo (Phyllostachys Pubescens) Forests along a Latitude Gradient in the Subtropical Region of China. PLoS ONE 2018, 13, e0193024. [Google Scholar] [CrossRef]
- Negi, V.S.; Joshi, B.C.; Pathak, R.; Rawal, R.S.; Sekar, K.C. Assessment of Fuelwood Diversity and Consumption Patterns in Cold Desert Part of Indian Himalaya: Implication for Conservation and Quality of Life. J. Clean. Prod. 2018, 196, 23–31. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Ji, Y.; Gao, J. Climate Factors Determine the Utilization Strategy of Forest Plant Resources at Large Scales. Front. Plant Sci. 2022, 13, 990441. [Google Scholar] [CrossRef]
- Pelletier, J.; Siampale, A.; Legendre, P.; Jantz, P.; Laporte, N.T.; Goetz, S.J. Human and Natural Controls of the Variation in Aboveground Tree Biomass in African Dry Tropical Forests. Ecol. Appl. 2017, 27, 1578–1593. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.K.; Dutta, B.K.; Sundriyal, R.C.; Das, A.K. Diversity, Representativeness and Biotic Pressure on Plant Species along Alpine Timberline of Western Arunachal Pradesh in the Eastern Himalaya, India. Curr. Sci. 2013, 105, 701–708. [Google Scholar]
- Maza, B.; Rodes-Blanco, M.; Rojas, E. Aboveground Biomass along an Elevation Gradient in an Evergreen Andean—Amazonian Forest in Ecuador. Front. For. Glob. Change 2022, 5, 738585. [Google Scholar] [CrossRef]
- Cheng, Z.; Aakala, T.; Larjavaara, M. Elevation, Aspect, and Slope Influence Woody Vegetation Structure and Composition but Not Species Richness in a Human-Influenced Landscape in Northwestern Yunnan, China. Front. For. Glob. Change 2023, 6, 1187724. [Google Scholar] [CrossRef]
- Du, Q.; Xu, J.; Wang, J.W.; Zhang, F.; Ji, B.Y. Correlation between Forest Carbon Distribution and Terrain Elements of Altitude and Slope. J. Zhejiang Agric. For. Univ. 2013, 30, 330–335. [Google Scholar] [CrossRef]
- Fan, Y.; Zhou, G.M.; Shi, Y.J.; Du, H.Q.; Zhou, Y.F.; Xu, X.J. Effects of Terrain on Stand Structure and Vegetation Carbon Storage of Phyllostachys edulis Forest. Sci. Silvae Sin. 2013, 49, 177–182. [Google Scholar]
- Moser-Reischl, A.; Uhl, E.; Rötzer, T.; Biber, P.; Con, T.V.; Tan, N.T.; Pretzsch, H. Effects of the Urban Heat Island and Climate Change on the Growth of Khaya Senegalensis in Hanoi, Vietnam. For. Ecosyst. 2018, 5, 37. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat Fragmentation and Its Lasting Impact on Earth’s Ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Manrique, S.M.; Vacaflor, P.; Cruz, N. Argentinian Yungas Forest Fragmentation: Effects on Aboveground Biomass, Microclimate and Carbon Storage. Open J. For. 2018, 8, 117–139. [Google Scholar] [CrossRef]
- Astiani, D.W.I.; Curran, L.M.; Mujiman, M.; Ratnasari, D.; Salim, R.; Lisnawaty, N. Edge Effects on Biomass, Growth, and Tree Diversity of a Degraded Peatland in West Kalimantan, Indonesia. Biodiversitas J. Biol. Divers. 2018, 19, 272–278. [Google Scholar] [CrossRef]
- Silva Junior, C.H.L.; Aragão, L.E.O.C.; Anderson, L.O.; Fonseca, M.G.; Shimabukuro, Y.E.; Vancutsem, C.; Achard, F.; Beuchle, R.; Numata, I.; Silva, C.A.; et al. Persistent Collapse of Biomass in Amazonian Forest Edges Following Deforestation Leads to Unaccounted Carbon Losses. Sci. Adv. 2020, 6, eaaz8360. [Google Scholar] [CrossRef] [PubMed]
- Yeneneh, N.; Elias, E.; Feyisa, G.L. Monitoring Soil Quality of Different Land Use Systems: A Case Study in Suha Watershed, Northwestern Highlands of Ethiopia. Environ. Syst. Res. 2024, 13, 7. [Google Scholar] [CrossRef]
- Hao, Y.; He, Z. Effects of Grazing Patterns on Grassland Biomass and Soil Environments in China: A Meta-Analysis. PLoS ONE 2019, 14, e0215223. [Google Scholar] [CrossRef] [PubMed]
- Pinard, M.A.; Putz, F.E. Retaining Forest Biomass by Reducing Logging Damage. Biotropica 1996, 28, 278–295. [Google Scholar] [CrossRef]
- Liu, X.; Feng, Y.; Hu, T.; Luo, Y.; Zhao, X.; Wu, J.; Maeda, E.E.; Ju, W.; Liu, L.; Guo, Q.; et al. Enhancing Ecosystem Productivity and Stability with Increasing Canopy Structural Complexity in Global Forests. Sci. Adv. 2024, 10, eadl1947. [Google Scholar] [CrossRef] [PubMed]
- Gogoi, A.; Sahoo, U.K.; Saikia, H. Vegetation and Ecosystem Carbon Recovery Following Shifting Cultivation in Mizoram-Manipur-Kachin Rainforest Eco-Region, Southern Asia. Ecol. Process. 2020, 9, 21. [Google Scholar] [CrossRef]
- Khaling, S.; Lepcha, P.Y. Shifting to Settled Agriculture: Experiences from Dzongu Valley, North Sikkim, India. In A Tradition in Transition: Understanding the Role of Shifting Cultivation for Sustainable Development of North East India; The Energy and Resources Institute (TERI) Press: New Delhi, India, 2022; p. 246. [Google Scholar]
- Pandey, D.K.; De, H.K.; Dubey, S.K.; Kumar, B.; Dobhal, S.; Adhiguru, P. Indigenous People’s Attachment to Shifting Cultivation in the Eastern Himalayas, India: A Cross-Sectional Evidence. For. Policy Econ. 2020, 111, 102046. [Google Scholar] [CrossRef]
- Ao, A.; Changkija, S.; Brearley, F.Q.; Tripathi, S.K. Plant Community Composition and Carbon Stocks of a Community Reserve Forest in North-East India. Forests 2023, 14, 245. [Google Scholar] [CrossRef]
- Buragohain, M.K.; Dar, A.A.; Babu, K.N.; Parthasarathy, N. Tree Community Structure, Carbon Stocks and Regeneration Status of Disturbed Lowland Tropical Rain Forests of Assam, India. Trees For. People 2023, 11, 100371. [Google Scholar] [CrossRef]
- Chaudhury, G.; Darji, C.B.; Basumatari, M.; Dutta, G.; Devi, A.; Bharadwaj, N. Stand Structure, Biomass and Carbon Stock along Disturbance Gradients in Differently Managed Tropical Forests of Assam, Northeast India. Trees For. People 2022, 9, 100296. [Google Scholar] [CrossRef]
- Mir, A.H.; Chaudhury, G.; Barbhuyan, H.S.A.; Sarma, K.; Upadhaya, K. Impact of Disturbance on Community Structure, Biomass and Carbon Stock in Montane Evergreen Forests of Meghalaya, Northeast India. Carbon Manag. 2021, 12, 215–233. [Google Scholar] [CrossRef]
- Duncanson, L.; Liang, M.; Leitold, V.; Armston, J.; Krishna Moorthy, S.M.; Dubayah, R.; Costedoat, S.; Enquist, B.J.; Fatoyinbo, L.; Goetz, S.J. The Effectiveness of Global Protected Areas for Climate Change Mitigation. Nat. Commun. 2023, 14, 2908. [Google Scholar] [CrossRef]
- Williams, B.A.; Beyer, H.L.; Fagan, M.E.; Chazdon, R.L.; Schmoeller, M.; Sprenkle-Hyppolite, S.; Griscom, B.W.; Watson, J.E.; Tedesco, A.M.; Gonzalez-Roglich, M. Global Potential for Natural Regeneration in Deforested Tropical Regions. Nature 2024, 636, 131–137. [Google Scholar] [CrossRef]
- Sharma, C.K.; Sarma, I. Issues of Conservation and Livelihood in a Forest Village of Assam. Int. J. Rural Manag. 2014, 10, 47–68. [Google Scholar] [CrossRef]
- Tynsong, H.; Dkhar, M.; Tiwari, B. Review: Traditional Ecological Knowledge of Tribal Communities of North East India. Biodiversitas J. Biol. Divers. 2020, 21, 3209–3224. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, M.; Li, G.; Wang, M.; Li, Z.; De Boeck, H.J. Changes of Aboveground and Belowground Biomass Allocation in Four Dominant Grassland Species across a Precipitation Gradient. Front. Plant Sci. 2021, 12, 650802. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Dean, T.J. Relationship of Aboveground Biomass Production, Site Index and Soil Characteristics in a Loblolly Pine Stands. In General Technical Report SRS–71 Asheville NC; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2004; pp. 368–371. [Google Scholar]
- Longepierre, M.; Feola Conz, R.; Barthel, M.; Bru, D.; Philippot, L.; Six, J.; Hartmann, M. Mixed Effects of Soil Compaction on the Nitrogen Cycle under Pea and Wheat. Front. Microbiol. 2022, 12, 822487. [Google Scholar] [CrossRef]
- Rao, L.G.G.; Joseph, B.; Sreemannarayana, B. Growth and Biomass Production of Some Important Multipurpose Tree Species on Rainfed Sandy Loam Soils. Indian For. 2000, 126, 772–781. [Google Scholar]
- Ye, C.; Zheng, G.; Tao, Y.; Xu, Y.; Chu, G.; Xu, C.; Chen, S.; Liu, Y.; Zhang, X.; Wang, D. Effect of Soil Texture on Soil Nutrient Status and Rice Nutrient Absorption in Paddy Soils. Agronomy 2024, 14, 1339. [Google Scholar] [CrossRef]
- Choudhury, B.U.; Fiyaz, A.R.; Mohapatra, K.P.; Ngachan, S. Impact of Land Uses, Agrophysical Variables and Altitudinal Gradient on Soil Organic Carbon Concentration of North-Eastern Himalayan Region of India. Land Degrad. Dev. 2016, 27, 1163–1174. [Google Scholar] [CrossRef]
- de Castilho, C.V.; Magnusson, W.E.; de Araújo, R.N.O.; Luizao, R.C.; Luizao, F.J.; Lima, A.P.; Higuchi, N. Variation in Aboveground Tree Live Biomass in a Central Amazonian Forest: Effects of Soil and Topography. For. Ecol. Manag. 2006, 234, 85–96. [Google Scholar] [CrossRef]
- Jia, D.; Wang, C.; Hakkenberg, C.R.; Numata, I.; Elmore, A.J.; Cochrane, M.A. Accuracy Evaluation and Effect Factor Analysis of GEDI Aboveground Biomass Product for Temperate Forests in the Conterminous United States. GISci. Remote Sens. 2024, 61, 2292374. [Google Scholar] [CrossRef]
- Berninger, A.; Lohberger, S.; Zhang, D.; Siegert, F. Canopy Height and Above-Ground Biomass Retrieval in Tropical Forests Using Multi-Pass X- and C-Band Pol-InSAR Data. Remote Sens. 2019, 11, 2105. [Google Scholar] [CrossRef]
- Jiang, P.; He, H.; Mao, Z.; Su, Y.; Zhang, M.; Zhu, M.; Xu, Z.; Yin, J.; Jiang, H.; Fang, S.; et al. Canopy Properties Predominately Explain Above-Ground Biomass Stock in Temperate Forests. J. Plant Ecol. 2025, 18, rtaf026. [Google Scholar] [CrossRef]
- Meyer, V.; Saatchi, S.; Clark, D.B.; Keller, M.; Vincent, G.; Ferraz, A.; Espírito-Santo, F.; d’Oliveira, M.V.N.; Kaki, D.; Chave, J. Canopy Area of Large Trees Explains Aboveground Biomass Variations across Neotropical Forest Landscapes. Biogeosciences 2018, 15, 3377–3390. [Google Scholar] [CrossRef]
- Dutta Roy, A.; Das, S.K.; Debnath, B. A Non-Destructive Approach to Develop Tree-Level Allometric Equations for Estimating Aboveground Biomass in the Forests of Tripura, Northeast India. Trop. Ecol. 2023, 64, 532–542. [Google Scholar] [CrossRef]
- Nath, A.; Tiwari, B.; Sileshi, G.; Sahoo, U.; Brahma, B.; Deb, S.; Devi, N.; Das, A.; Reang, D.; Chaturvedi, S.; et al. Allometric Models for Estimation of Forest Biomass in North East India. Forests 2019, 10, 103. [Google Scholar] [CrossRef]
- Dutta Roy, A.; Debbarma, S. Comparing the Allometric Model to Machine Learning Algorithms for Aboveground Biomass Estimation in Tropical Forests. Ecol. Front. 2024, 44, 1069–1078. [Google Scholar] [CrossRef]
- Dong, L.; Du, H.; Han, N.; Li, X.; Zhu, D.; Mao, F.; Zhang, M.; Zheng, J.; Liu, H.; Huang, Z. Application of Convolutional Neural Network on Lei Bamboo Above-Ground-Biomass (AGB) Estimation Using Worldview-2. Remote Sens. 2020, 12, 958. [Google Scholar] [CrossRef]


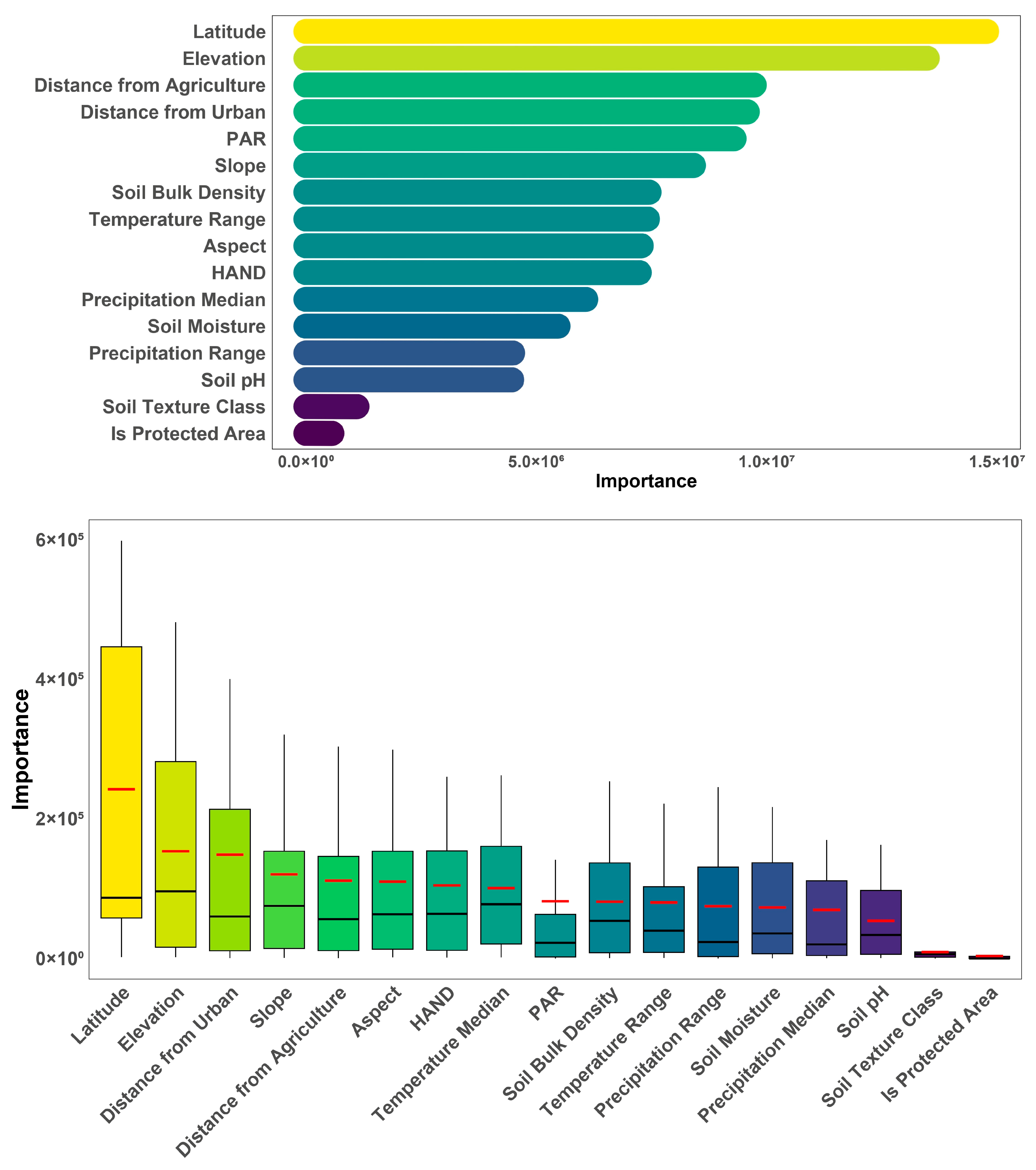
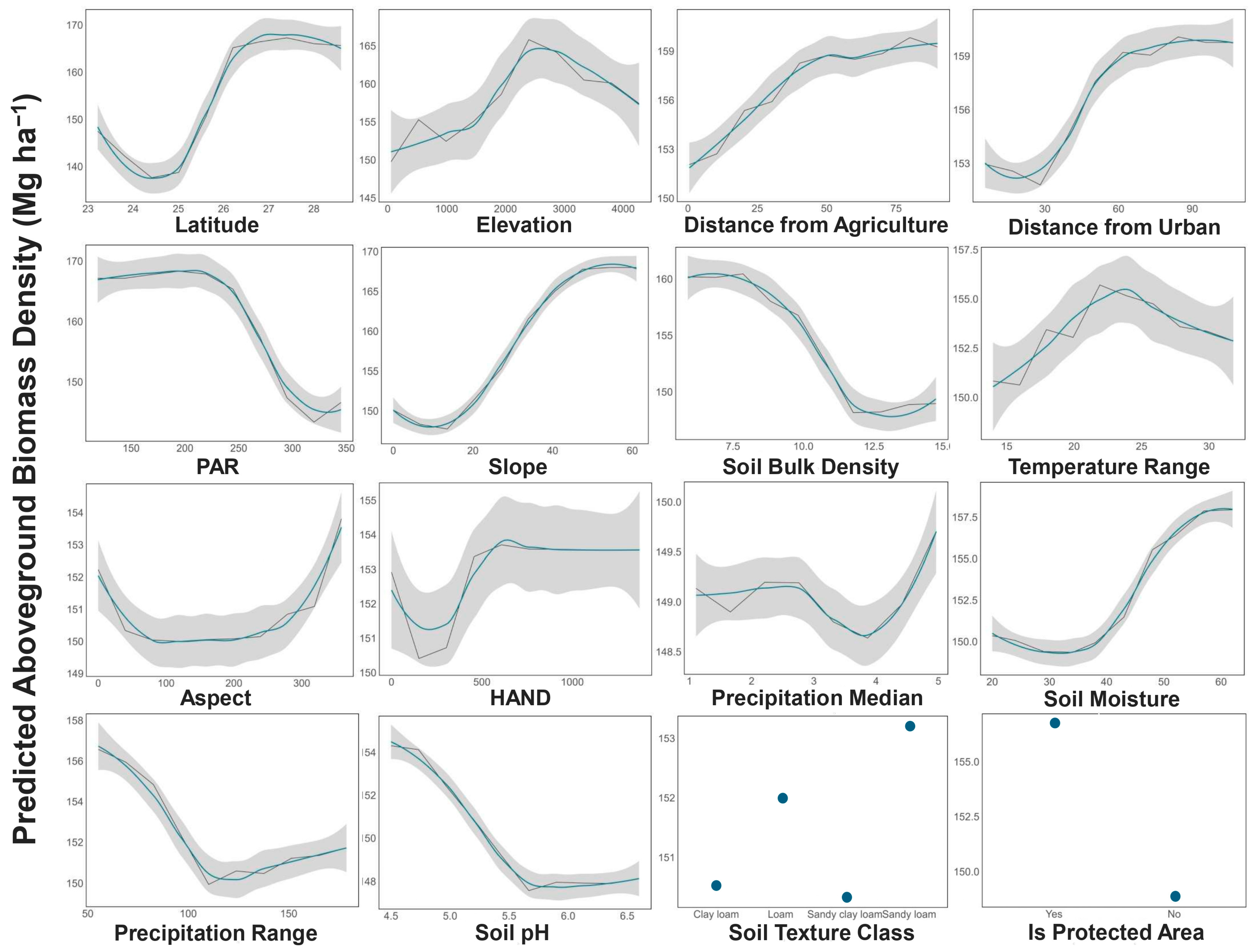

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State | GEDI Footprints | AGBD (Mg ha−1) | ||
|---|---|---|---|---|
| Median | Mean ± SD | Range (Min–Max) | ||
| Eastern Himalayas (Overall) | 30,257 | 129.1 | 149.6 ± 79.5 | 10.6 to 499.7 |
| Arunachal Pradesh | 4997 | 186.7 | 192.4 ± 87.7 | 10.9 to 498.7 |
| Assam | 4823 | 137.5 | 144.8 ± 58.2 | 57.3 to 497.9 |
| Manipur | 3790 | 103.0 | 102.8 ± 24.7 | 57.1 to 145.5 |
| Meghalaya | 4134 | 139.4 | 158.3 ± 77.4 | 57.1 to 493.6 |
| Mizoram | 3996 | 109.4 | 133.0 ± 76.3 | 57.2 to 496.6 |
| Nagaland | 2756 | 163.8 | 197.0 ± 109.2 | 44.5 to 499.7 |
| Sikkim | 731 | 201.1 | 218.0 ± 119.0 | 10.6 to 498.1 |
| Tripura | 2756 | 113.65 | 123.8 ± 49.9 | 57.1 to 421.1 |
| West Bengal (north Bengal districts) | 2274 | 109.46 | 108.7 ± 28.4 | 57.2 to 156.6 |
| Model Code | Ecoregion Name | GEDI Footprints | AGBD (Mg ha−1) | ||
|---|---|---|---|---|---|
| Median | Mean ± SD | Range (Min–Max) | |||
| EH | Eastern Himalayas (Overall) | 30,257 | 129.1 | 149.6 ± 79.5 | 10.6 to 499.7 |
| E01 | Brahmaputra valley semi-evergreen forests | 3189 | 124.6 | 136.5 ± 62.8 | 57.1 to 497.8 |
| E02 | Eastern Himalayan broadleaf forests | 1852 | 209.0 | 219.3 ± 110.9 | 24.3 to 498.3 |
| E03 | Eastern Himalayan subalpine conifer forests | 2646 | 183.1 | 184.8 ± 79.6 | 10.6 to 498.0 |
| E04 | Himalayan subtropical pine forests | 178 | 238.1 | 245.5 ± 119.7 | 26.0 to 494.4 |
| E05 | Lower Gangetic plains moist deciduous forests | 2369 | 108.2 | 117.2 ± 48.2 | 57.1 to 421.1 |
| E06 | Meghalaya subtropical forests | 6312 | 141.2 | 153.9 ± 69.0 | 57.0 to 493.5 |
| E07 | Mizoram-Manipur-Kachin rainforests | 6822 | 110.8 | 128.3 ± 67.7 | 57.2 to 496.6 |
| E08 | Northeast Himalayan subalpine conifer forests | 1052 | 170.2 | 172.9 ± 67.5 | 15.8 to 304.8 |
| E09 | Northeast India-Myanmar pine forests | 4733 | 121.7 | 152.6 ± 93.9 | 44.4 to 499.6 |
| E10 | Terai-Duar savanna and grasslands | 1104 | 113.9 | 111.7 ± 28.5 | 57.2 to 156.5 |
| Serial Number | Variable | Unit | Variable Parameters | ||
|---|---|---|---|---|---|
| Median | Mean ± SD | Range (Min–Max) | |||
| 1 | Elevation | m | 785 | 1068.0 ± 919.7 | 53 to 4268 |
| 2 | Slope | Degree (0–90°) | 16.07 | 15.6 ± 9.2 | 0 to 61.0 |
| 3 | Aspect | Degree (0–360°) | 180 | 179.9 ± 107.5 | 0 to 358.4 |
| 4 | Latitude | Coordinates | 26.07 | 25.9 ± 1.5 | 23.21 to 28.6 |
| 5 | Height above nearest drainage (HAND) | m | 38 | 76.4 ± 97.4 | 0 to 1461 |
| 6 | Median 5-day precipitation (1984–2024) | mm | 0.43 | 0.7 ± 0.9 | 0.01 to 4.9 |
| 7 | Precipitation range (1984–2024) | mm | 99.63 | 103.3 ± 21.4 | 55.2 to 178.7 |
| 8 | Median daily land surface temperature (2004–2024) | °C | 22.27 | 20.5 ± 5.3 | 1.6 to 27.5 |
| 9 | Land surface temperature range (2004–2024) | °C | 18.16 | 19.0 ± 3.0 | 13.9 to 31.8 |
| 10 | Photosynthetically active radiation (PAR) | E m–2 d–1 | 306.88 | 284.3 ± 56.6 | 118.9 to 345.0 |
| 11 | Distance from agricultural areas | km | 12.5 | 22.8 ± 25.8 | 0.5 to 89.9 |
| 12 | Distance from urban areas | km | 35.65 | 42.3 ± 22.4 | 5.7 to 107.0 |
| 13 | Soil moisture content | % | 36 | 37.1 ± 4.9 | 18 to 62 |
| 14 | Soil pH | - | 5.3 | 5.3 ± 0.3 | 4.5 to 6.6 |
| 15 | Soil bulk density | kg m–3 | 12 | 11.4 ± 1.4 | 5.9 to 14.7 |
| Serial Number | Variable | Category | Proportion (%) |
|---|---|---|---|
| 1 | Is it a protected area (PA)? | Yes | 12.9 |
| No | 87.1 | ||
| 2 | Soil texture class | Clay loam | 65.3 |
| Loam | 30.9 | ||
| Sandy clay loam | 2.8 | ||
| Sandy loam | 1.0 |
| Model Code | Selected Regression Hyperparameters | Model Performance Metrics | |||
|---|---|---|---|---|---|
| Variables per Split (mtry) | Number of Decision Trees (ntree) | R2 | RMSE (Mg ha−1) | %RMSE | |
| EH | 2 | 1400 | 0.41 | 60.2 | 40.3 |
| E01 | 2 | 200 | 0.28 | 52.2 | 38.4 |
| E02 | 6 | 90 | 0.30 | 93.2 | 42.2 |
| E03 | 3 | 120 | 0.27 | 67.9 | 36.1 |
| E04 | 12 | 8 | 0.31 | 101.7 | 49.5 |
| E05 | 2 | 120 | 0.16 | 43.6 | 36.8 |
| E06 | 2 | 375 | 0.28 | 59.1 | 38.2 |
| E07 | 3 | 450 | 0.35 | 53.5 | 42.0 |
| E08 | 2 | 48 | 0.22 | 61.8 | 35.7 |
| E09 | 2 | 300 | 0.59 | 62.1 | 40.3 |
| E10 | 2 | 60 | 0.10 | 27.3 | 24.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dutta Roy, A.; Ranglong, A.; Timilsina, S.; Das, S.K.; Watt, M.S.; de-Miguel, S.; Deb, S.; Sahoo, U.K.; Mohan, M. Spaceborne LiDAR Reveals Anthropogenic and Biophysical Drivers Shaping the Spatial Distribution of Forest Aboveground Biomass in Eastern Himalayas. Land 2025, 14, 1540. https://doi.org/10.3390/land14081540
Dutta Roy A, Ranglong A, Timilsina S, Das SK, Watt MS, de-Miguel S, Deb S, Sahoo UK, Mohan M. Spaceborne LiDAR Reveals Anthropogenic and Biophysical Drivers Shaping the Spatial Distribution of Forest Aboveground Biomass in Eastern Himalayas. Land. 2025; 14(8):1540. https://doi.org/10.3390/land14081540
Chicago/Turabian StyleDutta Roy, Abhilash, Abraham Ranglong, Sandeep Timilsina, Sumit Kumar Das, Michael S. Watt, Sergio de-Miguel, Sourabh Deb, Uttam Kumar Sahoo, and Midhun Mohan. 2025. "Spaceborne LiDAR Reveals Anthropogenic and Biophysical Drivers Shaping the Spatial Distribution of Forest Aboveground Biomass in Eastern Himalayas" Land 14, no. 8: 1540. https://doi.org/10.3390/land14081540
APA StyleDutta Roy, A., Ranglong, A., Timilsina, S., Das, S. K., Watt, M. S., de-Miguel, S., Deb, S., Sahoo, U. K., & Mohan, M. (2025). Spaceborne LiDAR Reveals Anthropogenic and Biophysical Drivers Shaping the Spatial Distribution of Forest Aboveground Biomass in Eastern Himalayas. Land, 14(8), 1540. https://doi.org/10.3390/land14081540