


Mid-Term Evaluation of Herbaceous Cover Restoration on Skid Trails Following Ground-Based Logging in Pure Oriental Beech (Fagus orientalis Lipsky) Stands of the Hyrcanian Forests, Northern Iran


 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Method
- Twenty-seven skid trail plots: 3 logging intervals × 3 traffic intensities × 3 replicates.
- Twenty-seven control plots: 3 logging intervals × 3 controls (paired with skid trail intensities) × 3 replicates.
2.3. Data Analysis
- Protected LSD (Least Significant Difference) for mean comparisons;
- Pairwise t-tests to contrast specific treatment levels.
3. Results
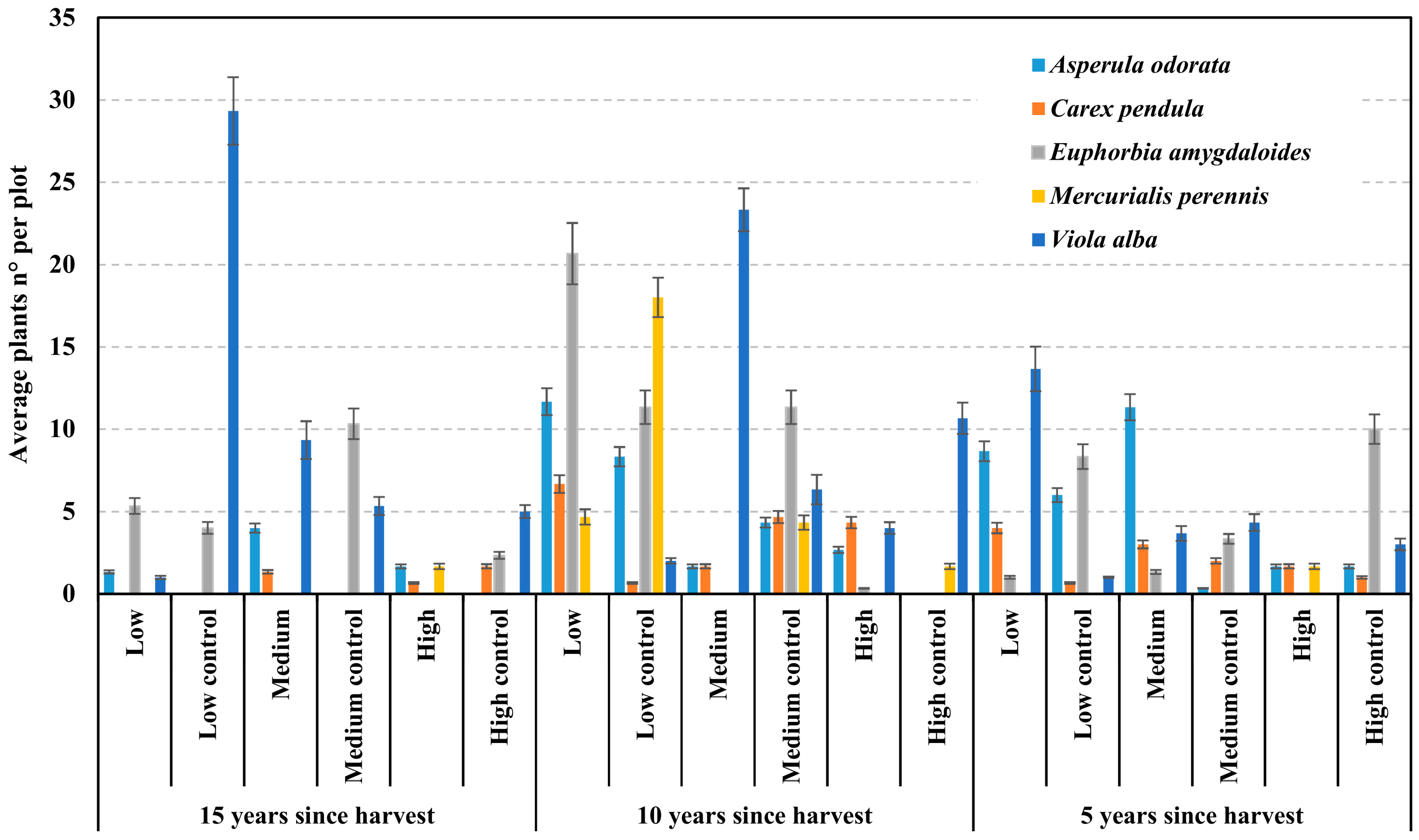
- Violaceae dominated, with 430 specimens (e.g., Viola alba).
- Euphorbiaceae (e.g., Euphorbia amygdaloides) and Rubiaceae (e.g., Asperula odorata) followed, with 269 and 196 individuals, respectively.
3.1. Number of Plants and Traffic Intensity
3.2. Number of Species and Traffic Intensity
4. Discussion
4.1. Number of Plants
4.2. Number of Species
- In the 5-year post-harvest period: The Simpson index indicated greater evenness on six skid trail plots in comparison to that for the control plots.
- In the 10-year post-harvest period: No significant differences in evenness were observed between skid trails and controls.
- In the 15-year post-harvest period: Skid trail plots exhibited higher evenness than the control plots, suggesting gradual recovery toward pre-disturbance conditions.
- Soil recovery dynamics: Post-logging soil compaction and nutrient cycling shifts.
- Light-regime changes: Altered canopy cover affecting shade-tolerant vs. pioneer species.
- Species interactions: Competitive exclusion, facilitation, or recruitment limitations.
- The temporal scales of recovery: How decades-long successional processes mediate diversity trajectories.
- Scale-dependent responses: Variations in species richness and community assembly across spatial gradients (e.g., microsites vs. landscape-level impacts).
- Mechanistic drivers: Genetic, functional, and environmental filters shaping post-logging recolonization.
5. Conclusions
- Low-traffic areas: Limit machinery use to designated paths, and enforce seasonal restrictions (e.g., avoiding wet periods) to minimize soil compaction. Promote natural recovery by protecting resilient species (e.g., clonal herbs).
- Medium-high-traffic areas: Prioritize the planting of species with high stress tolerance (e.g., deep-rooted perennials). Use brush mats or slash layers on skid trails to reduce direct soil impact.
- High traffic zones: Consider permanent trail closure after repeated use, coupled with active restoration (e.g., reseeding with native, disturbance-adapted species).
- Long-term monitoring: Track post-logging recovery beyond 15 years to assess resilience thresholds.
- Scale-dependent planning: Address microsite variability (e.g., light, soil compaction, etc.) to aid species recolonization.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chudy, R.P.; Cubbage, F.W. Research trends: Forest investments as a financial asset class. For. Policy Econ. 2020, 119, 102273. [Google Scholar] [CrossRef] [PubMed]
- Obst, C.; Hein, L.; Edens, B. National Accounting and the Valuation of Ecosystem Assets and Their Services. Environ. Monit. Assess. 2016, 64, 794. [Google Scholar] [CrossRef]
- Labelle, E.R.; Hansson, L.; Högbom, L.; Jourgholami, M.; Laschi, A. Strategies to Mitigate the Effects of Soil Physical Disturbances Caused by Forest Machinery: A Comprehensive Review. Curr. For. Rep. 2022, 8, 20–37. [Google Scholar] [CrossRef]
- Latterini, F.; Dyderski, M.K.; Horodecki, P.; Picchio, R.; Venanzi, R.; Lapin, K.; Jagodziński, A.M. The Effects of Forest Operations and Silvicultural Treatments on Litter Decomposition Rate: A Meta-Analysis. Curr. For. Rep. 2023, 9, 276–290. [Google Scholar] [CrossRef]
- Marchi, E.; Chung, W.; Visser, R.; Abbas, D.; Nordfjell, T.; Mederski, P.S.; McEwan, A.; Brink, M.; Laschi, A. Sustainable Forest Operations (SFO): A new paradigm in a changing world and climate. Sci. Total. Environ. 2018, 634, 1385–1397. [Google Scholar] [CrossRef]
- Latterini, F.; Dyderski, M.K.; Horodecki, P.; Rawlik, M.; Stefanoni, W.; Högbom, L.; Venanzi, R.; Picchio, R.; Jagodziński, A.M. A meta-analysis of the effects of ground-based extraction technologies on fine roots in forest soils. Land. Degrad. Dev. 2024, 35, 9–21. [Google Scholar] [CrossRef]
- Grünberg, J.; Ghaffariyan, M.R.; Jourgholami, M.; Labelle, E.R.; Kaakkurivaara, N.; Robert, R.C.G.; Kühmaier, M. Criteria for Assessing the Sustainability of Logging Operations. A Systematic Review. Curr. For. Rep. 2023, 9, 350–369. [Google Scholar] [CrossRef]
- Jourgholami, M.; Hosseiniala, E.A.; Latterini, F.; Venanzi, R.; Picchio, R. The Effects of Soil Compaction on the Growth and Architecture of the Seedlings of Species Commonly Used for Afforestation in Iran. Forests 2024, 15, 1090. [Google Scholar] [CrossRef]
- Vennin, S.; Montpied, P.; Behr, P.; Thimonier, A.; Dupouey, J.L. Mechanisation of forest operations drives long-term changes in plant communities. For. Ecol. Manag. 2025, 585, 122562. [Google Scholar] [CrossRef]
- Kiumarsi, F.; Jourgholami, M.; Jafari, M.; Lo Monaco, A.; Venanzi, R.; Picchio, R. Restoring soil properties in the Hyrcanian forests from machine induced compaction: Reforestation of N2-fixing black alder (Alnus glutinosa (L.) Gaertn.). Land. Degrad. Dev. 2024, 35, 2084–2096. [Google Scholar] [CrossRef]
- Samdaliri, H.; Jourgholami, M.; Salajegheh, A.; Sohrabi, H.; Venanzi, R.; Picchio, R.; Lo Monaco, A. The Impact of Group- and Single-Tree-Selection Cuttings on Runoff and Sediment Yield in Mixed Broadleaved Forests, Northern Iran. Sustainability 2024, 16, 1830. [Google Scholar] [CrossRef]
- Picchio, R.; Mederski, P.S.; Tavankar, F. How and how much do harvesting activities affect forest soil, regeneration and stands? Curr. For. Rep. 2020, 6, 115–128. [Google Scholar] [CrossRef]
- Picchio, R.; Jourgholami, M.; Zenner, E.K. Effects of forest harvesting on water and sediment yields: A review toward better mitigation and rehabilitation strategies. Curr. For. Rep. 2021, 7, 214–229. [Google Scholar] [CrossRef]
- Peterson, D.W.; Dodson, E.K. Post-fire logging produces minimal persistent impacts on understory vegetation in northeastern Oregon, USA. For. Ecol. Manag. 2016, 370, 56–64. [Google Scholar] [CrossRef]
- Mattson, L.R.; Coop, J.D.; Battaglia, M.A.; Cheng, A.S.; Sibold, J.S.; Viner, S. Post-spruce beetle timber salvage drives short-term surface fuel increases and understory vegetation shifts. Ecol. Manag. 2019, 437, 348–359. [Google Scholar] [CrossRef]
- Ding, Y.; Zang, R.; Lu, X.; Huang, J. The impacts of selective logging and clear-cutting on woody plant diversity after 40 years of natural recovery in a tropical montane rain forest, south China. Sci. Total Environ. 2017, 579, 1683–1691. [Google Scholar] [CrossRef]
- Makineci, E.; Sat Gungor, B.; Demir, M. Survived herbaceous plant species on compacted skid road in a fir (Abies bornmulleriana Mattf.) forest—A note. Transp. Res. D Trans. Environ. 2008, 13, 187–192. [Google Scholar] [CrossRef]
- Makineci, E.; Demir, M.; Yilmaz, E. Long-term harvesting effects on skid road in a fir (Abies bornmulleriana Mattf.) plantation forest. Build. Environ. 2007, 42, 1538–1543. [Google Scholar] [CrossRef]
- Buckley, D.S.; Crow, T.R.; Nauertz, E.A.; Schulz, K.E. Influence of skid trails and haul roads on understory plant richness and composition in managed forest landscapes in Upper Michigan, USA. For. Ecol. Manag. 2003, 175, 509–520. [Google Scholar] [CrossRef]
- Makineci, E.; Demit, M.; Comez, A.; Yilmaz, E. Effects of Timber Skidding on Chemical Characteristics of Herbaceous Cover, Forest Floor and Topsoil on Skidroad in an Oak (Quercus petraea L.) Forest. J. Ter. 2007, 44, 423–428. [Google Scholar] [CrossRef]
- Makineci, E.; Demit, M.; Comez, A.; Yilmaz, E. Chemical Characteristics of the Surface Soil, Herbaceous Cover and Organic Layer of a Compacted Skid Road in a Fir (Abies bornmulleriana Mattf.) Forest. Transp. Res. Part. D Tra. Env. 2007, 12, 453–459. [Google Scholar] [CrossRef]
- Mercier, P.; Aas, G.; Dengler, J. Effects of skid trails on understory vegetation in forests: A case study from Northern Bavaria (Germany). For. Ecol. Manag. 2019, 453, 117579. [Google Scholar] [CrossRef]
- Klein-Raufhake, T.; Hölzel, N.; Schaper, J.J.; Hortmann, A.; Elmer, M.; Fornfeist, M.; Linnemann, B.; Meyer, M.; Rentemeister, K.; Santora, L.; et al. Severity of topsoil compaction controls the impact of skid trails on soil ecological processes. J. Appl. Ecol. 2024, 61, 1817–1828. [Google Scholar] [CrossRef]
- Çesonienė, L.; Daubaras, R.; Kaškonas, P.; Kaškonienė, V.; Maruška, A.S.; Tiso, N.; Zych, M. Initial impact of clear-cut logging on dynamics of understory vascular plants and pollinators in Scots pine-dominated forests in Lithuania. Turk. J. Agric. For. 2018, 42, 433–444. [Google Scholar] [CrossRef]
- Mollinari, M.M.; Peres, C.A.; Edwards, D.P. Rapid recovery of thermal environment after selective logging in the Amazon. Agric. For. Meteorol. 2019, 278, 107637. [Google Scholar] [CrossRef]
- Matula, R.; Řepka, R.; Šebesta, J.; Pettit, J.L.; Chamagne, J.; Šrámek, M.; Horgan, K.; Maděra, P. Resprouting trees drive understory vegetation dynamics following logging in a temperate forest. Sci. Rep. 2020, 10, 9231. [Google Scholar] [CrossRef]
- Xiong, Q.; Li, L.; Luo, X.; He, X.; Zhang, L.; Pan, K.; Liu, C.; Sun, H. Driving forces for recovery of forest vegetation after harvesting a subalpine oak forest in eastern Tibetan Plateau. Environ. Sci. Pollut. Res. 2021, 28, 67748–67763. [Google Scholar] [CrossRef]
- Williamson, J.R.; Neilsen, W.A. The influence of forest site on rate and extent of soil compaction and profile disturbance of skid trails during ground-based harvesting. Can. J. For. Res. 2000, 30, 1196–1205. [Google Scholar] [CrossRef]
- Litschert, S.E.; MacDonald, L.H. Frequency and characteristics of sediment delivery pathways from forest harvest units to streams. For Ecol. Manag. 2009, 259, 143–150. [Google Scholar] [CrossRef]
- Ampoorter, E.; De Schrijver, A.; De Frenne, P.; Hrrmy, M.; Verheyen, K. Experimental assessment of ecological restoration options for compacted forest soils. Ecol. Eng. 2011, 37, 1734–1746. [Google Scholar] [CrossRef]
- DeArmond, D.; Ferraz, J.B.S.; Lovera, L.H.; de Souza, C.A.S.; Corrêa, C.; Spanner, G.C.; Lima, A.J.N.; dos Santos, J.; Higuchi, N. Impacts to soil properties still evident 27 years after abandonment in Amazonian log landings. For. Ecol. Manag. 2022, 510, 120105. [Google Scholar] [CrossRef]
- Rab, M.A. Recovery of soil physical properties from compaction and soil profile disturbance caused by logging of native forest in Victorian Central Highlands, Australia. For. Ecol. Manag. 2004, 191, 329–340. [Google Scholar] [CrossRef]
- DeArmond, D.; Ferraz, J.B.S.; de Oliveira, L.R.; Lima, A.J.N.; Falcão, N.P.D.S.; Higuchi, N. Soil compaction in skid trails still affects topsoil recovery 28 years after logging in Central Amazonia. Geoderma 2023, 434, 116473. [Google Scholar] [CrossRef]
- Mousavi, S.A.; Sarmadian, F.; Taati, A. Comparison of the FAO method and the analytical hierarchy process (AHP) to evaluate land suitability for dry wheat in Kohin region. J. Soil Res. (Soil Water Sci.) 2015, 30, 367–377. (In Persian) [Google Scholar]
- Closset-Kopp, D.; Hattab, T.; Decocq, G. Do drivers of forestry vehicles also drive herb layer changes (1970–2015) in a temperate forest with contrasting habitat and management conditions? J. Ecol. 2019, 107, 1439–1456. [Google Scholar] [CrossRef]
- Khanalizadeh, A.; Eshaghi Rad, J.; Amiri, G.Z.; Zare, H.; Schall, P.; Lexer, M.J. Effects of single tree selection cutting on vascular plant species diversity components in temperate forests of Hyrcanian region. Flora 2023, 305, 152341. [Google Scholar] [CrossRef]
- Picchio, R.; Tavankar, F.; Venanzi, R.; Lo Monaco, A.; Nikkooy, M. Study of forest road effect on tree community and stand structure in three Italian and Iranian temperate forests. Croat. J. For. Eng. 2018, 39, 57–70. [Google Scholar]
- Demir, M.; Makineci, E.; Yilmaz, E. Harvesting impacts on herbaceous understory, forest floor and top soil properties on skid road in a beech (Fagus orientalis Lipsky.) stand. J. Environ. Biol. 2007, 28, 427–432. [Google Scholar]
- Demir, M.; Makineci, E.; Gungor, B.S. Plant Species Recovery on a Compacted Skid Road. Sensors 2008, 8, 3123–3133. [Google Scholar] [CrossRef]
- Demir, M.; Makineci, E.; Yilmaz, E. Investigation of timber harvesting impacts on herbaceous cover, forest floor and surface soil properties on skid road in an oak (Quercus petraea L.) stand. Build. Environ. 2007, 42, 1194–1199. [Google Scholar] [CrossRef]
- Karamirad, S.; Abdi, A.; Majnounian, B.; Etemad, V.; Sohrabi, H. The impact of forest roads on herbaceous species diversity and tree regeneration establishment (Case study: Kheiroud forest). J. Wood Prod. (Iran. J. Nat. Resour.) 2016, 69, 29–40. [Google Scholar]
- Ghorbanalizadeh, A.; Akhani, H. Plant diversity of Hyrcanian relict forests: An annotated checklist, chorology and threat categories of endemic and near endemic vascular plant species. Plant Div. 2022, 44, 39–69. [Google Scholar] [CrossRef]
- Ramezani, E.; Klerk, P.D.; Naqinezhad, A.; Theuerkauf, M.; Joosten, H. Long-term dynamics of Oriental beech (Fagus orientalis Lipsky) stands in the Hyrcanian forests of northern Iran. Rev. Palaeobot. Palynol. 2023, 312, 104871. [Google Scholar] [CrossRef]
- Fornwalt, P.J.; Rhoades, C.C.; Hubbard, R.M.; Harris, R.L.; Faist, A.M.; Bowman, W.D. Short-term understory plant community responses to salvage logging in beetle-affected lodgepole pine forests. For. Ecol. Manag. 2018, 409, 84–93. [Google Scholar] [CrossRef]
- Moola, F.M.; Vasseur, L. The maintenance of understory residual flora with even-aged forest management: A review of temperate forests in northeastern North America. Environ. Rev. 2008, 16, 141–155. [Google Scholar] [CrossRef]
- Michalová, Z.; Morrissey, R.C.; Wohlgemuth, T.; Bače, R.; Fleischer, P.; Svoboda, M. Salvage-Logging after Windstorm Leads to Structural and Functional Homogenization of Understory Layer and Delayed Spruce Tree Recovery in Tatra Mts., Slovakia. Forests 2017, 8, 88. [Google Scholar] [CrossRef]
- Döbert, T.F.; Webber, B.L.; Sugau, J.B.; Dickinson, K.J.M.; Didham, R.K. Logging increases the functional and phylogenetic dispersion of understorey plant communities in tropical lowland rain forest. J. Ecol. 2017, 105, 1235–1245. [Google Scholar] [CrossRef]
- Eler, K.; Kermavnar, J.; Marinšek, A.; Kutnar, L. Short-term changes in plant functional traits and understory functional diversity after logging of different intensities: A temperate fir-beech forest experiment. Ann. For. Res. 2018, 61, 223–241. [Google Scholar] [CrossRef]
- Godefroid, S.; Koedam, N. Interspecific variation in soil compaction sensitivity among forest floor species. Biol. Conserv. 2004, 119, 207–217. [Google Scholar] [CrossRef]
- Crawford, L.J.; Heinse, R.; Kimsey, M.J.; Page-Dumroese, D.S. Soil Sustainability and Harvest Operations: A Review; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2021. [Google Scholar]
- Mohieddinne, H.; Brasseur, B.; Gallet-Moron, E.; Lenoir, J.; Kobaissi, A.; Horen, H. Assessment of soil compaction and rutting in managed forests through an airborne LiDAR technique. Land Degrad. Dev. 2023, 34, 1558–1569. [Google Scholar] [CrossRef]
- DeArmond, D.; Ferraz, J.B.S.; Higuchi, N. Natural recovery of skid trails: A review. Can. J. For. Res. 2021, 51, 948–961. [Google Scholar] [CrossRef]
- Naghdi, R.; Bagheri, I.; Basiri, R. Soil disturbances due to machinery traffic on steep skid trail in the north mountainous forest of Iran. J. For. Res. 2010, 21, 497–502. [Google Scholar] [CrossRef]
- Martiník, A.; Matoušková, M. Effect of thinning in birch stands: A review. For Ecol. Manag. 2025, 585, 122675. [Google Scholar] [CrossRef]
- Jahani, A. Risk assessment of forestry project implementation on spatial density changes of understory vegetation in forests using artificial neural network modeling approach. J. Spat. Anal. Environ. Hazards 2019, 6, 21–34. [Google Scholar]
- Lotfalian, M.; Riahifar, N.; Fallah, A.; Hodjati, S.M. Effects of roads on understory plant communities in a broadleaved forest in Hyrcanian zone. J. For. Sci. 2012, 58, 446–455. [Google Scholar] [CrossRef]
- Lee, M.; Manning, P.; Rist, J.; Power, S.A.; Marsh, C. A global comparison of grassland biomass responses to CO2 and nitrogen enrichment. Philos. Trans. R. Soc. B 2010, 365, 2047–2056. [Google Scholar] [CrossRef]
- Ghasemi Aghbash, F.; Abdi, A.; Heidari, M. Investigation of the effects of forest roads on the forest ecosystems of Iranian oak in terms of understory vegetation diversity and soil physical and chemical characteristics. J. Plant Ecosyst. Prot. 2018, 6, 59–76. [Google Scholar]
- Salehi, A.; Abkenar, K.; Basiri, R. Evaluation of soil physical properties recovery and natural regeneration establishment in skidding tracks (Case study: Now Asalem forest watershed). Iran. J. For. 2011, 3, 317–329. [Google Scholar]
- Zeng, S.L.; Zhang, T.T.; Gao, Y.; Li, B.; Fang, C.M.; Flory, S.L.; Zhao, B. Road effects on vegetation composition in a saline environment. J. Plant Ecol. 2011, 5, 206–218. [Google Scholar] [CrossRef]
- Cambi, M.; Certini, G.; Neri, F.; Marchi, E. The impact of heavy traffic on forest soils: A review. For. Ecol. Manag. 2015, 338, 124–138. [Google Scholar] [CrossRef]
- Naaf, T.; Kolk, J. Initial site conditions and interactions between multiple drivers determine herb-layer changes over five decades in temperate forests. For. Ecol. Manag. 2016, 366, 153–165. [Google Scholar] [CrossRef]
- Baeten, L.; Bauwens, B.; de Schrijver, A.; de Keersmaeker, L.; van Calster, H.; Vandekerkhove, K.; Roelandt, B.; Beeckman, H.; Verheyen, K. Herb layer changes (1954–2000) related to the conversion of coppice-with-standards forest and soil acidification. Appl. Veg. Sci. 2009, 12, 187–197. [Google Scholar] [CrossRef]
- Waite, S. Statistical Ecology in Practice: A Guide to Analyzing Environmental and Ecological Field Data; Prentice Hall Addison-Wesley Longman Ltd.: Boston, MA, USA, 1984. [Google Scholar]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Jourgholami, M.; Labelle, E.R.; Feghhi, J. Response of Runoff and Sediment on Skid Trails of Varying Gradient and Traffic Intensity over a Two-Year Period. Forests 2017, 8, 472. [Google Scholar] [CrossRef]
- Jourgholami, M.; Nasirian, A.; Labelle, E.R. Ecological Restoration of Compacted Soil Following the Application of Different Leaf Litter Mulches on the Skid Trail over a Five-Year Period. Sustainability 2018, 10, 2148. [Google Scholar] [CrossRef]
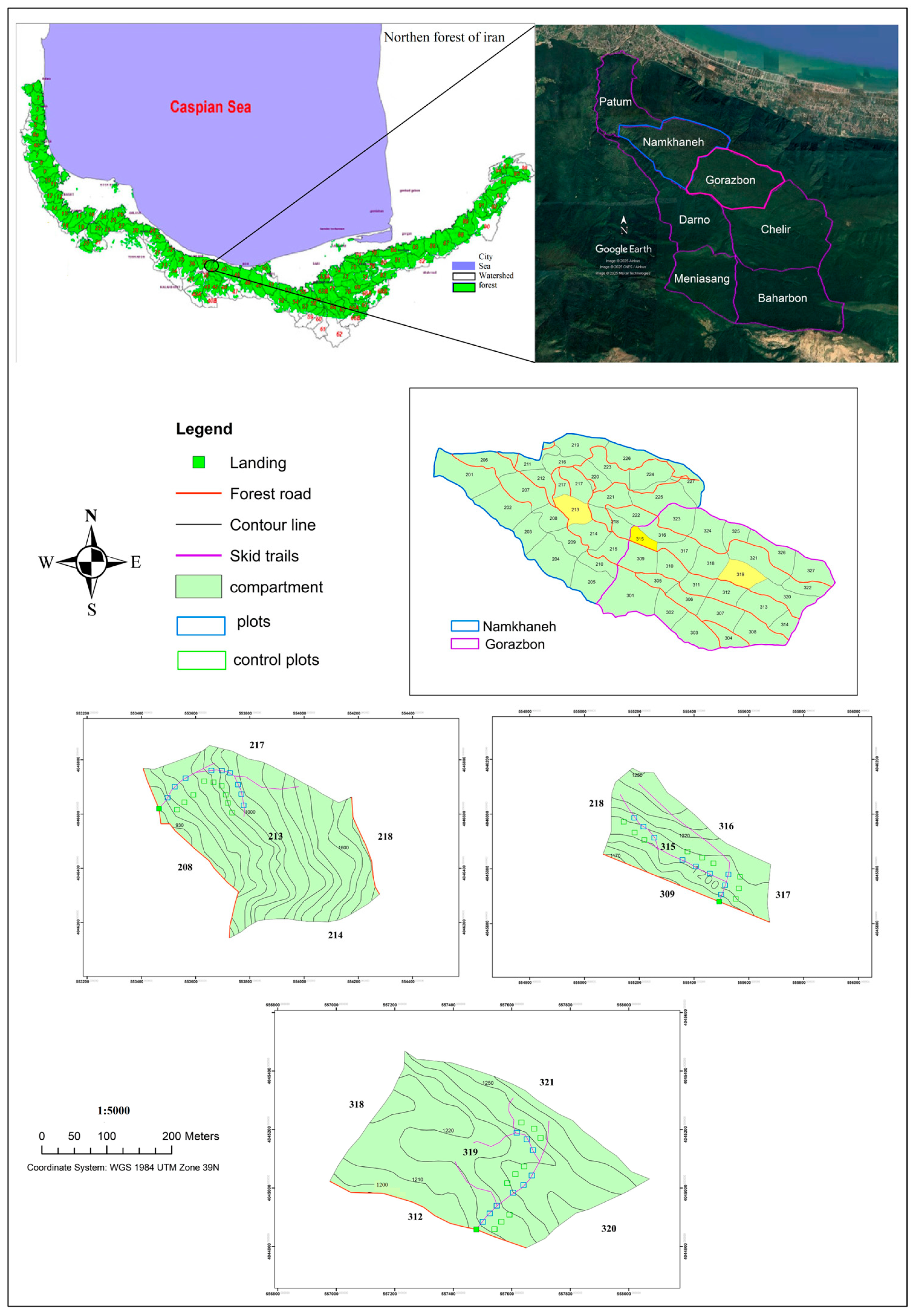

{kind=link}
{kind=link}
| Compartment No. | 213 | 315 | 319 |
|---|---|---|---|
| District | Namkhaneh | Gorazbon | Gorazbon |
| Time since harvest (years) | 15 | 5 | 10 |
| Forest type | Beech–hornbeam | Beech–hornbeam | Beech–hornbeam |
| Elevation (meters above sea level) | 980 | 1195 | 1230 |
| Average annual rainfall (mm) | 1081 | 1380 | 1380 |
| Soil type | Alfisol, classified as washed-brown forest soil with a loam–clay loam texture and a depth of 100 cm | Alfisol | Inceptisol with a depth of 145 cm |
| Main herbaceous plant species | Blackberry, holly, Asperula, spurge, violet, grasses, and clematis | Blackberry, holly, Asperula, Sanicula, male fern, mercury grass, spurge, and grasses | Blackberry and spurge, holly, Asperula, Sanicula, violet, and male ferns |
| General aspect | West to southwest | South to southwest | North and south |
| Slope (%) | 28 | 25 | 22 |
| Area (ha) | 34.7 | 20.03 | 42.74 |
| Scientific Name | Family | Biological Form | Chorotype | Growth Form | Number |
|---|---|---|---|---|---|
| Asperula odorata L. | Rubiaceae | Geo | PL | Broadleaf herbaceous | 196 |
| Brachypodium sylvaticum (Huds) | Gramineae | Hem | PL | Grass | 17 |
| Carex pendula L. | Cyperaceae | Geo | ES | Grass | 102 |
| Circaea lutetiana L. | Onagraceae | Geo | ES, IT, M | Broadleaf herbaceous | 4 |
| Drypotris affines L. | Aspidiaceae | Geo | ES | Fern | 39 |
| Epimedium pinnatum L. | Podophyllaceae | Geo | Hyr | Broadleaf herbaceous | 2 |
| Epipactis helleborine (L) Crantz | Orchidaceae | Geo | PL | Broadleaf herbaceous | 2 |
| Euphorbia amygdaloides L. | Euphorbiaceae | Geo | ES | Broadleaf herbaceous | 269 |
| Geranium sp. | Geraniaceae | Hem | ES | Broadleaf herbaceous | 1 |
| Hypericum androsaemum L. | Hypericaceae | Cha | ES | Broadleaf herbaceous | 41 |
| Lamium album L. | Lamiaceae/labiatae | Hem | E, IT | Broadleaf herbaceous | 23 |
| Lapsana communis L. | Asteraceae-Compositae | Th | E, IT | Broadleaf herbaceous | 2 |
| Luzula pilosa (Smith) DC. | Juncaceae | Geo | ES, M | Grass | 5 |
| Menta aquatic L. | Lamiaceae/labiatae | Hem | IT | Broadleaf herbaceous | 1 |
| Mentha sp. | Lamiaceae/labiatae | Hem | IT | Broadleaf herbaceous | 24 |
| Mercurialis perennis L. | Euphorbiaceae | Geo | ES, M | Broadleaf herbaceous | 96 |
| Oplismenus undulatifolius (Ard) | Gramineae | Hem | ES, M | Grass | 20 |
| Poa sp. | Gramineae | Th | ES, IT | Grass | 7 |
| Potentilla reptans L. | Rosaceae | Hem | ES, IT | Broadleaf herbaceous | 52 |
| Rubus hyrcanus Juz. | Rosaceae | Ph | ES | Broadleaf herbaceous | 33 |
| Sanicula europaea L. | Umbelliferae | Hem | PL | Broadleaf herbaceous | 5 |
| Scutellaria tournefortii Beneth. | Lamiaceae/labiatae | Th | IT | Broadleaf herbaceous | 4 |
| Solanum kieseritzkii C.A. Mey. | Solanaceae | Cha | ES | Broadleaf herbaceous | 30 |
| Viola alba Bess. | Violaceae | Hem | ES | Broadleaf herbaceous | 366 |
| Viola odorata L. | Violaceae | Hem | ES-M | Broadleaf herbaceous | 64 |
| Species | Skid Trail | Skid Trail Percentage | Control | Control Percentage |
|---|---|---|---|---|
| Asperula odorata L. | 134 | 68.37 | 62 | 31.63 |
| Brachypodium sylvaticum (Huds) | 0 | 0 | 17 | 100 |
| Carex pendula L. | 68 | 66.67 | 34 | 33.33 |
| Circaea lutetiana L. | 4 | 100 | 0 | 0 |
| Drypotris affines L. | 12 | 30.77 | 27 | 69.23 |
| Epimedium pinnatum L. | 0 | 0 | 2 | 100 |
| Epipactis helleborine (L) Crantz | 2 | 100 | 0 | 0 |
| Euphorbia amygdaloides L. | 86 | 31.97 | 183 | 68.03 |
| Geranium sp. | 2 | 100 | 0 | 0 |
| Hypericum androsaemum L. | 7 | 17.07 | 34 | 82.93 |
| Lamium album L. | 6 | 26.09 | 17 | 73.91 |
| Lapsana communis L. | 2 | 100 | 0 | 0 |
| Luzula pilosa (Smith) DC | 0 | 0 | 5 | 100 |
| Menta aquatica L. | 2 | 100 | 0 | 0 |
| Mentha sp. | 15 | 62.5 | 9 | 37.5 |
| Mercurialis perennis L. | 24 | 25 | 72 | 75 |
| Oplismenus undulatifolius (Ard) | 0 | 0 | 20 | 100 |
| Poa sp. | 7 | 100 | 0 | 0 |
| Potentilla reptans L. | 33 | 63.46 | 19 | 36.54 |
| Rubus hyrcanus Juz. | 5 | 15.15 | 28 | 84.85 |
| Sanicula europaea L. | 1 | 20 | 4 | 80 |
| Scutellaria tournefortii Beneth. | 4 | 100 | 0 | 0 |
| Solanum kieseritzkii C.A. Mey. | 15 | 50 | 15 | 50 |
| Viola alba Bess. | 165 | 45.08 | 201 | 54.92 |
| Viola odorata L. | 64 | 100 | 0 | 0 |
| Total | 656 | 46.69 | 749 | 53.31 |
| Herbaceous Species | n° S-T | n° C | p Value | Result |
|---|---|---|---|---|
| Asperula odorata L. | 14.9 ± 13.0 | 6.9 ± 9.5 | 0.08 | n.s |
| Carex pendula L. | 7.8 ± 6.3 | 3.6 ± 4.5 | 0.09 | n.s |
| Euphorbia amygdaloides L. | 9.6 ± 20.3 | 20.3 ± 13.1 | 0.94 | n.s |
| Mercurialis perennis L. | 2.7 ± 4.8 | 8.0 ± 17.8 | 0.12 | n.s |
| Potentilla reptans L. | 3.7 ± 5.4 | 2.1 ± 4.2 | 0.56 | n.s |
| Sanicula europaea L. | 0.1 ± 0.3 | 0.4 ± 1.0 | 0.07 | n.s |
| Solanum kieseritzkii C.A. Mey. | 1.7 ± 4.0 | 1.7 ± 2.3 | 0.59 | n.s |
| Viola alba Bess. | 7.1 ± 8.7 | 0.0 ± 0.0 | 0.99 | n.s |
| Brachypodium sylvaticum (Huds). | 0.0 ± 0.0 | 1.9 ± 5.7 | 0.04 | * |
| Circaea lutetiana L. | 0.4 ± 0.9 | 0.0 ± 0.0 | 0 | * |
| Drypotris affines L. | 1.3 ± 1.0 | 3.0 ± 3.5 | 0.05 | * |
| Epimedium pinnatum L. | 0.0 ± 0.0 | 0.2 ± 0.7 | 0.04 | * |
| Epipactis helleborine (L) Crantz. | 0.2 ± 0.7 | 0.0 ± 0.0 | 0.04 | * |
| Geranium sp. | 0.1 ± 0.3 | 0.0 ± 0.0 | 0.04 | * |
| Hypericum androsaemum L. | 0.8 ± 1.7 | 3.8 ± 5.5 | 0 | * |
| Lamium album L. | 0.7 ± 1.4 | 1.9 ± 3.0 | 0 | * |
| Lapsana communis L. | 0.2 ± 0.4 | 0.0 ± 0.0 | 0 | * |
| Luzula pilosa (Smith) DC. | 0.0 ± 0.0 | 0.2 ± 0.7 | 0.04 | * |
| Menta aquatica L. | 0.1 ± 0.3 | 0.0 ± 0.0 | 0.04 | * |
| Mentha sp. | 1.7 ± 2.8 | 1.0 ± 2.1 | 0.02 | * |
| Oplismenus undulatifolius (Ard). | 0.0 ± 0.0 | 2.2 ± 6.7 | 0.02 | * |
| Poa sp. | 0.8 ± 2.3 | 0.0 ± 0.0 | 0.04 | * |
| Rubus hyrcanus Juz. | 0.6 ± 1.1 | 3.1 ± 4.1 | 0 | * |
| Scutellaria tournefortii Beneth. | 0.1 ± 0.3 | 0.0 ± 0.0 | 0 | * |
| Viola odorata L. | 7.1 ± 8.7 | 0.0 ± 0.0 | 0 | * |
| Time | 15 Years Since Harvest (Compartment no. 213) | 10 Years Since Harvest (Compartment no. 319) | 5 Years Since Harvest (Compartment no. 315) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Traffic/Species | Low | Medium | High | Low | Medium | High | Low | Medium | High |
| Asperula odorata | 0.02 * | 0.03 * | 0.02 * | 0.88 | 0.08 | 0.02 * | 0.53 | 0.05 * | 1 |
| Carex pendula | - | 0.02 * | 0.09 | 0.13 | 0.07 | 0.02 * | 0.12 | 0.15 | 0.07 |
| Euphorbia amygdaloides | 0.23 | 0.05* | 0.02 * | 0.29 | 0.09 | 0.02 * | 0.04 | 0.18 | 0.05 * |
| Mercurialis perennis | - | - | 0.02 * | 0.05 * | 0.89 | 0.02 * | - | - | 0.02 * |
| Viola alba | 0.02 * | 0.29 | 0.12 | 0.02 * | 0.07 | 0.05 * | 0.04 | 0.56 | 0.02 * |
| Comparison of Statistical Results for the Traffic Intensity Factor for all Herbaceous Plants with Those for the Control | ||
|---|---|---|
| 1 | Low traffic intensity (average n° S-T 30.6 ± 21.5; average n° C 30.9 ± 19.3) | 0.62 |
| 2 | Medium traffic intensity (average n° S-T 17.6 ± 9.7; average n° C 24.7 ± 14.2) | 0.40 |
| 3 | High traffic intensity (average n° S-T 19.3 ± 9.7; average n° C 33.1 ± 18.9) | 0.02 * |
| Comparison of statistical results between traffic intensity levels | ||
| 1 | Low traffic intensity (average n° S-T 30.6 ± 21.5) vs. medium traffic intensity (average n° S-T 17.6 ± 9.7) | 0.93 |
| 2 | Low traffic intensity (average n° S-T 30.6 ± 21.5) vs. high traffic intensity (average n° S-T 19.3 ± 9.7) | 0.00 * |
| 3 | Medium traffic intensity (average n° S-T 17.6 ± 9.7) vs. high traffic intensity (average n° S-T 19.3 ± 9.7) | 0.00 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babaei-Ahmadabad, A.; Jourgholami, M.; Lo Monaco, A.; Venanzi, R.; Picchio, R. Mid-Term Evaluation of Herbaceous Cover Restoration on Skid Trails Following Ground-Based Logging in Pure Oriental Beech (Fagus orientalis Lipsky) Stands of the Hyrcanian Forests, Northern Iran. Land 2025, 14, 1387. https://doi.org/10.3390/land14071387
Babaei-Ahmadabad A, Jourgholami M, Lo Monaco A, Venanzi R, Picchio R. Mid-Term Evaluation of Herbaceous Cover Restoration on Skid Trails Following Ground-Based Logging in Pure Oriental Beech (Fagus orientalis Lipsky) Stands of the Hyrcanian Forests, Northern Iran. Land. 2025; 14(7):1387. https://doi.org/10.3390/land14071387
Chicago/Turabian StyleBabaei-Ahmadabad, Ali, Meghdad Jourgholami, Angela Lo Monaco, Rachele Venanzi, and Rodolfo Picchio. 2025. "Mid-Term Evaluation of Herbaceous Cover Restoration on Skid Trails Following Ground-Based Logging in Pure Oriental Beech (Fagus orientalis Lipsky) Stands of the Hyrcanian Forests, Northern Iran" Land 14, no. 7: 1387. https://doi.org/10.3390/land14071387
APA StyleBabaei-Ahmadabad, A., Jourgholami, M., Lo Monaco, A., Venanzi, R., & Picchio, R. (2025). Mid-Term Evaluation of Herbaceous Cover Restoration on Skid Trails Following Ground-Based Logging in Pure Oriental Beech (Fagus orientalis Lipsky) Stands of the Hyrcanian Forests, Northern Iran. Land, 14(7), 1387. https://doi.org/10.3390/land14071387