
_Kazoglou.png)
Interseeded Native Forbs Resilient Under Variable Grazing Regimen



Abstract
1. Introduction
2. Materials and Methods
2.1. Pasture Establishment
2.2. Pasture Treatments
2.3. Temperature and Precipitation
2.4. Cattle Management
2.5. NWSG and Forb Characteristics
2.6. Forage Mass
2.7. Forage Nutritive Composition
2.8. Statistical Analysis
3. Results
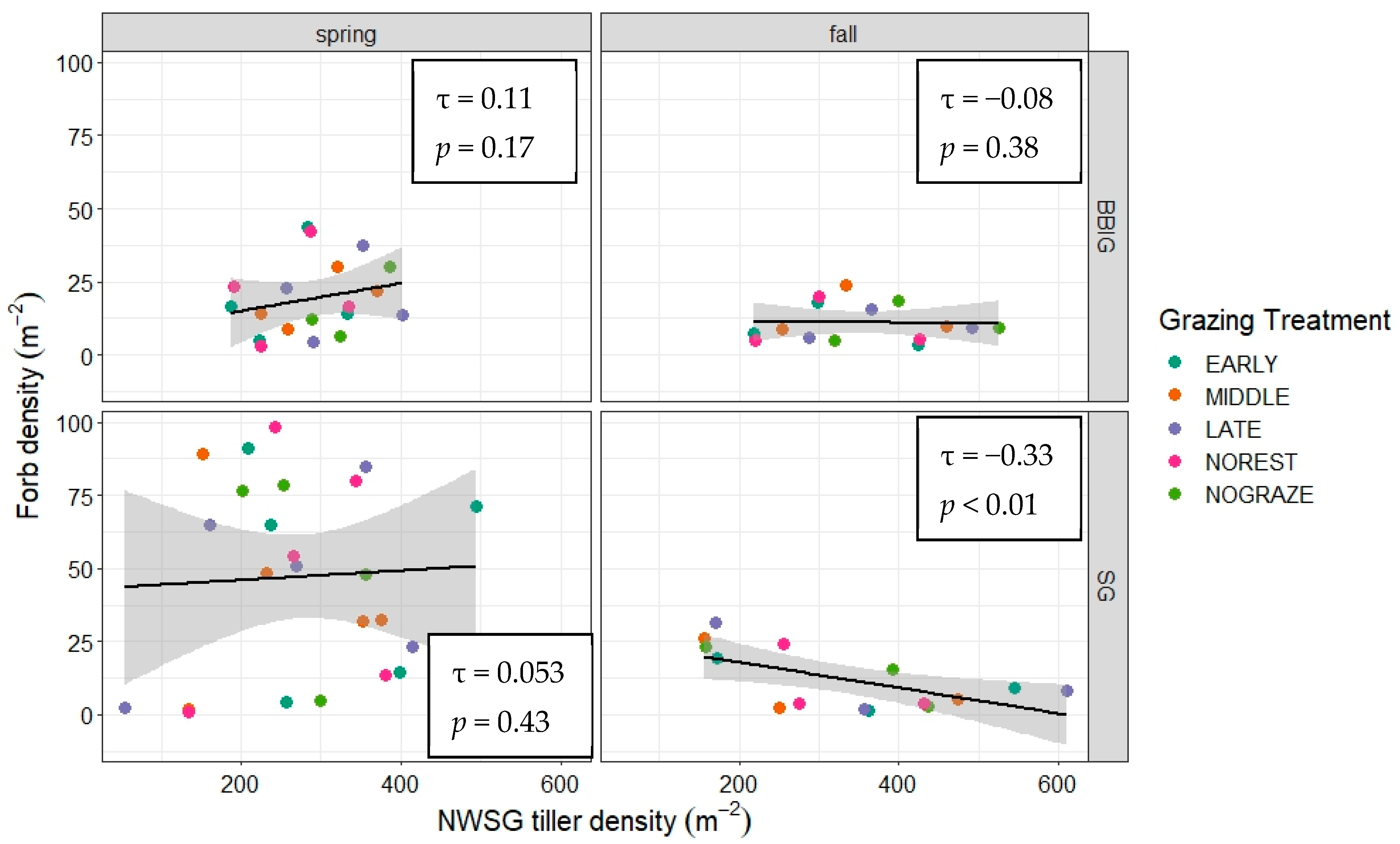
3.1. NWSG and Forb Characteristics
3.2. Forage Mass
3.3. Forage Nutritive Composition
4. Discussion
4.1. NWSG and Forb Characteristics
4.2. Forage Mass
4.3. Forage Nutritive Composition
4.4. Sustainability and Environmental Implications
4.5. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keyser, P.D.; Holcomb, E.D.; Lituma, C.M.; Bates, G.E.; Waller, J.C.; Boyer, C.N.; Travis Mulliniks, J. Forage Attributes and Animal Performance from Native Grass Inter-Seeded with Red Clover. Agron. J. 2016, 108, 373–383. [Google Scholar] [CrossRef]
- Backus, W.M.; Waller, J.C.; Bates, G.E.; Harper, C.A.; Saxton, A.; McIntosh, D.W.; Birckhead, J.; Keyser, P.D. Management of Native Warm-Season Grasses for Beef Cattle and Biomass Production in the Mid-South USA. J. Anim. Sci. 2017, 95, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Brazil, K.A.; Keyser, P.D.; Bates, G.E.; Saxton, A.M.; Holcomb, E.D. Continuous Grazing of Mixed Native Warm-Season Grass in the Fescue Belt. Agron. J. 2020, 112, 5067–5080. [Google Scholar] [CrossRef]
- Rushing, J.B.; Maples, J.G.; Rivera, J.D.; Lyles, J.C. Early-Season Grazing of Native Grasses Offers Potential Profitable Benefit. Agron. J. 2020, 112, 1057–1067. [Google Scholar] [CrossRef]
- Tracy, B.F.; Maughan, M.; Post, N.; Faulkner, D.B. Integrating Annual and Perennial Warm-Season Grasses in a Temperate Grazing System. Crop Sci. 2010, 50, 2171–2177. [Google Scholar] [CrossRef]
- Temu, V.W.; Rude, B.J.; Baldwin, B.S. Yield Response of Native Warm-Season Forage Grasses to Harvest Intervals and Durations in Mixed Stands. Agronomy 2014, 4, 90–107. [Google Scholar] [CrossRef]
- Rauzi, F.; Hanson, C.L. Water Intake and Runoff as Affected by Intensity of Grazing. J. Range Manag. 1966, 19, 351–356. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Milcu, A.; Sabais, A.C.W.; Bessler, H.; Brenner, J.; Engels, C.; Klarner, B.; Maraun, M.; Partsch, S.; Roscher, C.; et al. Plant Diversity Surpasses Plant Functional Groups and Plant Productivity as Driver of Soil Biota in the Long Term. PLoS ONE 2011, 6, e16055. [Google Scholar] [CrossRef]
- Aubault, H.; Webb, N.P.; Strong, C.L.; McTainsh, G.H.; Leys, J.F.; Scanlan, J.C. Grazing Impacts on the Susceptibility of Rangelands to Wind Erosion: The Effects of Stocking Rate, Stocking Strategy and Land Condition. Aeolian Res. 2015, 17, 89–99. [Google Scholar] [CrossRef]
- National Research Council Committee on the Status of Pollinators in North America. Status of Pollinators in North America; National Academy Press: Washington, DC, USA, 2007. [Google Scholar]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- van Engelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony Collapse Disorder: A Descriptive Study. PLoS ONE 2009, 4, e6481. [Google Scholar] [CrossRef] [PubMed]
- Shackelford, G.; Steward, P.R.; Benton, T.G.; Kunin, W.E.; Potts, S.G.; Biesmeijer, J.C.; Sait, S.M. Comparison of Pollinators and Natural Enemies: A Meta-Analysis of Landscape and Local Effects on Abundance and Richness in Crops. Biol. Rev. 2013, 88, 1002–1021. [Google Scholar] [CrossRef] [PubMed]
- Weiner, C.N.; Werner, M.; Linsenmair, K.E.; Blüthgen, N. Land-Use Impacts on Plant-Pollinator Networks: Interaction Strength and Specialization Predict Pollinator Declines. Ecology 2014, 95, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Grab, H.; Branstetter, M.G.; Amon, N.; Urban-Mead, K.R.; Park, M.G.; Gibbs, J.; Blitzer, E.J.; Poveda, K.; Loeb, G.; Danforth, B.N. Agriculturally Dominated Landscapes Reduce Bee Phylogenetic Diversity and Pollination Services. Science 2019, 363, 282–284. [Google Scholar] [CrossRef]
- Keyser, P.D.; Buehler, D.A.; Fike, J.H.; Finke, D.L.; Fuhlendorf, S.D.; Martin, J.A.; Naumann, H.D.; Smith, S.R. The Birds and the Bees: Producing Beef and Conservation Benefits on Working Grasslands. Agronomy 2022, 12, 1934. [Google Scholar] [CrossRef]
- Richwine, J.D.; Keyser, P.D.; Zobel, J.M.; Walker, F.R. Native Forbs Interseeded into Native Grass Pastures Persist under Grazing. Grass Forage Sci. 2024, 79, 328–341. [Google Scholar] [CrossRef]
- Locke, J.C.; French, J.T.; Gonzalez, C.E.; Harveson, L.A.; Warnock, B.J.; Gray, S.S. The Effects of Continuous and Rotational Livestock Grazing on Forb Quality and Quantity: Implications for Pronghorn Habitat Management. Rangel. Ecol. Manag. 2021, 77, 75–81. [Google Scholar] [CrossRef]
- McIntosh, D.; Aderson-Husmoen, B.J.; Kern-Lunbery, R.; Goldblatt, P.; Lemus, R.; Griggs, T.; Bauman, L.; Boone, S.; Shewmaker, G.; Teutsch, C. Guidelines for Optimal Use of NIRSC Forage and Feed Calibrations in Membership Laboratories, 2nd ed.; The University of Tennessee Press: Knoxville, TN, USA, 2022. [Google Scholar]
- McIntosh, D.; Bates, G.; Berghazi, P.; Labbe, N.; Eckelkamp, E.; Lemus, R.; Anderson, B. The Impact of Moisture Content on Forage Nutritive Predictions Using near Infrared Spectroscopy (NIRS) on Prepared Ground Samples and How This Affects Producers and Researchers. In Proceedings of the Western Alfalfa and Hay Symposium, Reno, NV, USA, 19–21 November 2019. [Google Scholar]
- Matthew, C.; Hernandez-Garay, A.; Hodgson, J. Making Sense of the Link between Tiller Density and Pasture Production. In Proceedings of the New Zealand Grassland Association; New Zealand Grassland Association: Timaru, New Zealand, 1996; pp. 83–87. [Google Scholar]
- Prigge, J.L.; Bisangwa, E.; Richwine, J.D.; Swilling, K.J.; Keyser, P.D. Blooming and Forage Characteristics of Twelve Native Forbs Subjected to Repeated Defoliation. Agronomy 2023, 14, 28. [Google Scholar] [CrossRef]
- Kulakow, P.A. Variation in Illinois Bundleflower (Desmanthus Illinoensis (Michaux) MacMillan): A Potential Perennial Grain Legume. Euphytica 1999, 110, 7–20. [Google Scholar] [CrossRef]
- Harris, C.; Balfour, N.J.; Ratnieks, F.L.W. Seasonal Variation in the General Availability of Floral Resources for Pollinators in Northwest Europe: A Review of the Data. Biol. Conserv. 2024, 298, 110774. [Google Scholar] [CrossRef]
- Ravetto Enri, S.; Probo, M.; Farruggia, A.; Lanore, L.; Blanchetete, A.; Dumont, B. A Biodiversity-Friendly Rotational Grazing System Enhancing Flower-Visiting Insect Assemblages While Maintaining Animal and Grassland Productivity. Agric. Ecosyst. Environ. 2017, 241, 1–10. [Google Scholar] [CrossRef]
- Jurado, E.; Flores, J. Is Seed Dormancy under Environmental Control or Bound to Plant Traits? J. Veg. Sci. 2005, 16, 559–564. [Google Scholar] [CrossRef]
- Sheley, R.L.; Half, M.L. Enhancing Native Forb Establishment and Persistence Using a Rich Seed Mixture. Restor. Ecol. 2006, 14, 627–635. [Google Scholar] [CrossRef]
- Watson, B.L.; Lukas, S.B.; Morris, L.R.; DeBano, S.J.; Schmalz, H.J.; Leffler, A.J. Forb Community Response to Prescribed Fire, Livestock Grazing, and an Invasive Annual Grass in the Pacific Northwest Bunchgrass Prairie. Appl. Veg. Sci. 2021, 24, e12619. [Google Scholar] [CrossRef]
- Sicangco, C.K.; Bavdekar, S.; Subhash, G.; Putz, F.E. Active Space Garnering by Leaves of a Rosette Plant. Curr. Biol. 2022, 32, R352–R353. [Google Scholar] [CrossRef]
- Maccherini, S.; Santi, E. Long-Term Experimental Restoration in a Calcareous Grassland: Identifying the Most Effective Restoration Strategies. Biol. Conserv. 2012, 146, 123–135. [Google Scholar] [CrossRef]
- Crider, F.J. Root-Growth Stoppage Resulting from Defoliation of Grass; No. 1102; US Department of Agriculture: Washington, DC, USA, 1955.
- Vermeire, L.T.; Russell, M.L. Seasonal Timing of Fire Alters Biomass and Species Composition of Northern Mixed Prairie. Rangel. Ecol. Manag. 2018, 71, 714–720. [Google Scholar] [CrossRef]
- Buxton, D.R. Quality-Related Characteristics of Forages as Influenced by Plant Environment and Agronomic Factors. Anim. Feed Sci. Technol. 1996, 59, 37–49. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Beef Cattle, 8th ed.; National Academy Press: Washington, DC, USA, 2016; ISBN 9780309317023. [Google Scholar]
- Zou, C.; Wang, K.; Wang, T.; Xu, W. Overgrazing and Soil Carbon Dynamics in Eastern Inner Mongolia of China. Ecol. Res. 2007, 22, 135–142. [Google Scholar] [CrossRef]
- Liu, M.; Ouyang, S.; Tian, Y.; Wen, S.; Zhao, Y.; Li, X.; Tao, B.Y.; Kuzyakov, Y.; Xu, X. Effects of Rotational and Continuous Overgrazing on Newly Assimilated C Allocation. Biol. Fertil. Soils 2021, 57, 193–202. [Google Scholar] [CrossRef]
- Tracy, B.F.; Sanderson, M.A. Forage Productivity, Species Evenness and Weed Invasion in Pasture Communities. Agric. Ecosyst. Environ. 2004, 102, 175–183. [Google Scholar] [CrossRef]
- Tracy, B.F.; Bauer, R.B. Evaluating Mob Stocking for Beef Cattle in a Temperate Grassland. PLoS ONE 2019, 14, e0226360. [Google Scholar] [CrossRef] [PubMed]
- Billman, E.D.; Williamson, J.A.; Soder, K.J.; Andreen, D.M.; Skinner, R.H. Mob and Rotational Grazing Influence Pasture Biomass, Nutritive Value, and Species Composition. Agron. J. 2020, 112, 2866–2878. [Google Scholar] [CrossRef]
- Prigge, J.L.; Bisangwa, E.; Richwine, J.D.; Sykes, V.R.; Ivey, J.L.Z.; Keyser, P.D. Native Forbs Provide Pollinator Resources and Improve Forage Nutrient Composition, Animal Performance, and Pasture Productivity. Agronomy 2024, 14, 2184. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Species | Abbreviation | Seeding Rate (PLS kg ha−1) | SEEDS m−2 * |
|---|---|---|---|---|
| Black-eyed Susan | Rudbeckia hirta L. | BESU | 0.56 | 194 |
| Eastern purple coneflower | Echinacea purpurea (L.) Moench | PURC | 0.7 | 18 |
| Illinois bundleflower § | Desmanthus illinoensis (Michx.) MacMill. | ILBF | 1.26 | 23 |
| Lanceleaf coreopsis | Coreopsis lanceolata L. | LCOR | 1.12 | 54 |
| Maximilian sunflower | Helianthus maximiliani Schrad. | MSUN | 0.56 | 308 |
| Oxeye sunflower | Heliopsis helianthoides (L.) Sweet | OSUN | 0.28 | 07 |
| Partridge pea § | Chamaecrista fasciculata (Michx.) Greene | PPEA | 0.56 | 08 |
| Plains coreopsis | Coreopsis tinctoria Nutt. | PLAC | 0.56 | 392 |
| Purple prairie clover § | Dalea purpurea (Vent.) Rydb. | PUPC | 0.56 | 37 |
| Dixie ticktrefoil § | Desmodium tortuosum (Sw.) DC. | TTFL | 0.56 | 17 |
| Upright prairie coneflower | Ratibida columnifera (Nutt.) Woot & Standl. | UPPC | 0.28 | 46 |
| Year | ||||||
|---|---|---|---|---|---|---|
| Grazing Treatment | 2022 § | |||||
| 2018 | 2019 | 2020 | 2021 | SG | BBIG | |
| Early rest * | 31 May to 20 Jun | 30 May to 19 Jun | 9 Jun to 26 Jun | 3 Jun to 23 Jun | 12 May to 2 Jun | 1 Jun to 22 Jun |
| Middle rest | 21 Jun to 11 Jul | 20 Jun to 10 Jul | 27 Jun to 17 Jul | 24 Jun to 15 Jul | 3 Jun to 23 Jun | 23 Jun to 13 Jul |
| Late rest | 12 Jul to 2 Aug | 12 Jul to 31 Jul | 18 Jul to 7 Aug | 16 Jul to 5 Aug | 24 Jun to 14 Jul | 14 Jul to 3 Aug |
| No rest | 17 May to 15 Aug | 16 May to 14 Aug | 28 May to 17 Aug | 19 May to 20 Aug | 28 Apr to 29 Jul | 18 May to 18 Aug |
| No graze | 17 May to 15 Aug | 16 May to 14 Aug | 28 May to 17 Aug | 19 May to 20 Aug | 28 Apr to 29 Jul | 18 May to 18 Aug |
| Total grazing days | 91 days | 91 days | 82 days | 94 days | 93 days | 93 days |
| Species | Abundance (Total Plants) * | 2022 Plants m−2 § | Abundance Rank | 2022 Plants Rank (Persistence) | Flowering Window Rank ‡ | Grazing Rank ⁑ | Rank Average | Overall Rank ⁂ | |
|---|---|---|---|---|---|---|---|---|---|
| SG | BESU | 1550 | 0.8 | 2 | 2 | 1 | 6 | 2.8 | 1 |
| LCOR | 2639 | 0 | 1 | 5 | 3 | 3 | 3.0 | 2 | |
| OSUN | 305 | 0.8 | 5 | 2 | 4 | 2 | 3.3 | 3 | |
| PURC | 364 | 0.2 | 4 | 4 | 2 | 4 | 3.5 | 4 | |
| TTFL | 245 | 0.9 | 6 | 1 | 8 | 1 | 4.0 | 5 | |
| MSUN | 112 | 0.4 | 7 | 3 | 5 | 5 | 5.0 | 6 | |
| PLAC | 741 | 0.2 | 3 | 4 | 6 | 7 | 5.0 | 7 | |
| PPEA | 32 | 0 | 8 | 5 | 7 | 7 | 6.8 | 8 | |
| UPPC | 20 | 0 | 9 | 5 | 9 | 7 | 7.5 | 9 | |
| ILBF | 5 | 0 | 10 | 5 | 10 | 7 | 8.0 | 10 | |
| PUPC | 2 | 0 | 11 | 5 | 10 | 7 | 8.3 | 11 | |
| BBIG | TTFL | 762 | 3.1 | 3 | 1 | 6 | 1 | 2.8 | 1 |
| PURC | 243 | 0.9 | 6 | 2 | 1 | 3 | 3.0 | 2 | |
| LCOR | 937 | 0.4 | 1 | 4 | 4 | 4 | 3.3 | 3 | |
| BESU | 828 | 0.8 | 2 | 3 | 2 | 6 | 3.3 | 4 | |
| OSUN | 311 | 0.8 | 5 | 3 | 3 | 2 | 3.3 | 5 | |
| MSUN | 55 | 0.2 | 8 | 5 | 5 | 5 | 5.8 | 6 | |
| PLAC | 341 | 0.1 | 4 | 6 | 7 | 7 | 6.0 | 7 | |
| UPPC | 63 | 0 | 7 | 7 | 8 | 7 | 7.3 | 8 | |
| ILBF | 12 | 0 | 9 | 7 | 10 | 7 | 8.3 | 9 | |
| PPEA | 6 | 0 | 10 | 7 | 9 | 7 | 8.3 | 10 | |
| PUPC | 0 | 0 | 11 | 7 | 10 | 7 | 8.8 | 11 |
| Predictor | F | df | p-Value | |
|---|---|---|---|---|
| Total forage mass | ||||
| SG | Grazing treatment | 9.34 | 3 | <0.01 |
| Month | 4.89 | 3 | <0.01 | |
| Year | 35.53 | 3 | <0.01 | |
| BBIG | Grazing treatment | 8.48 | 3 | <0.01 |
| Month | 22.71 | 3 | 0.03 | |
| Year | 3.06 | 3 | <0.01 | |
| Grazing treatment × Month * | 5.74 | 8 | <0.01 | |
| Grazing treatment × Year | 2.76 | 9 | <0.01 | |
| Forb forage mass | ||||
| SG & | Grazing treatment | 14.94 | 4 | <0.01 |
| BBIG | Month | 0.81 | 3 | 0.49 |
| Year | 19.84 | 3 | <0.01 | |
| Grazing treatment × Month | 4.19 | 12 | <0.01 | |
| Grazing treatment × Year | 6.91 | 12 | <0.01 | |
| CP | ||||
| SG | Grazing treatment | 2.68 | 3 | 0.05 |
| Month | 141.11 | 3 | <0.01 | |
| Year | 31.04 | 3 | <0.01 | |
| Month × Year | 13.97 | 9 | <0.01 | |
| BBIG | Grazing treatment | 4.22 | 3 | <0.01 |
| Month | 56.71 | 3 | <0.01 | |
| Year | 13.77 | 3 | <0.01 | |
| Month × Year | 11.13 | 9 | <0.01 | |
| NDF | ||||
| SG | Grazing treatment | 5.15 | 3 | <0.01 |
| Month | 130.41 | 3 | <0.01 | |
| Year | 40.73 | 3 | <0.01 | |
| Month × Year | 12.11 | 9 | <0.01 | |
| BBIG | Grazing treatment | 7.66 | 3 | <0.01 |
| Month | 39.71 | 3 | <0.01 | |
| Year | 3.60 | 3 | 0.01 | |
| Month × Year | 11.76 | 9 | <0.01 | |
| ADF | ||||
| SG | Grazing treatment | 2.10 | 3 | <0.01 |
| Month | 103.03 | 3 | <0.01 | |
| Year | 5.14 | 3 | <0.01 | |
| Month × year | 18.63 | 9 | <0.01 | |
| BBIG | Grazing treatment | 1.81 | 3 | 0.15 |
| Month | 29.40 | 3 | <0.01 | |
| Year | 10.19 | 3 | <0.01 | |
| Month × Year | 6.93 | 9 | <0.01 | |
| Grazing Treatment | Year | ||||
|---|---|---|---|---|---|
| 2018 | 2019 | 2021 | 2022 | ||
| –––––––––––––––––––––kg ha−1–––––––––––––––––––––– | |||||
| BBIG | Early rest | 2011 bcd⁑ | 2673 bcd | 7917 a | 1910 bcd |
| Middle rest | 1368 cd | 2586 bcd | 3502 b | 832 cd | |
| Late rest | 1372 cd | 2189 bcd | 3567 bc | 454 d | |
| No rest | 1398 bcd | 2422 bcd | 1957 bcd | 999 cd | |
| SG | Early rest | 4031 a§ | 4442 a | 5773 a | 1527 a |
| Middle rest | 2271 b | 2582 b | 3618 b | 565 b | |
| Late rest | 2323 b | 2637 b | 3683 b | 540 b | |
| No rest | 2017 b | 2310 b | 3295 b | 420 b | |
| Grazing Treatment | Year | |||
|---|---|---|---|---|
| 2018 | 2019 | 2021 | 2022 | |
| –––––––––––––––––––––kg ha−1–––––––––––––––––––––– | ||||
| Early rest | 2006 b⁑ | 825 cd | 227 cd | 0 cd |
| Middle rest | 1064 bcd | 870 bcd | 839 bcd | 167 cd |
| Late rest | 1058 bc | 520 cd | 1563 bc | 361 cd |
| No rest | 1078 bc | 1056 bc | 459 cd | 132 d |
| No graze | 3154 a | 951 c | 1052 bc | 798 cd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prigge, J.L.; Richwine, J.D.; Bisangwa, E.; Keyser, P.D. Interseeded Native Forbs Resilient Under Variable Grazing Regimen. Land 2025, 14, 989. https://doi.org/10.3390/land14050989
Prigge JL, Richwine JD, Bisangwa E, Keyser PD. Interseeded Native Forbs Resilient Under Variable Grazing Regimen. Land. 2025; 14(5):989. https://doi.org/10.3390/land14050989
Chicago/Turabian StylePrigge, Jessica L., Jonathan D. Richwine, Eric Bisangwa, and Patrick D. Keyser. 2025. "Interseeded Native Forbs Resilient Under Variable Grazing Regimen" Land 14, no. 5: 989. https://doi.org/10.3390/land14050989
APA StylePrigge, J. L., Richwine, J. D., Bisangwa, E., & Keyser, P. D. (2025). Interseeded Native Forbs Resilient Under Variable Grazing Regimen. Land, 14(5), 989. https://doi.org/10.3390/land14050989