
Soil Organic Carbon Content and Density in Response to Pika Outbreaks Along the Altitudinal Gradient in Alpine Meadows of the Qinghai–Tibet Plateau, West China



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Vegetation Survey
2.3. Soil Sampling and Analysis
2.4. Data Processing
3. Results and Analysis
3.1. Impacts of Plateau Pika Outbreaks on Vegetation and Soil Properties Along the Altitudinal Gradient
3.2. Effects of Plateau Pika Outbreaks on Soil Organic Carbon Along Altitudinal Gradient
3.3. Key Drivers of SOC and SOCD
3.4. Pika Outbreak Impacts on SOC and SOCD
4. Discussion
4.1. Impacts of Plateau Pika Outbreaks on Vegetation and Soil Physicochemical Properties
4.2. Effects of Plateau Pika Outbreaks on SOC Content and Density
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, S.; Li, Y.; Zhang, Z.; Wang, S.; Liu, X.; Li, C.; Li, G. Distribution characteristics of soil water-soluble organic carbon in typical natural grasslands of the Three-River Source Region. Northwest Agric. J. 2024, 33, 2146–2155. [Google Scholar]
- Huang, P. Effects of Degradation and Artificial Establishment of Alpine Meadows on Soil Microorganisms and Carbon Cycling Functional Genes. Ph.D. Dissertation, Jiangxi Normal University, Nanchang, China, 2024. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, H.; Wang, Q. Spatial heterogeneity of soil carbon in alpine grasslands on the eastern edge of the Qinghai-Tibet Plateau. J. Sichuan Norm. Univ. 2016, 39, 602–607. [Google Scholar] [CrossRef]
- Li, H.; Yuan, H.; Song, H.; Zhang, L.; Duan, L.; Sheng, H. Research progress on soil organic carbon and its components under different utilization methods. Acta Agric. Jiangxi 2020, 32, 57–63. [Google Scholar] [CrossRef]
- Wang, Y. Overview of plant diversity and conservation strategies in the Qinghai-Tibet Plateau ecosystem. Qinghai J. Anim. Husb. Vet. Med. 2015, 45, 47–49. [Google Scholar]
- Du, K.; Kang, Y.; Zhang, D.; Su, J. Effects of different grazing methods on organic carbon and nitrogen pools in Qilian Mountain alpine meadows. Acta Agrestia Sin. 2020, 28, 1412–1420. [Google Scholar]
- Zhao, X. Effects of Enclosure on Soil and Organic Carbon Pool in Alpine Meadows of the Three-River Source Region. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2014. [Google Scholar]
- Chang, X. Spatial Distribution and Influencing Factors of Soil Organic Carbon in Alpine Grasslands of the Three-River Source Region. Ph.D. Dissertation, Graduate University of Chinese Academy of Sciences, Beijing, China, 2012. [Google Scholar]
- Cao, J. Effects of Grassland Management Changes on Soil Organic Carbon and Nitrogen Pools in Qilian Mountain. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2010. [Google Scholar]
- Ma, W.; Liu, Q.; Li, G.; Chang, W. Soil particulate organic carbon content and dynamics at different vegetation degradation stages in Gahai Wetland, Gansu. Acta Pedol. Sin. 2025, 62, 189–200. [Google Scholar]
- Zheng, X.; Chen, M.; Liu, Y.; Liu, Y.; Shi, Y.; Li, F.; Zhang, J. Analysis of current status of soil organic carbon loss. Mod. Chem. Ind. 2020, 40, 7–11. [Google Scholar]
- Li, N.; Guo, T.; Zhang, Q.; Xu, T.; Xu, S. Response of carbon balance in alpine grassland ecosystems to grazing intensity on the Qinghai-Tibet Plateau. J. Domest. Anim. Ecol. 2024, 45, 1–7. [Google Scholar] [CrossRef]
- Li, Y.; Wu, J.; Wu, Z.; Xiu, Y.; Wu, C.; Gong, J.; Zhao, J.; Li, L.; Yu, S.; Sun, F.; et al. Soil organic carbon mineralization and its temperature sensitivity under natural succession restoration of alpine desertified grasslands. Acta Agrestia Sin. 2024, 23, 2686–2694. [Google Scholar] [CrossRef]
- Wang, Y. Effects of Plateau Pika Disturbance on Soil Organic Carbon Fractions and Carbon Cycle Enzyme Activities in Alpine Meadows. Master’s Thesis, Lanzhou University, Lanzhou, China, 2024. [Google Scholar]
- Sun, F.; Long, R.; Guo, Z.; Liu, W.; Gan, Y.; Chen, W. Effects of rodent activities on plant communities and soil environment in alpine meadows. Pratacultural Sci. 2011, 28, 146–151. [Google Scholar]
- Sun, F.; Guo, Z.; Shang, Z.; Long, R. Effects of plateau pika burrow density on soil physicochemical properties in alpine meadows. Acta Pedol. Sin. 2010, 47, 378–383. [Google Scholar] [CrossRef]
- Wang, Y.; Pang, X.; Xiao, Y.; Jia, T.; Wang, Q.; Yu, C.; Guo, Z. Effects of plateau pika disturbance on the relationship between plant diversity and soil nutrients in alpine meadows. Acta Ecol. Sin. 2016, 36, 5485–5496. [Google Scholar] [CrossRef]
- Yu, C.; Jia, T.T.; Pang, X.P.; Guo, Z. Effects of plateau pika disturbance intensity on soil carbon and nitrogen distribution in alpine meadows. Acta Pedol. Sin. 2016, 53, 768–778. [Google Scholar] [CrossRef]
- Song, Z.; Li, X.; Su, X.; Kazhecairang; Ma, G. Spatial distribution patterns and succession laws of disturbance patches in areas with outbreaks of plateau pika and plateau zokor populations. Acta Ecol. Sin. 2023, 43, 2949–2958. [Google Scholar] [CrossRef]
- Duan, C. Study on the Mechanism of Patchy Degradation and Restoration of Alpine Meadows in the Source Region of the Yellow River. Ph.D. Dissertation, Qinghai University, Xining, China, 2023. [Google Scholar]
- Zhang, H. Effects of Plateau Zokor Mounds on Plant Communities and Soil Nutrients in Alpine Meadows. Master’s Thesis, Lanzhou University, Lanzhou, China, 2019. [Google Scholar]
- Zhou, R.; Hua, R.; Hua, M.; Zhang, F.; Ye, G.; Tang, Z.; Hua, L. Spatiotemporal dynamic characteristics of plant community structure in alpine meadows under plateau pika disturbance. Grassl. Turf. 2021, 41, 1–7. [Google Scholar]
- Bao, S. Soil Agrochemical Analysis; China Agriculture Press: Beijing, China, 1999. [Google Scholar]
- Su, X.; Li, X.; Sun, H.; Song, Z.; Li, J.; Zhang, J. Effects of plateau pika and mowing disturbances on plant communities and soil characteristics in alpine meadows. Acta Ecol. Sin. 2024, 44, 10189–10199. [Google Scholar] [CrossRef]
- Shi, H.; Yu, J. Effects of plateau pika burrow density on vegetation and soil moisture in alpine Kobresia meadows. Chin. J. Grassl. 2010, 32, 109–112, 116. [Google Scholar]
- Zhou, J.; Zhao, H.; Yuan, X.; Yang, Y.; Luo, K.; Guo, Z. Characteristics of plant communities in alpine meadows at the edge of the Qinghai-Tibet Plateau. Pratacultural Sci. 2013, 30, 1066–1070. [Google Scholar]
- Zhao, M.; Jiang, W.; Kong, W.; Geng, Q. Diversity and distribution characteristics of plant communities in alpine and high-altitude regions. Guizhou Agric. Sci. 2023, 51, 124–133. [Google Scholar]
- Dong, S. Study on Community Stability and Regulation Mechanisms of Perennial Grass Mixed-Sowing Grasslands in Alpine Regions. Ph.D. Dissertation, Gansu Agricultural University, Lanzhou, China, 2001. [Google Scholar]
- Zhang, F.; Li, Y.; Wang, S.; Zhao, X. Responses of soil organic matter, total nitrogen, and total phosphorus contents to different land use patterns in alpine meadows of the Qinghai-Tibet Plateau. Chin. J. Agrometeorol. 2009, 30, 323–326, 334. [Google Scholar] [CrossRef]
- Wei, C.; Shen, G.; Pei, Z.; Ren, M.; Lu, J.; Wang, Q.; Wang, W. Effects of different plant cultivations on soil physicochemical properties and fine root growth in saline-alkali soils of the Songnen Plain. Bull. Bot. Res. 2015, 35, 759–764. [Google Scholar] [CrossRef]
- Li, H.; He, J.; Wang, L.; Song, D.; Liu, G.; Wang, L.; Li, S. Vegetation species diversity and soil physicochemical properties during the restoration process of alpine grasslands in the eastern Qilian Mountains. Res. Soil Water Conserv. 2025, 32, 110–120. [Google Scholar] [CrossRef]
- Wang, C.; Long, R.; Wang, Q.; Jing Ze Shang, Z.; Ding, L. Distribution of soil organic matter, nitrogen, phosphorus and changes in productivity along different altitude gradients in alpine meadows and their relationships with environmental factors. Acta Prataculturae Sin. 2005, 14, 15–20. [Google Scholar] [CrossRef]
- Ai, L.; Wu, J.; Liu, J.; Qiang, W.; Tian, Z.; Chang, X.; Li, H.; Liu, X. Soil organic carbon and total nitrogen contents and their relationships with altitude, vegetation, and climatic factors: A case study on the northern slope of the middle section of Qilian Mountains. Chin. Hortic. Abstr. 2010, 26, 27–34. [Google Scholar] [CrossRef]
- He, Z.; Zhu, F. Spatial distribution of soil nutrients and microorganisms along different altitude gradients in Xuefeng Mountain. Chin. Agric. Sci. Bull. 2011, 27, 73–78. [Google Scholar]
- Xu, M. Heterogeneity of Soil Nitrogen and Mineralization Characteristics in Alpine Shrub Meadows of Western Sichuan. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2019. [Google Scholar] [CrossRef]
- Zhang, H. Analysis of soil organic matter and total nitrogen content at different altitudes in Lushan Mountain. Shenzhou 2019, 29, 225–227. [Google Scholar]
- Ye, H.; Mi, M. Research progress on interactions between plateau pika and other species. J. Hunan Ecol. Sci. 2023, 10, 113–119. [Google Scholar]
- Liu, W.; Ma, Y.; Wu, Y.; Li, X. Effects of plateau pika disturbance on soil nutrients and microbial biomass carbon and nitrogen in the Qinghai Lake Basin. J. Beijing Norm. Univ. 2018, 54, 254–260. [Google Scholar] [CrossRef]
- Liu, W.; Xu, Q.; Wang, X.; Zhao, J.; Zhou, L. Effects of plateau pika digging activities on soil nitrogen content. Acta Theriol. Sin. 2010, 30, 35–44. [Google Scholar] [CrossRef]
- Ma, J. Effects of Plant Species Diversity on Soil Organic Carbon in Alpine Meadows of the Qinghai-Tibet Plateau. Master’s Thesis, Lanzhou Jiaotong University, Lanzhou, China, 2024. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Y.; Xie, E.; Peng, Y.; Lu, F. Research progress and prospects on spatiotemporal changes of soil organic carbon. J. Agro-Environ. Sci. 2020, 39, 673–679. [Google Scholar] [CrossRef]
- Xu, M. Effects of Altitude Gradient and Land Use Types on Soil Organic Carbon and Microbial Communities in Southeastern Tibet. Ph.D. Dissertation, China Agricultural University, Beijing, China, 2016. [Google Scholar]
- Qiu, S.; Lin, S.; Wang, W. Characteristics and influencing factors of soil organic carbon pools at different altitude gradients in eastern China: A review. Sci. Soil Water Conserv. 2022, 20, 142–150. [Google Scholar] [CrossRef]
- Liu, Y.; Bai, W.; Yin, P.; Feng, Y.; Zhang, J. Effects of exogenous nitrogen addition on soil nutrients and plant community biomass in alpine swamp meadows of the Yangtze River Source Region. Acta Agrestia Sin. 2020, 28, 483–491. [Google Scholar]
- Wang, Y.; Tian, W.; Wang, J.; Qu, W.; Sun, X.; Yang, S. Correlation and estimation models between soil organic matter, total nitrogen levels, and soil carbon emissions. Guizhou Agric. Sci. 2024, 52, 41–48. [Google Scholar] [CrossRef]
- Liu, C.; Duan, Y.; Wang, L.; Wang, Y.; Guo, Z. Effects of plateau pika disturbance on the relationship between plant species diversity and soil ecological stoichiometry in alpine meadows. Acta Prataculturae Sin. 2023, 32, 157–166. [Google Scholar] [CrossRef]
- Xu, X.; Wang, Y.; Wang, X.; Li, J.; Li, J.; Yang, D.; Guo, Z.; Pang, X. Consequences of plateau pika disturbance on plant-soil carbon and nitrogen in alpine meadows. Front. Plant Sci. 2024, 15, 1362125. [Google Scholar] [CrossRef]
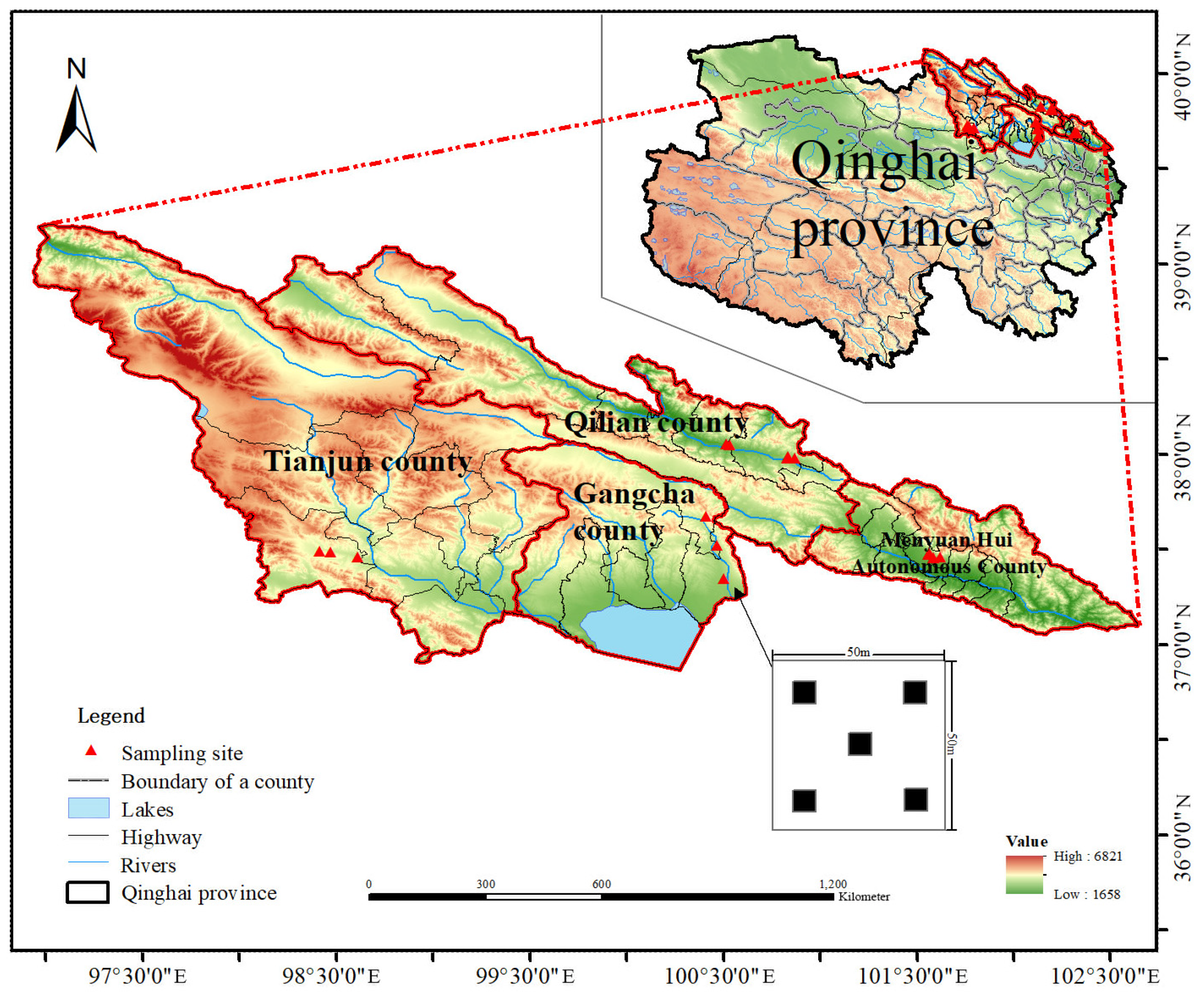
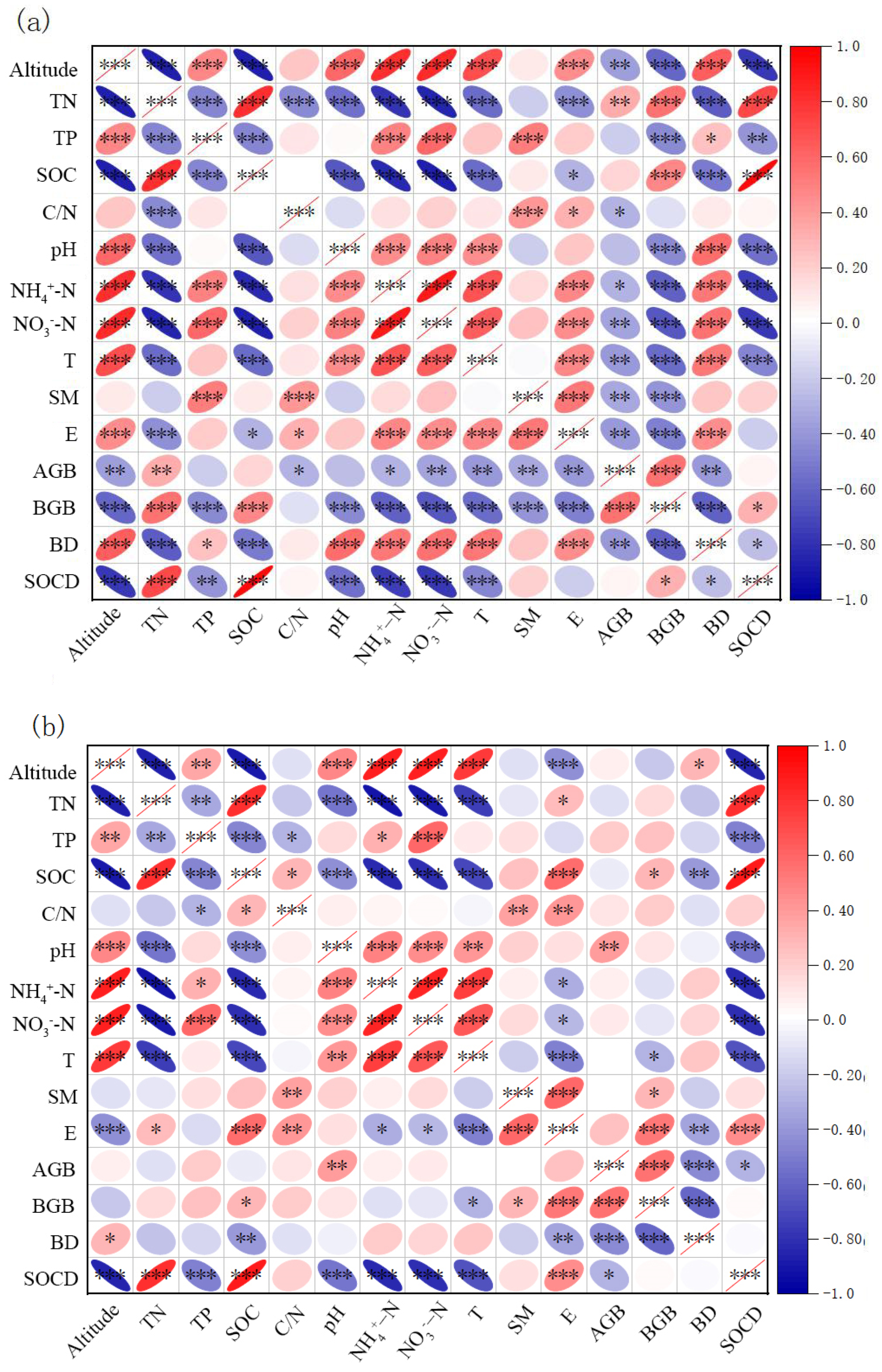
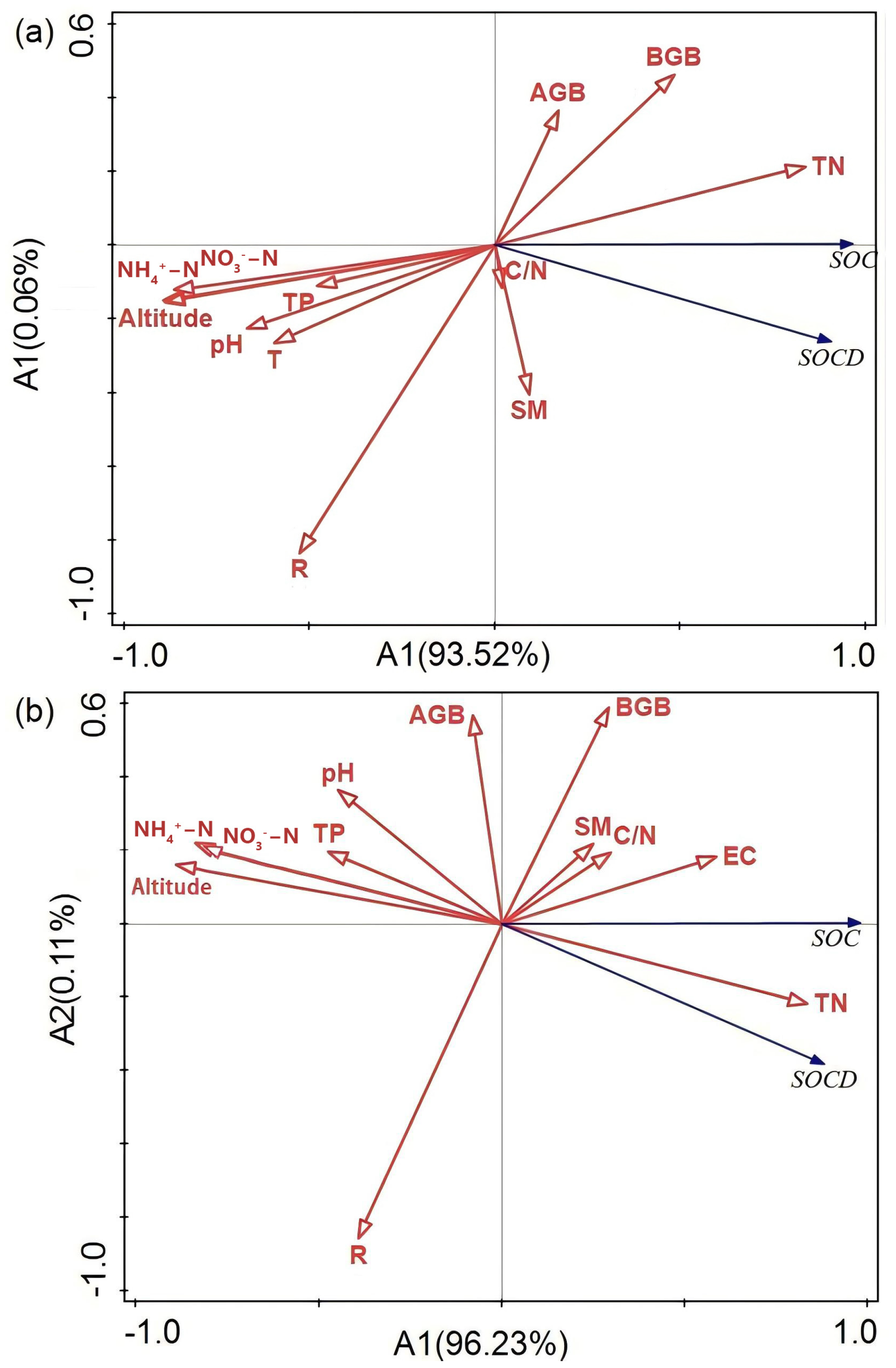
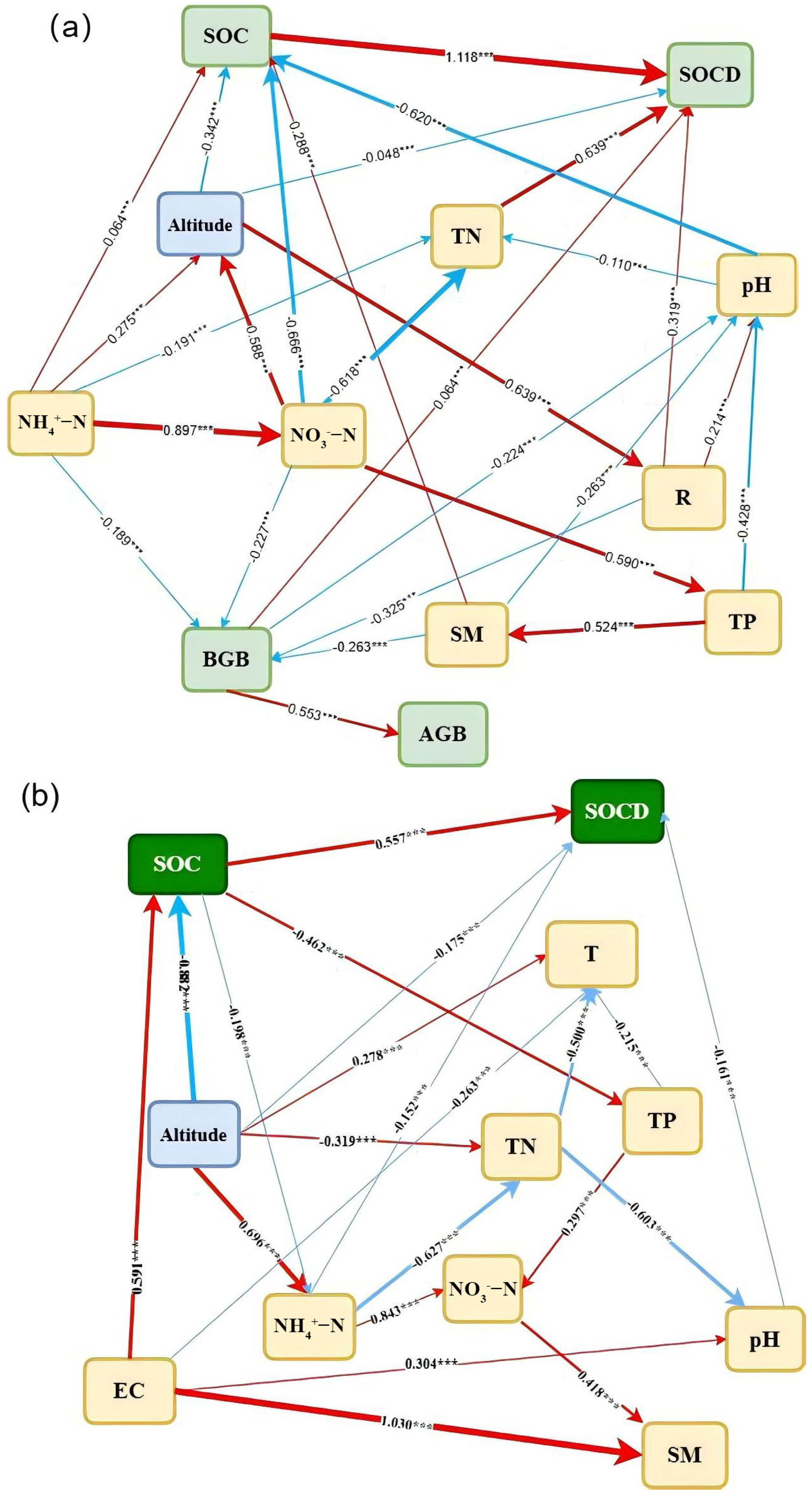

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | Sample Site | Longitudes | Longitude | Elevation (m) | Slope Direction (°) | Average Annual Rainfall (mm) | Average Annual Temperature (°C) | Proportion of Patchily Degraded Areas in an Alpine Meadow |
|---|---|---|---|---|---|---|---|---|
| 1 | Menyuan county | 101.5879 | 37.4572 | 3221 | 259.695 | 517.6 | 1.5 | 64% |
| 2 | Menyuan county | 101.5861 | 37.4573 | 3194 | 223.091 | 517.6 | 1.5 | 67% |
| 3 | Menyuan county | 101.6213 | 37.4626 | 3476 | 247.011 | 517.6 | 1.5 | 63% |
| 4 | Menyuan county | 101.561 | 37.474 | 3152 | 120.619 | 517.6 | 1.5 | 69% |
| 5 | Menyuan county | 101.5649 | 37.4818 | 3182 | 115.427 | 517.6 | 1.5 | 61% |
| 6 | Qilian county | 100.5329 | 38.0532 | 3248 | 186.71 | 420 | 1 | 63% |
| 7 | Qilian county | 100.8363 | 37.9918 | 3312 | 208.413 | 420 | 1 | 68% |
| 8 | Qilian county | 100.5159 | 38.0572 | 3147 | 166.504 | 420 | 1 | 79% |
| 9 | Qilian county | 100.8309 | 37.9814 | 3280 | 18.4349 | 420 | 1 | 77% |
| 10 | Qilian county | 100.871 | 37.9821 | 3361 | 136.042 | 420 | 1 | 65% |
| 11 | Gonchak county | 100.4690 | 37.5246 | 3589 | 248.166 | 360 | −1.5 | 87% |
| 12 | Gonchak county | 100.4089 | 37.6747 | 3711 | 257.671 | 360 | −1.5 | 91% |
| 13 | Gonchak county | 100.5029 | 37.3494 | 3384 | 243.361 | 360 | −1.5 | 89% |
| 14 | Tianjun county | 98.4708 | 37.486 | 3673 | 191.174 | 409.3 | 0.4 | 95% |
| 15 | Tianjun county | 98.4123 | 37.4917 | 3705 | 96.1188 | 409.3 | 0.4 | 90% |
| 16 | Tianjun county | 98.6115 | 37.4603 | 3584 | 160.623 | 409.3 | 0.4 | 87% |
| Elevation/m | Treatments | TN/g·kg−1 | TP/g·kg−1 | pH | C/N | Ammonium Nitrogen/mg·kg−1 | Nitrate Nitrogen/mg·kg−1 | EC | SOC/g·kg−1 | SOCD/g·m3 |
|---|---|---|---|---|---|---|---|---|---|---|
| 3205 | Outbreak areas | 6.34 ± 2.32 aA | 0.30 ± 0.07 bB | 7.42 ± 0.27 bA | 10.20 ± 6.19 aA | 1.30 ± 0.91 bA | 0.96 ± 0.60 bA | 0.09 ± 0.03 bB | 53.97 ± 6.54 bB | 4.22 ± 1.24 bA |
| Non-outbreak area | 6.68 ± 1.11 aA | 0.61 ± 0.19 bA | 7.75 ± 0.34 aA | 11.59 ± 5.59 aA | 0.99 ± 0.42 bA | 1.01 ± 0.59 dA | 0.32 ± 0.27 aA | 73.97 ± 29.88 aA | 5.56 ± 2.38 bA | |
| 3248 | Outbreak areas | 6.59 ± 1.09 aA | 0.47 ± 0.04 bA | 7.35 ± 0.46 bA | 11.80 ± 3.09 aA | 1.25 ± 0.59 bA | 4.00 ± 1.88 bA | 0.16 ± 0.03 aA | 77.15 ± 7.24 aA | 7.40 ± 0.84 aA |
| Non-outbreak area | 6.25 ± 0.70 aA | 0.39 ± 0.04 cA | 7.50 ± 0.37 aA | 11.33 ± 2.10 aA | 0.80 ± 0.35 bA | 2.72 ± 1.04 cA | 0.19 ± 0.05 bA | 71.07 ± 8.70 aA | 7.07 ± 0.57 aA | |
| 3562 | Outbreak areas | 1.20 ± 0.16 bA | 1.33 ± 0.36 aA | 7.67 ± 0.12 abA | 12.95 ± 2.80 aA | 59.83 ± 11.58 aA | 83.45 ± 19.70 aA | 0.18 ± 0.02 aA | 15.52 ± 3.99 cA | 1.67 ± 0.42 cA |
| Non-outbreak area | 1.35 ± 0.17 bA | 1.04 ± 0.28 aA | 7.89 ± 0.06 aA | 11.37 ± 4.38 aA | 64.98 ± 7.80 aA | 66.23 ± 14.34 aA | 0.18 ± 0.02 bA | 14.95 ± 4.66 bA | 1.42 ± 0.47 cA | |
| 3655 | Outbreak areas | 1.19 ± 0.20 bA | 0.46 ± 0.22 bA | 8.12 ± 0.24 aA | 11.79 ± 2.38 aA | 63.22 ± 25.88 aA | 71.35 ± 12.18 aA | 0.20 ± 0.11 aA | 13.66 ± 2.03 cA | 1.61 ± 0.24 cA |
| Non-outbreak area | 1.18 ± 0.33 bA | 0.46 ± 0.15 bcA | 8.12 ± 0.19 aA | 12.70 ± 3.21 aA | 68.54 ± 22.87 aA | 47.52 ± 8.84 bB | 0.13 ± 0.03 cA | 14.12 ± 2.29 bA | 1.41 ± 0.23 cA |
| Elevation/m | Treatments | T/°C | SM/% | Volume Weight/g·cm3 | Above-Ground Biomass/g·m2 | Below-Ground Biomass/g·m2 |
|---|---|---|---|---|---|---|
| 3205 | Outbreak areas | 11.89 ± 1.66 cA | 16.33 ± 3.80 bB | 0.78 ± 0.19 bA | 2.26 ± 0.74 cA | 1.88 ± 6.84 bA |
| Non-outbreak area | 10.94 ± 1.84 bA | 25.87 ± 11.32 bA | 0.78 ± 0.19 aA | 3.11 ± 1.11 cA | 5.68 ± 4.37 cB | |
| 3248 | Outbreak areas | 15.11 ± 4.74 bA | 25.57 ± 3.42 aA | 0.96 ± 0.08 abA | 5.36 ± 1.45 aB | 5.17 ± 1.15 aB |
| Non-outbreak area | 13.53 ± 4.77 bA | 26.14 ± 4.81 bA | 1.01 ± 0.13 aA | 11.99 ± 1.45 aA | 13.21 ± 1.18 bA | |
| 3562 | Outbreak areas | 18.24 ± 1.94 abA | 27.79 ± 2.52 aA | 1.09 ± 0.16 abA | 4.28 ± 1.41 bB | 0.68 ± 0.17 cB |
| Non-outbreak area | 18.83 ± 1.65 aA | 31.39 ± 1.72 aA | 0.96 ± 0.11 aA | 8.04 ± 1.20 bA | 6.48 ± 1.80 cA | |
| 3655 | Outbreak areas | 22.55 ± 2.91 aA | 17.83 ± 5.88 bA | 1.18 ± 0.11 aA | 3.12 ± 1.47 cB | 0.52 ± 0.28 cB |
| Non-outbreak area | 22.23 ± 3.52 aA | 22.62 ± 3.78 bA | 1.00 ± 0.11 aA | 8.15 ± 2.16 bA | 11.53 ± 1.36 aA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, W.; Zhang, J.; Li, X. Soil Organic Carbon Content and Density in Response to Pika Outbreaks Along the Altitudinal Gradient in Alpine Meadows of the Qinghai–Tibet Plateau, West China. Land 2025, 14, 981. https://doi.org/10.3390/land14050981
Yao W, Zhang J, Li X. Soil Organic Carbon Content and Density in Response to Pika Outbreaks Along the Altitudinal Gradient in Alpine Meadows of the Qinghai–Tibet Plateau, West China. Land. 2025; 14(5):981. https://doi.org/10.3390/land14050981
Chicago/Turabian StyleYao, Wenzhi, Jing Zhang, and Xilai Li. 2025. "Soil Organic Carbon Content and Density in Response to Pika Outbreaks Along the Altitudinal Gradient in Alpine Meadows of the Qinghai–Tibet Plateau, West China" Land 14, no. 5: 981. https://doi.org/10.3390/land14050981
APA StyleYao, W., Zhang, J., & Li, X. (2025). Soil Organic Carbon Content and Density in Response to Pika Outbreaks Along the Altitudinal Gradient in Alpine Meadows of the Qinghai–Tibet Plateau, West China. Land, 14(5), 981. https://doi.org/10.3390/land14050981