

Does a Time-Lagged Effect Exist Between Landscape Pattern Changes and Giant Panda Density?
Abstract
1. Introduction
2. Materials and Methods
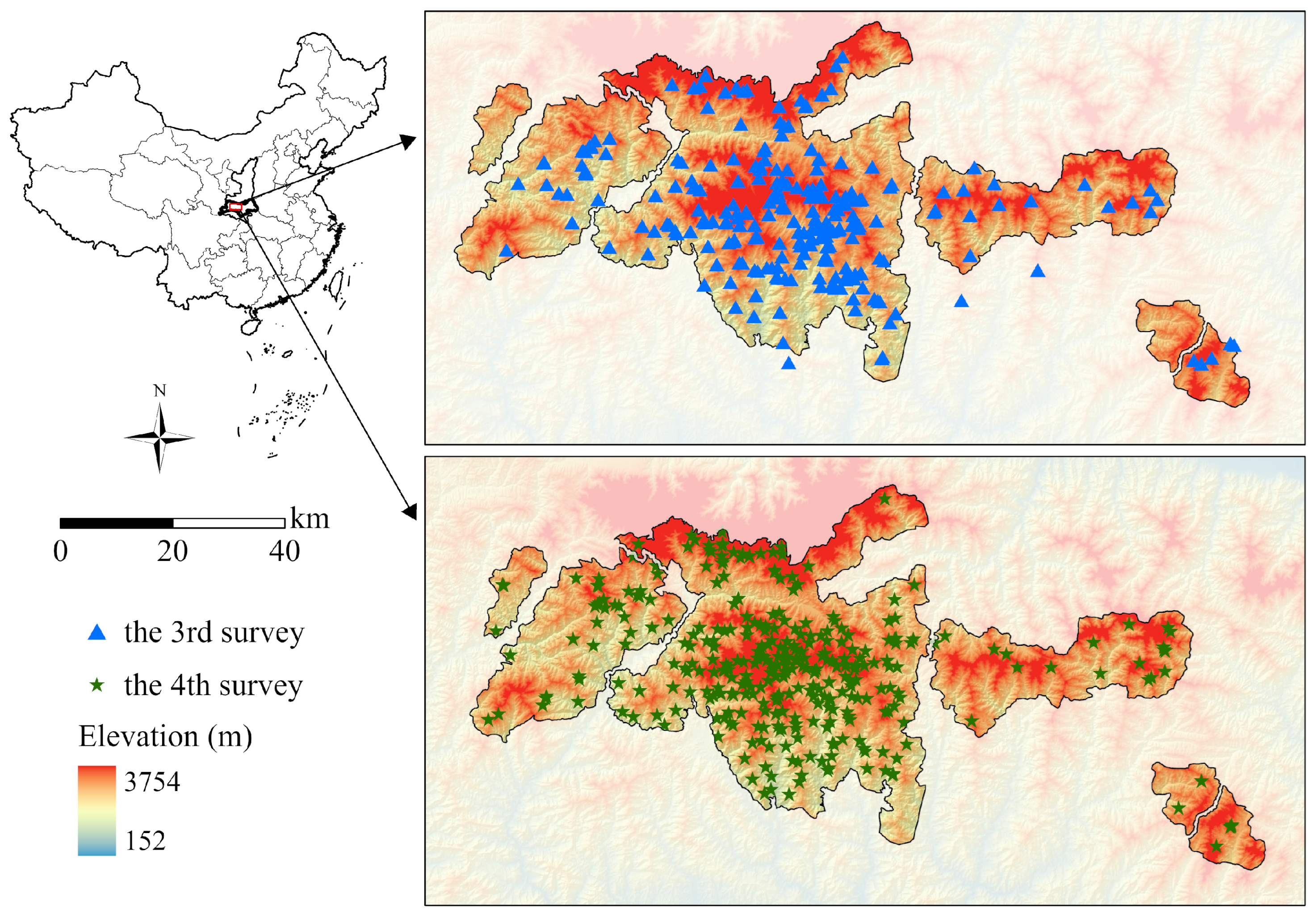
2.1. Study Area
2.2. Random Forest Classification and Regression Models
2.3. Giant Panda Density Data
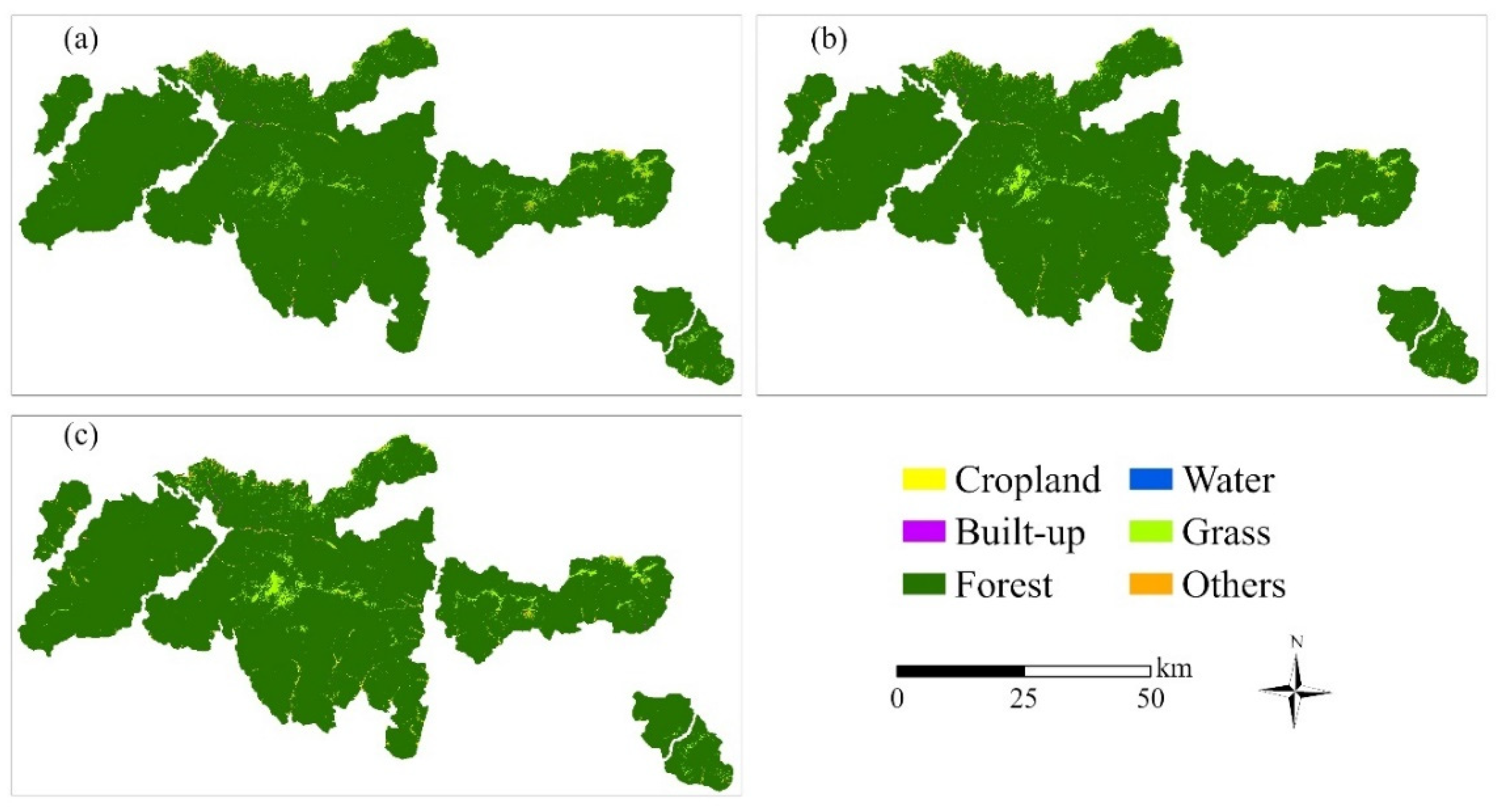
2.4. Land Cover Classification
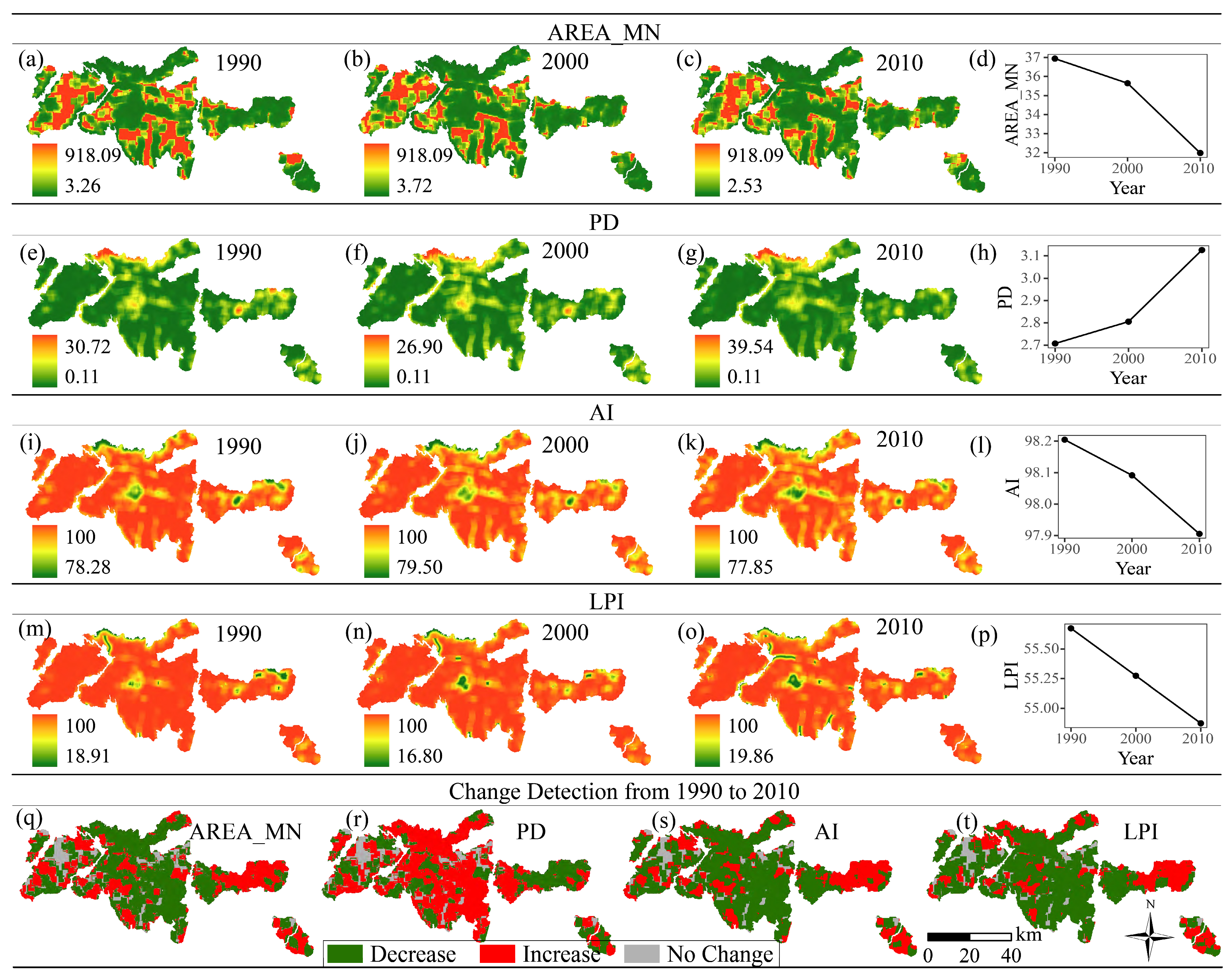
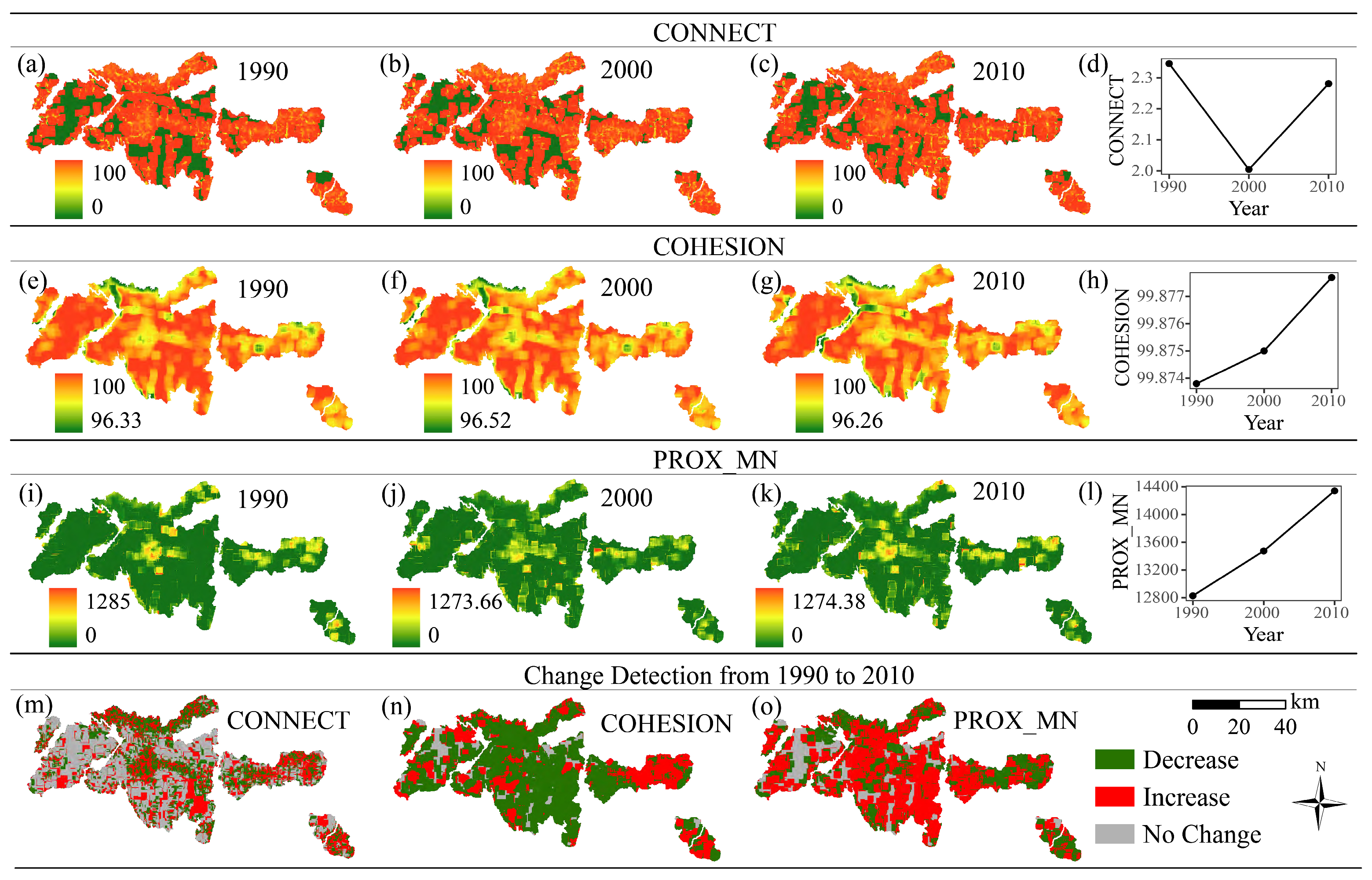
2.5. Landscape Metric Determination
2.6. Time Lag Effects of Landscape Metrics on Giant Panda Density
3. Results
3.1. Land Use and Land Cover Change
3.2. Changes in Landscape Metrics
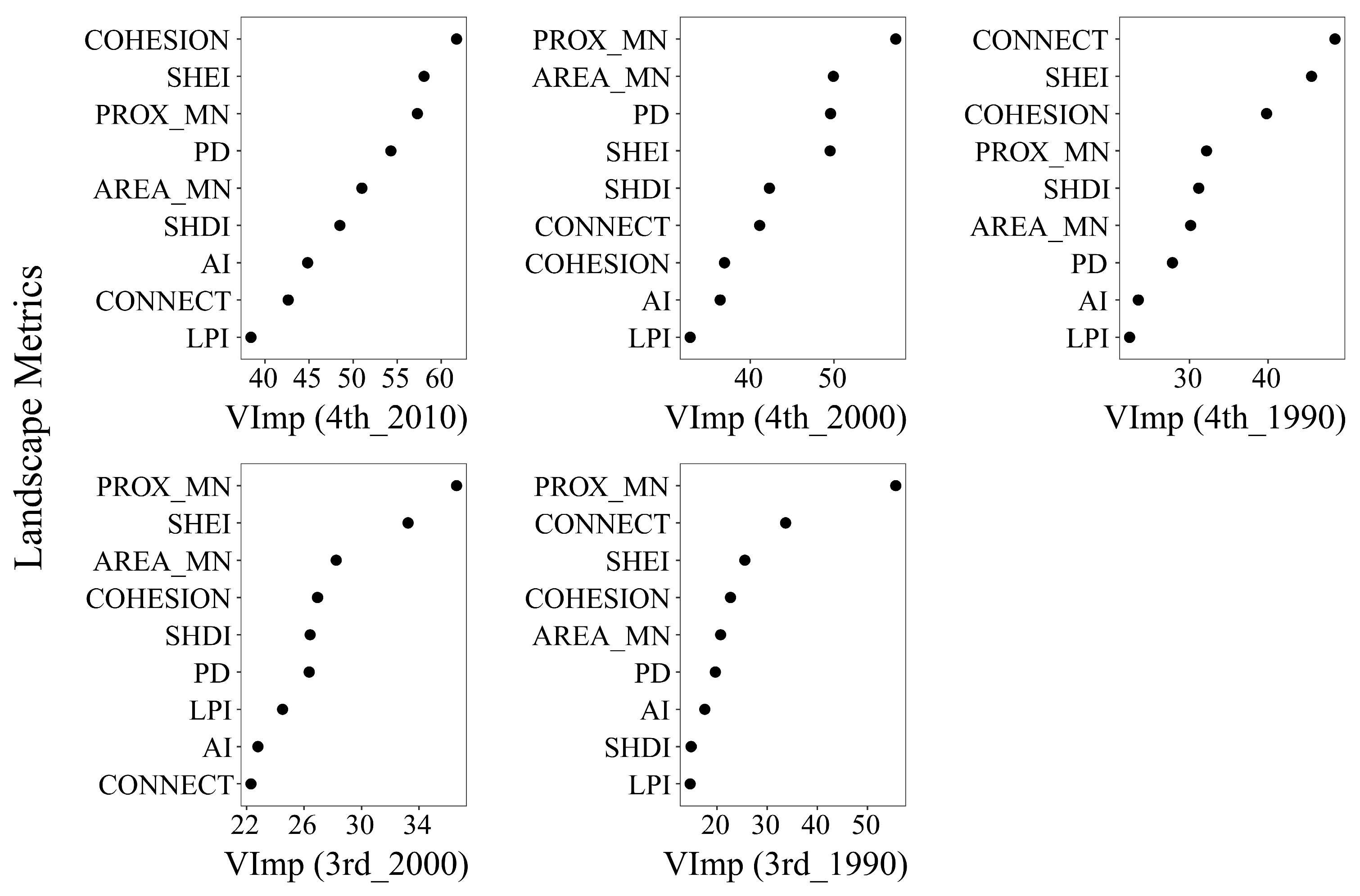
3.3. Does a Time Lag Effect Exist?
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Huang, Q.; Fei, Y.; Yang, H.; Gu, X.; Songer, M. Giant Panda National Park, a step towards streamlining protected areas and cohesive conservation management in China. Global Ecol. Conserv. 2020, 22, e00947. [Google Scholar] [CrossRef]
- Tuanmu, M.N.; Vina, A.; Winkler, J.A.; Li, Y.; Xu, W.; Ouyang, Z.; Liu, J. Climate-change impacts on understorey bamboo species and giant pandas in China’s Qinling Mountains. Nat. Clim. Chang. 2013, 3, 249–253. [Google Scholar] [CrossRef]
- Li, X.; Mao, F.; Du, H.; Zhou, G.; Xing, L.; Liu, T.; Han, N.; Liu, Y.; Zhu, D.E.; Zheng, J.; et al. Spatiotemporal evolution and impacts of climate change on bamboo distribution in China. J. Environ. Manag. 2019, 248, 109265. [Google Scholar] [CrossRef]
- Li, R.Q.; Xu, M.; Wong, M.H.G.; Qiu, S.; Sheng, Q.K.; Li, X.H.; Song, Z.M. Climate change-induced decline in bamboo habitats and species diversity: Implications for giant panda conservation. Divers. Distrib. 2015, 21, 379–391. [Google Scholar] [CrossRef]
- Tang, J.; Swaisgood, R.R.; Owen, M.A.; Zhao, X.; Wei, W.; Pilfold, N.W.; Wei, F.; Yang, X.; Gu, X.; Yang, Z.; et al. Climate change and landscape-use patterns influence recent past distribution of giant pandas. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200358. [Google Scholar] [CrossRef]
- Li, Y.; Rao, T.; Luo, G.; Price, M.L.; Liu, Y.; Ran, J. Giant pandas are losing their edge: Population trend and distribution dynamic drivers of the giant panda. Glob. Chang. Biol. 2023, 29, 4480–4495. [Google Scholar] [CrossRef]
- Wang, Y.; Lan, T.; Deng, S.; Zang, Z.; Zhao, Z.; Xie, Z.; Xu, W.; Shen, G. Forest-cover change rather than climate change determined giant panda’s population persistence. Biol. Conserv. 2022, 265, 109436. [Google Scholar] [CrossRef]
- Alfaya, P.; De Pablo, C.T.L.; Alonso, G. Is landscape fragmentation always detrimental for species conservation? The case of the Iberian lynx in central Spain. Ecol. Complex. 2022, 49, 100985. [Google Scholar] [CrossRef]
- Khan, T.U.; Mannan, A.; Hacker, C.E.; Ahmad, S.; Siddique, M.A.; Khan, B.U.; Din, E.U.; Chen, M.; Zhang, C.; Nizami, M.; et al. Use of GIS and Remote Sensing Data to Understand the Impacts of Land Use/Land Cover Changes (LULCC) on Snow Leopard (Panthera uncia) Habitat in Pakistan. Sustainability 2021, 13, 3590. [Google Scholar] [CrossRef]
- Peng, Y.; Mi, K.; Wang, H.; Liu, Z.; Lin, Y.; Sang, W.; Cui, Q. Most suitable landscape patterns to preserve indigenous plant diversity affected by increasing urbanization: A case study of Shunyi District of Beijing, China. Urban For. Urban Green. 2019, 38, 33–41. [Google Scholar] [CrossRef]
- Chan, A.N.; Wittemyer, G.; McEvoy, J.; Williams, A.C.; Cox, N.; Soe, P.; Grindley, M.; Shwe, N.M.; Chit, A.M.; Oo, Z.M.; et al. Landscape characteristics influence ranging behavior of Asian elephants at the human-wildlands interface in Myanmar. Mov. Ecol. 2022, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Schindler, S.; von Wehrden, H.; Poirazidis, K.; Wrbka, T.; Kati, V. Multiscale performance of landscape metrics as indicators of species richness of plants, insects and vertebrates. Ecol. Indic. 2013, 31, 41–48. [Google Scholar] [CrossRef]
- Rios, E.; Benchimol, M.; Dodonov, P.; De Vleeschouwer, K.; Cazetta, E. Testing the habitat amount hypothesis and fragmentation effects for medium- and large-sized mammals in a biodiversity hotspot. Landsc. Ecol. 2021, 36, 1311–1323. [Google Scholar] [CrossRef]
- Lombardi, J.V.; Tewes, M.E.; Perotto-Baldivieso, H.L.; Mata, J.M.; Campbell, T.A. Spatial structure of woody cover affects habitat use patterns of ocelots in Texas. Mammal Res. 2020, 65, 555–563. [Google Scholar] [CrossRef]
- Chatterjee, P.; Mukherjee, T.; Dutta, R.; Sharief, A.; Kumar, V.; Joshi, B.D.; Chandra, K.; Thakur, M.; Sharma, L.K. Future simulated landscape predicts habitat loss for the Golden Langur (Trachypithecus geei): A range level analysis for an endangered primate. Sci. Total Environ. 2022, 826, 154081. [Google Scholar] [CrossRef]
- Gao, B.; Gong, P.; Zhang, W.; Yang, J.; Si, Y. Multiscale effects of habitat and surrounding matrices on waterbird diversity in the Yangtze River Floodplain. Landsc. Ecol. 2021, 36, 179–190. [Google Scholar] [CrossRef]
- Fernández, V.P.; Rodríguez-Gómez, G.B.; Molina-Marín, D.A.; Castaño-Villa, G.J.; Fontúrbel, F.E. Effects of landscape configuration on the occurrence and abundance of an arboreal marsupial from the Valdivian rainforest. Rev. Chil. Hist. Nat. 2022, 95, 3. [Google Scholar] [CrossRef]
- Capellesso, E.S.; da Rosa, C.M.; Magnago, L.F.S.; Marques, R.; Marques, M.C.M. Habitat amount is a driver for biodiversity, but not for the carbon stock in post-logging natural regenerating areas in Tropical Atlantic Forest. Biol. Conserv. 2022, 273, 109673. [Google Scholar] [CrossRef]
- Wang, T.; Ye, X.; Skidmore, A.K.; Toxopeus, A.G. Characterizing the spatial distribution of giant pandas (Ailuropoda melanoleuca) in fragmented forest landscapes. J. Biogeogr. 2010, 37, 865–878. [Google Scholar] [CrossRef]
- Liu, X.; Wu, P.; Shao, X.; Songer, M.; Cai, Q.; Zhu, Y.; He, X. Spatiotemporally monitoring forest landscape for giant panda habitat through a high learning-sensitive neural network in Guanyinshan Nature Reserve in the Qinling Mountains, China. Environ. Earth Sci. 2017, 76, 589. [Google Scholar] [CrossRef]
- Qin, Q.; Huang, Y.; Liu, J.; Chen, D.; Zhang, L.; Qiu, J.; Tan, H.; Wen, Y. The Landscape Patterns of the Giant Panda Protection Area in Sichuan Province and Their Impact on Giant Pandas. Sustainability 2019, 11, 5993. [Google Scholar] [CrossRef]
- Majumdar, P.; Mondal, B.; Debnath, S.; Sarkar, S.; Ghosh, U. Effect of fear and delay on a prey-predator model with predator harvesting. Comp. Appl. Math. 2022, 41, 357. [Google Scholar] [CrossRef]
- Cheng, M.; Wang, Z.; Wang, S.; Liu, X.; Jiao, W.; Zhang, Y. Determining the impacts of climate change and human activities on vegetation change on the Chinese Loess Plateau considering human-induced vegetation type change and time-lag effects of climate on vegetation growth. Int. J. Digital Earth 2024, 17, 2336075. [Google Scholar] [CrossRef]
- Ma, M.; Wang, Q.; Liu, R.; Zhao, Y.; Zhang, D. Effects of climate change and human activities on vegetation coverage change in northern China considering extreme climate and time-lag and -accumulation effects. Sci. Total Environ. 2023, 860, 160527. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, X.; Jia, R.; Peng, X.; Zhang, X.; Yang, M.; Li, Z.; Guo, J.; Chen, Y.; Yin, W.; et al. The Spatiotemporal Pattern and Its Determinants of Hemorrhagic Fever With Renal Syndrome in Northeastern China: Spatiotemporal Analysis. Jmir Public Health Surveill. 2023, 9, e42673. [Google Scholar] [CrossRef]
- Coutts, S.R.; Helmstedt, K.J.; Bennett, J.R. Invasion lags: The stories we tell ourselves and our inability to infer process from pattern. Divers. Distrib. 2018, 24, 244–251. [Google Scholar] [CrossRef]
- Wei, W.; Han, H.; Zhou, H.; Hong, M.; Cao, S.; Zhang, Z. Microhabitat use and separation between giant panda (Ailuropoda melanoleuca), takin (Budorcas taxicolor), and goral (Naemorhedus griseus) in Tangjiahe Nature Reserve, China. Folia Zool. 2018, 67, 198–206. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Q.; Cui, B.; Chen, G.; Xie, T.; Yang, W. Ecological time lags in biodiversity response to habitat changes. J. Environ. Manag. 2023, 346, 118965. [Google Scholar] [CrossRef]
- Zani, D.; Lischke, H.; Lehsten, V. The role of dispersal limitation in the forest biome shifts of Europe in the last 18,000 years. J. Biogeogr. 2024, 51, 1438–1457. [Google Scholar] [CrossRef]
- Chen, Y.X.; Zeng, C.J.; Fang, S.G. Y-Chromosome microdissection and Y-linked genes identification of the giant panda (Ailuropoda melanoleuca). J. Anim. Plant Sci. 2022, 32, 1478–1485. [Google Scholar] [CrossRef]
- Yang, S.; Lan, T.; Wei, R.; Zhang, L.; Lin, L.; Du, H.; Huang, Y.; Zhang, G.; Huang, S.; Shi, M.; et al. Single-nucleus transcriptome inventory of giant panda reveals cellular basis for fitness optimization under low metabolism. BMC Biol. 2023, 21, 222. [Google Scholar] [CrossRef] [PubMed]
- Nomoto, H.A.; Alexander, J.M. Drivers of local extinction risk in alpine plants under warming climate. Ecol. Lett. 2021, 24, 1157–1166. [Google Scholar] [CrossRef]
- Li, J.; Liu, F.; Xue, Y.; Zhang, Y.; Li, D. Assessing vulnerability of giant pandas to climate change in the Qinling Mountains of China. Ecol. Evol. 2017, 7, 4003–4015. [Google Scholar] [CrossRef]
- Sánchez, A.C.; Salazar, A.; Oviedo, C.; Bandopadhyay, S.; Mondaca, P.; Valentini, R.; Briceño, N.B.R.; Guzmán, C.T.; Oliva, M.; Guzman, B.K.; et al. Integrated cloud computing and cost effective modelling to delineate the ecological corridors for Spectacled bears (Tremarctos ornatus) in the rural territories of the Peruvian Amazon. Global Ecol. Conserv. 2022, 36, e02126. [Google Scholar] [CrossRef]
- Ma, L.; Li, M.; Ma, X.; Cheng, L.; Du, P.; Liu, Y. A review of supervised object-based land-cover image classification. ISPRS J. Photogramm. Remote Sens. 2017, 130, 277–293. [Google Scholar] [CrossRef]
- Zhao, Q.; Yu, S.; Zhao, F.; Tian, L.; Zhao, Z. Comparison of machine learning algorithms for forest parameter estimations and application for forest quality assessments. For. Ecol. Manag. 2019, 434, 224–234. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Sun, C.; Jin, X.; Yuan, W.; Wang, W.; Cao, Y.; Lu, X. The 3rd Comprehensive Survey Report on Giant Panda in Shaani Province; Xi’an Map Press: Xi’an, China, 2007. [Google Scholar]
- Zhou, L.G.; Zhang, X.M.; Jiu, Q.; Meng, X.M. Report of the Fourth Survey on Giant Panda in Qinling Mountains, Shaanxi Province; Shaanxi Science and Technology Press: Xi’an, China, 2017. [Google Scholar]
- McGarigal, K.; Cushman, S.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical Maps. Computer Software Program Produced by the Authors. 2023. Available online: https://www.fragstats.org (accessed on 25 November 2024).
- Mahmoudzadeh, H.; Masoudi, H.; Jafari, F.; Khorshiddoost, A.M.; Abedini, A.; Mosavi, A. Ecological networks and corridors development in urban areas: An example of Tabriz, Iran. Front. Environ. Sci. 2022, 10, 969266. [Google Scholar] [CrossRef]
- Yang, Y. Evolution of habitat quality and association with land-use changes in mountainous areas: A case study of the Taihang Mountains in Hebei Province, China. Ecol. Indic. 2021, 129, 107967. [Google Scholar] [CrossRef]
- Renó, V.; Novo, E. Forest depletion gradient along the Amazon floodplain. Ecol. Indic. 2019, 98, 409–419. [Google Scholar] [CrossRef]
- Almeida-Gomes, M.; Lira, P.K.; Severo-Neto, F.; de Souza, F.L.; Valente-Neto, F. Evidence of taxonomic but not functional diversity extinction debt in bird assemblages in an urban area in the Cerrado hotspot. Landsc. Urban Plann. 2025, 253, 105219. [Google Scholar] [CrossRef]
- Balasubramaniam, T.; Mohotti, W.A.; Sabir, K.; Nayak, R. Feature engineering on climate data with machine learning to understand time-lagging effects in pasture yield prediction. Ecol. Inf. 2025, 86, 103011. [Google Scholar] [CrossRef]
- Deng, X.; Li, Z. A review on historical trajectories and spatially explicit scenarios of land-use and land-cover changes in China. J. Land Use Sci. 2016, 11, 709–724. [Google Scholar] [CrossRef]
- Chen, D.; Jin, X.; Zhang, X.; Zhu, Q.; Zhang, Z.; Hu, S.; Chen, Y.; Zhao, Q. Giant panda habitat restoration requires more than just planting bamboo and trees. Restor. Ecol. 2023, 31, e13817. [Google Scholar] [CrossRef]
- Li, C.; Bao, Z.-Q.; Luo, X.-R.; Wu, W.; Yu, J.-J.; Hou, R.; Owens, J.R.; Xu, Q.; Gu, X.-D.; Yang, H.; et al. Does high vegetation coverage equal high giant panda density? Zool. Res. 2022, 43, 608–611. [Google Scholar] [CrossRef]
- Bai, W.; Huang, Q.; Zhang, J.; Stabach, J.; Huang, J.; Yang, H.; Songer, M.; Connor, T.; Liu, J.; Zhou, S.; et al. Microhabitat selection by giant pandas. Biol. Conserv. 2020, 247, 108615. [Google Scholar] [CrossRef]
- Lindborg, R.; Eriksson, O. Historical landscape connectivity affects present plant species diversity. Ecology 2004, 85, 1840–1845. [Google Scholar] [CrossRef]
- Bu, H.; McShea, W.J.; Wang, D.; Wang, F.; Chen, Y.; Gu, X.; Yu, L.; Jiang, S.; Zhang, F.; Li, S. Not all forests are alike: The role of commercial forest in the conservation of landscape connectivity for the giant panda. Landsc. Ecol. 2021, 36, 2549–2564. [Google Scholar] [CrossRef]
- Osterhout, M.J.; Stewart, K.M.; Wakeling, B.F.; Schroeder, C.A.; Blum, M.E.; Brockman, J.C.; Shoemaker, K.T. Effects of large-scale gold mining on habitat use and selection by American pronghorn. Sci. Total Environ. 2024, 921, 170750. [Google Scholar] [CrossRef]
- Vanthomme, H.P.A.; Nzamba, B.S.; Alonso, A.; Todd, A.F. Empirical selection between least-cost and current-flow designs for establishing wildlife corridors in Gabon. Conserv. Biol. 2019, 33, 329–338. [Google Scholar] [CrossRef]
- Wang, F.; McShea, W.J.; Wang, D.; Li, S.; Zhao, Q.; Wang, H.; Lu, Z. Evaluating Landscape Options for Corridor Restoration between Giant Panda Reserves. PLoS ONE 2014, 9, e105086. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Lu, D.; Zhao, Y.; Zhou, M.; Chen, G. Spatial patterns of vegetation coverage change in giant panda habitat based on MODIS time-series observations and local indicators of spatial association. Ecol. Indic. 2021, 124, 107418. [Google Scholar] [CrossRef]
- Jia, W.; Yan, S.; He, Q.; Li, P.; Fu, M.; Zhou, J. Giant Panda Microhabitat Study in the Daxiangling Niba Mountain Corridor. Biology 2023, 12, 165. [Google Scholar] [CrossRef] [PubMed]
- Hämäläinen, A.; Fahrig, L. Time-lag effects of habitat loss, but not fragmentation, on deadwood-dwelling lichens. Landsc. Ecol. 2024, 39, 111. [Google Scholar] [CrossRef]
- Kuussaari, M.; Bommarco, R.; Heikkinen, R.K.; Helm, A.; Krauss, J.; Lindborg, R.; Öckinger, E.; Pärtel, M.; Pino, J.; Rodà, F.; et al. Extinction debt: A challenge for biodiversity conservation. Trends Ecol. Evol. 2009, 24, 564–571. [Google Scholar] [CrossRef]
- Chen, Y.; Shen, T.J. A general framework for predicting delayed responses of ecological communities to habitat loss. Sci. Rep. 2017, 7, 998. [Google Scholar] [CrossRef]


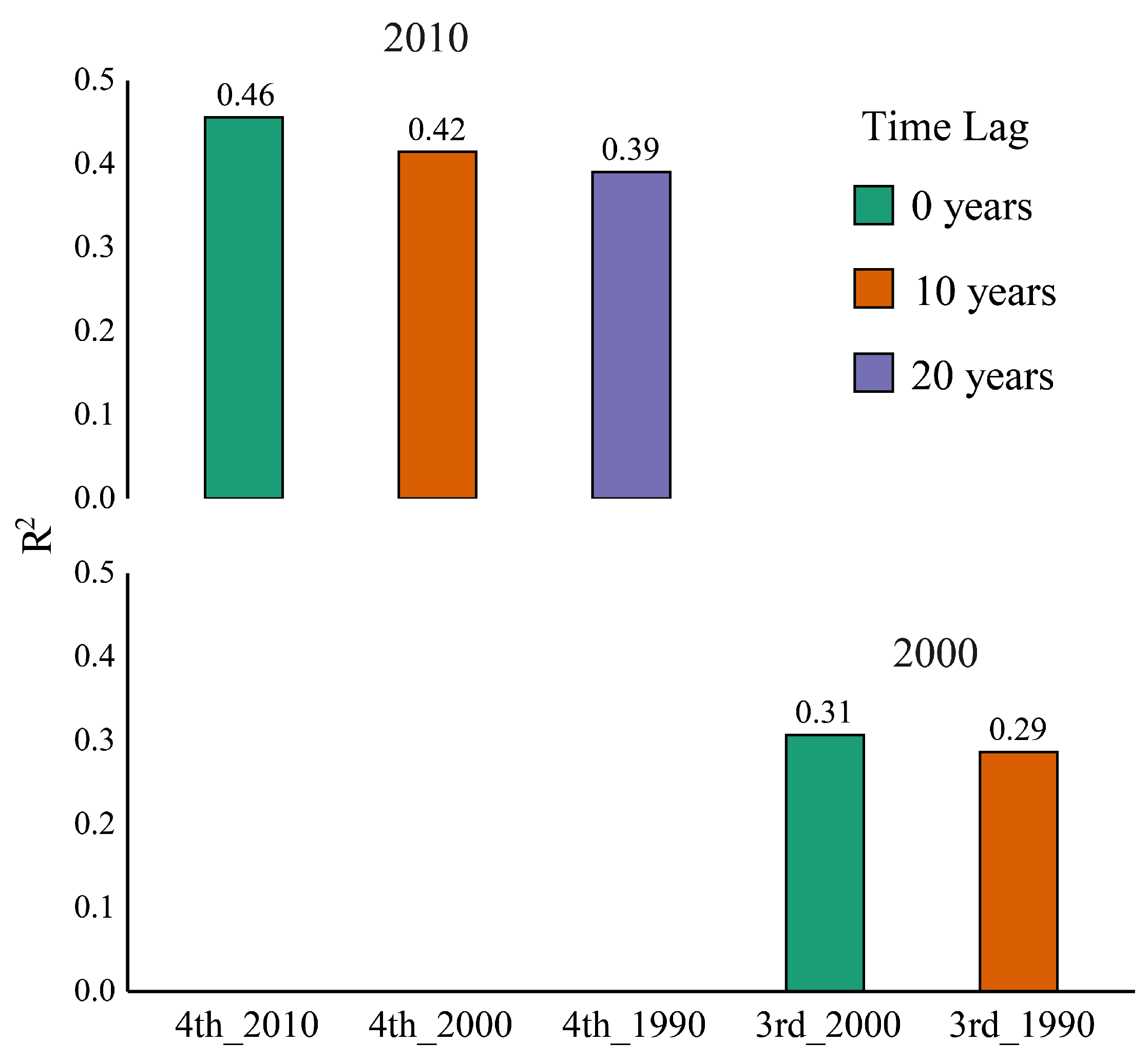

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Cover | Description |
|---|---|
| Built-up | Urban and rural residential areas and other industrial, mining, and transportation lands |
| Cropland | Land used for growing crops. |
| Forest | Forestry land for growing trees, shrubs, bamboo, and coastal mangroves. |
| Grass | Grasslands that mainly grow herbaceous plants with a coverage of more than 5%, including shrub grasslands dominated by grazing and open forest grasslands with a canopy density of less than 10%. |
| Others | Land that is currently unused, including land that is difficult to use. |
| Water | Land for natural terrestrial waters and water conservancy facilities. |
| LULCC | 1990 | 2000 | 2010 | |||
|---|---|---|---|---|---|---|
| PA (%) | UA (%) | PA (%) | UA (%) | PA (%) | UA (%) | |
| built-up | 85.90 | 92.45 | 86.89 | 91.94 | 85.98 | 95.38 |
| cropland | 95.66 | 91.11 | 96.82 | 91.75 | 97.82 | 88.95 |
| forest | 99.73 | 99.38 | 99.83 | 99.52 | 99.84 | 99.48 |
| grass | 93.58 | 96.53 | 95.83 | 97.01 | 94.91 | 96.98 |
| others | 96.49 | 98.16 | 94.64 | 98.07 | 94.96 | 98.54 |
| water | 94.74 | 97.41 | 93.53 | 97.43 | 91.89 | 98.03 |
| Overall (%) | 97.38 | 97.55 | 97.44 | |||
| Kappa | 0.9544 | 0.9578 | 0.9558 | |||
| LULCC | 1990 | 2000 | 2010 | 1990–2010 | ||||
|---|---|---|---|---|---|---|---|---|
| Area (km2) | Area (%) | Area (km2) | Area (%) | Area (km2) | Area (%) | Area (km2) | Area (%) | |
| built-up | 2.31 | 0.06 | 2.18 | 0.06 | 2.40 | 0.07 | 0.09 | 3.90 |
| cropland | 8.87 | 0.25 | 15.80 | 0.44 | 22.62 | 0.63 | 13.75 | 155.02 |
| forest | 3500.90 | 97.36 | 3492.01 | 97.11 | 3478.53 | 96.74 | −22.37 | −0.64 |
| grass | 76.89 | 2.14 | 78.52 | 2.18 | 83.39 | 2.32 | 6.50 | 8.45 |
| others | 6.55 | 0.18 | 6.96 | 0.19 | 8.10 | 0.23 | 1.55 | 23.66 |
| water | 0.34 | 0.01 | 0.39 | 0.01 | 0.83 | 0.02 | 0.49 | 144.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Q.; Zhu, Q.; Huang, J.; Cui, Y.; Liu, Y.; Chen, D.; Jin, X. Does a Time-Lagged Effect Exist Between Landscape Pattern Changes and Giant Panda Density? Land 2025, 14, 1075. https://doi.org/10.3390/land14051075
Zhao Q, Zhu Q, Huang J, Cui Y, Liu Y, Chen D, Jin X. Does a Time-Lagged Effect Exist Between Landscape Pattern Changes and Giant Panda Density? Land. 2025; 14(5):1075. https://doi.org/10.3390/land14051075
Chicago/Turabian StyleZhao, Qingxia, Qifeng Zhu, Jiqin Huang, Yueduo Cui, Yutai Liu, Dong Chen, and Xuelin Jin. 2025. "Does a Time-Lagged Effect Exist Between Landscape Pattern Changes and Giant Panda Density?" Land 14, no. 5: 1075. https://doi.org/10.3390/land14051075
APA StyleZhao, Q., Zhu, Q., Huang, J., Cui, Y., Liu, Y., Chen, D., & Jin, X. (2025). Does a Time-Lagged Effect Exist Between Landscape Pattern Changes and Giant Panda Density? Land, 14(5), 1075. https://doi.org/10.3390/land14051075