
Impact of Future Urban Expansion on Endemic Species in China at the Species Level


Abstract
1. Introduction
2. Materials and Methods
2.1. Land Use/Cover Data
2.2. Species Data
2.3. Extent of Suitable Habitat Analysis
2.4. Fragmentation of ESH
2.5. Countryside Species–Area Relationship Model
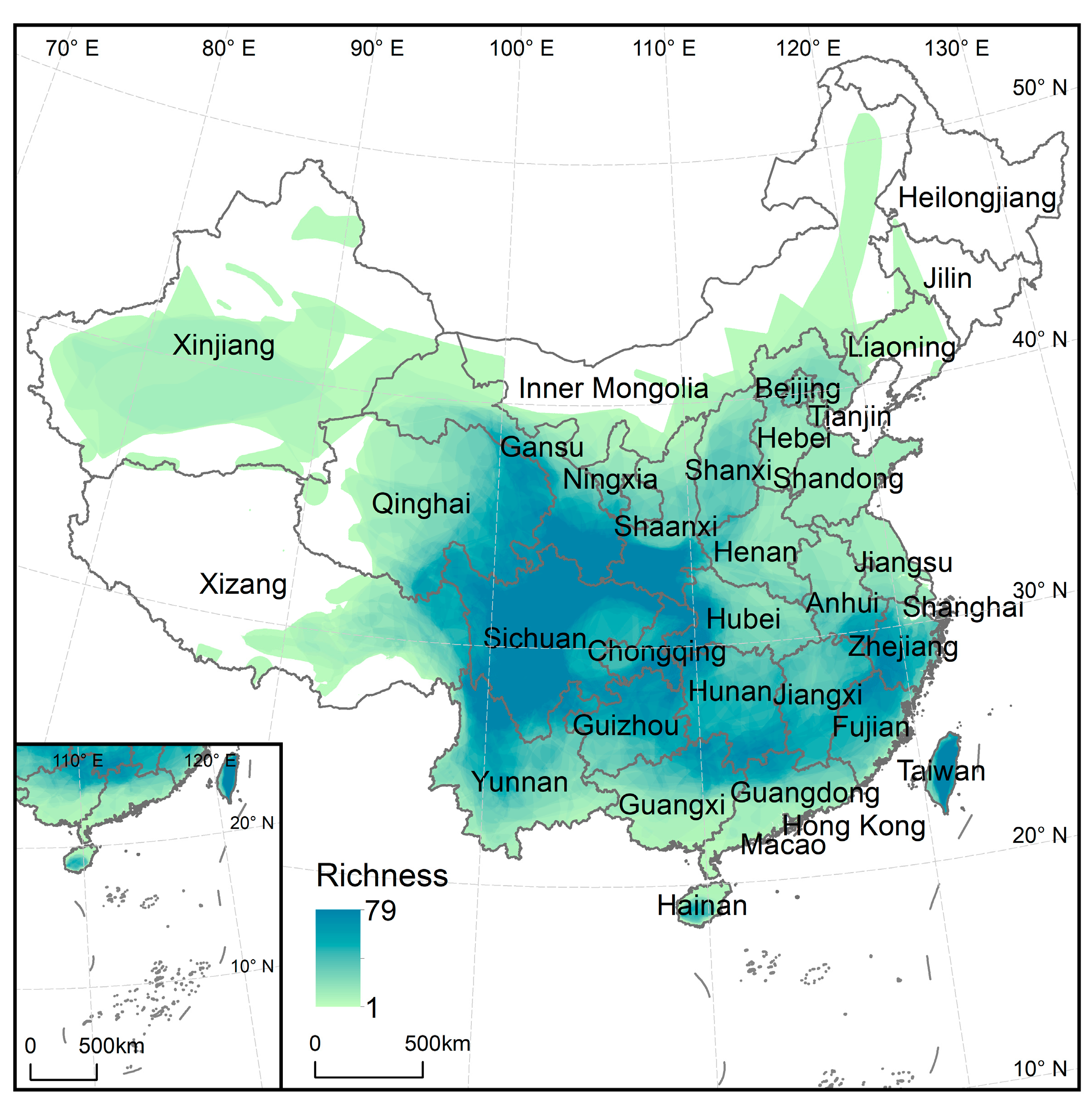
3. Results
3.1. Urban Expansion from 2015 to 2050
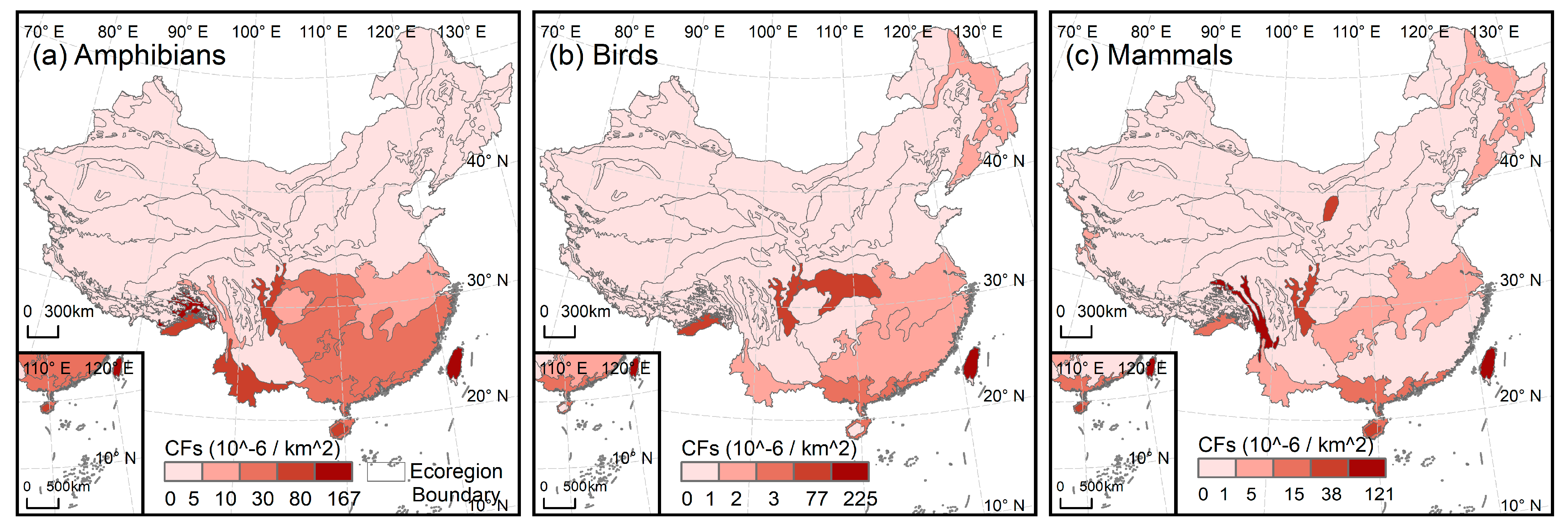
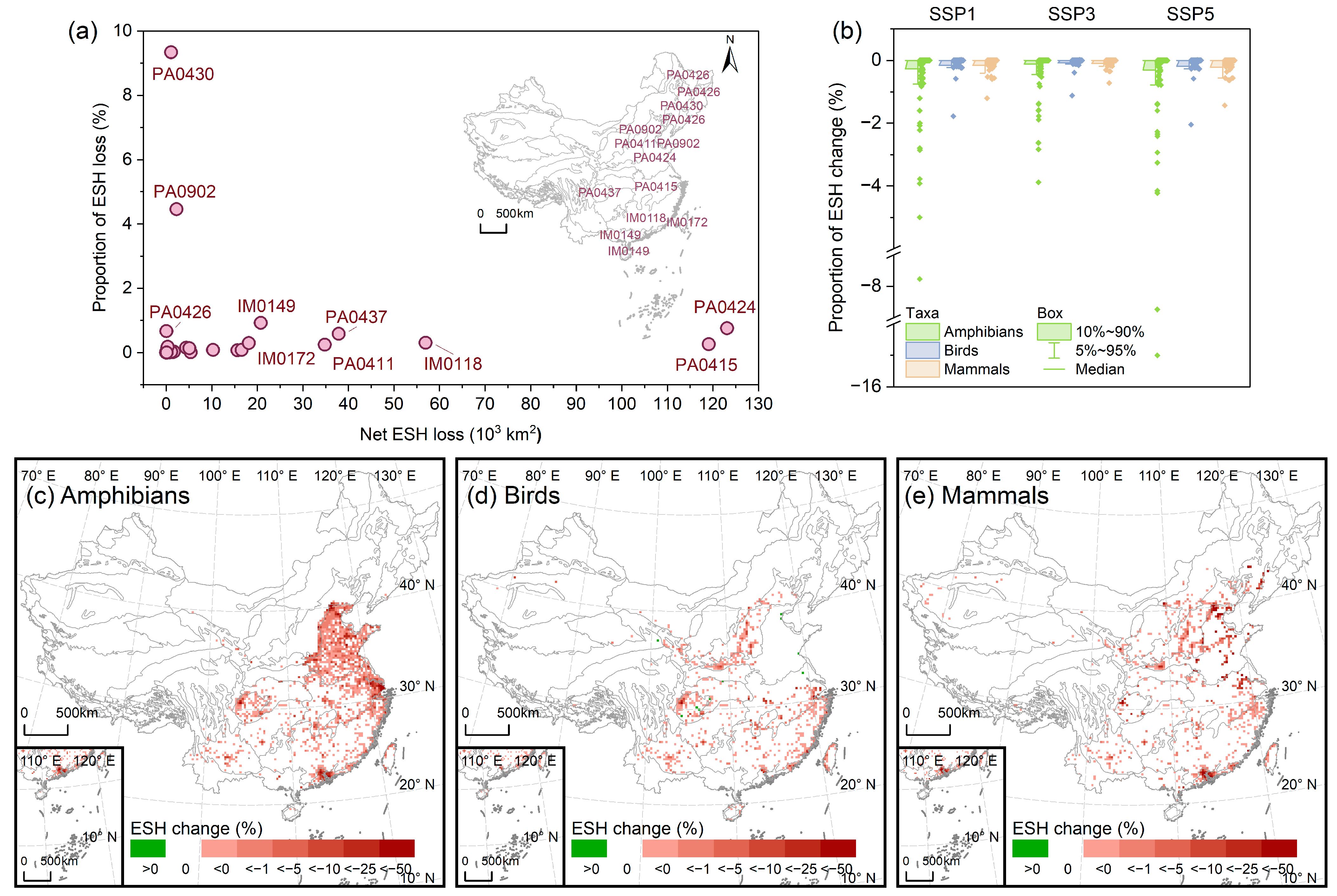
3.2. Impact of Urban Expansion on ESH
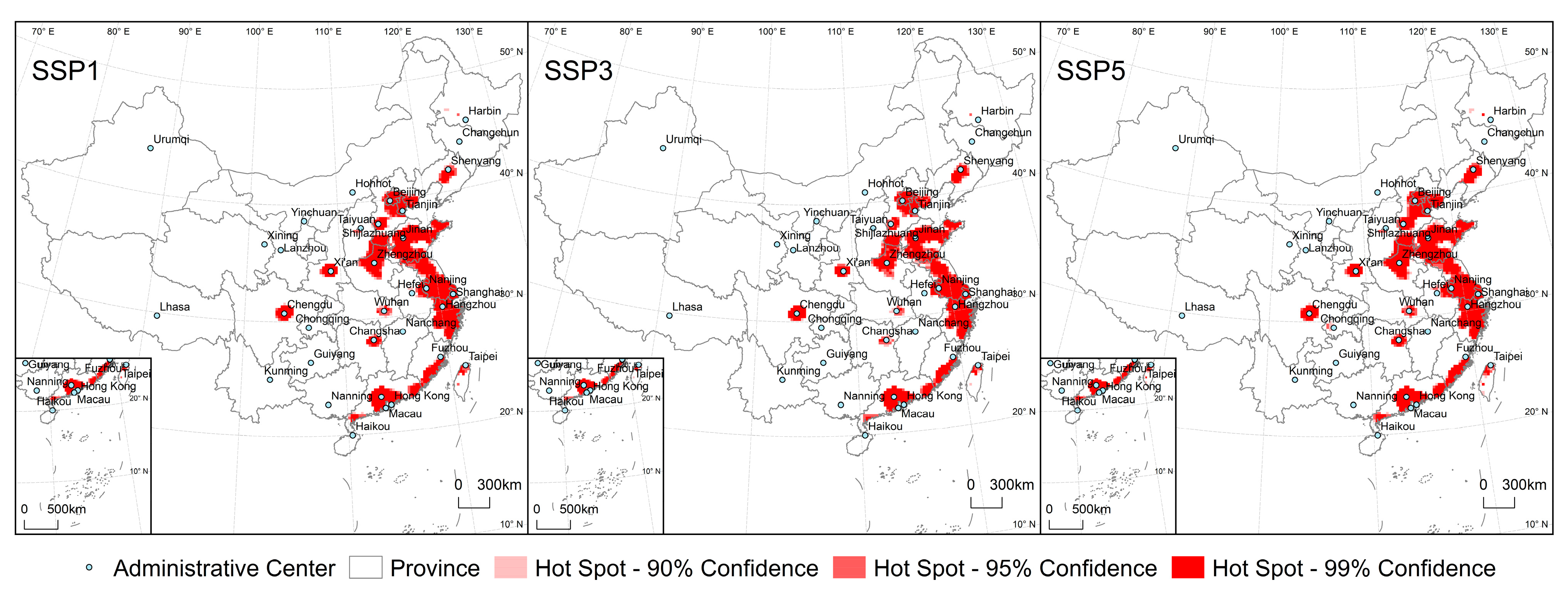
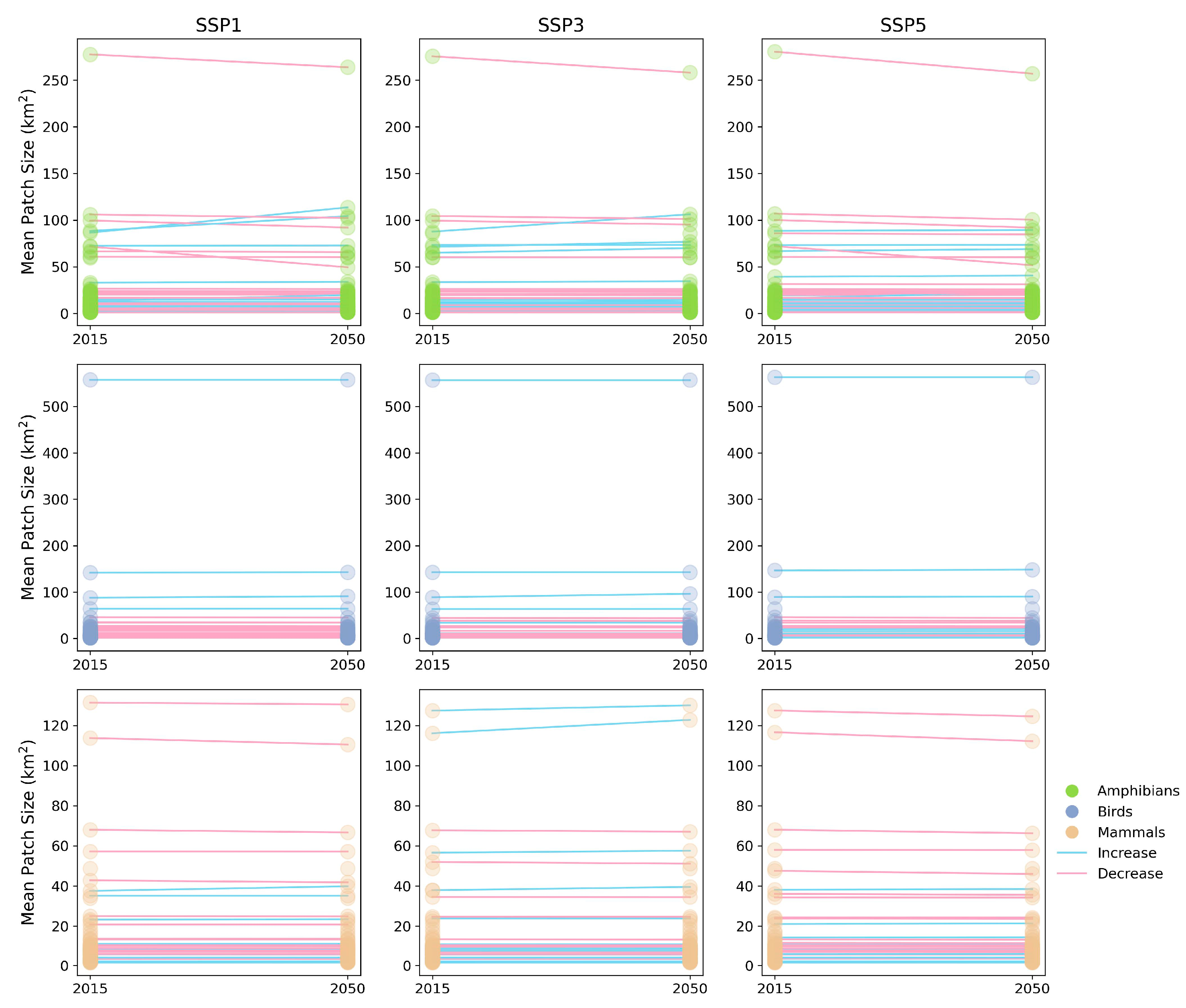
3.3. Impact of Urban Expansion on ESH Fragmentation
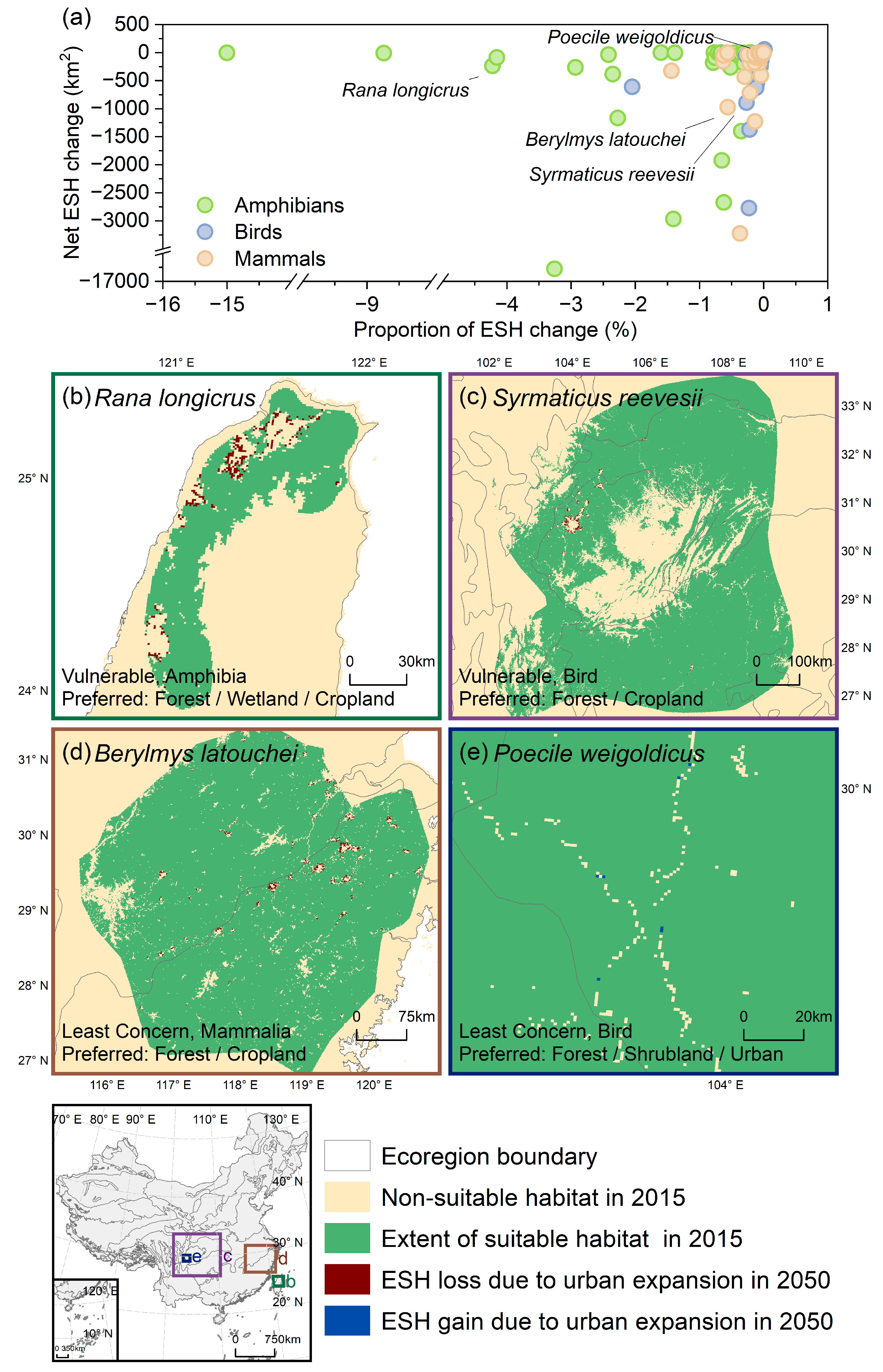
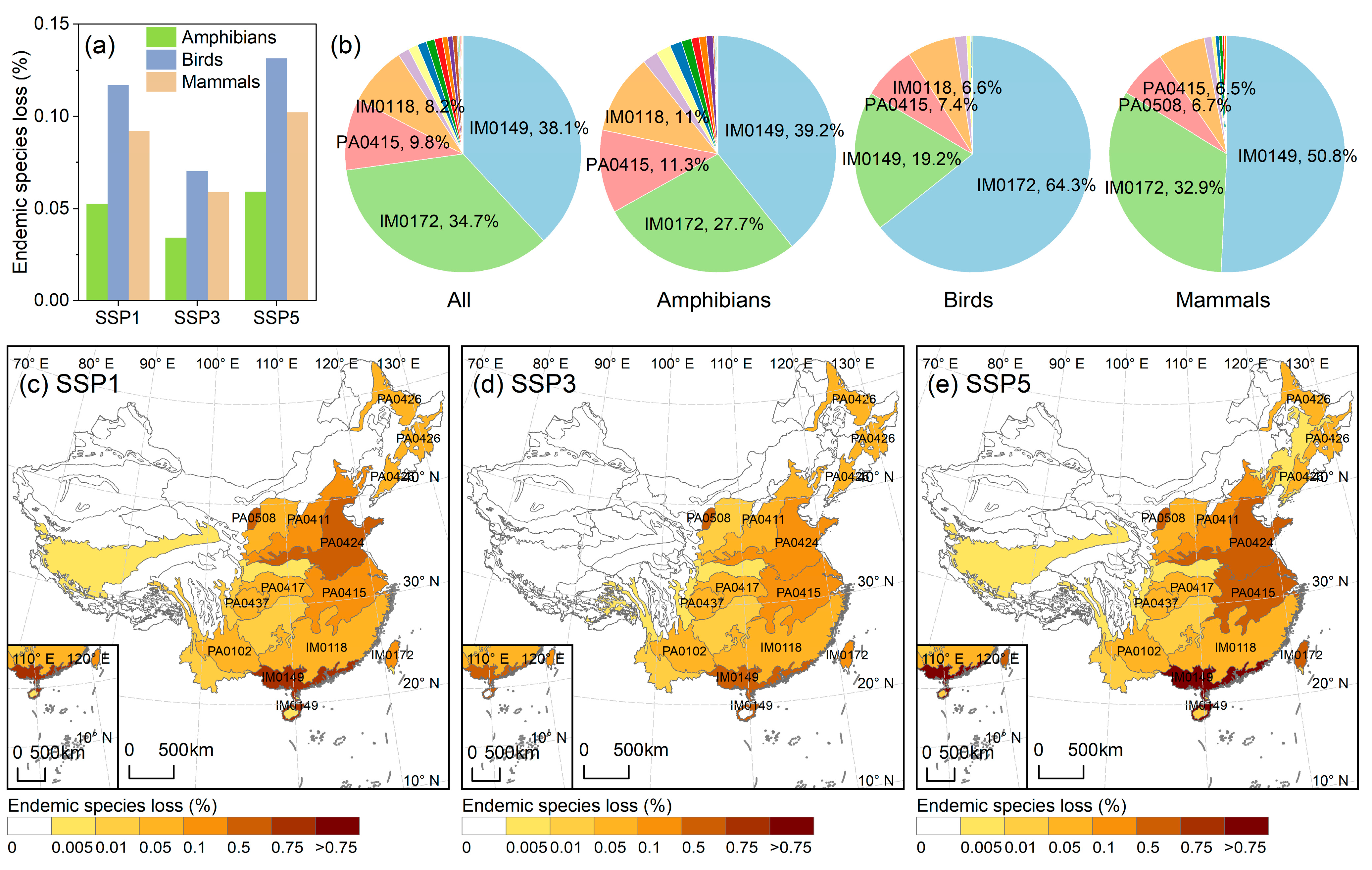
3.4. The Extinction Risk of Ecoregion-Level Endemic Species
4. Discussion
4.1. The Impact of Urban Expansion on Endemic Species
4.2. Implications for Management and Protection
4.3. Limitations and Future Perspectives
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, S.; Zhou, Y.; Yu, R.; Xu, X.; Xu, M.; Li, G.; Wang, W.; Yang, Y. China’s biodiversity conservation in the process of implementing the sustainable development goals (SDGs). J. Clean. Prod. 2022, 338, 130595. [Google Scholar] [CrossRef]
- Pereira, H.M.; Martins, I.S.; Rosa, I.M.D.; Kim, H.; Leadley, P.; Popp, A.; van Vuuren, D.P.; Hurtt, G.; Quoss, L.; Arneth, A.; et al. Global trends and scenarios for terrestrial biodiversity and ecosystem services from 1900 to 2050. Science 2024, 384, 458–465. [Google Scholar] [CrossRef]
- IPBES. Global Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2019. [Google Scholar]
- Tittensor, D.P.; Walpole, M.; Hill, S.L.L.; Boyce, D.G.; Britten, G.L.; Burgess, N.D.; Butchart, S.H.M.; Leadley, P.W.; Regan, E.C.; Alkemade, R.; et al. A mid-term analysis of progress toward international biodiversity targets. Science 2014, 346, 241–244. [Google Scholar] [CrossRef]
- Marques, A.; Martins, I.S.; Kastner, T.; Plutzar, C.; Theurl, M.C.; Eisenmenger, N.; Huijbregts, M.A.J.; Wood, R.; Stadler, K.; Bruckner, M.; et al. Increasing impacts of land use on biodiversity and carbon sequestration driven by population and economic growth. Nat. Ecol. Evol. 2019, 3, 628–637. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, Biodiversity, and Conservation: The impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Simkin, R.D.; Seto, K.C.; McDonald, R.I.; Jetz, W. Biodiversity impacts and conservation implications of urban land expansion projected to 2050. Proc. Natl. Acad. Sci. USA 2022, 119, e2117297119. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Fang, C.; Li, Y.; Wang, Z.; Sun, S.; He, S.; Qi, W.; Bao, C.; Ma, H.; Fan, Y.; et al. Global impacts of future urban expansion on terrestrial vertebrate diversity. Nat. Commun. 2022, 13, 1628. [Google Scholar] [CrossRef]
- Xu, J.; Ling, Y.; Sun, Y.; Jiang, Y.; Shen, R.; Wang, Y. How do different processes of habitat fragmentation affect habitat quality?—Evidence from China. Ecol. Indic. 2024, 160, 111880. [Google Scholar] [CrossRef]
- Zhou, W.; Zhang, S.; Yu, W.; Wang, J.; Wang, W. Effects of Urban Expansion on Forest Loss and Fragmentation in Six Megaregions, China. Remote Sens. 2017, 9, 991. [Google Scholar] [CrossRef]
- Ren, Q.; He, C.; Huang, Q.; Zhang, D.; Shi, P.; Lu, W. Impacts of global urban expansion on natural habitats undermine the 2050 vision for biodiversity. Resour. Conserv. Recycl. 2023, 190, 106834. [Google Scholar] [CrossRef]
- Wurz, A.; Grass, I.; Lees, D.C.; Rakotomalala, A.A.N.A.; Sáfián, S.; Martin, D.A.; Osen, K.; Loos, J.; Benasoavina, E.; Alexis, T.; et al. Land-use change differentially affects endemic, forest and open-land butterflies in Madagascar. Insect Conserv. Divers. 2022, 15, 606–620. [Google Scholar] [CrossRef]
- McDonald, R.I.; Güneralp, B.; Huang, C.-W.; Seto, K.C.; You, M. Conservation priorities to protect vertebrate endemics from global urban expansion. Biol. Conserv. 2018, 224, 290–299. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Seto, K.C.; Güneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Natl. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef] [PubMed]
- Güneralp, B.; Seto, K.C. Futures of global urban expansion: Uncertainties and implications for biodiversity conservation. Environ. Res. Lett. 2013, 8, 014025. [Google Scholar] [CrossRef]
- Walters, G.; Ngagnia Ndjabounda, E.; Ikabanga, D.; Biteau, J.P.; Hymas, O.; White, L.J.T.; Ndong Obiang, A.M.; Ndong Ondo, P.; Jeffery, K.J.; Lachenaud, O.; et al. Peri-urban conservation in the Mondah forest of Libreville, Gabon: Red List assessments of endemic plant species, and avoiding protected area downsizing. Oryx 2016, 50, 419–430. [Google Scholar] [CrossRef]
- Sanz, V.; Caula, S. Assessing bird assemblages along an urban gradient in a Caribbean island (Margarita, Venezuela). Urban Ecosyst. 2015, 18, 729–746. [Google Scholar] [CrossRef]
- Bai, X.; Shi, P.; Liu, Y. Society: Realizing China’s urban dream. Nature 2014, 509, 158–160. [Google Scholar] [CrossRef]
- Hu, Y.n.; Connor, D.S.; Stuhlmacher, M.; Peng, J.; Turner Ii, B.L. More urbanization, more polarization: Evidence from two decades of urban expansion in China. npj Urban Sustain. 2024, 4, 33. [Google Scholar] [CrossRef]
- Li, X.; Chen, Y. Projecting the future impacts of China’s cropland balance policy on ecosystem services under the shared socioeconomic pathways. J. Clean. Prod. 2020, 250, 119489. [Google Scholar] [CrossRef]
- Ministry of Ecology and Environment of the People’s Republic of China; Chinese Academy of Sciences. China’s Red List of Biodiversity: Vertebrates. Biodivers. Sci. 2015, 24, 500–551. [Google Scholar]
- O’Neill, B.C.; Kriegler, E.; Ebi, K.L.; Kemp-Benedict, E.; Riahi, K.; Rothman, D.S.; van Ruijven, B.J.; van Vuuren, D.P.; Birkmann, J.; Kok, K.; et al. The roads ahead: Narratives for shared socioeconomic pathways describing world futures in the 21st century. Glob. Environ. Change 2017, 42, 169–180. [Google Scholar] [CrossRef]
- van Vuuren, D.P.; Stehfest, E.; Gernaat, D.E.H.J.; Doelman, J.C.; van den Berg, M.; Harmsen, M.; de Boer, H.S.; Bouwman, L.F.; Daioglou, V.; Edelenbosch, O.Y.; et al. Energy, land-use and greenhouse gas emissions trajectories under a green growth paradigm. Glob. Environ. Change 2017, 42, 237–250. [Google Scholar] [CrossRef]
- Fujimori, S.; Hasegawa, T.; Masui, T.; Takahashi, K.; Herran, D.S.; Dai, H.; Hijioka, Y.; Kainuma, M. SSP3: AIM implementation of Shared Socioeconomic Pathways. Glob. Environ. Change 2017, 42, 268–283. [Google Scholar] [CrossRef]
- Kriegler, E.; Bauer, N.; Popp, A.; Humpenöder, F.; Leimbach, M.; Strefler, J.; Baumstark, L.; Bodirsky, B.L.; Hilaire, J.; Klein, D.; et al. Fossil-fueled development (SSP5): An energy and resource intensive scenario for the 21st century. Glob. Environ. Change 2017, 42, 297–315. [Google Scholar] [CrossRef]
- Chen, G.; Li, X.; Liu, X.; Chen, Y.; Liang, X.; Leng, J.; Xu, X.; Liao, W.; Qiu, Y.; Wu, Q.; et al. Global projections of future urban land expansion under shared socioeconomic pathways. Nat. Commun. 2020, 11, 537. [Google Scholar] [CrossRef]
- Pesaresi, M.; Huadong, G.; Blaes, X.; Ehrlich, D.; Ferri, S.; Gueguen, L.; Halkia, M.; Kauffmann, M.; Kemper, T.; Lu, L.; et al. A Global Human Settlement Layer From Optical HR/VHR RS Data: Concept and First Results. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2013, 6, 2102–2131. [Google Scholar] [CrossRef]
- Liu, X.; Liang, X.; Li, X.; Xu, X.; Ou, J.; Chen, Y.; Li, S.; Wang, S.; Pei, F. A future land use simulation model (FLUS) for simulating multiple land use scenarios by coupling human and natural effects. Landsc. Urban Plan. 2017, 168, 94–116. [Google Scholar] [CrossRef]
- Pontius, R.G.; Boersma, W.; Castella, J.C.; Clarke, K.; de Nijs, T.; Dietzel, C.; Duan, Z.; Fotsing, E.; Goldstein, N.; Kok, K.; et al. Comparing the input, output, and validation maps for several models of land change. Ann. Reg. Sci. 2008, 42, 11–37. [Google Scholar] [CrossRef]
- Li, X.; Chen, G.; Liu, X.; Liang, X.; Wang, S.; Chen, Y.; Pei, F.; Xu, X. A New Global Land-Use and Land-Cover Change Product at a 1-km Resolution for 2010 to 2100 Based on Human-Environment Interactions. Ann. Am. Assoc. Geogr. 2017, 107, 1040–1059. [Google Scholar] [CrossRef]
- Chen, Y.M.; Li, X.; Liu, X.P.; Ai, B. Modeling urban land-use dynamics in a fast developing city using the modified logistic cellular automaton with a patch-based simulation strategy. Int. J. Geogr. Inf. Sci. 2014, 28, 234–255. [Google Scholar] [CrossRef]
- Ren, Q.; He, C.; Huang, Q.; Shi, P.; Zhang, D.; Güneralp, B. Impacts of urban expansion on natural habitats in global drylands. Nat. Sustain. 2022, 5, 869–878. [Google Scholar] [CrossRef]
- Menéndez, R.; Megías, A.G.; Hill, J.K.; Braschler, B.; Willis, S.G.; Collingham, Y.; Fox, R.; Roy, D.B.; Thomas, C.D. Species richness changes lag behind climate change. Proc. R. Soc. B Biol. Sci. 2006, 273, 1465–1470. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Gao, Z.; Su, Y.; Lin, S.; Jiang, H. Geographical and temporal origins of terrestrial vertebrates endemic to Taiwan. J. Biogeogr. 2018, 45, 2458–2470. [Google Scholar] [CrossRef]
- Qiao, L.; Wen, G.; Qi, Y.; Lu, B.; Hu, J.; Song, Z.; Fu, J. Evolutionary melting pots and reproductive isolation: A ring-shaped diversification of an odorous frog (Odorrana margaratea) around the Sichuan Basin. Mol. Ecol. 2018, 27, 4888–4900. [Google Scholar] [CrossRef]
- Powers, R.P.; Jetz, W. Global habitat loss and extinction risk of terrestrial vertebrates under future land-use-change scenarios. Nat. Clim. Change 2019, 9, 323–329. [Google Scholar] [CrossRef]
- Li, F.; Wu, S.; Liu, H.; Yan, D. Biodiversity loss through cropland displacement for urban expansion in China. Sci. Total. Environ. 2024, 907, 167988. [Google Scholar] [CrossRef]
- Halley, J.M.; Sgardeli, V.; Triantis, K.A. Extinction debt and the species–area relationship: A neutral perspective. Glob. Ecol. Biogeogr. 2014, 23, 113–123. [Google Scholar] [CrossRef]
- Chaudhary, A.; Brooks, T.M. National Consumption and Global Trade Impacts on Biodiversity. World Dev. 2019, 121, 178–187. [Google Scholar] [CrossRef]
- Proença, V.; Pereira, H.M. Species–area models to assess biodiversity change in multi-habitat landscapes: The importance of species habitat affinity. Basic Appl. Ecol. 2013, 14, 102–114. [Google Scholar] [CrossRef]
- Pereira, H.M.; Ziv, G.U.Y.; Miranda, M. Countryside Species—Area Relationship as a Valid Alternative to the Matrix—Calibrated Species—Area Model. Conserv. Biol. 2014, 28, 874–876. [Google Scholar] [CrossRef]
- Drakare, S.; Lennon, J.J.; Hillebrand, H. The imprint of the geographical, evolutionary and ecological context on species–area relationships. Ecol. Lett. 2006, 9, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Leclère, D.; Obersteiner, M.; Barrett, M.; Butchart, S.H.M.; Chaudhary, A.; De Palma, A.; DeClerck, F.A.J.; Di Marco, M.; Doelman, J.C.; Dürauer, M.; et al. Bending the curve of terrestrial biodiversity needs an integrated strategy. Nature 2020, 585, 551–556. [Google Scholar] [CrossRef]
- Bren d’Amour, C.; Reitsma, F.; Baiocchi, G.; Barthel, S.; Guneralp, B.; Erb, K.H.; Haberl, H.; Creutzig, F.; Seto, K.C. Future urban land expansion and implications for global croplands. Proc. Natl. Acad. Sci. USA 2017, 114, 8939–8944. [Google Scholar] [CrossRef]
- Oliver, T.H.; Marshall, H.H.; Morecroft, M.D.; Brereton, T.; Prudhomme, C.; Huntingford, C. Interacting effects of climate change and habitat fragmentation on drought-sensitive butterflies. Nat. Clim. Change 2015, 5, 941–945. [Google Scholar] [CrossRef]
- Wang, X.; Cumming, S.G. Modeling configuration dynamics of harvested forest landscapes in the Canadian boreal plains. Landsc. Ecol. 2009, 24, 229–241. [Google Scholar] [CrossRef]
- Wang, X.; Blanchet, F.G.; Koper, N. Measuring habitat fragmentation: An evaluation of landscape pattern metrics. Methods Ecol. Evol. 2014, 5, 634–646. [Google Scholar] [CrossRef]
- Rogan, J.E.; Lacher, T.E. Impacts of Habitat Loss and Fragmentation on Terrestrial Biodiversity. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Willmer, J.N.G.; Püttker, T.; Prevedello, J.A. Global impacts of edge effects on species richness. Biol. Conserv. 2022, 272, 109654. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Chen, R.; Carruthers-Jones, J.; Carver, S.; Wu, J. Constructing urban ecological corridors to reflect local species diversity and conservation objectives. Sci. Total Environ. 2024, 907, 167987. [Google Scholar] [CrossRef] [PubMed]
- Tambosi, L.R.; Martensen, A.C.; Ribeiro, M.C.; Metzger, J.P. A Framework to Optimize Biodiversity Restoration Efforts Based on Habitat Amount and Landscape Connectivity. Restor. Ecol. 2014, 22, 169–177. [Google Scholar] [CrossRef]
- Yang, X.; Tan, X.; Chen, C.; Wang, Y. The influence of urban park characteristics on bird diversity in Nanjing, China. Avian Res. 2020, 11, 45. [Google Scholar] [CrossRef]
- Chiquet, C.; Dover, J.W.; Mitchell, P. Birds and the urban environment: The value of green walls. Urban Ecosyst. 2013, 16, 453–462. [Google Scholar] [CrossRef]
- Luedtke, J.A.; Chanson, J.; Neam, K.; Hobin, L.; Maciel, A.O.; Catenazzi, A.; Borzée, A.; Hamidy, A.; Aowphol, A.; Jean, A.; et al. Ongoing declines for the world’s amphibians in the face of emerging threats. Nature 2023, 622, 308–314. [Google Scholar] [CrossRef]
- Purvis, A.; Agapow, P.-M.; Gittleman, J.L.; Mace, G.M. Nonrandom Extinction and the Loss of Evolutionary History. Science 2000, 288, 328–330. [Google Scholar] [CrossRef]
- Rodrigues, A.S.L.; Andelman, S.J.; Bakarr, M.I.; Boitani, L.; Brooks, T.M.; Cowling, R.M.; Fishpool, L.D.C.; da Fonseca, G.A.B.; Gaston, K.J.; Hoffmann, M.; et al. Effectiveness of the global protected area network in representing species diversity. Nature 2004, 428, 640–643. [Google Scholar] [CrossRef]
- McDonald, R.I.; Mansur, A.V.; Ascensão, F.; Colbert, M.l.; Crossman, K.; Elmqvist, T.; Gonzalez, A.; Güneralp, B.; Haase, D.; Hamann, M.; et al. Research gaps in knowledge of the impact of urban growth on biodiversity. Nat. Sustain. 2019, 3, 16–24. [Google Scholar] [CrossRef]
- Zecheng, W.; Xinsheng, Z.; Lijuan, C.; Yinru, L.; Ziliang, G.; Jinzhi, W.; Jing, L.; Xiajie, Z.; Rumiao, W.; Wei, L. Identification and optimization of urban wetland ecological networks in highly urbanized areas: A case study of Haidian District, Beijing. Ecol. Indic. 2025, 170, 113028. [Google Scholar] [CrossRef]
- Fernández, I.C.; Simonetti, J.A. Small mammal assemblages in fragmented shrublands of urban areas of Central Chile. Urban Ecosyst. 2013, 16, 377–387. [Google Scholar] [CrossRef]
- Pinheiro, R.T.; Dornas, T.; Reis, E.D.; Barbosa, M.D.; Rodello, D. Birds of the urban area of Palmas, TO: Composition and conservation. Rev. Bras. Ornitol. 2008, 16, 339–347. [Google Scholar]
- Smith, C.V.; Gilbert, T.C.; Woodfine, T.; Kraaijeveld, A.; Chege, G.; Kimiti, D.; Low-Mackey, B.; Mutinda, M.; Ngene, S.; Rubenstein, D.; et al. Population and habitat connectivity of Grevy’s zebra Equus grevyi, a threatened large herbivore in degraded rangelands. Biol. Conserv. 2022, 274, 109711. [Google Scholar] [CrossRef]
- Mi, X.; Feng, G.; Hu, Y.; Zhang, J.; Chen, L.; Corlett, R.T.; Hughes, A.C.; Pimm, S.; Schmid, B.; Shi, S.; et al. The global significance of biodiversity science in China: An overview. Natl. Sci. Rev. 2021, 8, nwab032. [Google Scholar] [CrossRef] [PubMed]
- Semenchuk, P.; Kalt, G.; Kaufmann, L.; Kastner, T.; Matej, S.; Bidoglio, G.; Erb, K.-H.; Essl, F.; Haberl, H.; Dullinger, S.; et al. The global biodiversity footprint of urban consumption: A spatially explicit assessment for the city of Vienna. Sci. Total Environ. 2023, 861, 160576. [Google Scholar] [CrossRef]
- Keil, P.; Storch, D.; Jetz, W. On the decline of biodiversity due to area loss. Nat. Commun. 2015, 6, 8837. [Google Scholar] [CrossRef]
- Jin, X.; Qian, S.; Yuan, J. Identifying urban rewilding opportunity spaces in a metropolis: Chongqing as an example. Ecol. Indic. 2024, 160, 111778. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Type | ESA CCI-LC Class Value |
|---|---|
| Urban | 190 |
| Cropland | 10/11/12/20/30 |
| Forest | 40/50/60/61/62/70/71/72/80/81/82/90/100 |
| Grassland | 110/130 |
| Shrubland | 120/121/122 |
| Bare land | 140/150/151/152/153/200/201/202/220 |
| Water and Wetland | 160/170/180/210 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Wu, X.; Liu, X. Impact of Future Urban Expansion on Endemic Species in China at the Species Level. Land 2025, 14, 1005. https://doi.org/10.3390/land14051005
Liu K, Wu X, Liu X. Impact of Future Urban Expansion on Endemic Species in China at the Species Level. Land. 2025; 14(5):1005. https://doi.org/10.3390/land14051005
Chicago/Turabian StyleLiu, Kangyao, Xinxin Wu, and Xiaoping Liu. 2025. "Impact of Future Urban Expansion on Endemic Species in China at the Species Level" Land 14, no. 5: 1005. https://doi.org/10.3390/land14051005
APA StyleLiu, K., Wu, X., & Liu, X. (2025). Impact of Future Urban Expansion on Endemic Species in China at the Species Level. Land, 14(5), 1005. https://doi.org/10.3390/land14051005