Abstract
Soil microbes are critical in regulating the growth and function of eucalyptus plantations. The mechanisms underlying soil microbial communities’ response to different eucalyptus plantation management practices remain elusive. In this study, we compiled datasets containing 2744 observations across global eucalyptus-planted regions and analyzed the effects of five management practices (i.e., burning, residual removal, fertilization, mixed planting, and controlling planting years) on soil microbial biomass, diversity, and structures. Our results showed that fungal community alpha diversity responds more sensitively to management practices than bacterial community alpha diversity on eucalyptus plantations. Although the implementation of management practices significantly increased the content of most soil nutrients and microbial biomass elements (excluding burning), these practices did not necessarily improve soil microbial biomass and diversity, particularly among fungal communities. Burning, fertilization, and mixing eucalyptus with nitrogen-fixing species significantly decreased the diversity of fungal communities, which were mainly impacted by soil organic carbon and total potassium content. Compared to the four other management practices, mixing eucalyptus with nitrogen-fixing species favored the growth of bacterial communities and the storage of microbial biomass nitrogen, making it the most effective management practice. However, attention should also be paid to the protection of fungal communities. In addition, these management practices significantly changed microbial community structures, which were positively correlated with the microbial biomass elements carbon and nitrogen and, to a lesser extent, soil microbial alpha diversity. Our results highlight the importance of prioritizing mixing eucalyptus with nitrogen-fixing species as a management practice and safeguarding fungal community diversity during its implementation and suggest that microbial diversity development associated with soil organic carbon and potassium contents should be given priority in eucalyptus plantation management.
1. Introduction
Eucalyptus trees exhibit rapid growth, even in nutrient-poor soils, outperforming species such as Acacia, Pinus, and other common plantation species [1,2,3,4]. These trees are capable of efficiently absorbing nutrients from highly weathered soils in tropical and subtropical regions, enabling them to produce substantial biomass in a relatively short period [5]. Consequently, eucalyptus plantations have been successfully established across various soil types, providing both commercial and subsistence opportunities for rural smallholders [6]. Furthermore, owing to its significant carbon sequestration potential, eucalyptus plays a vital role in maintaining carbon balance under global climate change conditions [7,8]. These advantages have thus enabled eucalyptus to become one of the most widely utilized species for afforestation efforts globally, with a broad distribution across Australia, Asia, Africa, the Americas, and other regions [8]. However, the results of a number of studies have indicated that during the growth of eucalyptus, it can consume large amounts of water and nutrients, substantially depleting soil resources and impacting the global flux of nitrogen dioxide [9,10]. The allelopathic effects of eucalyptus can reduce understory plant biomass and diversity, which not only diminishes soil resource availability but also induces deterioration in local microclimates, leading to a loss of biodiversity [11,12,13,14]. At present, a series of management practices have been implemented around the globe to mitigate the adverse impacts of eucalyptus and optimize its benefits. These practices include burning, fertilization, mixed planting, residue removal, and controlling planting age [15,16,17,18,19]. The results of previous studies suggest that artificial interventions on eucalyptus plantations are beneficial for eucalyptus growth and soil fertility [17,19,20,21]. However, the results of other studies indicate that such interventions may negatively impact understory and soil biodiversity, thereby hindering the sustainable development of eucalyptus plantations [15,16,18,22]. At present, the effects of different management practices on eucalyptus plantations remain unclear.
Microorganisms play a vital role in ecosystems, with them being widely involved in the processes of carbon and nitrogen cycling, in addition to soil structure formation [15,17,19]. Compared to soil properties, they exhibit higher sensitivity to environmental changes, and variations in their biomass, diversity, and structure are considered early indicators of soil ecosystem changes [23,24]. A series of management practices implemented by personnel on eucalyptus plantations can directly or indirectly (by affecting soil properties and aboveground vegetation) impact the biomass, structure, and diversity of soil microorganism communities [18,19,20,21,25]. For example, burning and residual removal can effectively mitigate the accumulation of surface fuels while concurrently diminishing the influx of carbon and nitrogen nutrients into the soil via leaching from litter, resulting in a substantial reduction in microbial biomass and alterations in microbial community structures [16,22,24,25,26]. However, the results of some studies suggest that the thermal effects of fire can stimulate the germination of numerous plant species within ecosystems [27] and stimulate spore germination of heat-tolerant species, which induces soil microbial activity [28,29]. In contrast, removing accumulated litter from a forest area can enhance soil aeration and water infiltration, in addition to promoting the growth of a wider variety of plants [30]. These factors, in turn, provide a wider range of ecological niches and nutritional resources for soil microorganisms, ultimately enhancing soil microbial diversity and activity [25,30]. Fertilization and mixing nitrogen-fixing species with eucalyptus mainly impact the microbial community through the addition of soil nitrogen supply and the enhancement of soil nitrogen utilization efficiency [17,18,19,31,32]. The introduction of nitrogen-fixing species can enhance aboveground vegetation diversity by promoting photosynthesis, further altering microbial community diversity and structure [18,20,33]. However, although fertilization has the potential to enhance soil nutrient contents [34,35], its influence on microorganisms is not consistently beneficial [36,37,38]. In their study, Hasselquist et al. [38] found that the enrichment of the nitrogen fertilizer inhibits saprophytic fungi while activating certain mycorrhizal fungi. Regarding timber, the determination of eucalyptus rotation periods often hinges on considerations of economic profitability and carbon sequestration potential, with limited regard for their effects on soil quality [21,39]. Shorter rotation periods typically yield higher productivity but lead to soil nutrient imbalances [20,40,41]. Conversely, longer rotation periods allow for the gradual opening of eucalyptus canopies, which promotes understory biodiversity and accelerates litter decomposition rates [42]. These processes benefit soil nutrient reserves [39,43] and support the resilience and development of microbial communities [21]. Nonetheless, the results of some studies suggest that with the aging of eucalyptus stands, the allelopathic effects lead to notable declines in the diversity of undisturbed vegetation and the content of soil nutrients, thereby impacting soil enzyme activity, microbial diversity, and community structure [44,45,46,47].
The inconsistent results presented in the above studies may stem from variations in management practices, plantation age, and climatic conditions at the location under study. Individual studies, unfortunately, often lack the capacity to comprehensively account for all of these influencing factors. To address the above issues, some researchers have employed meta-analysis to evaluate the effects of artificial management practices on forest soil properties and microbial communities. For instance, Guo et al. [48] and Curtright et al. [49] assessed the impact of mixed-species planting on soil properties, enzyme activity, and microbial biomass in plantation forests. The results of studies by Wang et al. [26] and Zhou et al. [50] demonstrated that nitrogen fertilization led to a significant decrease in microbial diversity and shift in microbial community composition in plantation forests. Furthermore, the results of several studies have highlighted the effects of forest residue harvesting on the soil nutrient content and microbial biomass [51,52]. However, the authors of these studies predominantly focused on individual management practices within global forests, primarily investigating their influence on soil properties, enzyme activity, microbial diversity, and community structure. At present, there are no studies specifically focusing on eucalyptus plantations that examine in detail the effects of artificial management practices on microbial biomass, diversity, and community structure across different microbial taxa. In light of this gap, it is necessary to conduct a meta-analysis of multiple studies to determine the effect of management practices on soil microorganisms on eucalyptus plantations, specifically in the different microbial taxa. The results of this study can provide scientific guidance for subsequent eucalyptus plantation management strategies and contribute to sustainable development.
To accomplish the above research aims, we conducted a meta-analysis of 2744 observations, including soil properties, enzyme activity, microbial biomass, microbial community diversity, and structure. The management practices implemented on eucalyptus plantations included burning, fertilization, mixed planting, residue removal, and control of planting age, which are five common practices. The primary aims of this study were as follows: (1) to analyze the effects of different management practices on the biomass of different soil microbial taxa, diversity, and structure on eucalyptus plantations on a global scale and (2) to explore the key factors driving different soil microbial taxa biomass, diversity, and structure on eucalyptus plantations globally. Through the following study, we aim to provide scientific recommendations for the subsequent management of eucalyptus plantations.
2. Materials and Methods
2.1. Data Collection
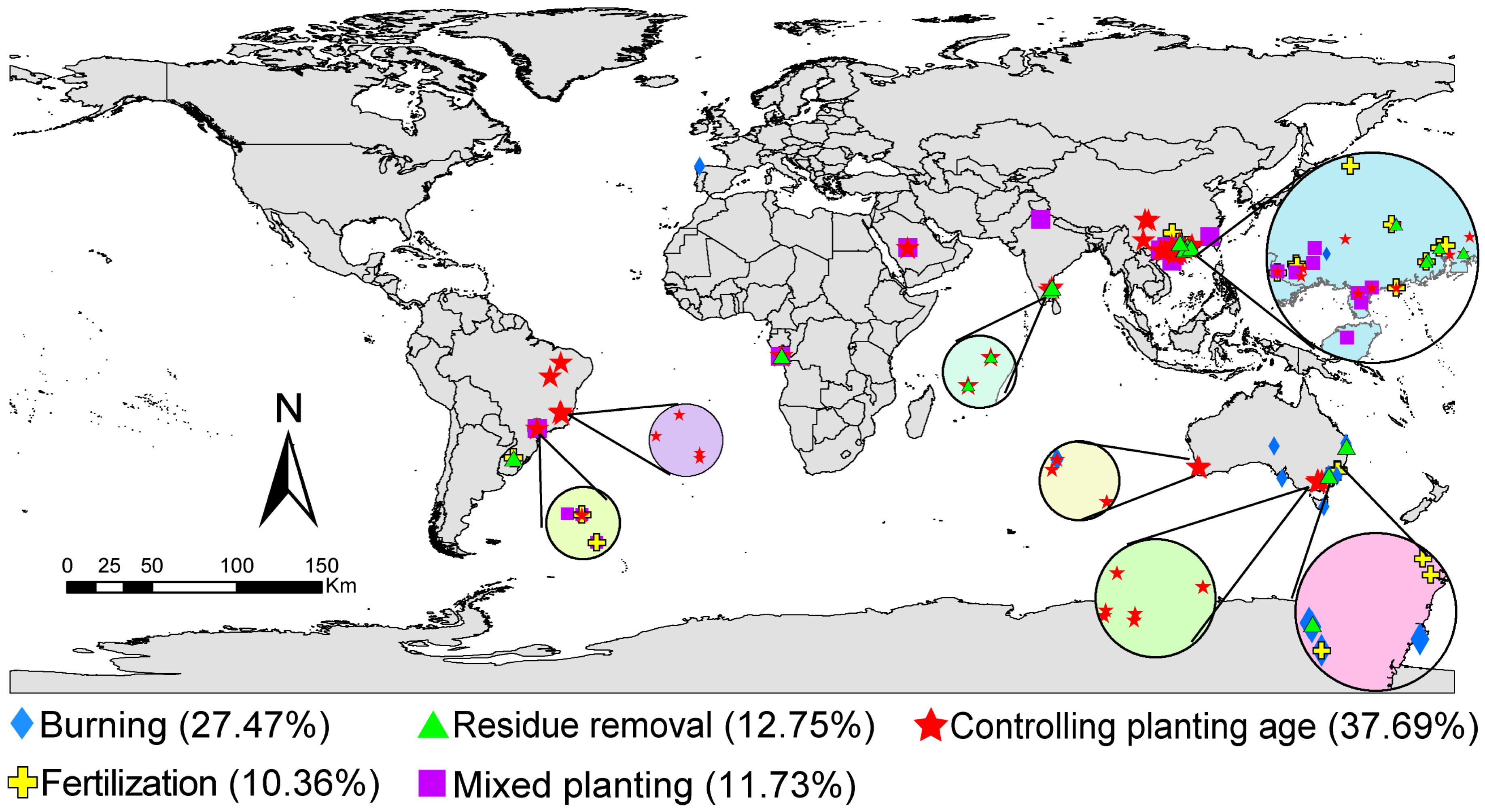
We collected all articles published prior to May 2023 from Web of Science (https://apps.webofknowledge.com (accessed on 30 April 2023)) and the China Knowledge Resource Integrated Database (https://www.cnki.net (accessed on 30 April 2023)) addressing the following topics: eucalyptus plantations and soil microorganisms, soil fungi, and soil bacteria. In total, 2744 observations were collected from 103 articles (Figure 1 and Supplementary S1). To mitigate potential biases arising from variations across studies, we implemented stringent inclusion criteria during data collection. These criteria were as follows: (1) The article reports at least one microbial indicator, including microbial biomass elements (microbial biomass carbon (MBC), microbial biomass nitrogen (MBN), or microbial biomass phosphorus (MBP)), microbial composition biomass (PLFA, bacterial biomass, fungal biomass, Gram-positive bacterial biomass, Gram-negative bacterial biomass, or Actinomycetes biomass), alpha diversity index, beta diversity, and community structure; (2) the availability of sample size, mean, and standard deviation, either reported directly or calculated from tables, digitized charts, or textual descriptions; (3) the inclusion of both field experiments and laboratory cultures; and (4) for factorial experiments, the selection of only one management practice as the control and treatment factor, excluding any interaction factors. Each study underwent independent screening by two reviewers to determine compliance with these criteria. In cases of disagreement, additional reviewers were engaged to reach consensus. Data extraction from the figures presented in the papers was conducted using GetData Graph Digitizer software (version 2.20, http://www.getdata-graph-digitizer.com/index.php (accessed on 30 April 2023)) to extract the mean values and standard deviation.
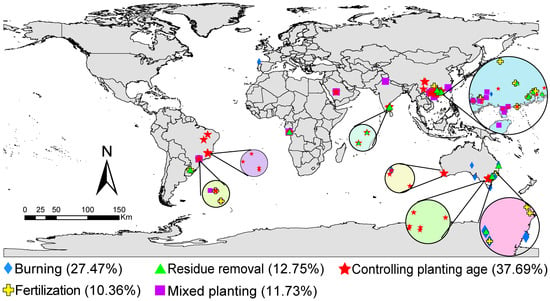
Figure 1.
Soil sampling sites.
The information collected by our group also includes site locations (latitude, longitude, and altitude), climate conditions (mean annual temperature (MAT) and mean annual precipitation (MAP)), soil physical properties (bulk density (BD) and soil moisture content (SMC)), soil chemical properties (total carbon (TC), soil organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), total potassium (TK), and pH), soil enzymes (catalase, urease, and invertase), and experimental forcing factors (burning, fertilization, mixed planting, residue removal, and controlling planting age).
2.2. Data Calculations
For each case study, the effect of the practice on each variable was assessed and computed after transformation into natural logarithms (Table S1):
where and represent the mean values of the respective variable in the control and treatment groups, respectively. The change in soil pH demonstrated the impact of the practice on soil pH; the difference in pH between the treatment and control was calculated based on the method outlined in a previous meta-analysis [53]. The corresponding sample variance () was calculated as follows:
where and represent the replicate numbers of the treatment and control groups, respectively, and and represent the standard deviation of the treatment and control groups, respectively.
Microbial alpha diversity is generally evaluated using the Shannon and Richness indices [26,54]. Notably, the Chao, ACE, and OTU indices are commonly reported metrics for microbial richness in the literature.
The methodologies employed for analyzing microbial community structure in the reviewed articles predominantly encompassed redundancy analysis, principal coordinate analysis, principal component analysis, and non-metric multidimensional scaling analysis. These methods revealed discernible differences in soil microbial community structure between the control and treatment conditions [55]. The approach of Zhou et al. [56] was used to assess microbial community structure. Specifically, sample positions along the first two ordination axes were extracted, the “vegan” package was utilized to compute the Euclidean distance among different samples [57], and the RR of the microbial community structure () and the RR of beta diversity () were calculated as follows:
where is the distance within the control, is the distance within the treatment, is the distance between the control and the treatment, and , , and represent the means of , , and .
indicates that the practice has no effect on microbial community structure; indicates that there is a notable alteration in the community structure; and indicates that beta diversity was increased by the management practice.
2.3. Statistical Analyses
Egger’s regression test and sensitivity analyses were conducted to test for potential publication bias and to determine the robustness of the meta-analysis (Table S2) [58]. Any outliers were excluded before analyses [51]. In addition, we assessed the heterogeneity of effect sizes using formal Cochran’s Q-test to determine whether the variability in the observed effect sizes was larger than that expected by chance (Table S2) [59]. Based on Cochran’s Q-test results, we chose either a random effects or fixed effects model to assess the impact of management practices on soil properties and microbial indicators using the “Metafor” package [60]. The significance of the difference between management practice treatment and control was determined for each variable based on whether the CI of the RR overlapped with zero [56].
Model selection relied on AIC correction. The relative importance of a given predictor was determined by summing the Akaike weights across all models in which the predictor appeared. Consequently, predictors featured in models with higher Akaike weights received greater importance values, indicating stronger overall support across all models. For this process, the “gmulti” package was utilized [61]. Four types of candidate predictors were considered: (1) climate factors, (2) soil properties, (3) soil enzymes, and (4) management practices. Lastly, we employed linear regression models to investigate the associations between the RR of microbial indicators and predictor variables, in addition to the relationship between the RR of microbial indicators (microbial biomass and alpha diversity) and beta diversity and structure. All statistical analyses were conducted in R software (Version 4.0.2).
3. Results
3.1. Variation in Soil Microbial Community Biomass, Diversity, and Structure
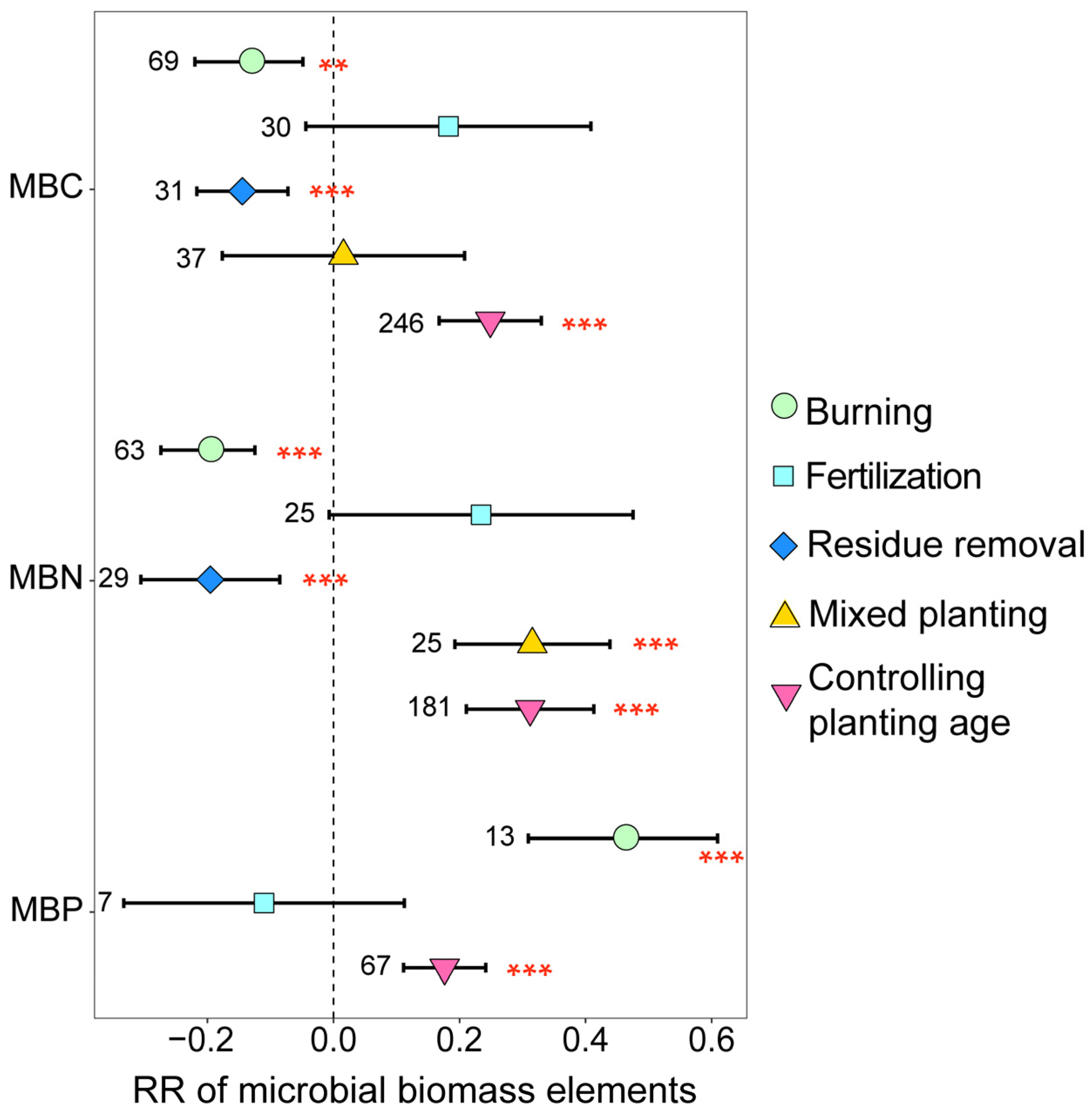
The soil microbial biomass elements of eucalyptus plantations were significantly influenced by the management practices employed (Figure 2). Both burning and residue removal significantly decreased MBC and MBN; in comparison, burning notably increased MBP (p < 0.05). Mixed planting significantly increased MBN (p < 0.001). Additionally, with the increase in planting age, MBC, MBN, and MBP significantly increased (p < 0.05). These findings demonstrate that implementing management practices on eucalyptus plantations does not always significantly increase the abundance of soil microbial biomass elements, with MBN being more sensitive to environmental changes compared to MBC.
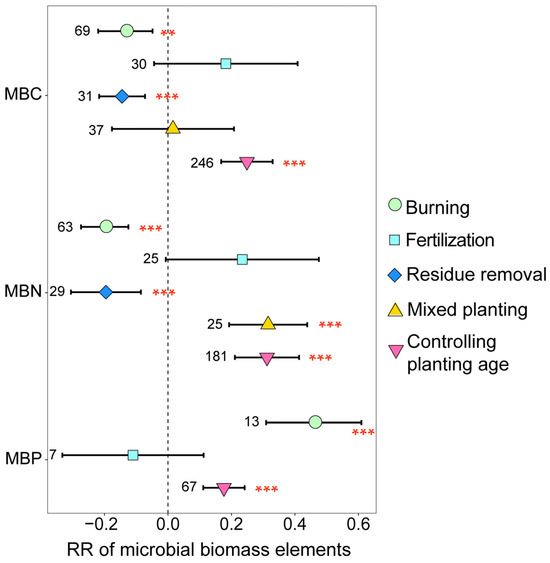
Figure 2.
The response ratios of soil microbial biomass elements under different management practices. The values beside the bars are the corresponding number of observations. *** and ** indicates p < 0.001 and p < 0.01, respectively.
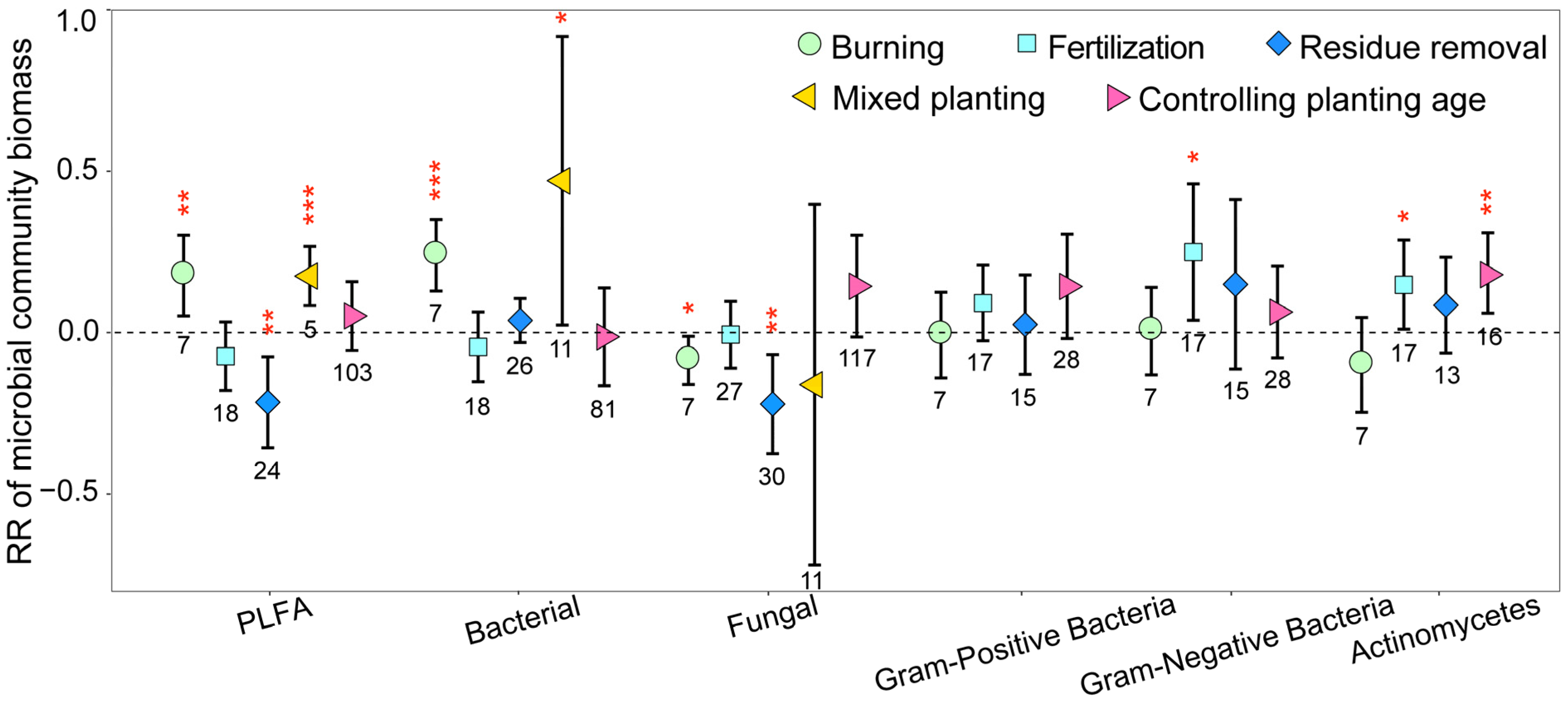
The microbial biomass of different species on eucalyptus plantations was also influenced by management practices employed, with the biomass of most species significantly increasing as a result of these practices, whereas fungal biomass significantly decreased (Figure 3). After burning, total microbial and bacterial biomass significantly increased; in comparison, fungal biomass significantly decreased (p < 0.05). Residue removal significantly decreased total microbial and fungal biomass (p < 0.01), and mixed planting significantly increased total microbial and bacterial biomass (p < 0.05). In addition, fertilization significantly increased Gram-negative bacterial biomass; in comparison, the increase in planting age significantly decreased Actinomycetes biomass (p < 0.05).
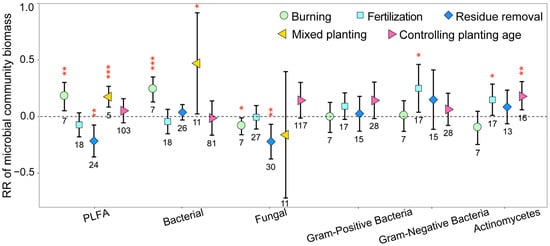
Figure 3.
The response ratios of soil microbial biomass of different species under different management practices. The values shown at the bottom of the bars represent the corresponding number of observations. ***, ** and * indicates p < 0.001, p < 0.01 and p < 0.05, respectively.
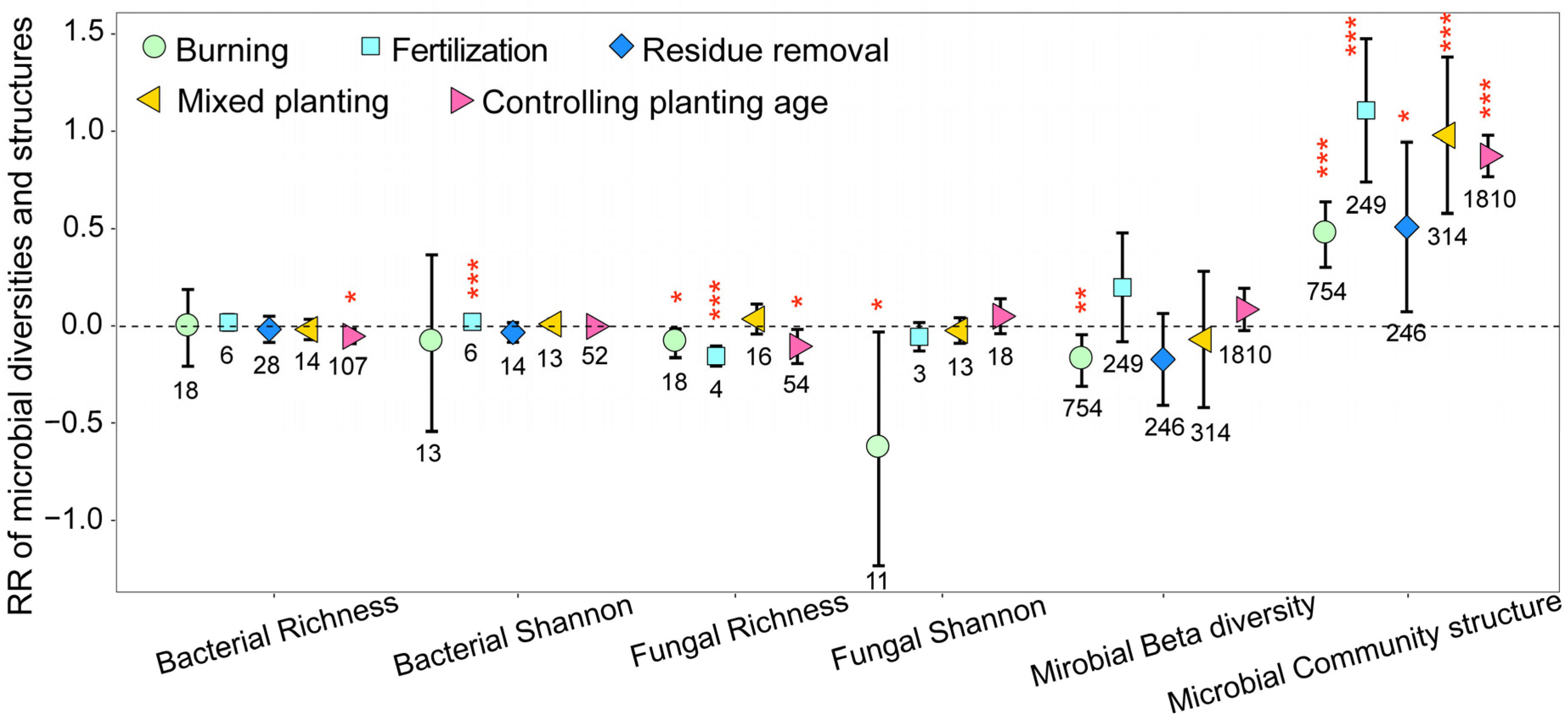
The different management practices significantly decreased fungal community richness on eucalyptus plantations, with it being important to note that these practices may not consistently have a positive effect on the alpha diversity of bacterial communities (Figure 4). In addition, the management practices employed could possibly even reduce microbial beta diversity. Nonetheless, all management practices significantly impacted the soil microbial community structure (Figure 4).
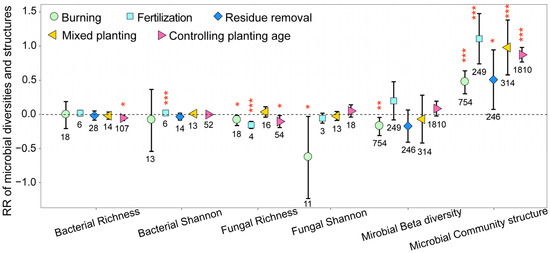
Figure 4.
The response ratios of soil microbial alpha diversity and community structures under different management practices. The values shown at the bottom of the bars represent the corresponding number of observations. ***, ** and * indicates p < 0.001, p < 0.01 and p < 0.05, respectively.
As shown in Figure 4, burning significantly decreased the diversity and richness of soil fungal communities on eucalyptus plantations while having no effect on the diversity and richness of bacterial communities. These results suggest that the post-burning environment is unfavorable for the survival of fungal communities. The absolute values of the RR of bacterial and fungal Shannon indices were greater than those of the RR of bacterial and fungal richness indices, suggesting that rare species were more sensitive to burning. Similarly, the absolute values of the RR of bacterial and fungal richness indices on the mixed planting and older-age plantations were greater than those of the RR of bacterial and fungal Shannon indices, indicating that abundant species were more sensitive to the practice of mixed planting and increased planting age. In addition, with the increase in planting age, the richness of both soil fungal and bacterial communities significantly decreased (p < 0.05). In addition, we found that fungal community alpha diversity responds more strongly than bacterial community alpha diversity to different management practices.
Under all management practices, the RR of microbial community structures was greater than 0, particularly for fertilization, mixed planting, and increased planting age, which indicated that management practices significantly modified the microbial community structures, and microbial community structures were more sensitive than microbial alpha diversity to different management practices.
3.2. Variation in Soil Properties and Enzymes
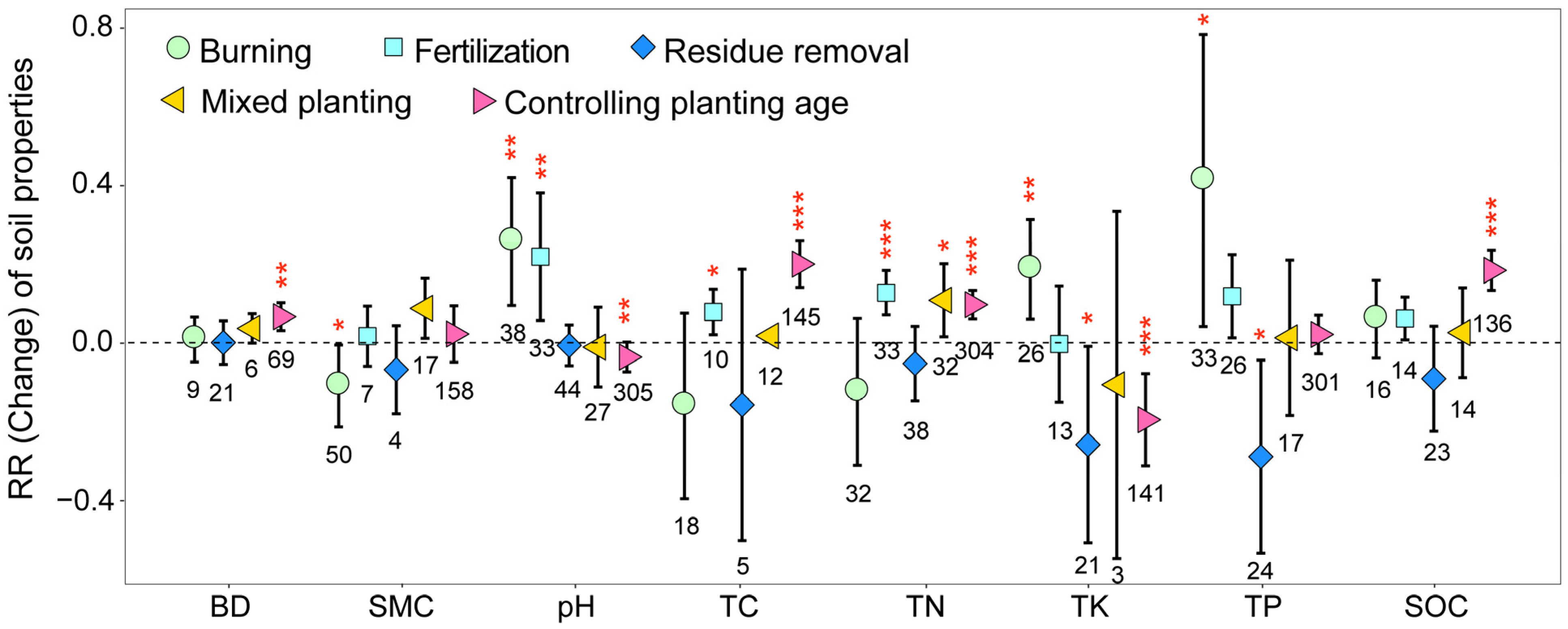
The implementation of different management practices on eucalyptus plantations significantly influenced the soil nutrient content while concurrently having no significant impact on soil aeration and the moisture content (Figure 5). Burning significantly increased soil TK, TP, and pH while significantly decreasing SMC (p < 0.05). Residue removal significantly decreased soil TK and TP; in comparison, mixed planting significantly increased soil TN (p < 0.05). Fertilization significantly increased soil TC, TN, and pH (p < 0.05). In addition, as planting age increased, soil BD, TC, TN, and SOC exhibited significant increases; in comparison, soil TK and pH showed significant decreases (p < 0.05). These results illustrated that most artificial management practices significantly increased the soil nutrient content on eucalyptus plantations.
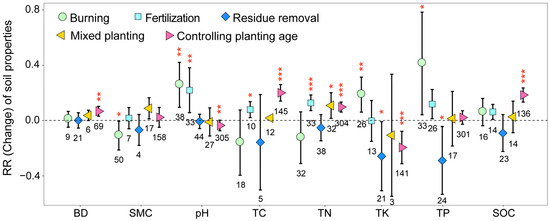
Figure 5.
Response ratios of soil properties under different management practices. The values shown at the bottom of the bars represent the corresponding number of observations. BD: soil bulk density; SMC: soil moisture content; TC: total carbon; TN: total nitrogen; TP: total phosphorus; TK: total potassium; SOC: soil organic carbon. ***, ** and * indicates p < 0.001, p < 0.01 and p < 0.05, respectively.
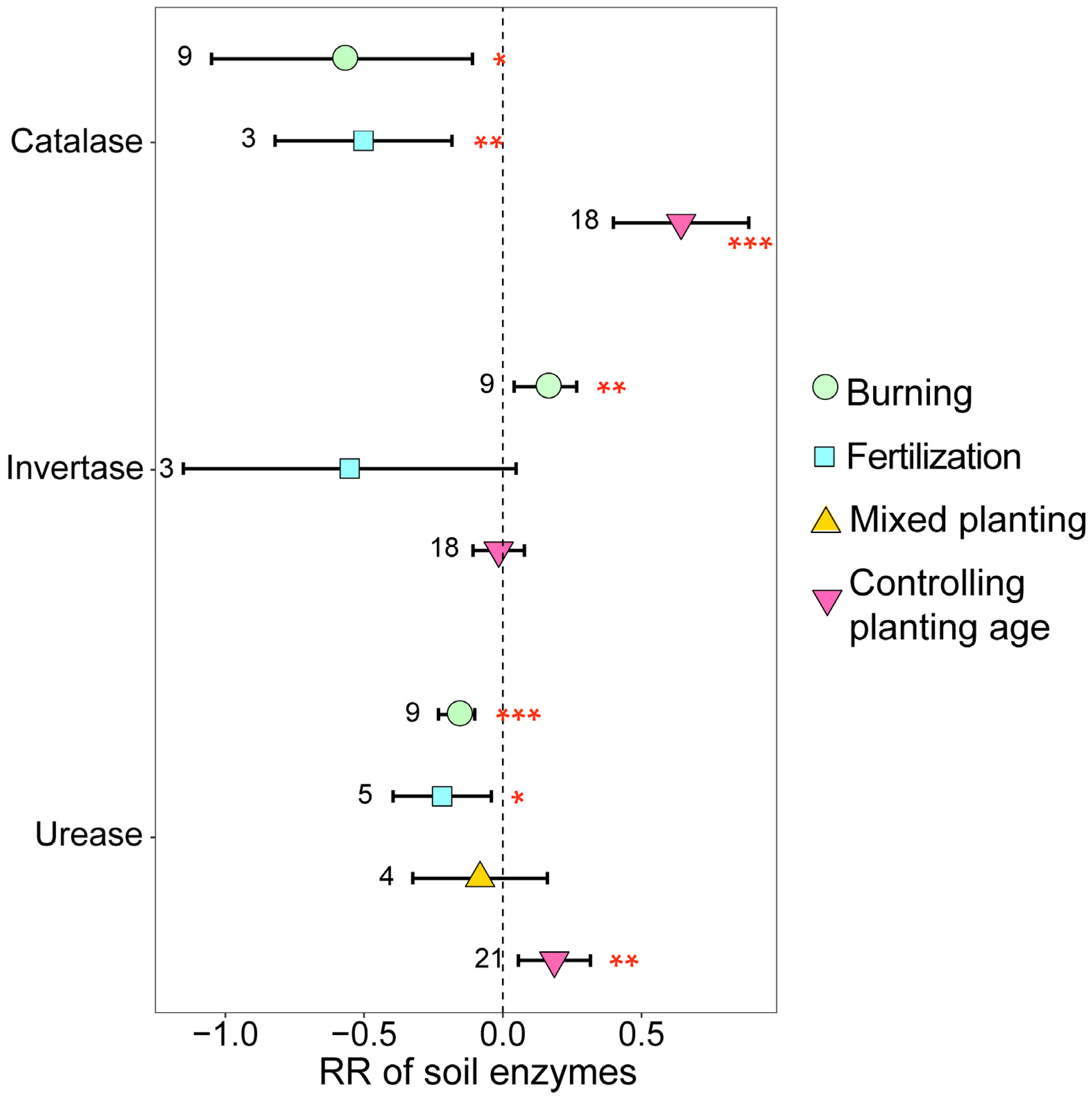
The different management practices also exerted a significant influence on soil enzyme activity on eucalyptus plantations (Figure 6). Burning and fertilization significantly decreased soil catalase and urease activities while concurrently significantly increasing soil invertase activity (p < 0.05). Conversely, with the increase in planting age, catalase and urease contents significantly increased (p < 0.05).
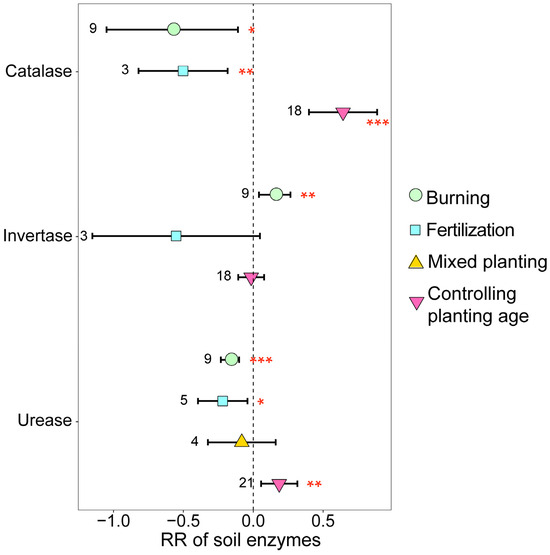
Figure 6.
The response ratios of soil enzymes under different management practices. The values beside the bars are the corresponding number of observations. ***, ** and * indicates p < 0.001, p < 0.01 and p < 0.05, respectively.
3.3. Mechanisms of Soil Microbial Biomass, Diversity, and Structural Changes
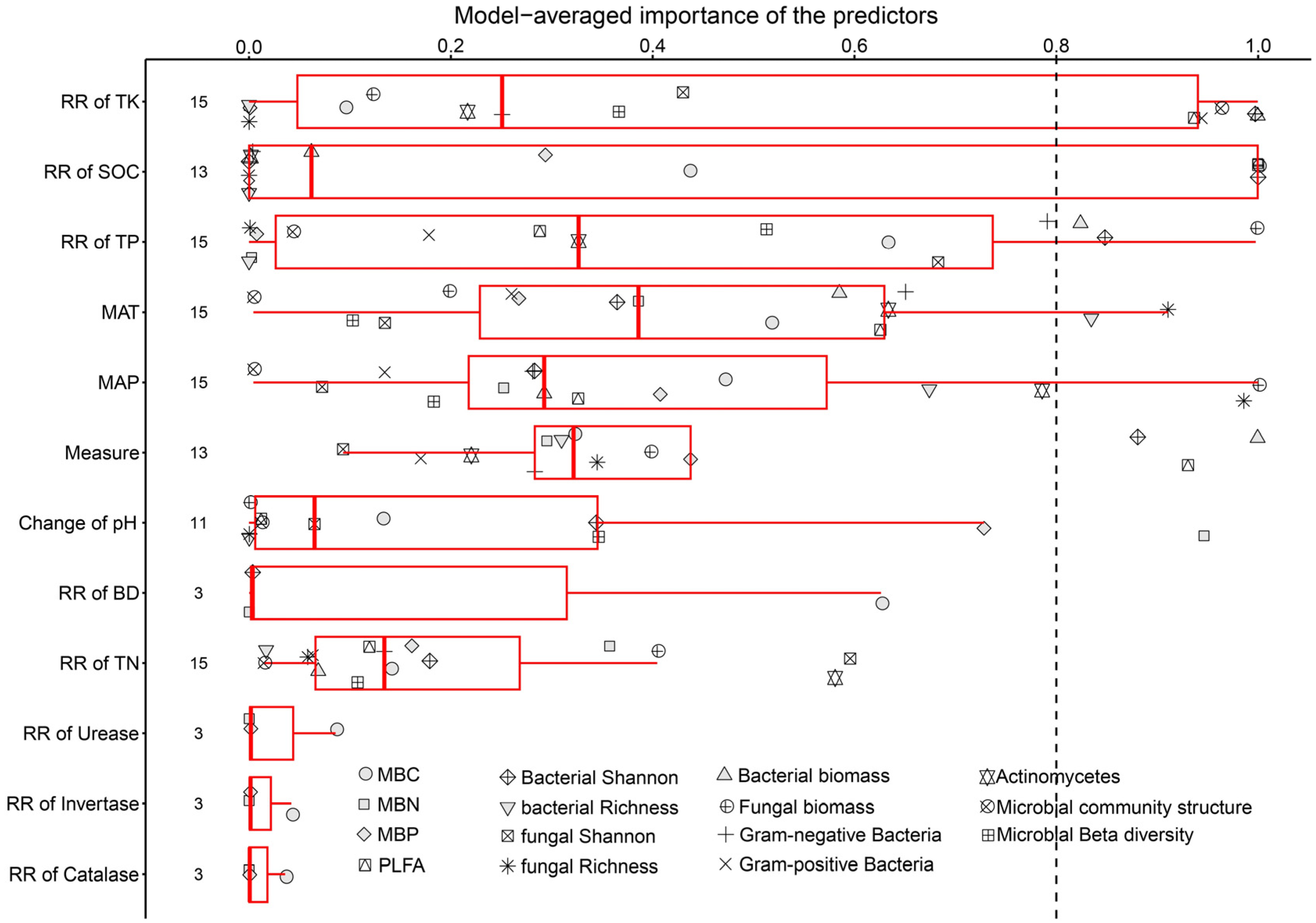
Through model selection analysis, we identified that SOC and TK are crucial factors for soil microbes. Moreover, certain soil microbes were also affected by TP, MAT, and MAP (Figure 7 and Figure 8).
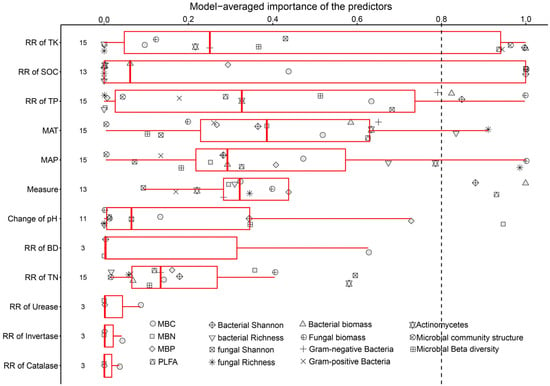
Figure 7.
Factors influencing the response of microbial biomass, alpha diversity, and community structure. BD: soil bulk density; TN: total nitrogen; TP: total phosphorus; TK: total potassium; SOC: soil organic carbon; MAT: mean annual temperature; MAP: mean annual precipitation.
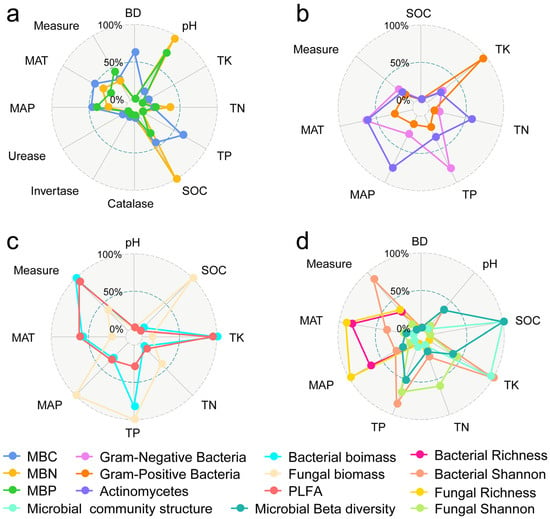
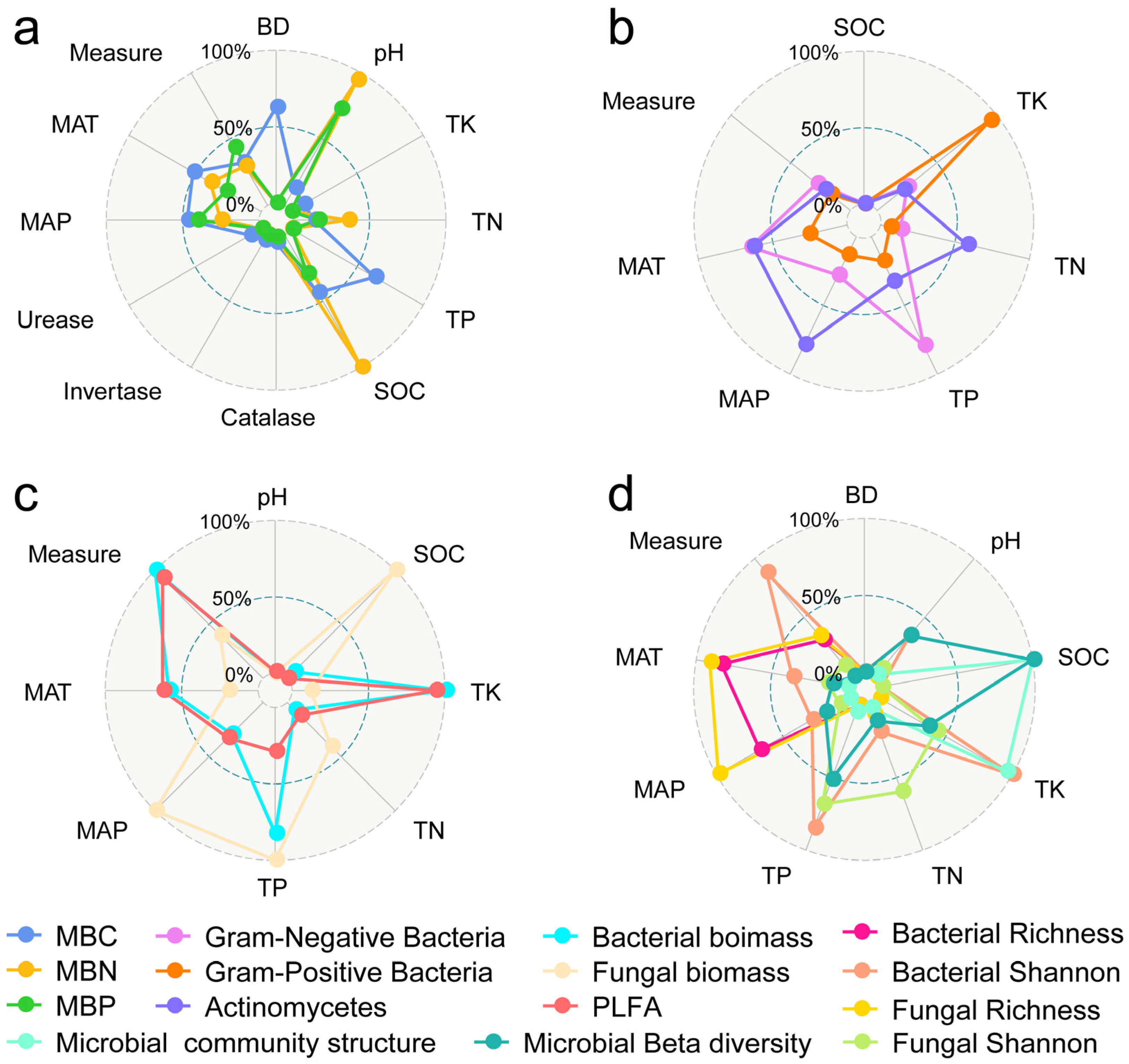
Figure 8.
Factors influencing the response of microbial biomass elements (a), microbial biomass of different species (b,c), alpha diversity, and community structure (d). BD: soil bulk density; TN: total nitrogen; TP: total phosphorus; TK: total potassium; SOC: soil organic carbon; MAT: mean annual temperature; MAP: mean annual precipitation.
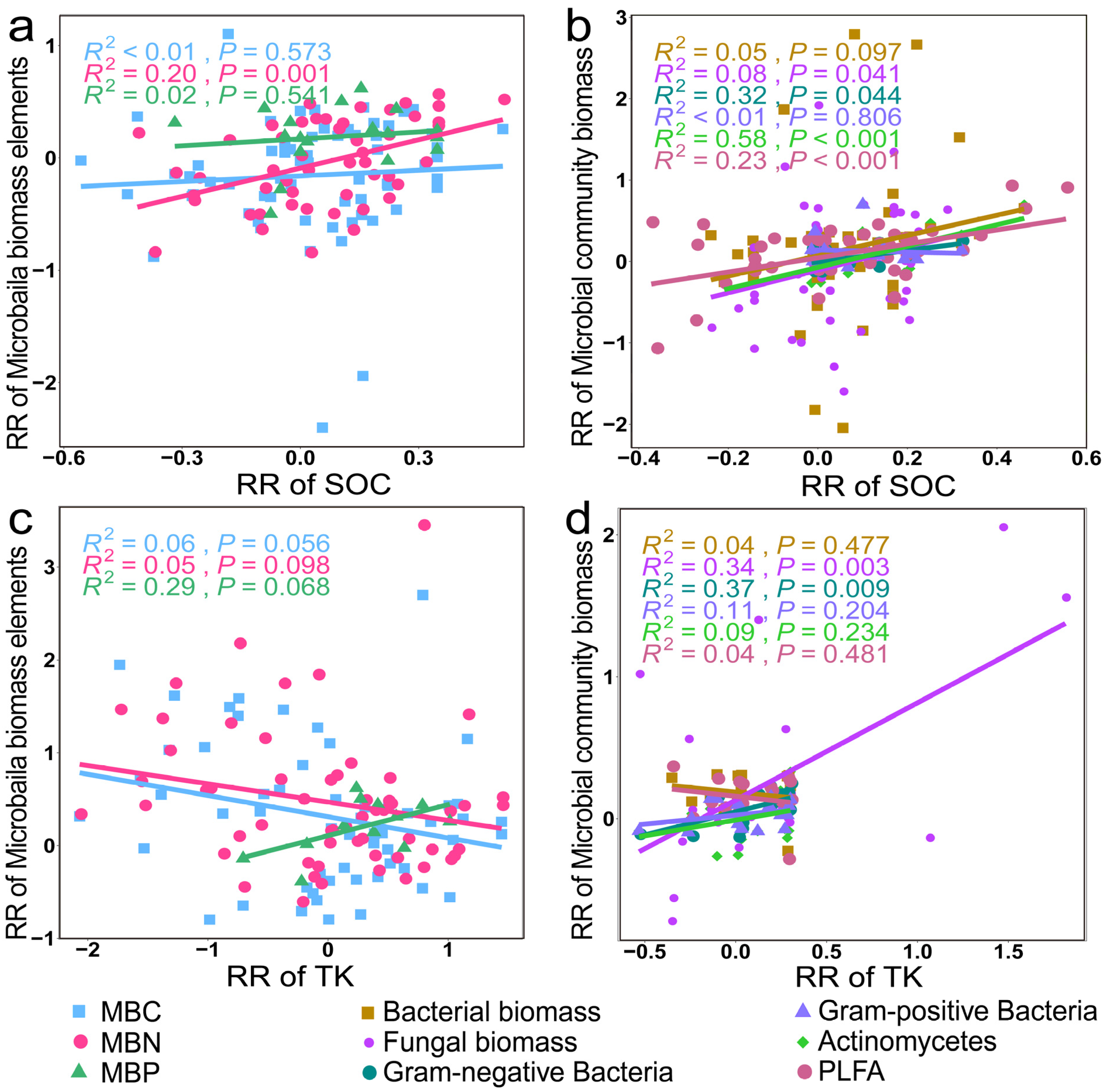
We conducted further regression analyses and observed a significant negative correlation between the RR of SOC and the RR of MBC (Figure 9a, p = 0.001). With an increase in the RR of TK and SOC, there was a concurrent significant increase in the RR of fungal and Gram-negative bacteria biomass (Figure 9b,d, p < 0.05). In addition, with an increase in the RR of SOC, the RR of Actinomycetes and the total microbial biomass exhibited concurrent significant increases (Figure 9b, p < 0.001).
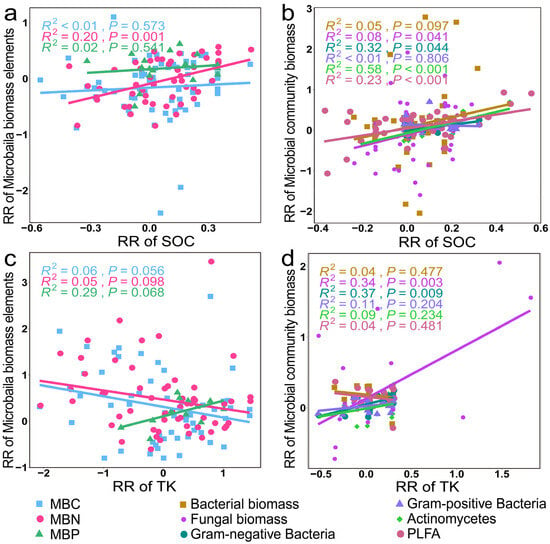
Figure 9.
Relationships between the RR of microbial biomass elements and the microbial biomass of different species with the RR of SOC (a,b) and TK (c,d).
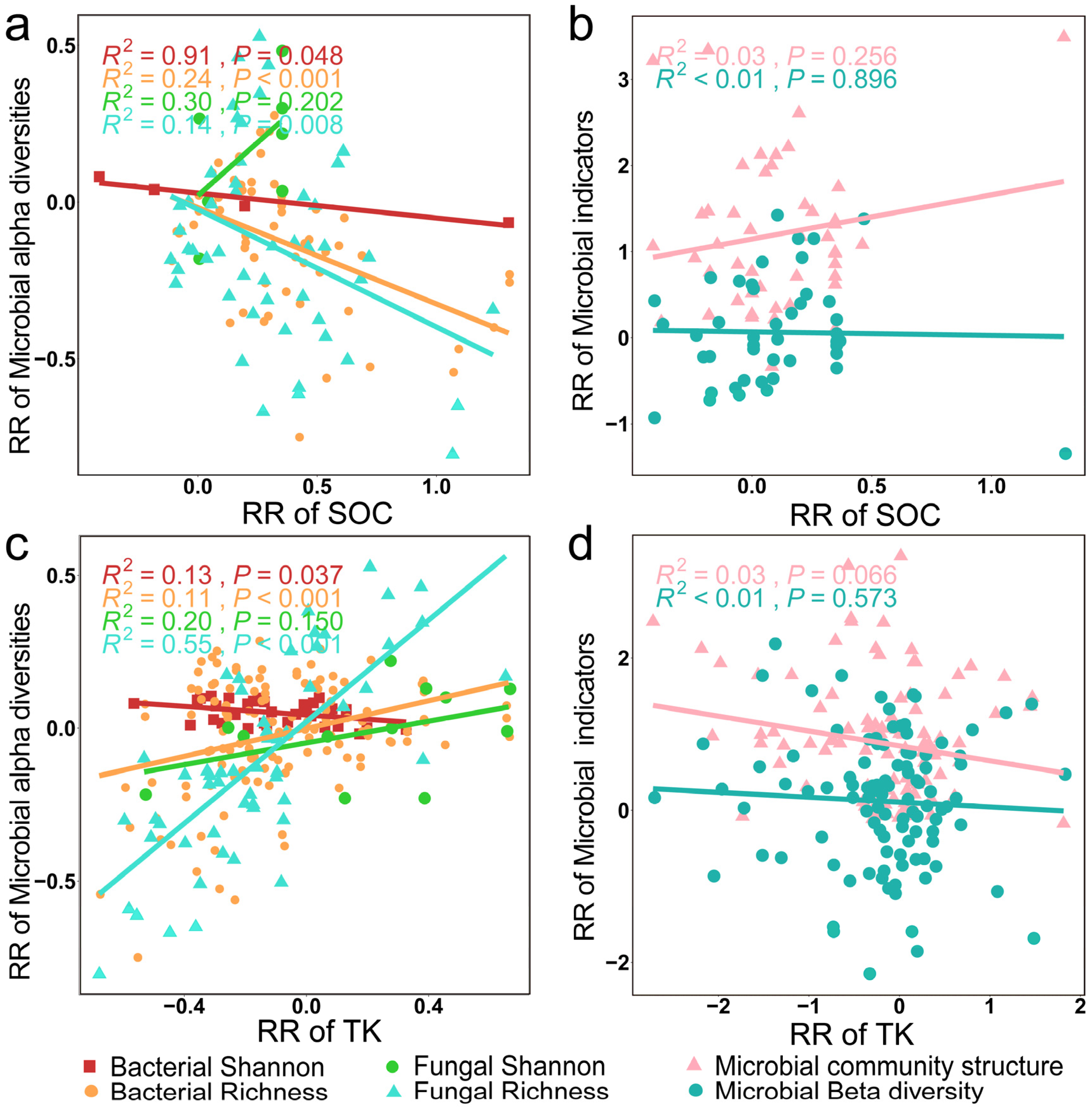
Soil microbial alpha diversity indices also exhibited significant correlations with SOC and TK (Figure 9a,c, p < 0.01). A discernible diminishing trend was observed in the RR of soil bacterial alpha diversity indices as the RR of SOC and TK increased (Figure 10a,c, p < 0.05). In particular, bacterial and fungal richness indices exhibited a negative correlation with SOC (Figure 10a, p < 0.01) while showing a concurrent positive correlation with TK (Figure 10c, p < 0.001).
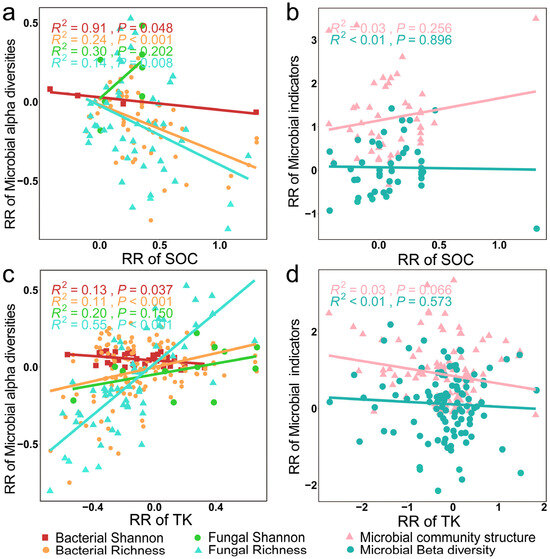
Figure 10.
Relationships between the RR of microbial alpha diversity and microbial community structures with the RR of SOC (a,b) and TK (c,d).
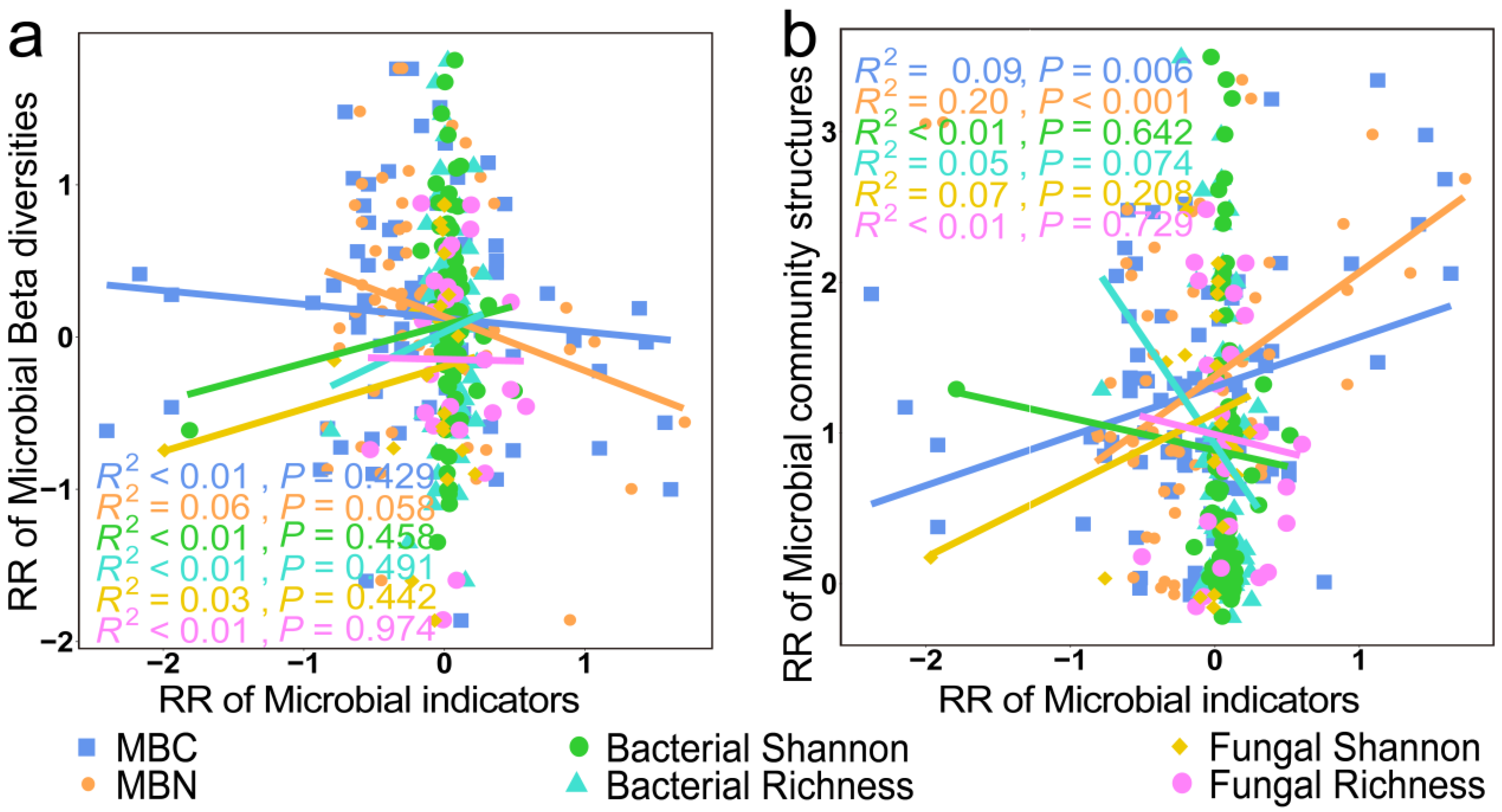
As shown in Figure 11, we found a significant negative correlation between the RR of microbial community structures and the RR of MBC and MBN (p < 0.01). However, there was no significant correlation between the RR of microbial community structure and the RR of alpha diversity (p > 0.05).
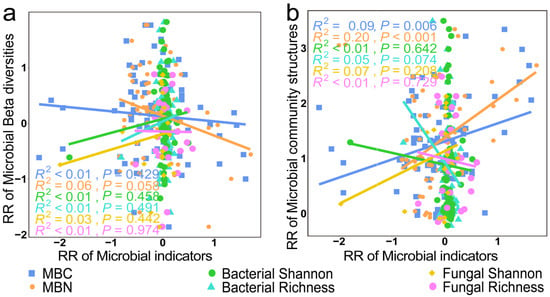
Figure 11.
Relationships between the RR of microbial beta diversity (a) and microbial community structure (b) with the RR of microbial biomass elements and microbial alpha diversity.
4. Discussion
4.1. Factors Affecting Microbial Community Biomass, Diversity, and Structure
Both burning and residue removal had negative impacts on the soil MBC and MBN, particularly inhibiting the growth of fungal communities. The reduction in soil MBC and MBN was attributed to the decrease in SOC. The decrease in SOC after burning may be due to the reduction in peroxidase and urease enzyme contents as these enzymes play a vital role in soil carbon and nitrogen release, respectively [22]. The results of a previous study demonstrated that peroxidase and urease enzyme contents exhibited a significantly negative correlation with soil pH [62]. These findings indicate that, with the increase in soil pH, induced by the leaching of alkaline salts from the ash and charcoal in addition to the denaturation of organic acids, the peroxidase and urease enzyme contents will decrease, ultimately leading to a decline in SOC [22,63]. In contrast, the decrease in SOC following residue removal may be due to the direct loss of nutrients leached from the litter [15].
In addition, both burning and residue removal led to a reduction in fungal biomass; an important difference, however, was the fact that burning resulted in an increase in bacterial biomass. This discrepancy was attributed to bacteria showing a preference for utilizing readily available carbon substrates, whereas fungi displayed a tendency to degrade more complex carbon substrates [64]. Intense burning is often accompanied by deterioration in intricate carbon substrates such as cellulose and lignin [63], consequently augmenting the accessibility of simpler carbon substrates. From the above results, it can be concluded that the practice of burning promotes bacterial growth rather than fungal growth. Moreover, burning led to an increase in soil pH, and under elevated pH conditions, bacterial growth surpassed fungal growth [65,66]. Furthermore, burning significantly reduced the diversity and richness of fungal communities but had no significant effect on bacterial communities. This difference in response may be attributed to the greater fire tolerance of bacteria compared to fungi [22,67,68]. Soil temperatures above 70 °C are lethal to fungi [69]; in comparison, temperatures exceeding 200 °C may only eliminate certain bacterial species [70]. However, after residue removal, fungal biomass exhibited a significant decrease, while bacterial biomass remained relatively unchanged. This phenomenon may have arisen from the exposure of soil to air after the removal of surface residues, thereby transforming the anaerobic environment into one that is aerobic, which was unfavorable for fungal growth [18]. Consequently, this resulted in a substantial decrease in fungal biomass [71]. Overall, the complete removal of all residues and the practice of burning are detrimental to soil microbial growth, with burning negatively impacting the preservation of fungal community diversity.
Fertilization, mixing eucalyptus with nitrogen-fixing species, and controlling planting age primarily influenced soil health by impacting soil TC and TN contents. However, these management practices enhanced soil nutrient abundance through different pathways. Introducing nitrogen-fixing species alongside eucalyptus can stimulate the proliferation of nitrogen-fixing bacteria, which symbiotically fix atmospheric nitrogen into the soil through root symbiosis [72], thereby substantially elevating the TN content in eucalyptus plantation soil [17,19,32]. With the increasing age of plants, there was a gradual increase in the richness of understory plants and root yield, accompanied by accelerated litter decomposition rates. These processes contributed to enhanced soil aggregation and permeability [21,42,73], consequently leading to significant increases in TC, TN, and SOC on eucalyptus plantations [30,43]. Fertilization directly increases soil nutrient contents through the addition of nutrients to the soil.
The soil microbial biomass, diversity, and community structure exhibit contrasting responses to different management practices due to the varying methods of nutrient addition. For example, the biomass of Gram-negative bacteria significantly increased following fertilization, whereas the biomass of Gram-positive bacteria remained unchanged. This discrepancy may be attributed to the rapid response of Gram-negative bacteria to readily available carbon from fresh litter inputs; in comparison, Gram-positive bacteria must adapt to utilizing more complex carbon sources [74]. Mixing eucalyptus with nitrogen-fixing species led to a significant increase in MBN and total microbial biomass. This result was likely due to mixed planting increasing litter diversity, which provided sufficient carbon and nitrogen sources for microbial growth [75,76], in addition to creating a more diversified overall environment [77]. Moreover, we found that soil bacterial biomass significantly increased, whereas fungal biomass remained relatively stable. This discrepancy may have arisen from the greater changes in soil TN compared to TC, resulting in a decrease in soil C:N, which was more favorable for bacterial growth than fungal growth [20]. With the increase in planting age, the augmentation of the soil nutrient content and the secretion of allelochemicals notably enhanced soil microorganism activity [78,79], consequently resulting in a significant increase in soil MBC, MBN, and MBP. Actinomycetes, widely distributed in soil, play a crucial role in promoting the growth and development of eucalyptus [16,80,81]. In soils with older plantation ages, the biomass of Actinomycetes tended to be higher, potentially due to the increase in SOC with increasing plantation age, which in turn led to an increase in Actinomycetes biomass (Figure 9b). With the increase in planting age, there was a significant decrease in the richness of both fungal and bacterial communities. This finding could be attributed to the proliferation of environmental opportunists, such as organic decomposers, in response to abundant carbon resources. These opportunists may potentially suppress the reproduction of other species, leading to the disappearance of certain species and a reduction in richness [82]. Our findings also indicated a significant decrease in both bacterial and fungal community richness with increasing carbon resources (Figure 10a). Changes in bacterial and fungal community richness were also related to soil TK, exhibiting a significant decrease in richness as TK decreased (Figure 10c). In addition, ventilation also constrained bacterial community diversity [21]. As planting age increased, intensified canopy shading and litter accumulation created an anoxic environment, thereby diminishing bacterial community richness [18]. Overall, burning negatively affected fungal growth, diversity, MBC, and MBN while promoting bacterial community growth; residue removal hindered fungal growth and MBC and MBN storage; mixing eucalyptus with nitrogen-fixing species favored bacterial community growth and MBN storage; fertilization reduced fungal diversity but promoted bacterial diversity; and controlling stand age negatively affected both bacterial and fungal diversity while concurrently benefiting Actinomycetes growth and MBC, MBN, and MBP storage. From the above results, it can be concluded that mixing eucalyptus with nitrogen-fixing species is the most beneficial management practice; however, attention should be paid to properly protecting fungal communities, with residue removal being the least advantageous management practice.
The alpha diversity of fungal communities was more sensitive to the different management practices than that of bacterial communities, a finding that may be related to the different habits and physiological structures of fungi and bacteria. Fungi typically possess relatively complex cellular structures, including nuclei and cell walls, whereas bacteria typically possess less complex cellular structures and frequently establish biofilms in the soil environment. From the above findings, it is evident that compared to bacteria, fungi are less adaptable to environmental pressures [83]. Additionally, bacteria play a broader role in ecosystem biochemical cycles [84], whereas fungi are primarily involved in organic matter decomposition [85]; such factors result in bacteria being more adaptable to environmental changes compared to fungi [86].
Different management practices exert a significant impact on the structures of microbial communities, although it is important to note that there may not always be a direct correlation between microbial community structures and alpha diversity (Figure 11). Furthermore, microbial community structures were more sensitive to management practices than microbial alpha diversity (Figure 4). The reason for this finding is the fact that while certain species may have decreased in number or disappeared in response to the different management practices, another set of species may have increased in number or appeared, thereby altering the microbial community composition without necessarily affecting its diversity. However, significant changes in the internal composition of microbes may occur.
4.2. Limitations and Future Directions
While we were able to gather a substantial number of foundational data in the present study, few authors of existing studies have comprehensively addressed all of the indicators collected by our group. For the current dataset, constructing an accurate mechanistic framework to explore the changes in soil microbial communities on eucalyptus plantations following the implementation of various management practices remains a challenge. To address this limitation, we hope that the authors of future studies can expand our dataset. In addition, we hope that the authors of future studies can incorporate a broader range of environmental variables to more precisely evaluate the response of soil microbial communities to different management practices. Meanwhile, our study has limited focus on the influence of eucalyptus species on management practices. Although we selected common management practices applicable to most eucalyptus plantations and aimed to minimize the impact of eucalyptus species on these practices, we still hope to collect additional data in the future to address this drawback in our research. Moreover, we hope that future studies can include data on microbial functions and taxonomy, enabling a deeper investigation into how different management strategies influence the function of soil microbial communities. Such factors represent a critical component for accurately predicting and managing soil ecosystem functions and should not be overlooked.
5. Conclusions
On eucalyptus plantations, fungal community alpha diversity responds more sensitively to different management practices than bacterial community alpha diversity. Although the implementation of management practices significantly increased most soil nutrient contents and microbial biomass elements (except for burning), these practices did not necessarily improve soil microbial biomass and diversity, particularly among fungal communities. Specifically, burning negatively affected fungal growth, diversity, and microbial biomass carbon and nitrogen contents while simultaneously promoting bacterial community growth; residue removal hindered fungal growth and microbial biomass carbon and nitrogen storage; mixing eucalyptus with nitrogen-fixing species favored bacterial community growth and microbial biomass nitrogen storage; fertilization reduced fungal diversity but promoted bacterial diversity; and controlling stand age negatively affected both bacterial and fungal diversity while benefiting Actinomycetes growth and microbial biomass carbon, nitrogen, and phosphorus storage. Thus, mixing eucalyptus with nitrogen-fixing species is the most beneficial management practice; however, care should be taken to protect fungal communities, while residue removal is the least advantageous. Furthermore, our study indicated that variations of soil microbial diversity were primarily influenced by soil organic carbon and total potassium contents, suggesting that these should be prioritized when implementing management practices. Additionally, the implementation of management practices also significantly altered microbial community structures by positively affecting microbial biomass carbon and microbial biomass nitrogen instead of soil microbial alpha diversity. However, the impact of different practices on microbial community structure differed, burning mainly affected rare species structures, whereas mixed planting and increasing planting age mainly effected an abundant species structure. Therefore, different management practices require attention to different microbial community structures.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/land14040692/s1, Supplementary S1 is the source of the data, Table S1 is the results of the practice on each variable effect, and Table S2 is the results of potential publication bias analyses.
Author Contributions
Conceptualization: Y.X., W.L. and M.Z.; methodology: Y.X., W.L. and M.Z.; resources: J.Z., M.Z. and L.W.; software: L.W., Z.C. and X.L.; writing—original draft preparation, Y.X.; writing—review and editing, Y.X., W.L. and M.Z.; funding acquisition, W.L. and Q.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been funded by the National Key R&D Program of China (Nos. 2022YFF1303301 and 2022YFF1302603), the National Natural Science Fund of China (Nos. U24A20178 and 42201133), Gansu Provincial Science and Technology Planning Project (Nos. 23ZDFA018, 23ZDKA017 and 22JR5RA072), Gansu Province Intellectual Property Plan Program (No. 23ZSCQD001), Excellent Doctoral Student Program of Gansu Province (No. 24JRRA110), and the National Grass Industry Technology Innovation Center (preparation) Key Innovation Platform Construction Project (CCPTZX2024GJ04).
Data Availability Statement
Data will be made available on request.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Laclau, J.P.; Ranger, J.; Boullet, J.P.; Nzila, J.D.; Deleporte, P. Nutrient cycling in a clonal stand of Eucalyptus and an adjacent savanna ecosystem in Congo. 1. Chemical composition of rainfall and stem flow solutions. For. Ecol. Manag. 2003, 176, 105–119. [Google Scholar]
- Laclau, J.P.; Ranger, J.; Boullet, J.P.; Nzila, J.D.; Deleporte, P. Nutrient cycling in a clonal stand of Eucalyptus and an adjacent savanna ecosystem in Congo. 2. Chemical composition of soil solutions. For. Ecol. Manag. 2003, 180, 527–544. [Google Scholar]
- Pate, J.S.; Verboom, W.H. Contemporary biogenic formation of clay pavement by eucalypts: Further support for the phytotarium concept. Ann. Bot. 2009, 103, 673–685. [Google Scholar]
- Phalan, B.; Bertzky, M.; Butchart, S.H.M.; Donald, P.F.; Scharlemann, J.P.W.; Stattersfield, A.J.; Balmfordet, A. Crop expansion and conservation priorities in tropical countries. PLoS ONE 2013, 8, e51759. [Google Scholar]
- Eufrade Junior, H.J.; Melo, R.X.; Sartori, M.M.P.; Guerra, S.P.S.; Ballarin, A.W. Sustainable use of eucalypt biomass grown on short rotation coppice for bioenergy. Biomass Bioenerg. 2016, 90, 15–21. [Google Scholar]
- Jagger, P.; Pender, J. The role of trees for sustainable management of less-favoured lands: The case of eucalyptus in Ethiopia. For. Policy Econ. 2003, 5, 83–95. [Google Scholar]
- Nouvellon, Y.; Epron, D.; Marsden, C.; Kinana, A.; Maire, G.L.; Deleporte, P.; SaintAndre, L.; Bouillet, J.P.; Laclau, J.P. Age-related changes in litter inputs explain annual trends in soil CO2 effluxes over a full Eucalyptus rotation after afforestation of a tropical savannah. Biogeochemistry 2012, 111, 515–533. [Google Scholar]
- Zhang, Y.X.; Wang, X.J. Geographical spatial distribution and productivity dynamic change of Eucalyptus plantations in China. Sci. Rep. 2021, 11, 19764. [Google Scholar]
- Laclau, J.P.; Deleporte, P.; Ranger, J.; Bouilleti, J.P.; Kazotti, G. Nutrient dynamics throughout the rotation of Eucalyptus clonal stands in Congo. Ann. Bot. 2003, 91, 879–892. [Google Scholar]
- Li, H.F.; Fu, S.L.; Zhao, H.T.; Xia, H.P. Effects of understory removal and N-fixing species seeding on soil N2O fluxes in four forest plantations in southern China. Soil Sci. Plant Nut. 2010, 56, 541–551. [Google Scholar]
- Florentine, S.K.; Fox, J.E.D. Allelopathic effects of Eucalyptus victrix L. on Eucalyptus species and grasses. Allelopath 2003, 11, 77–83. [Google Scholar]
- Zhang, C.L.; Fu, S.L. Allelopathic effects of eucalyptus and the establishment of mixed stands of eucalyptus and native species. For. Ecol. Manag. 2009, 258, 1391–1396. [Google Scholar]
- Kara, O.; Bolat, I.; Çakıroğlu, K.; Öztürk, M. Plant canopy effects on litter accumulation and soil microbial biomass in two temperate forests. Biol. Fertil. Soils 2008, 45, 193–198. [Google Scholar]
- Xue, Y.Y. Research on the Characteristics and Influencing Factors of Soil Microbial Communities in Forest and Cultivated Land; Northwest Institute of Eco-Environment and Resource, Chinese Academy of Science: Lanzhou, China, 2022. [Google Scholar]
- Wang, Y.Z.; Zheng, J.Q.; Xu, Z.H.; Abdullah, K.M.; Zhou, Q.X. Effects of changed litter inputs on soil labile carbon and nitrogen pools in a eucalyptus-dominated forest of southeast Queensland, Australia. J. Soil Sediment. 2019, 19, 1661–1671. [Google Scholar]
- Mercedes, M.O.; Michael, B.; Richard, J.P.D.; Mark, K.J.O.; Miriam, M.R. Fire and land use impact soil properties in a Mediterranean dry sclerophyll woodland. J. Environ. Manag. 2020, 324, 116245. [Google Scholar]
- Jiang, Y.S.; Luo, C.L.; Zhang, D.Y.; Ostle, N.J.; Cheng, Z.N.; Ding, P.; Shen, C.D.; Zhang, G. Radiocarbon evidence of the impact of forest-to-plantation conversion on soil organic carbon dynamics on a tropical island. Geoderma 2020, 375, 114484. [Google Scholar]
- Qu, Z.L.; Liu, B.; Ma, Y.; Sun, H. Differences in bacterial community structure and potential functions among Eucalyptus plantations with different ages and species of trees. Appl. Soil Ecol. 2020, 149, 103515. [Google Scholar]
- Pereira, A.P.A.; Santana, M.C.; Zagatto, M.R.G.; Brandani, C.B.; Wang, J.T.; Verma, J.P.; Singh, B.K.; Cardoso, E.J.B.N. Nitrogen-fixing trees in mixed forest systems regulate the ecology of fungal community and phosphorus cycling. Sci. Total Environ. 2021, 758, 143711. [Google Scholar]
- Huang, X.M.; Liu, S.R.; Wang, H.; Hu, Z.D.; Li, Z.G.; You, Y.M. Changes of soil microbial biomass carbon and community composition through mixing nitrogen-fixing species with Eucalyptus urophylla in subtropical China. Soil Biol. Biochem. 2014, 73, 42–48. [Google Scholar]
- Xu, Y.X.; Du, A.; Wang, Z.C.; Zhu, W.K.; Li, C.; Wu, L.C. Effects of different rotation periods of Eucalyptus plantations on soil physiochemical properties, enzyme activities, microbial biomass and microbial community structure and diversity. For. Ecol. Manag. 2020, 456, 117683. [Google Scholar]
- Liu, X.; Chen, C.R.; Wang, W.J.; Hughes, J.M.; Lewis, T.; Hou, E.Q.; Shen, J.P. Soil environmental factors rather than denitrification gene abundance control N2O fluxes in a wet sclerophyll forest with different burning frequency. Soil Biol. Biochem. 2013, 57, 292–300. [Google Scholar] [CrossRef]
- Gunina, A.; Smith, A.R.; Godbold, D.L.; Jones, D.L.; Kuzyakov, Y. Response of soil microbial community to afforestation with pure and mixed species. Plant Soil 2017, 412, 357–368. [Google Scholar] [CrossRef]
- Wang, D.J.; Abdullaha, K.M.; Xu, Z.H.; Wang, W.J. Water extractable organic C and total N: The most sensitive indicator of soil labile C and N pools in response to the prescribed burning in a suburban natural forest of subtropical Australia. Geoderma 2020, 377, 114586. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Wang, J.C.; Weng, Y.L.; Chen, X.L.; Wu, L.C. Soil characteristics of Eucalyptus urophylla×Eucalyptus grandis plantations under different management practices for harvest residues with soil depth gradient across time. Ecol. Indic. 2020, 117, 106530. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.W.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Palmer, H.D.; Denham, A.J.; Ooi, M.K.J. Fire severity drives variation in post-fire recruitment and residual seed bank size of Acacia species. Plant Ecol. 2018, 219, 527–537. [Google Scholar] [CrossRef]
- Wang, C.; Wang, G.; Wang, Y.; Rafique, R.; Ma, L.; Hu, L.; Luo, Y. Fire alters vegetation and soil microbial community in alpine meadow. Land Degrad. Dev. 2015, 27, 1379–1390. [Google Scholar] [CrossRef]
- Bárcenas-Moreno, G.; García-Orenes, F.; Mataix-Solera, J.; Mataix-Beneyto, J. Plant community influence on soil microbial response after a wildfire in Sierra Nevada National Park (Spain). Sci. Total Environ. 2016, 573, 1265–1274. [Google Scholar] [CrossRef]
- Mendham, D.S.; Sankaran, K.V.; OConnell, A.M.; Grove, T.S. Eucalyptus globulus harvest residue management effects on soil carbon and microbial biomass at 1 and 5 years after plantation establishment. Soil Biol. Biochem. 2002, 34, 1903–1912. [Google Scholar] [CrossRef]
- Rachid, C.T.C.C.; Balieiro, F.C.; Peixoto, R.S.; Pinheiro, Y.A.S.; Piccolo, M.C.; Chaer, G.M.; Rosado, A.S. Mixed plantations can promote microbial integration and soil nitrate increases with changes in the N cycling genes. Soil Biol. Biochem. 2013, 66, 146–153. [Google Scholar] [CrossRef]
- Pereira, A.P.A.; Durrer, A.; Gumierec, T.; Gonçalves, J.L.M.; Robina, A.; Bouillet, J.P.; Wang, J.T.; Vermab, J.P.; Singh, B.K.; Cardoso, E.J.B.N. Mixed Eucalyptus plantations induce changes in microbial communities and increase biological functions in the soil and litter layers. For. Ecol. Manag. 2019, 433, 332–342. [Google Scholar]
- Xu, Y.X.; Ren, S.Q.; Liang, Y.F.; Du, A.; Li, C.; Wang, Z.C.; Zhu, W.K.; Wu, L.C. Soil nutrient supply and tree species drive changes in soil microbial communities during the transformation of a multi-generation Eucalyptus plantation. Appl. Soil Ecol. 2021, 166, 103991. [Google Scholar]
- Bini, D.; dos Santos, C.A.; Bouillet, J.P.; Goncalves, J.L.D.M.; Cardoso, E.J.B.N. Eucalyptus grandis and Acacia mangium in monoculture and intercropped plantations: Evolution of soil and litter microbial and chemical attributes during early stages of plant development. Appl. Soil Ecol. 2013, 63, 57–66. [Google Scholar] [CrossRef]
- Qian, Z.Z.; Zhu, K.X.; Zhuang, S.Y.; Tang, L.Z. Soil nutrient cycling and bacterial community structure in response to various green manures in a successive Eucalyptus (Eucalyptus urophylla × Eucalyptus grandis) plantation. Land Degrad. Dev. 2022, 33, 2809–2821. [Google Scholar]
- Wang, Q.K.; Wang, S.L.; Liu, Y.X. Responses to N and P fertilization in a young Eucalyptus dunnii plantation: Microbial properties, enzyme activities and dissolved organic matter. Appl. Soil Ecol. 2008, 40, 484–490. [Google Scholar]
- Kjoller, R.; Nilsson, L.O.; Hansen, K.; Schmidt, I.K.; Vesterdal, L.; Gundersen, P. Dramatic changes in ectomycorrhizal community composition, root tip abundance and mycelial production along a stand-scale nitrogen deposition gradient. New Phytol. 2012, 194, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Hasselquist, N.J.; Hgberg, P. Dosage and duration effects of nitrogen additions on ectomycorrhizal sporocarp production and functioning: An example from two N-limited boreal forests. Ecol. Evol. 2014, 4, 3015–3026. [Google Scholar] [CrossRef]
- Asante, P.; Armstrong, G.W.; Adamowicz, W.L. Carbon sequestration and the optimal forest harvest decision: A dynamic programming approach considering biomass and dead organic matter. J. For. Econ. 2011, 17, 3–17. [Google Scholar]
- Cavagnaroa, T.R.; Cunningham, S.C.; Fitzpatrick, S. Pastures to woodlands: Changes in soil microbial communities and carbon following reforestation. Appl. Soil Ecol. 2016, 107, 24–32. [Google Scholar] [CrossRef]
- Heyde, M.V.D.; Bunce, M.; Dixon, K.; Wardell-Johnson, G.; White, N.E.; Nevill, P. Changes in soil microbial communities in post mine ecological restoration: Implications for monitoring using high throughput DNA sequencing. Sci. Total Environ. 2020, 749, 142262. [Google Scholar]
- Trogisch, S.; He, J.S.; Hector, A.; Scherer-Lorenzen, M. Impact of species diversity, stand age and environmental factors on leaf litter decomposition in subtropical forests in China. Plant Soil 2015, 400, 337–350. [Google Scholar]
- Sun, H.; Wang, Q.; Liu, N.; Li, L.; Zhang, C.; Liu, Z.; Zhang, Y. Effects of different leaf litters on the physicochemical properties and bacterial communities in Panax ginseng-growing soil. Appl. Soil Ecol. 2017, 111, 17–24. [Google Scholar]
- Wang, J.; Yang, S.; Ma, T.F.; Raza, W.; Jing, L.; Howland, J.G.; Huang, Q.W.; Shen, Q.R. Impacts of inorganic and organic fertilization treatments on bacterial and fungal communities in a paddy soil. Appl. Soil Ecol. 2017, 112, 42–50. [Google Scholar]
- Wang, S.; Li, T.; Zheng, Z. Distribution of microbial biomass and activity within soil aggregates as affected by tea plantation age. CATENA 2017, 153, 1–8. [Google Scholar]
- Liu, D.; Huang, Y.; Sun, H.; An, S. The restoration age of Robinia pseudoacacia plantation impacts soil microbial biomass and microbial community structure in the Loess Plateau. CATENA 2018, 165, 192–200. [Google Scholar]
- Zhu, L.; Wang, X.; Chen, F.; Li, C.; Wu, L. Effects of the successive planting of Eucalyptus urophylla on the soil bacterial and fungal community structure, diversity, microbial biomass, and enzyme activity. Land Degrad. Dev. 2019, 30, 636–646. [Google Scholar]
- Guo, J.H.; Feng, H.L.; McNie, P.; Liu, Q.Y.; Xu, X.; Pan, C.; Yan, K.; Feng, L.; Goitom, E.A.; Yu, Y.C. Species mixing improves soil properties and enzymatic activities in Chinese fir plantations: A meta-analysis. CATENA 2023, 220, 106723. [Google Scholar]
- Curtright, A.J.; Tiemann, L.K. Intercropping increases soil extracellular enzyme activity: A meta-analysis. Agric. Ecosyst. Environ. 2021, 319, 107489. [Google Scholar]
- Zhou, Z.H.; Wang, C.K.; Zheng, M.H.; Jiang, L.F.; Luo, Y.Q. Patterns and mechanisms of responses by soil microbial communities to nitrogen addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar]
- Waring, B.; Gee, A.; Liang, G.P.; Adkins, S. A quantitative analysis of microbial community structure-function relationships in plant litter decay. iScience 2022, 25, 104523. [Google Scholar]
- Achat, D.L.; Deleuze, C.; Landmann, G.; Pousse, N.; Ranger, J.; Augusto, L. Quantifying consequences of removing harvesting residues on forest soils and tree growth—A meta-analysis. For. Ecol. Manag. 2015, 348, 124–141. [Google Scholar]
- Meng, C.; Tian, D.S.; Zeng, H.; Li, Z.L.; Yi, C.X.; Niu, S.L. Global soil acidification impacts on belowground processes. Environ. Res. Lett. 2019, 14, 074003. [Google Scholar]
- Delgado-Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [PubMed]
- Paliy, O.; Shankar, V. Application of multivariate statistical techniques in microbial ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar]
- Zhou, Z.H.; Wang, C.K.; Luo, Y.Q. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 3072. [Google Scholar] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; OHara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community ecology package. R. Package Version 2019, 2, 5–6. [Google Scholar]
- Yuan, C.P.; Gao, B.L.; Peng, Y.T.; Gao, X.; Fan, B.B.; Chen, Q. A meta-analysis of heavy metal bioavailability response to biochar aging: Importance of soil and biochar properties. Sci. Total Environ. 2021, 756, 144058. [Google Scholar]
- Wang, J.; Ding, C.; Heino, J.; Jiang, X.; Tao, J.; Ding, L.; Su, W.; Huang, M.; He, D. What explains the variation in dam impacts on riverine macroinvertebrates? A global quantitative synthesis. Environ. Res. Lett. 2020, 15, 124028. [Google Scholar]
- Viechtbauer, W. Conducting meta-analyses in R with the metafor package. J. Stati. Softw. 2010, 36, 1–48. [Google Scholar]
- Calcagno, V.; de Mazancourt, C. glmulti: An R package for easy automated model selection with (generalized) linear models. J. Stat. Softw. 2010, 34, 1–29. [Google Scholar]
- Luo, Y.J. Study on Soil Microorganism and Soil Enzyme Activity Along a Chronosequence of Eucalyptus plantation. Master’s Thesis, Guangxi Normal University, Guilin, China, 2014. [Google Scholar]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [PubMed]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar]
- Chen, L.; Jiang, Y.; Liang, C.; Luo, Y.; Xu, Q.; Han, C.; Zhao, Q.; Sun, B. Competitive interaction with keystone taxa induced negative priming under biochar amendments. Microbiome 2019, 7, 77. [Google Scholar]
- Xue, Y.Y.; Liu, W.; Feng, Q.; Zhu, M.; Wang, L.G.; Chen, Z.X.; Zhang, J.T. Variations and controls of soil microbial necromass carbon in grasslands along aridity gradients. Ecol. Indic. 2023, 156, 111188. [Google Scholar]
- González-pérez, J.A.; González-Vila, F.J.; Almendros, G.; Knicker, H. The effect of fire on soil organic matter: A review. Environ. Int. 2004, 30, 855–870. [Google Scholar] [PubMed]
- Hart, S.C.; DeLuca, T.H.; Newman, G.S.; MacKenzie, M.D.; Boyle, S.I. Post-fire vegetative dynamics as drives of microbial community structure and function in forest soil. For. Ecol. Manag. 2005, 220, 166–184. [Google Scholar]
- Raison, R.J. Modification of the soil environment by vegetation fires, with particular reference to nitrogen transformations: A review. Plant Soil 1979, 51, 73–108. [Google Scholar]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on belowground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar]
- Chen, X.; Hu, Y.; Xia, Y.; Zheng, S.; Ma, C.; Rui, Y.; He, H.; Huang, D.; Zhang, Z.; Ge, T.; et al. Contrasting pathways of carbon sequestration in paddy and upland soils. Glob. Change Biol. 2021, 27, 2478–2490. [Google Scholar]
- Paula, R.R.; Bouillet, J.P.; Gonçalves, J.L.M.; Trivelin, P.C.O.; Balieiro, F.C.; Nouvellon, Y.; Oliveira, J.C.; Júnior, J.C.D.; Bordron, B.; Laclau, J.P. Nitrogen fixation rate of Acacia mangium wild at mid rotation in Brazil is higher in mixed plantations with Eucalyptus grandis hill ex maiden than in monocultures. Ann. Forest Sci. 2018, 75, 14. [Google Scholar]
- So, H.B.; Grabski, A.; Desborough, P. The impact of 14 years of conventional and no-till cultivation on the physical properties and crop yields of a loam soil at Grafton NSW, Australia. Soil Tillage Res. 2009, 104, 180–184. [Google Scholar]
- Blagodatskaya, E.V.; Blagodatsky, S.A.; Anderson, T.H.; Kuzyakov, Y. Priming effects in Chernozem induced by glucose and N in relation to microbial growth strategies. Appl. Soil Ecol. 2007, 37, 95–105. [Google Scholar]
- Demoling, F.; Figueroa, D.; Bååth, E. Comparison of factors limiting bacterial growth in different soils. Soil Biol. Biochem. 2007, 39, 2485–2495. [Google Scholar]
- Cizungu, L.; Staelens, J.; Huygens, D.; Walangululu, J.; Muhindo, D.; Van Cleemput, O.; Boeckx, P. Litterfall and leaf litter decomposition in a central African tropical mountain forest and Eucalyptus plantation. For. Ecol. Manag. 2014, 326, 109–116. [Google Scholar]
- Rottstock, T.; Joshi, J.; Kummer, V.; Fischer, M. Higher plant diversity promotes higher diversity of fungal pathogens, while it decreases pathogen infection per plant. Ecology 2014, 95, 1907–1917. [Google Scholar]
- Wardle, D.A.; Jonsson, M.; Bansal, S.; Bardgett, R.D.; Gundale, M.J.; Metcalfe, D.B. Linking vegetation change, carbon sequestration and biodiversity: Insights from island ecosystems in a long term natural experiment. J. Ecol. 2012, 100, 16–30. [Google Scholar]
- Zhong, J.D.; Pan, P.; Xiao, S.H.; Ouyang, X.Z. Influence of Eucalyptus plantation on soil organic carbon and its fractions in severely degraded soil in Leizhou Peninsula, China. Forests 2022, 13, 1606. [Google Scholar] [CrossRef]
- Lombao, A.; Barreiro, A.; Carballas, T.; Fontúrbel, M.T.; Martín, A.; Vega, J.A.; Fernández, C.; Díaz-Raviña, M. Changes in soil properties after a wildfire in Fragas do Eume Natural Park (Galicia, NW Spain). CATENA 2015, 135, 409–418. [Google Scholar]
- Oliveira, F.C.C.; Ferreira, G.W.D.; Dungait, J.A.J.; Araújo, E.F.; Soares, E.M.B.; Silva, I.R. Eucalypt harvest residue management influences microbial community structure and soil organic matter fractions in an afforested grassland. Soil Till. Res. 2021, 205, 104787. [Google Scholar]
- Bhattacharyya, R.; Rabbi, S.M.; Zhang, Y.; Young, I.M.; Jones, A.R.; Dennis, P.G.; Menzies, N.W.; Kopittke, P.M.; Dalal, R.C. Soil organic carbon is significantly associated with the pore geometry, microbial diversity and enzyme activity of the macro-aggregates under different land uses. Sci. Total Environ. 2021, 778, 146286. [Google Scholar]
- Flemming, H.C.; Wuertz, S. Bacteria and archaea on earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [PubMed]
- Sui, X.; Zhang, R.T.; Yang, L.B.; Xu, N.; Zhong, H.X.; Wang, J.F.; Ni, H.W. Bacterial community functional diversity of Calamagrostis angustifolia wetland in Sanjiang Plain. Res. Environ. Sci. 2016, 29, 1479–1486. (In Chinese) [Google Scholar]
- Courty, P.E.; Buee, M.; Diedhiou, A.G.; Frey Klett, P.; Le Tacon, F.; Rineau, F.; Turpault, M.P.; Uroz, S.; Garbaye, J. The role of ectomycorrhizal communities in forest ecosystem processes: New perspectives and emerging concepts. Soil Biol. Biochem. 2010, 42, 679–698. [Google Scholar]
- Cai, Y. Distribution and Accumulation of Microbial-Derived Organic Carbon in Soils: The Regulatory Mechanism of Minerals and Substrates; University of Chinese Academy of Sciences in Chinese: Beijing, China, 2020. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).